01.03.2004.
Cytoszkielet
Ruchliwość - zdolność żywych systemów do wykonywania mechanicznej pracy kosztem energii metabolicznej.
Rodzaje ruchów:
Ruch wewnątrzkomórkowy ( funkcja: rozprowadzanie związków wewnątrz komórki, cytokineza, ruch chromosomów, tworzenie ściany komórkowej, ruch organelli komórkowych, wici, rzęsek, skurcz komórek mięśniowych)
Ruchy komórki w środowisku
Cytoszkielet: cytoplazmatyczny system filamentów białkowych eukariotów (nie mówimy o cytoszkielecie u prokariotów!).
Elementy cytoszkiletu:
Mikrotubule φ 24nm Kanał wewnętrzny φ 15nm
Filamenty pośrednie φ 10nm (funkcja podporowa)
Mikrofilamenty φ 5-7 nm (ruch)
Przyczyną ruchu są procesy polimeryzacji i depolimeryzacji elementów cytoszkieletu (oprócz filamentów pośrednich). Z cytoszkieletem współdziałają białka motoryczne.
Mikrotubule:
Funkcja:
funkcja podporowa, wzmacniająca chromosomów pobliżu błony komórkowej
organizują wnętrze komórki (kompartmentacja)
rozdział chromosomów podczas kariokinezy w anafazie, w cytokinezie
stanowią strukturę wici i rzęsek (jedność budowy: taka sama wić u wiciowca jak u plemnika, a rzęska u orzęsków taka jak w jajowodzie ssaków)
wyznaczają tory (kanały) transportowe w ruchu wewnątrzkomórkowym, budują ściany komórkowe (pęcherzyki z aparatu Gologiego tworzą fragmoplast poruszając się po szlakach wyznaczonych przez mikrotubule.).
Budowa i zasada działania: Dimer tubuliny składa się z dwóch monomerów: tubuliny α i tubuliny β (te podjednostki w cytozolu tworzą bazę do budowy MT). Każdy dołączany dimer tubuliny jest połączony z GTP i wiążą się one ze sobą znacznie ściślej, niż dimery tubuliny zawierające GDP. Stąd MT, które mają świeżo przyłączone dimery tubuliny ze związanym GTP (tzw. czapeczką GTP)wykazują tendencję do kontynuowania wzrost. Niemniej jednak od czasu do czasu, szczególnie jeśli wzrost MT jest powolnyy, podjednostki w czapeczce będą hydrolizowały GTP do GDP, zanim nowa podjednostka z GTP zostanie przyłączona. Tym sposobem następuje utrata czapeczki. Podjednostki zawierające GDP są znacznie mniej ściśle związane z polimerem i są łatwiej uwalniane z jego wolnego końca, tak że MT zaczyna się skracać.
Mikrotubule mogą być stabilizowane przez białka, które łączą się z końcem „+” (proces wpływający na położenie układów MT w komórce).
Mikrotubula (tak jak kij) ma dwa końce: „+” i „-”. Na końcu „+” zachodzi polimeryzacja (koniec β-tubulinowy), na końcu „-” depolimeryzacja (koniec α-tubulinowy).
Istnieją dwie pule mikrotubul: krótko- i długożyjąca.
Krótko żyjąca - tworzą sieć mikrotubul w czasie interfazy, a w czasie mitozy - wrzeciono podziałowe.
Długo żyjąca, stabilna - buduje rzęski i wici.
Rzęski:9x2 - dziewięć dubletów, w każdym dublecie 1 pełna i 1 niepełna mikrotubula + 2 (jeden dublet w środku)
Centrum organizacji mikrotubul (MTOC)inaczej centrum nukleacji:
U zwierząt: centrosom, zawiera struktury o kształcie pierścienia utworzone przez γ - tubulinę, a każdy taki pierścień służy jako punkt startowy do wzrostu jednej mikrotubuli, tzw. miejsce nukleacji. Dimery αβ-tubuliny dołączają się do γ-tybuliny tak, że koniec minus każdej mikrotubuli jest osadzony w centrosomie( wyjątkiem są komórki nerwowe). W centrosomie znajdują się centriole, ale ich funkcja nie jest znana (do powstania MT wystarczą pierścienie γ-tubuliny).
Zbiór mikrotubul zakotwiczonych w centrosomie ulega ciagłym zmianom , w miarę jak nowe mikrotubule wyrastaja a stare skracaja się - tzw dynamiczna niestabilność
W centrosomie dwie centriole (9 dupletów + 2 w środku: tylko jedna mikrotubula w parze kompletna: 13 dimerów). Zwykle znajduje się koło jądra komórkowego.
Tak samo w przypadku wici i rzęsek: wolno polimeryzujące i depolimeryzujące mikrotubule układzie 9 dubletów i 2 w centrum (ciałko podstawowe). Aksonema - zorganizowany zespół stabilnych mikrotubul w wici i rzęsce.
U roślin: brak centrioli, mikrotubule są generowane przez otoczkę jądrową, błonę komórkową, błonę organelli (tam właśnie znaleziono γ-tubulinę).
Na terenie ciałka podstawowego: 9 tripletów mikrotubul - brak centralnych.
Białka związane z mikrotubulami: białka motoryczne, które wiążą się z filamentami aktynowymi lub mikrotubulami i zużywają energię, aby przemieszczać się wzdłuż filamentu aktynowego czy też mikrotubuli w jednym kierunku. Energię uzyskują z powtarzających się cykli hydrolizy ATP (wiązanie, uwalnianie i ponowne wiązanie z MT). Oba typy białek motorycznych zbudowane są z 2 łańcuchów ciężkich (głów), którymi przemieszcza się po mikrotubulach przy użyciu ATP i z 2 łańcuchów lekkich (ogony), które związane są z substancjami transportowanymi - cargo.
Dyneina: ruch przebiega w kierunku „-”
Kinezyna: ruch przebiega w kierunku „+”
Oba typy białek motorycznych mikrotubul istnieja w wielu fomach, a każda taka forma transportuje inny rodzaj cargo
Wici oraz rzęski zawierają stabilne mikroktubule przemieszczane przez dyneinę rzęskową, która generuje ruch zginający rdzenia
Filamenty pośrednie: nie są zaangażowane w ruch (dlatego nie będziemy o nich zbyt dużo mówić), pełnią wyłącznie funkcję wzmacniającą, często łączą się (współpracują) z mikrotubulami.
Tworzenie: monomer (który jest podstawą filamentu pośredniego) + polimeraza → filamenty pośredni
Tworzą siec wewnątrz cytoplazmy otaczając jądro komórkowe i rozciągając się aż do krańców komórki (zabezpieczają komórkę przed mechanicznym stresem)
Filamenty pośrednie blaszki jądrowej leżą po jądrowej stronie wewnątrznej warstwy otoczki jądrowej, białka budujące filamenty to laminy
Demontaż i ponowne formowanie blaszki jądrowej po podziale komórki następuje w wyniku fosforylacji i defosforylacji lamin przez kinazy białkowe
Ich duża wytrzymałość odgrywa rolę przy tworzeniu połączeń międzykomórkowych (np. desmosomów).
Mikrofilamenty = filamenty aktynowe: ( występują głównie tuż pod błoną komórkową - kora komórki)
Budowa: (duże podobieństwo do mikrotubule)
aktyna to monomer, który po polimeryzacji tworzy filamenty aktynowe - F-aktynę (do 10% aktyny w komórkach mięśniowych, w pozostałych od 1% do 5%), jest bardzo stabilna ewolucyjnie (silnie konserwowana), 6 różnych genów aktyn: 3 aktyny α (w mięśniach), aktyna β (w niemięśniowych komórkach) i aktyna γ (w mięśniach gładkich)
Filamenty aktynowe rzadko występują w komórce pojedynczo, najczęściej są umiejscowione w poprzecznie powiązanych pęczkach i sieciach, które są znacznie silniejsze niż pojedyncze filamenty.
bieguny „+” i „-”, a polimeryzacja i depolimeryzacja podstawą ruchu);polimeryzacja od strony „+” („tępy koniec”, zachodzi w obecności jonów: Mg2+, Na+, K+.
poszczególne elementy łączą białka: cross-linking-proteins, np. fascyna
każdy wolny monomer aktynowy niesie ściśle związany trifosforan nukleozydu, w tym przypadku ATP, który jest hydrolizowany do ADP wkrótce po przyłączeniu monomeru aktyny do mikrofilamenty (hydroliza związanego ATP do ADP w filamencie aktynowym zmniejsza wytrzymałość połączeń między monomerami oraz stabilność polimeru - tak jak w MT)
Funkcja: kompleks aktyna + miozyna (różne klasy: I, II, V, zbudowana także z czterech łańcuchów: 2 ciężkich i 2 lekkich) bierze udział w:
biorą udział w ruchu cyrkulacyjnym wokół komórki pod błoną, wokół centralnej wakuoli; wakuola może być poprzedzielana mostkami cytoplazmatycznymi, w których też ma miejsce ruch
ruch wewnatrzkomórkowy nie musi być na bazie MF, ale także MT
w neuronie: w synapsie - MF, w aksonie - MT (koniec „+” ku zakończeniu aksonu)
Ruchu fibroblastów (wolne komórki, np. neutrofile):
wypychanie powierzchni komórki do przodu (polimeryzacja aktyny), wysuwane są:
lamellipodia - cienkie, blaszkowate wypustki zawierające przestrzenna siec filamentów aktynowych z końcami plus umiejscowionymi blisko błony komórkowej
filopodia - cienkie, sztywne, palczaste wypustki wysuwane także przez wiele komórek, zarówno na końcu prowadzącym, jak i gdziekolwiek na swojej powierzchni (także zorientowane tak, że ich końce plus skierowane na zewnątrz)
Wszystkie aktynozależne białka motoryczne należą do rodziny miozyn, wiążą one i hydrolizyja ATP, co dostarcza energie do ich ruchu wzdłuż mikrofilamentów aktynowych od końca minus do końca plus filamentu
Ruchu komórek mięśniowych (sarkomerów): dzięki ślizgowi główek miozynowych(zablokowanie obu końców MF odpowiednimi białkami) między szkieletem aktynowym
Mikrofilamenty aktynowe (z miozyną II) tworzą przewężenie pierwotne (zaciskanie się pierścienia)
Kontynuują ruch pęcherzyków: najpierw po mikrotubulach dzięki kinezynie, a potem po mikrofilamentach dzięki miozynie Transport wewnątrzkomórkowy w większości na bazie mikrofilamentów (w szczególności u zwierząt).
Białka motoryczne związane z mikrofilamentami: 10 klas rodzin genów różnych rodzajów miozyn (2 ciężkie łańcuchy - „głowy” do przesuwu po mikrofilamencie i „ogonek” do transportu). Białka motoryczne przekształcają energię chemiczną z ATP w energię mechaniczną
Podstawa ruchu: polimeryzacja i depolimeryzacja mikrotubul i mikrofilamentów
Transport: białka motoryczne przekształcają energię chemiczną z ATP w energię kinetyczną
RUCH AMEBOIDALNY (Amoeba proteus):
Budowa „aparatu ruchu” u ameby:
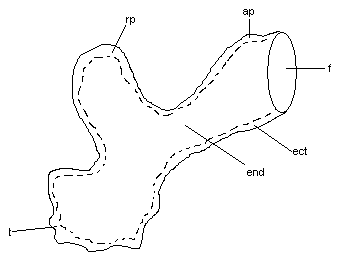
zróżnicowanie cytoplazmy na peryferyjny zżelifikowany cylinder ektoplazmatyczny (ect) i wewnętrzną płynną endoplazmę (end)
cylinder otwarty w strefie czołowej (f) wysuwanej nibynóżki (pseudopodia) (ap)
cylinder zamknięty w tyle (t)i na końcu nibynóżki wycofywanej (rp).
U ameby brak MT!
Płynność morfologiczna, anatomiczna (panta rhei)
Komórki heterotaktyczne - zawieszone w płynie, ruch, ale brak przemieszczania
Ameba nie ma narządów ruchu tylko tymczasowe pseudopodia (różny kształt, wielkość). Ruch wewnątrzkomórkowy i całej komórki są ze sobą skorelowane, ale nie jest tak, że ruch wewnętrzny powoduje lokomocję.
Aby była lokomocja muszą być spełnione warunki:
Polaryzacja komórki (F - front, T - uroid)
Adhezja do podłoża (punkt podparcia) (minipodia - przyczepiają się do podłoża, guzki adhezyjne - gotowość odruchu w różnych kierunkach)
Cytoszkielet (generacja siły niezbędnej do ruchu)
Teoria Masta:
Żel ektoplazmatyczny tworzy kurczliwy cylinder zamknięty z tyłu, a otwarty z przodu (porównanie do wyciskania pasty z tubki). Skurcz cylindra, powodując wzrost ciśnienia hydrostatycznego, tłoczy endoplazmę do przodu.
W amebie: aktyna, miozyna (nie ma tubuliny)!
Cytoszkielet w części ektoplazmatycznej łączy się z organellami, a przez białka błonowe łączy się z błoną (bo to ona jest dla niego podstawą ruchu)
Istniały jak dotąd 3 teorie ruchu ameb (dwie pierwsze nie prawdziwe):
teoria skurczu tylnego: kurczliwość ograniczona do korteksu tylnego (cc), korteks ścian bocznych niekurczliwy
teoria skurczu przedniego: żelfikująca się endoplazma ciągnie zawartość komórki (polimeryzacja, sol -> żel)
Teoria ogólnego kortykalnego (prof. Grembecki)
Skurcz izotoniczny w okolicach wycofywanych i izometryczny w przyrastających:
Rozdzielenie funkcji:
sterującej (nibynóżka wiodąca - określa kierunek, szybkość): skurcz izotoniczny: f-aktyna przekształca się w g-aktynę: zmiana kształtu → ciśnienie hydrostatyczne przepycha cytoplazmę do przodu - powoduje to pofałdowanie uroidu
motorycznej (kurczliwy korteks): skurcz izometryczny tam gdzie ameba przyczepia się do podłoża: odkleja się cytoszkielet o od błony i przefiltrowuje cytoplazmę do przodu (działa jak sito) (porównanie do otwartej tuby) (nie zmienia kształtu)
W wysuwanej nibynóżce - błona cytoplazmatyczna i cytoszkielet nie przylegają do siebie, cytoszkielet przefiltrowuje cytoplazmę -> elementy aktynowe przechodzą i tworzy się nowy cytoszkielet itd. (kolejne kroki)
skurcz w tylnej części komórki - solfikacja -> przepływ do części przedniej
pod samym frontem - miozyna I; mutant zaw. Tylko miozynę I wytwarza nóżki, ale się nie przemieszcza
Ameba kroczy (a nie płynie), średnio co 2 sekundy krok nibynóżki. Układ filamentów aktynowych przemieszcza się dzięki miozynie (miozyna II niezbędna do ruchu) wzdłuż tuby ektoplazmatycznej.
Depolimeryzacja aktyny w uroidzie, dimery do przodu i w nibynóżce polimeryzacja.
Białka towarzyszące:
α-aktynina i filamina: łączą filamenty w sieć - szkielet sieciowy
filamina, profilina i białka z rodziny żelsolin: zależy od nich polimeryzacja i depolimeryzacja
talina, winkuliny, α-aktynina, poksyna: stabilne połączenie cytoszkieletu z błoną w miejscach adhezji
miozyna I, spektryna, olkiny, pofiryna, MARKS, kompleks spektryna-ankilina: dynamiczne połączenia cytoszkieletu z błoną
Podczas pinocytozy - lejki pinocytarne, kiepska adhezja do podłoża. Fagocytoza - dobra adhezja do podłoża - fagosom.
6
Wyszukiwarka