Potencjał spoczynkowy to różnica potencjałów (napięcie) między obiema stronami błony.
Potencjały spoczynkowe mają wartości ujemne, które w komórkach nerwowych wahają się między –65 mV a –90
mV. Powstanie potencjału spoczynkowego jest spowodowane przede wszystkim tendencją jonów potasu do
przepływania zgodnie z gradientem stężenia tych jonów z wnętrza na zewnątrz błony komórkowej. Powoduje to
pozostanie niewielkiego nadmiaru ładunków ujemnych po wewnętrznej stronie błony. Inne jony (np. sodu) jedynie
w niewielkim stopniu wpływają na wartości potencjału spoczynkowego. Siła elektrochemiczna, powodująca ruch
jonu poprzez błonę komórkową, jest różnicą między potencjałem spoczynkowym a potencjałem równowagi dla
danego jonu. Potencjał równowagi jonu jest to taki potencjał, przy którym wypływ tego rodzaju jonów z komórki
jest równy ich wpływowi do jej wnętrza.
Em = [RT/zF] * ln (przewodność: PNa[Na]out+PK[K]out+PCl[Cl]in/ to samo w konfiguracji in in out)
Potencjał czynnościowy (czyli iglicowy) - przejściowa zmiana potencjału błonowego komórki, związana z
przekazywaniem informacji. Bodźcem do powstania potencjału czynnościowego jest zmiana potencjału
elektrycznego w środowisku zewnętrznym komórki. Wędrujący potencjał czynnościowy nazywany jest impulsem
nerwowym. Faza depolaryzacji i repolaryzacji potencjału czynnościowego (iglica) trwa nie więcej niż 1 ms i osiąga
maksymalnie wartości około +30 mV. Hiperpolaryzacja następcza może trwać kilkadziesiąt milisekund. W trakcie
potencjału czynnościowego neurony stają się niepobudliwe, zaś później, podczas hiperpolaryzującego potencjału
następczego ich pobudliwość jest zmniejszona. Zjawiska te nazywamy refrakcją bezwzględną i względną. Ze
względu na okres refrakcji bezwzględnej oraz refrakcji względnej komórki nerwowe człowieka nie mogą generować
potencjałów czynnościowych z dowolną częstotliwością. Jednak w najbardziej sprzyjających okolicznościach
częstotliwość potencjałów czynnościowych może dojść do 100 impulsów na sekundę.
Miejscem powstawania potencjału czynnościowego w komórce nerwowej jest tzw. wzgórek aksonowy, skąd
potencjał iglicowy rozprzestrzenia się po powierzchni błony komórkowej wypustki osiowej- aksonu. Generowaniem
potencjałów czynnościowych rządzi zasada: "wszystko albo nic": do zapoczątkowania potencjału czynnościowego
niezbędny jest bodziec o intensywności wystarczającej do zdepolaryzowania neuronu powyżej określonej wartości
progowej; wszystkie potencjały czynnościowe w danej komórce osiągają tę samą amplitudę.
Kod częstotliwości działa dzięki: ilości wydzielanego neurotransmitera, potencjałowi receptorowemu
(tłumaczonemu na częstotliwość), potencjałowi czynnościowemu.
Szczyt nie przekroczy 58 mV (pot. Równowagi Na+)
Przebieg potencjału czynnościowego w komórce nerwowej
Jeśli dokomórkowe prądy kationów przeważają nad odkomórkowymi, następuje depolaryzacja od poziomu
potencjału spoczynkowego (ok. -70 mV) do potencjału krytycznego czyli progowego (ok. -50 mV). Po osiągnięciu
potencjału progowego, następuje otwarcie bramkowanych elektrycznie (zależne od napięcia) kanałów
przewodzących odkomórkowo kationy potasowe, oraz kanałów przewodzących kationy sodowe do wnętrza
komórki. Wskutek różnicy stężeń i potencjałów kationy sodowe szybko napływają do wnętrza komórki, niwelując
różnicę potencjałów pomiędzy środowiskiem zewnątrz- i wewnątrzkomórkowym do zera (depolaryzują błonę
komórkową). Jest to początek tzw. potencjału iglicowego. Ze względu na dużą początkową różnicę stężeń, mimo
wyrównania potencjałów, kationy sodowe napływają w dalszym ciągu do wnętrza komórki, powodując odwrotną
polaryzację błony, tzw. nadstrzał dochodzący do +35 mV. Następuje to stosunkowo szybko; w ciągu ok. 2 ms
następuje już całkowita inaktywacja kanałów sodowych. Po rozpoczęciu aktywacji sodowej, w odpowiedzi na
depolaryzację, następuje aktywacja potasowa, czyli otwarcie kanałów dla odkomórkowego prądu kationów
potasowych. Powoduje to zmianę potencjału wnętrza komórki z powrotem na ujemny, czyli repolaryzację. W tym
czasie nie jest możliwe ponowne pobudzenie komórki, jest to okres refrakcji bezwzględnej. W czasie trwania
potencjału iglicowego, a zwłaszcza pod koniec - proporcje kationów po obu stronach błony są odwrócone (Na+
wewnątrz, K+ na zewnątrz komórki). Przez cały czas trwania potencjału czynnościowego, aktywna jest pompa
sodowo-potasowa oraz inne układy transportujące jony. Poziom aktywności pompy sodowo-potasowej jest
bowiem uzależniony m. in. od stężenia kationów sodowych we wnętrzu komórki. Zatem w czasie trwania
potencjału iglicowego pompa działa z najwyższą możliwą prędkością. W ciągu kolejnych kilkudziesięciu milisekund
przywraca proporcje kationów sodowych i potasowych po obu stronach błony komórkowej. Jednak początkowo
błona jest jeszcze w stanie hiperpolaryzacji - różnica potencjałów przekracza wartość spoczynkową. Jest to okres
refrakcji względnej, kiedy komórkę da się pobudzić, ale dużo większym bodźcem - ze względu na większą
"odległość" od poziomu potencjału krytycznego. W pewnym stopniu, w różnych tkankach, w opisanych procesach
biorą udział również inne jony, głównie chlorkowe (Cl-) oraz wapniowe (Ca2+).
Wraz ze zmianą potencjału zmienia się stężenie jonów wapnia. Kanały jonowe są wysoce selektywne. Potencjał
spoczynkowy: - 60 mV.
Na+
K+
Cl-
Ca2+
Zewnątrz
140
5
147
1
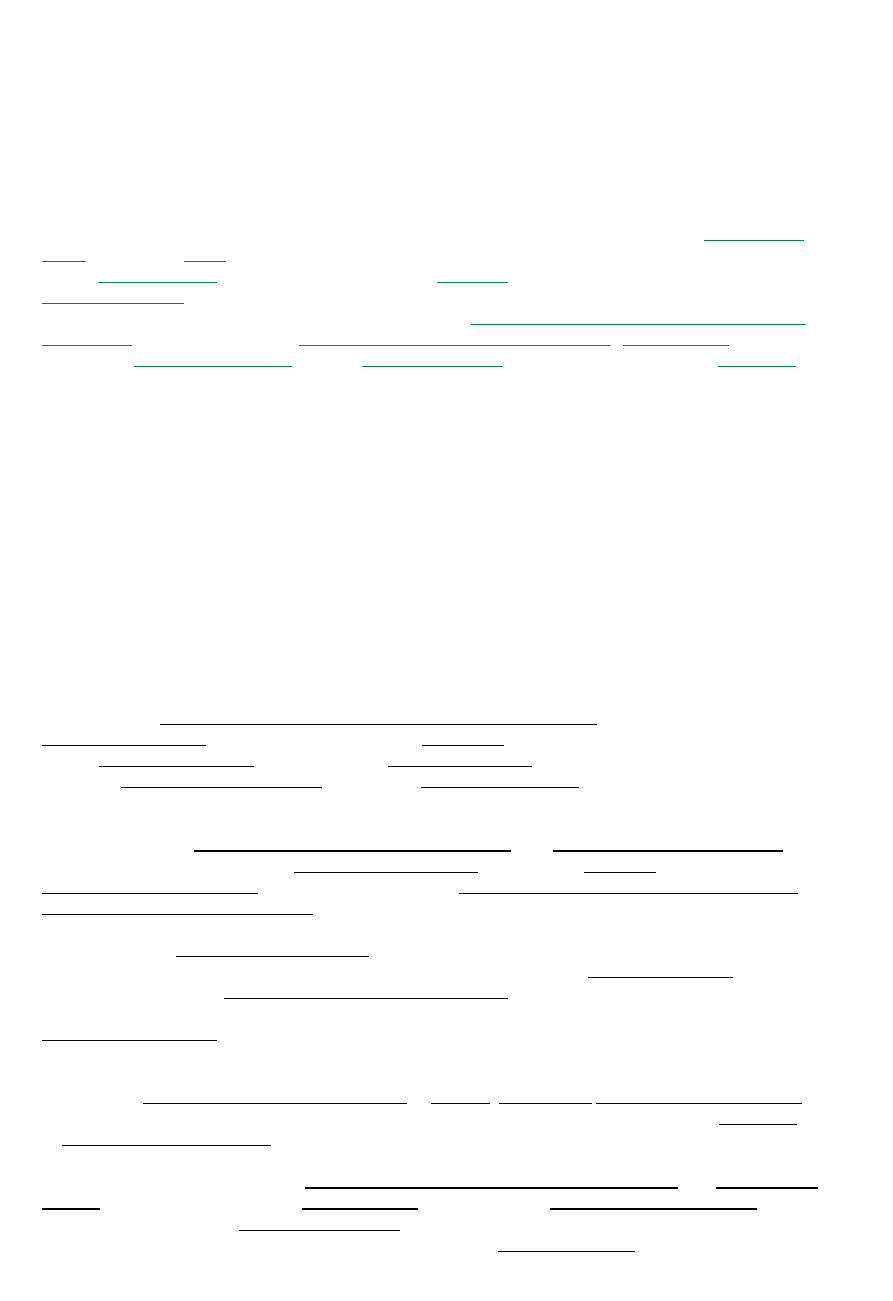
Wewnątrz
14
140
Jony potasu przemieszczały się na zewnątrz póki działał gradient, następnie minus był tak silny, że nie wypuścił
więcej.
Potencjał równowagi równy dla danych jonów, ruch zostaje przy nim zatrzymany: -84 mV.
Potencjał spoczynkowy jest zbliżony dla jonów K+, ponieważ komórka najbardziej przepuszcza te właśnie jony (w
stanie spoczynkowym) – można powiedzieć, że neurony pracują jak elektroda potasowa.
Ek+ = -84 mV
ENa+ = 58 mV
ECl- = -58 mV
ECa2+ = 116 mV
Potencjał równowagi – jeśli po dwu stronach przegrody przepuszczalnej dla jonów wytworzyć różnicę stężeń tych
jonów, to na skutek dyfuzji będą one przechodziły z przedziału o stężeniu wyższym do przedziału o stężeniu
niższym. Przepływ jonów pomiędzy przedziałami można zatrzymać wytwarzając pomiędzy nimi odpowiednią
różnicę potencjałów. Pole elektryczne będzie powodowało ruch jonów (migrację) w stronę przeciwną do kierunku
ich ruchu związanego z dyfuzją. W ten sposób może dojść do równowagi pomiędzy strumieniem dyfuzyjnym i
migracyjnym. Tym samym całkowity strumień przez przegrodę będzie równy zero i stężenia jonów w obu
przedziałach przestaną się zmieniać. Wartość różnicy potencjałów przy której dochodzi do takiej równowagi
nazywamy potencjałem równowagi. Zależy od stosunku jonów na zewnątrz i wewnątrz.
Eion=58 ln [(Ion)ext/(Ion)int]
Dla K+: dużo w środku, ucieka na zewnątrz, zgodnie z gradientem, do dużego minusa w środku (więcej nie
wypuści
Dla Na+: dużo na zewnątrz, wchodzi do środka, bo w środku minus (siła elektryczna), do mniejszego
stężenia (siła chemiczna), aż do ładunku + w komórce (więcej nie wpuści)
Dla Ca2+: wchodzi do środka, bo dużo na zewnątrz
K+ i Cl- odpowiadają za przywracanie stanu spoczynkowego, Na+ i Ca+ za potencjał czynnościowy. Czynniki
ograniczające przewodność K i Na prowadzą do ograniczenia pracy mięśni
Jeśli zwiększymy stężenie K to pot. Spoczynkowy się zwiększy, zanika więc możliwość generowania potencjałów
czynnościowych.
Depolaryzacja – zmniejszenie elektroujemnego potencjału elektrycznego błony komórkowej spowodowane
napływem przez kanały jonowe w błonie komórkowej jonów sodu do cytoplazmy komórki (potencjał zmienia się
średnio od −80 mV do +10 mV). Prowadzi ona do pobudzenia komórki nerwowej lub mięśniowej. Jeżeli wartość
potencjału przekroczy wartość progową to dojdzie do przekazania informacji.
Repolaryzacja – proces odwrotny do depolaryzacji. Zespół zjawisk fizykochemicznych następujący po przejściu
impulsu nerwowego, przywracający polaryzację błony komórkowej, czyli różnicę potencjału elektrycznego
(potencjał spoczynkowy) między jej zewnętrzną a wewnętrzną powierzchnią. Wewnątrz komórki ładunek staje się
ujemny, a na zewnątrz dodatni. Istotny jest w tym procesie ruch jonów sodu z wnętrza komórki na zewnątrz, a
jonów potasu w przeciwnym kierunku.
Hiperpolaryzacja - stan nadmiernej polaryzacji błony komórkowej w następstwie zwiększenia elektroujemności
wewnątrz komórki; trwa stosunkowo krótko. Błona komórkowa osiąga wtedy potencjał ok. -80 mV. Podczas
hiperpolaryzacji komórka nie jest zdolna do przewodzenia impulsu, przekazywania informacji.
Prąd płynący przez błonę:
Im = Ii (prąd jonowy) + Ic (prąd pojemnościowy
, nie przechodzi przez błonę, pozwala
tylko na odpowiednie ustawienie jonów)
Rezystancja – zależy od otwarcia kanałów jonowych, w Ohmach, nie równa się przewodności (w Siemensach).
Błona daje doskonałą izolację, białka zmniejszają rezystancję, przewodność wzrasta. Rezystancja zmienia się wraz
ze stanem czynnościowym błony.
Patch clamp – metoda polegająca na rejestracji aktywacji pojedynczych kanałów jonowych oraz aktywacji całej
komórki. Do komórki wprowadza się cienką elektrodę, która pozwala na rejestrację aktywacji kanałów. Umożliwia
wykonywanie pomiarów na pojedynczych kanałach; doprowadzić należy do stałego kontaktu pipety z błoną i
następnie rejestracji prądów przez przylegającą pow. Błony, przy ustalonym napięciu między elektrodami
pomiarowymi. Dzięki małej powierzchni badania i małej ilości kanałów na niej występujących można uzyskać tą
metodą charakterystyki transportowe pojedynczych jonów.
Voltage clamp – metoda stabilizacji napięcia; pozwala na pomiar prądów przepływających przez błonę komórki
nerwowej w czasie jej pobudzenia.
Hiperpolaryzacja zamyka wszystkie kanały, a depolaryzacja otwiera.
Akustyka – dział fizyki i techniki obejmujący zjawiska związane z powstawaniem, propagacją i oddziaływaniem fal
akustycznych. Ze względu na różnorodność działów akustyka jest obecnie traktowana jako nauka
interdyscyplinarna obejmująca oprócz akustyki ogólnej, zajmującej się zagadnieniami podstawowymi, również
szereg działów akustyki stosowanej, zajmujących się praktycznym zastosowaniem zjawisk akustycznych.
Słuch – zmysł umożliwiający odbieranie (percepcję) fal dźwiękowych; „słyszeć” to posiadać zdolność detekcji fal i
rozpoznawania ich cech fizycznych.
Fale - materii; mechaniczne; elektromagnetyczne.
Fala dźwiękowa - to zaburzenie mechaniczne rozchodzące się w ośr. Sprężystym obdarzonym masą; zaburzone bo
przenosi energię mechaniczną przez drgające cząsteczki ośrodka (zgęszczenia i rozrzedzenia). Fale te to fale
ciśnienia ośrodka, fale podłużne (drgania mają kierunek oscylacji zgodny z kierunkiem ruchu fali). Źródłem
dźwięków słyszalnych są ciała wprawione w drgania o dostatecznej energii wywołującej wrażenie słuchowe.
ƛ = c * T
ƛ = c/f
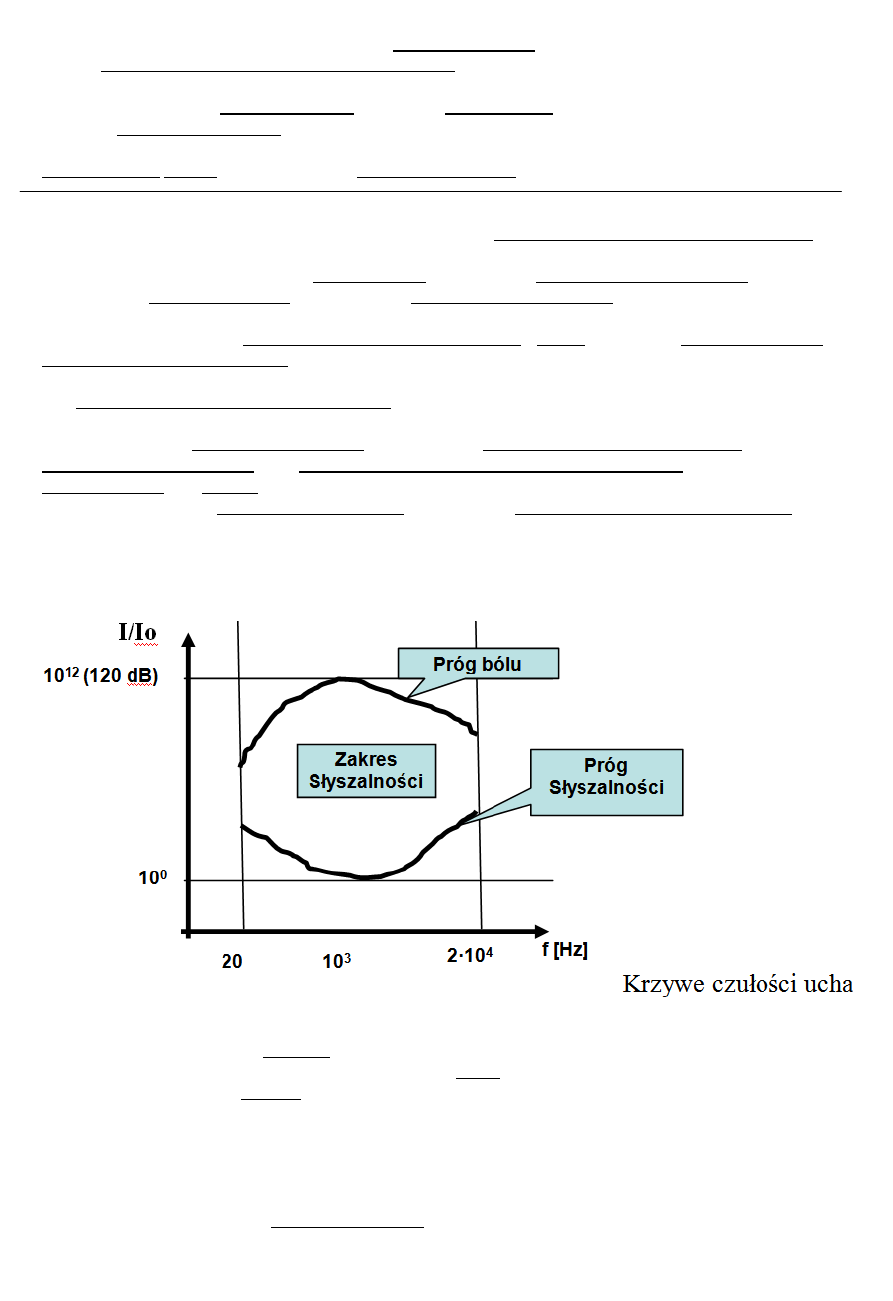
Ucho ludzkie reaguje na dźwięki o częstotliwości od 20 do 20 000 Hz
Własności dźwięków
− Obiektywne:
o częstość [Hz] -> wysokość
o charakter krzywej falowej (kształt drgań) -> barwa
o natężenie -> głośność
− Subiektywne (związane z wrażeniami odczuwanymi przez człowieka):
o wysokość
o barwa
o głośność
Infradźwięki – fale akustyczne o częstościach niższych (odczuwanie tylko wibracji), poniżej 16 Hz. Infradźwięki mają
bardzo dużą długość fali - powyżej 17 m, przez to słabo tłumione mogą rozchodzić się na znaczne odległości.
Drugim problemem jest ich słabe tłumienie poprzez ekrany akustyczne. Wytwarzane naturalnie (wodospady,
tornada), bądź przez duże maszyny (wiatraki, odrzutowce, silniki rakietowe).
Ultradźwięki – fale akustyczne o częstościach wyższych, słyszalne przez nietoperze, pochłaniane przez włosy, nie są
odbijane,
powyżej 20 kHz. Niektóre zwierzęta mogą emitować i słyszeć ultradźwięki, np. pies, szczur, delfin,
wieloryb, chomik czy nietoperz.
Wytwarzanie:
• mechaniczne - układy drgające (struny, płytki sprężyste, piszczałki). Wykorzystują one drgania samego
tworzywa albo przepływ gazów czy cieczy. Typowe przykłady to syreny ultradźwiękowe i piszczałka
Pohlmana-Janowskiego, wykorzystywana do wytwarzania rozmaitych emulsji w chemii i biotechnologii
• termiczne - poprzez wyładowania elektryczne w płynach i gazach, poprzez ciągle lub impulsowe
podnoszenie temperatury przewodników prądu.
• magnetostrykcja - zmiana długości rdzenia elektromagnesu pod wpływem zmiennego prądu
przepuszczanego przez solenoid nawinięty na ten rdzeń.
• odwrócenie efektu piezoelektrycznego - polega na doprowadzeniu do przeciwległych płaszczyzn kryształu
kwarcu lub innego minerału szybko zmiennego napięcia elektrycznego. Prowadzi to do rozszerzenia lub
skurczenia płytki i do powstania drgań o odpowiedniej częstotliwości.
• optyczne - laserem można wytworzyć fale sprężyste w szerokim zakresie częstotliwości ultradźwiękowych
aż do zakresu hiperdźwiękowego.
Ultradźwięki dźwięki małej długości fali pozwalają na uzyskanie dokładnych obrazów przedmiotów. Urządzenie,
które umożliwia obserwację głębin morskich to sonar. Jego zastosowanie to lokalizacja wszystkich obiektów
zanurzonych w wodzie. Sonary wykorzystywano w okrętach podwodnych. Ultradźwięki znajdują także
zastosowanie w medycynie. Za pomocą urządzenia generującego i rejestrującego fale ultradźwiękowe
(ultrasonograf) można uzyskać obraz narządów wewnętrznych.
Ultradźwięki pozwalają też na pomiar odległości przy pomocy dalmierza ultradźwiękowego, w zakresie od 1 do 10
m. Jeżeli wykorzysta się silne źródło ultradźwięków, to mogą one niszczyć, rozgrzewać niektóre materiały, co
pozwala na obróbkę powierzchniową wytwarzanych przedmiotów (obróbka ultradźwiękowa).
Wiele gatunków posługuje się nimi w celu echolokacji. Na przykład większość nietoperzy wytwarza ultradźwięki,
wiele gatunków posiada również duże i bardzo sprawne uszy. Są one zdolne do wykrywania owadów latających w
ciemnościach (ćmy). Ultradźwięki wykorzystują również walenie. Wieloryby używają ich do echolokacji w podobny
sposób jak to się odbywa w technice morskiej. Dzięki temu mogą namierzać ławice ryb lub plankton.
Najdoskonalszy zmysł echolokacji posiadają delfiny. Na ich głowach znajduje się rezonator pozwalający na
generowanie precyzyjnie ukierunkowanego strumienia ultradźwięków. Jednocześnie ogromne mózgi delfinów są w
stanie przetworzyć uzyskane w ten sposób dane w trójwymiarowy model otoczenia.
Różny skład drgań harmonicznych – skład różnych podstawowych barw, czyli zbiór fal o różnych barwach. Ciśnienie
akustyczne – opisuje natężenie dźwięku [Pa]
Ton – dźwięk prosty, mający sinusoidalny przebieg o ściśle określonej częstotliwości, amplitudzie i fazie; wiązka
monochromatyczna. Żaden ze składników mowy ludzkiej nie jest tonem. Każdy dźwięk zależy od natężenia
występujących w nim tonów prostych na pasma częstotliwości. Basy – tony o niższych częstotliwościach i małym
natężeniu
Dźwięk – zsumowanie tonów.
Wrażenie słuchowe, spowodowane falą akustyczną rozchodzącą się w ośrodku
sprężystym (ciele stałym, cieczy, gazie). Twierdzenie Fourniera: każdy dźwięk okresowy o częstotliwości v można
przedstawić jako sumę tonów o częstotliwościach v, v2, v3 itd. I odpowiednio dobranych amplitudach. Dźwięki o
naturze nieokreślonej to szmery, hałasy itp.
Barwa dźwięku – cecha, która pozwala odróżnić brzmienia różnych instrumentów, głosu; uzalezniona od ilości,
rodzaju i natężenia fal.
Pogłos (rewerberacja) – zjawisko stopniowego zanikania energii dźwięku po ucichnięciu źródła, związane z
występowaniem dużej liczby fal odbitych od powierzchni pomieszczenia. Ucho ludzkie odczuwa pogłos jako
przedłużenie dźwięku. Pogłos jest określany ilościowo przez czas pogłosu, czyli ilość sekund potrzebną na spadek
energii akustycznej o 60 dB.
Hałas – dźwięki zazwyczaj o nadmiernym natężeniu (zbyt głośne) w danym miejscu i czasie.
Szmer - zjawisko akustyczne wywołane przez nieregularne drgania dźwięku, obejmujące wszelkie odgłosy o
nieokreślonej wysokości (huk, trzask, szum). Szmerami są również odgłosy niektórych instrumentów perkusyjnych,
np. bębna, trójkąta, gongu, kastanietów itp
Natężenie dźwięku – natężenie jest proporcjonalne do ilości energii, ta natomiast do kwadratu amplitudy, czyli
natężenie jest proporcjonalne do kwadratu amplitudy drgań źródła:
I = E/St [J/m2*s = W/m2]
I~E -> I~A2 -> E~A2
Głośność – cecha wrażenia słuchowego, która umożliwia odróżnianie dźwięków cichszych i głośniejszych. Jest
pojęciem psychoakustycznym i nie może być utożsamiana z parametrami fizycznymi, chociaż od nich zależy, np. od
ciśnienia, struktury widmowej, czasu trwania. Wrażenie głośności określa się przez poziom głośności w fonach
(poziom głośności dowolnego dźwięku w fonach jest liczbowo równy poziomowi natężenia - wyrażonego w
decybelach - tonu o częstotliwości 1 kHz, którego głośność jest równa głośności tego dźwięku.) lub przez głośność
w sonach.
L [dB]
fony
poz. natężenia dźwięku
głośność
głośność
poz. głośności
Poziom natężenia dźwięku – logarytmiczna miara natężenia dźwięku w stosunku do pewnej umownie przyjętej
wartości odniesienia, wyrażana w decybelach.
L = 10 log10 (I/Io)
– prawo Webera-Fechnera: wrażenie jest wprost proporcjonalne do logarytmu ze stosunku
bodźców. Dźwięk wydaje się głośniejszy gdy ma wyższe natężenie.
Bel – jednostka miary wielkości ilorazowych. W wypadku pomiaru mocy (często w zastosowaniach w elektronice):
Pb = log (P/Po)
Często stosuje się jednostkę podwielokrotną, decybel:
1 dB = 1/10 B
Rozchodzenie się fal akustycznych – rozchodzą się w ośrodkach substancjonalnych, różnymi prędkościami (fala
mechaniczna, którą trzeba odnieść do ośrodka).
Dźwięk w cieczkach i ciałach stałych:
V= √ E / g
najszybciej rozchodzi się w stali, najwolniej w powietrzu (330
m/s), także
v = λ f
Pochłanianiu ulegają fale przechodząc przez ośrodek substancjonalny. Natężenie fali zmienia się wtedy
ekspotencjalnie:
I = Io *e^ -Ex
Wartość współczynnika E (pochłaniania) zależy od częstotliwości drgań i rodzaju ośrodka. Pochłanianie zwiększa się
wraz ze wzrostem częstotliwości drgań, a wsp. Pochłaniania jest proporcjonalny do V^3 i do lepkości
E ~ 1/V3, E~n
Odbicie i załamanie dźwięków zachodzi na granicy ośrodków. Opór akustyczny ośrodka to iloczyn rozchodzenia się
fali i gęstości ośrodka:
z = V * g
Jeżeli opory sąsiadujących ośrodków są podobne, do przeważa załamanie fali (większość przechodzi z jednego
ośrodka do drugiego). Jeśli opory są różne przeważa odbicie.
Zjawisko Dopplera - zjawisko obserwowane dla fal, polegające na powstawaniu różnicy częstotliwości wysyłanej
przez źródło fali oraz zarejestrowanej przez obserwatora, który porusza się względem źródła fali.
f = c / λ
− obserwator zbliża się do dźwięku:
f = c + u / λ = c + u / c*T =1/T =fo
f=(c + u/c) * fo =(1+u/c)*fo
Obserwator usłyszy dźwięk wyższy – liczba zarejestrowanych przez ucho drgań jest większa od drgań
wychodzących
− obserwator oddala się:
obserwator w miejscu oddalenia usłyszy dźwięk niższy, częstotliwość jest subiektywna
− źródło zbliża się do obserwatora:
obserwator zarejestruje podwyższony dźwięk (podwyższa się f), zmienia się długość fali
− źródło oddala się od ruchomego obserwatora:
zmniejsza się częstotliwość

Rezonans magnetyczny - działanie rezonansu magnetycznego opiera się na wykorzystaniu właściwości protonów
wodoru, które w odpowiednio silnym polu magnetycznym ulegają namagnesowaniu, pobierają impulsy fal
elektromagnetycznych i wysyłają impuls podczas zaniku pobudzenia. Budowa tkanek ludzkich charakteryzuje się
różnym stopniem zawartości wody, co też powoduje różne natężenie sygnału w badaniu. Wysyłane impulsy są
przekształcane za pomocą urządzeń elektronicznych w obraz diagnostyczny pojawiający się na monitorze.
Obrazowanie magnetyczno-rezonansowe – nieinwazyjne, opiera się na zjawisku jądrowego rezonansu
magnetycznego. Zjawisko to może zajść w próbce zawierającej jądra o różnym od zera spinie umieszczonej w
silnym stałym polu magnetycznym. W takich warunkach próbka ulega częściowej polaryzacji opisywanej wektorem
magnetyzacji. Jeśli tak spolaryzowana próbka zostanie poddana działaniu innego pola magnetycznego,
zaobserwować można oddziaływanie między polem a magnetyzacją próbki. Jeśli różne części próbki znajdują się w
różnych polach, nie można mówić o jednej częstości odbieranego sygnału, mamy do czynienia z wieloma
częstościami a najczęściej z ciągłym jej widmem. Jeśli mapa w jakim znajduje się próbka jest znana, informacja
przestrzenna może zostać odkodowana a zebrane widma mogą zostać zamienione na obraz próbki. Modulowanie
pola głównego i jednoczesny pomiar sygnału rezonansu magnetycznego są podstawą metody obrazowania MR.
Odkodowanie obrazu nazywane jest rekonstrukcją. Jądrem rezonansowym najczęściej wykorzystywanym w
obrazowaniu MR jest proton — jądro atomu wodoru mające spin połówkowy i występujące powszechnie w
obiektach biologicznych w cząsteczkach wody.
Spektroskopia NMR, Spektroskopia Magnetycznego Rezonansu Jądrowego – jedna z najczęściej stosowanych
obecnie technik spektroskopowych w chemii i medycynie. Spektroskopia ta polega na wzbudzaniu spinów
jądrowych znajdujących się w zewnętrznym polu magnetycznym poprzez szybkie zmiany pola magnetycznego, a
następnie rejestrację promieniowania elektromagnetycznego powstającego na skutek zjawisk relaksacji, gdzie
przez relaksację rozumiemy powrót układu spinów jądrowych do stanu równowagi termodynamicznej. NMR jest
zatem jedną ze spektroskopii emisyjnych.
Spektroskopia EPR czyli spektroskopia elektronowego rezonansu paramagnetycznego lub elektronowy rezonans
spinowy (ESR) – technika pozwalająca na wykrycie związków posiadających jeden lub więcej niesparowanych
elektronów, takich jak wolne rodniki, jony metali przejściowych, czy pierwiastki ziem rzadkich.
Podstawowe fizyczne założenia techniki są analogiczne do tych wykorzystywanych w spektroskopii NMR, ale
badane są spiny elektronów, a nie spiny jąder atomowych. Z powodu różnic w masie pomiędzy jądrami a
elektronami, w technice EPR używane są słabsze pola magnetyczne i wyższe częstotliwości promieniowania
mikrofalowego niż w spektroskopii NMR. Dla elektronów, rezonans paramagnetyczny w polu magnetycznym o
wartości ok. 0,3 T zachodzi przy częstotliwości ok. 10 GHz (tzw. pasmo X).
Spektroskopia EPR jest wykorzystywana m.in. w fizyce ciała stałego do identyfikacji wolnych rodników, w chemii do
badań przebiegu reakcji oraz w biologii i medycynie do śledzenia znaczników spinowych. Metoda EPR jest też
stosowana do datowania w badaniach archeologicznych, np. wieku szkliwa zębów.
Aby badać układy biologiczne zaprojektowano mało reaktywne molekuły mogące wiązać się w specyficznych
miejscach w komórce czy białku. Pozwala to na otrzymanie informacji o otoczeniu danej próbki spinowej.
Wzrok – widzenie umożliwiają fotoreceptory zlokalizowane w wyspecjalizowanych narządach (oko). Towarzyszą im
dodatkowe struktury usprawniajcie percepcje i chroniące właściwy narząd wzroku (aparat ochronny: (brwi,
powieki, rzęsy i narząd łzowy)). Następnie bodziec jest przekazywany do dalszych obszarów układu nerwowego,
gdzie jest analizowany. U organizmów wyższych mówi się o drodze wzrokowej przekazującej informacje do kory
wzrokowej.
Rozróżniamy przedmioty jaśniejsze i ciemniejsze oceniając ilość światła; określamy barwy poprzez czułość na
długość fali; ustalamy kształt przedmiotów na podstawie odległości od obserwatora.
Widzenie – detekcja, czyli formowanie obrazu. Analiza amplitudy, długości fali i rozmieszczenia źródeł światła (też
tych odbijających). Widzimy światło odbite i rozproszone. Światło jest falą elektromagnetyczną.
Oko – narząd receptorowy umożliwiający widzenie. W najprostszym przypadku chodzi o zdolność wykrywania
pewnego zakresu promieniowania elektromagnetycznego. Bardziej skomplikowane oczy są w stanie dostarczyć
informacje o kierunku padania światła, jego intensywności oraz kształtach obiektów.
Światło przechodzi przez przednią część twardówki – rogówkę; wpada do oka przez źrenicę regulowaną tęczówką –
kolorową częścią oka; przechodzi przez soczewkę, która załamuje promienie świetlne; przechodzi przez ciało
szkliste; promienie padają na wewnętrzną warstwę oka – siatkówkę (gdzie powstaje odwrócony obraz), składającą
się z fotoreceptorów – czopków (kolor) i pręcików (kształt i ruch). Plamka żółta – największe skupisko czopków;
plamka ślepa – tam nie ma fotoreceptorów, od niej wychodzi nerw wzrokowy.
Siatkówka oka ssaka jest wrażliwa na UV, ale promienie nie przenikają z powodu soczewki działającej jak filtr.
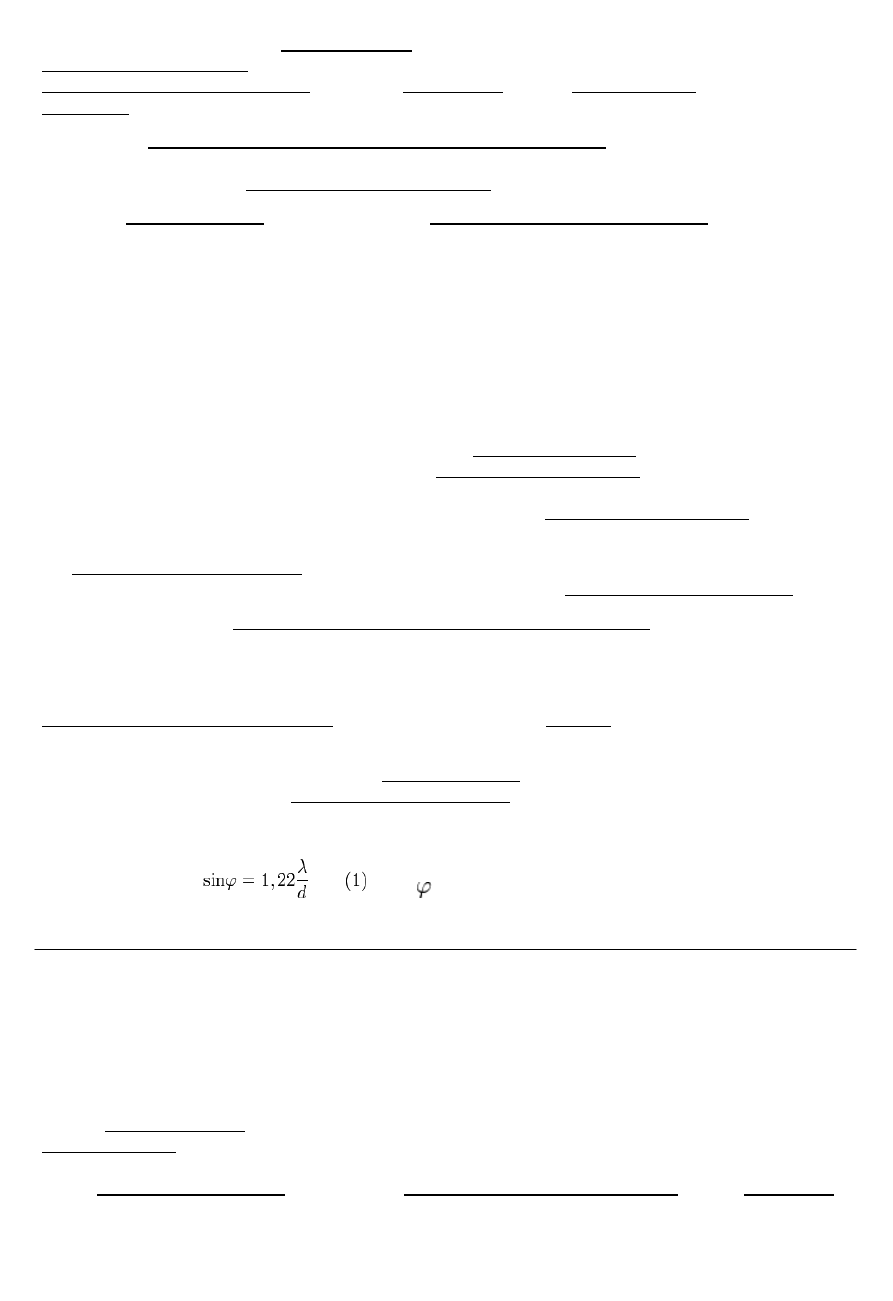
Układ optyczny oka – składa się z rogówki i soczewki, przesłoną jest tęczówka, która chroni przed silnym światłem i
eliminuje aberracje sferyczne. Na środku wsp. załamania – 1,4, na zewnątrz 1,33.
Oś optyczna to środek symetrii układu optycznego, oś widzenia to oś, którą promienie tworzą dochodząc do
receptorów.
Akomodacja – zdolność soczewki do zmiany kształtu lub odległości od siatkówki. Punkt bliży wzrokowej to
najbliższy punkt jaki oko jest w stanie widzieć, punkt dali wzrokowej to najbliższy punkt powyżej którego soczewka
nie akomoduje. Akomodacja szwankuje przy słabym oświetleniu.
Refrakcja – zdolność skupiająca; w ukł. Optycznym jest sumą wszystkich zdolności skupiających i wynosi 63 D. Oko
miarowe dąży aby So = ∞, czyli R dąży do 0 -> R=1/∞
R = 1/So
– odległość punktu dalekiego [dioptrie]
Starowzroczność – sztywnienie soczewki, problem z akomodacją
Astygmatyzm – krzywizna rogówki
Aberracje pozaoczne – nie mają wpływu na widzenie; oko radzi sobie samo z aberracjami sferycznymi i
chromatycznymi.
Krótkowzroczność zmierzchowa – kiedy źrenica się powiększa pojawia się aberracja sferyczna.
Dalekowzroczność - wada refrakcyjna wzroku. Jest wynikiem zbyt małych rozmiarów przednio-tylnych oka (zbyt
krótką gałką oczną) w stosunku do jego siły łamiącej lub niewystarczającą siłą łamiącą układu optycznego oka (np.
zbyt płaską rogówką) w stosunku do jego długości.
Promienie równoległe, które w akomodującym oku miarowym
ogniskowane są na siatkówce, w nieakomodującym oku nadwzrocznym ogniskowane są za siatkówką.
Krótkowzroczność - jedna z najczęściej spotykanych wad refrakcyjnych wzroku polegająca na tym, że tor optyczny
oka nieprawidłowo skupia promienie świetlne. Promienie równoległe, które w akomodującym oku miarowym
ogniskowane są na siatkówce, w nieakomodującym oku krótkowzrocznym ogniskowane są przed siatkówką.
Aberracja chromatyczna – różne długości fal nie mogą być zogniskowane jednocześnie. W soczewkach tworzonych
przez człowieka koryguje się to poprzez stosowanie soczewek złożonych, mających kilka współczynników refrakcji.
Światło niebieskie ogniskuje najszybciej, najlepiej zaś wyeliminować fale najkrótsze (największy wsp. refrakcji).
Problem nie dotyczy owadów – oczy złożone nie ogniskują.
Człowiek nie widzi płaszczyzny polaryzacji – pszczoły wykorzystują ją do nawigacji (niebieskie niebo to światło
spolaryzowane, białe chmury to światło niespolaryzowane).
U niektórych węży receptorami podczerwieni są narządy policzkowe, tzw. Oczy termiczne. Mogą służyć do
percepcji źródła podczerwieni, wąż atakuje źródło promieniowania. Takie narządy nie mogą działać pod wodą –
woda ma dużą pojemność cieplną, wysokie przewodnictwo cieplne, w konsekwencji jest słaba penetracja
promieniowania podczerwonego.
Zdolność rozdzielcza -
, gdzie
-minimalny kąt między promieniami, których obrazy mają
być rozróżnialne, czyli inaczej – ich odległość kątowa;, d – średnica otworu
Właściwości optyczne materiału wynikają ze zjawisk:
− Absorpcji
− Załamania
− Odbicia
− Rozpraszania
− Własności elektrycznych
Maser - wzmacnia mikrofale za pomocą emisji wymuszonej promieniowania elektromagnetycznego. Emituje fale o
f< od podczerwieni.
Laser - generator promieniowania, wykorzystujący inwersję obsadzeń i emisję wymuszoną. Emituje podczerwień
światła widzialnego i wyższych zakresów częstotliwości.
Emisja wymuszona – jeden z 3 procesów oddziaływania światła z materią, w wyniku tego zjawiska zmienia się stan
kwantowy atomu. Emisja fotonu przez materię zachodzi w wyniku oddziaływania z innym fotonem, warunek to
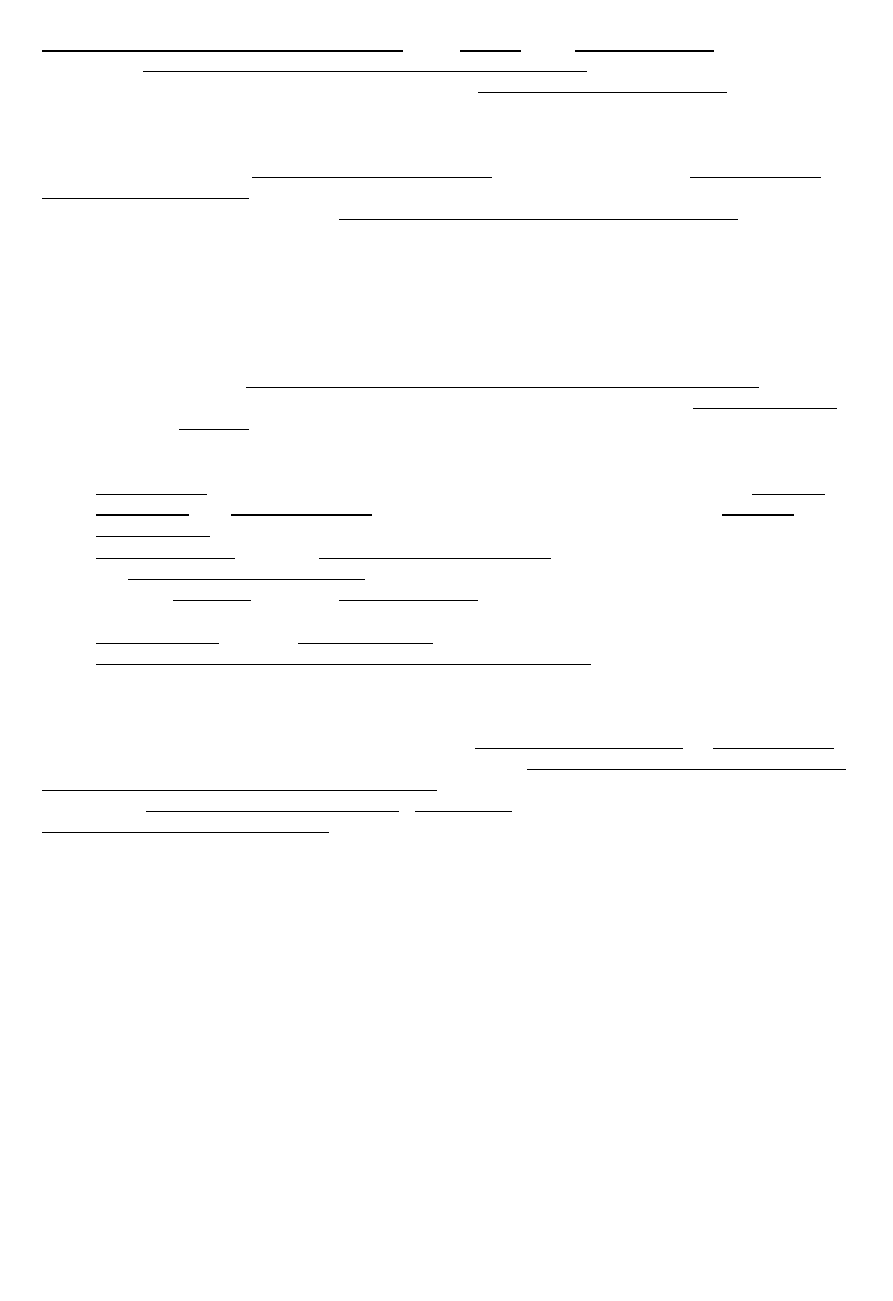
równość energii fotonu z energią wbudzenia atomu. Foton inicjujący emisję nie jest pochłaniany, tylko wyzwala
proces. Foton emitowany ma f, E, Falę i polaryzację taką jak foton wywołujący emisję (są spójne), kierunek ruchu
ten sam. Światło spójne to złożone z takich fotonów. Zjawisko odwrotne do pochłaniania fotonów. Zachodzi gdy
atom wzbudzony zderza się z fotonem.
Inwersja obsadzeń – konieczna do zajścia emisji wymuszonej. Należy zapewnić w ośrodku wzmacniającym
odpowiednie warunki: więcej elektronów w stanie wzbudzonym, materiał musi pozwalać na odpowiednio długi
czas przebywania elektronów na danym poziomie wzbudzonym (poziom niestabilny; foto wyzwala wiele fotonów
na raz, ta sama f). Uzyskuje się za pomocą oświetlania światłem, laserem bądź światłem błyskowym.
Absorbcja – kolejny proces oddziaływania światła z materią, w wyniku absorpcji fotonu o częstości V spełnia się
równanie E2-E1 = hV, czyli E = hV
Emisja spontaniczna – ostatni proces oddziaływania światła z materią; w wyniku procesu atom emituje foton i
przechodzi ze stanu wzbudzonego w podstawowy.
Promieniowanie laserowe – spójne w czasie i przestrzeni, spolaryzowane, wiązka o małej rozbieżności
(skolimowana, zbiór promieni równoległych, nie ulega zmianie przy oddalaniu się od źródła), monochromatyczna.
Lasery najlepsze to niebieskie – krótkie fale, mała f, mieści dużo informacji, niebieska fala mało się ugina.
Części lasera
− Ośrodek czynny: zespół atomów, jonów lub cząsteczek, w odpowiednich warunkach zachodzi kwantowe
wzmocnienie, czyli powielenie fotonów (akcja laserowa); decyduje o parametrach lasera, określa dł.
Emitowanej fali;
− Rezonator optyczny: umożliwia wybranie odpowiednich fotonów (wzmocnienie); układ optyczny pełniący
rolę sprzężenia zwrotnego dodatniego dla światła o wybranym kierunku i długości fali; dwa dokładnie
wykonane zwierciadła, może zajść wielokrotne odbicie, wtedy przebiegają tylko fotony mogące wywołać
akcję laserową;
− Układ pompujący: dostarcza energię do ośrodka czynnego, który wytwarza inwersję obsadzeń.
Przeniesienie jak największej liczby elektronów do stanu wzbudzonego. Pompowanie odbywa się: błysk
lampy, innego lasera, przepływ pędu w gazie, r-cje chemiczne, zderzenia atomów, wystrzelenie wiązki e.
Oddziaływanie na tkanki – zależy od parametrów lasera: moc, dł. Fali, czas trwania impulsu lub czas naświetlania i
pole powierzchni przekroju wiązki padającej; oraz własności obiektu: wsp. absorpcji i rozpraszania promieniowania,
gęstość, ciepło właściwe, wsp. przewodnictwa cieplnego.
Jeśli dominuje efekt biochemiczny nad termicznym – biostymulacja laserowa (mała moc promieniowania). Jeśli
dominuje termiczny – efekt koagulacyjny, czyli chirurgia laserowa.
Efekt fototermiczny – po absorpcji tkanka nagrzewa się i ulega denaturacji. Wysyłanie krótkich impulsów o dużej
mocy powoduje rozrywanie wiązań bez nagrzewania, czyli usunięcie tkanki bez niszczenia innych.
Efekt fotojonizujący - współistnienie z ef. Fototermicznym, krótkie impulsy o dużej gęstości mocy.
Siwert jest jednostką pochodną układu SI wielkości fizycznych odnoszących się do działania promieniowania
jonizującego na organizmy żywe. Siwert jest względnie dużą jednostką – u człowieka już po przekroczeniu dawki
skutecznej 1 Sv promieniowania gamma dla całego ciała może wystąpić ostry zespół popromienny, mogący prowadzić
do śmierci. J/kg
Bekerel, Bq to jednostka miary aktywności promieniotwórczej w układzie SI (jednostka pochodna układu SI). Próbka ma
aktywność 1 Bq, gdy zachodzi w niej jeden rozpad promieniotwórczy na sekundę. Na przykład, jeśli dane ciało generuje
1000 cząstek alfa na sekundę poprzez rozpad alfa (i jednemu rozpadowi odpowiada emisja jednej cząstki alfa), to
aktywność tego ciała jest równa 1000 Bq, czyli 1 kBq. Miano bekerela to s-1.
Grej (Gy) – jednostka dawki pochłoniętej w układzie SI (Jednostka pochodna układu SI). Jest to ilość energii
promieniowania (w dżulach) pochłoniętej przez kilogram materii. 1 Gy = 1 m²·s-2 lub 1 Gy = 1 J/kg

Kiur (Ci) – nieużywana pozaukładowa jednostka miary radioaktywności. W układzie SI zastąpiona została bekerelem
(Bq). Przelicznik jest następujący:
1 Ci miał odpowiadać aktywności 1 g czystego izotopu radu o liczbie masowej 226. Współczesne dokładniejsze badania
wykazały, że radioaktywność 1 g radu 226 wynosi w rzeczywistości 3,66·1010 Bq.
Rad – jednostka dawki absorbowanej (dawki pochłoniętej) promieniowania jonizującego. Dawka absorbowana przez
napromieniowane ciało o masie 1 g, jeżeli energia przekazana temu ciału przez cząstki jonizujące równa jest 100 erg czyli
10-5 J.
1 rad = 0,01 Gy (grej) = 0,01 J/kg = 100 erg/g
Rem – jednostka równoważnika dawki promieniowania jonizującego pochłoniętego przez organizm. 1 rem jest
odpowiednikiem dawki równoważnej równej 0,01 Sv (0,01 Gy dawki pochłoniętej promieniowania gamma) pod
względem działania biologicznego.
Wyszukiwarka
Podobne podstrony:
ściąga biofizyka temat 20, Medycyna
Sciaga biofiza cwiczenie 5, I ROK STOMATOLOGIA SUM ZABRZE, BIOFIZYKA
ściąga biofiz wszystko
Ściąga biofizyka 1
Sciaga z biofizyki
ściąga biofizyka 13 2
biofizyka ściąga
ściąga statystyka, MEDYCYNA - ŚUM Katowice, I ROK, Biofizyka
biofizyka ściaga, Fizyka Medyczna UŚ Katowice, Biofizyka
biofizyka sciaga
Semestr II - Biofizyka - Ściąga - Notatki od Dominiki - Kolokwium - Maj 2011, R4
Semestr II - Biofizyka - Ściąga - Notatki od Dominiki - Kolokwium - Maj 2011, R5
Biofizyka ściąga, Anatomia człowieka
Semestr II - Kolokwium - Całość - Biofizyka - ściąga - pełna, UJK.Fizjoterapia, - Notatki - Rok I -,
Semestr I - Kolokwium I - ściąga, UJK.Fizjoterapia, - Notatki - Rok I -, Biofizyka, Materiały na zal
Kolokwium - ŚCIĄGA - 8-10.04.2011, UJK.Fizjoterapia, - Notatki - Rok I -, Biofizyka, Materiały na za
Ściąga - pojęcia z biofizyki, Naukowe, Fizjoterapia, Biofizyka
sciaga Gochy, FIZJOTREAPIA, rok 1, semestr 2, biofizyka
więcej podobnych podstron