Zagadnienia na egzamin z przedmiotu ,,BIOCHEMIA”
kier. Behawiorystyka
Wydziału Biologii i Hodowli Zwierząt UP w Lublinie
Wyjaśnić pojęcia: metabolizm, katabolizm, anabolizm, fosforylacja oksydacyjna, fosforylacja substratowa.
METABOLIZM- całokształt reakcji biochemicznych zachodzących w komórkach związany z przepływem: materii, energii i informacji genetycznej, zapewniający organizmowi wzrost, ruch, rozmnażanie, wrażliwość.
KATABOLIZM- dysymilacja; rozkład; ogół reakcji chemicznych metabolizmu prowadzący do rozpadu złożonych związków chemicznych na prostsze cząsteczki; energia wyzwolona z tych związków jest kumulowana w przenośniku ATP. Reakcja egzoenergetyczna, uwalniająca energię, w substratach jest więcej energii niż w produktach; np. oddychanie wewnątrzkomórkowe.
ANABOLIZM-asymilacja; przyswajanie; synteza; grupa reakcji chemicznych, w wyniku których z prostych substratów powstają związki złożone, gromadzące energię. Reakcje te wymagają dostarczenia energii, w wyniku czego w produktach jest więcej energii niż w substratach; np. biosynteza białek, cukrów, tłuszczy.
FOSFORYLACJA OKSYDACYJNA (UTLENIAJĄCA)- cykl reakcji przyłączania reszty kwasu ortofosforowego do związków chemicznych, połączone ze zmianą stopnia utlenienia atomu, do którego ta grupa bezpośrednio się przyłącza; przebiega w mitochondrium, wymaga obecności tlenu jako akceptora elektronów; energia uwalniana podczas utleniania zredukowanych nukleotydów przekształcana jest w energię ATP.
FOSFORYLACJA SUBSTRATOWA- reakcja chemiczna, podczas której reszta fosforanowa zostaje przeniesiona ze związku ufosforylowanego - substratu - bezpośrednio na ADP przez enzymy, najczęściej z grupy kinaz. Ten sposób wytwarzania ATP nie wymaga udziału tlenu i zachodzi np. w glikolizie oraz cyklu Krebsa. Jest wykorzystywana np. w mechanizmach skurczu mięśni, aktywnego transportu przez błony, przekazywania sygnału w komórce.
Biologiczna rola wody w organizmie.
Woda stanowi główny końcowy produkt metabolizmu spalania pokarmów. Służy albo jako reagent, albo jako produkt w wielu reakcjach. Stanowi doskonały rozpuszczalnik. 2/3 całkowitej wody w organizmie to płyn wewnątrzkomórkowy, reszta to płyn pozakomórkowy. Regulacja równowagi wodnej zależy głównie od podwzgórza i czynności nerek. Dodatkowo woda odznacza się dobrym przewodnictwem cieplnym i wysokim ciepłem parowania, co zabezpiecza organizm przed przegrzaniem i umożliwia sprawną termoregulacje; ma wysokie napięcie powierzchniowe, co sprawia, że spełnia ważną role w tworzeniu błon cytoplazmatycznych; stanowi środek transportu wewnątrzustrojowego, np.: produktów przemiany materii, hormonów ,witamin, enzymów; uczestniczy w regulacji temperatury, ciśnienia osmotycznego, pH; stanowi płynne środowisko, niezbędne do usuwania końcowych produktów przemiany materii.
Biopierwiastki i ich rola w procesach biochemicznych: sód, potas,
Sód (Na) -chroni przed utratą płynów, ma właściwości zasadowe, równoważy w organizmie gospodarkę kwasowo-zasadową i wpływa na gospodarkę wodno-elektrolitową, utrzymywanie prawidłowego ciśnienia osmotycznego, wchodzi w skład osocza, bierze udział w przewodzeniu impulsów nerwowych.
Potas (K) - wpływa na gospodarkę wodną organizmu, ciśnienie osmotyczne, równowagę kwasowo-zasadową i pobudliwość nerwowo-mięśniową, podwyższa stopień uwodnienia koloidów komórkowych, aktywator wielu enzymów; jest niezbędnym składnikiem do syntezy białek w rybosomach.
Biochemia wapnia regulacja przemiany wapnia w organizmie
Wapń występuje głównie w kościach, lecz mała jego część znajduje się w płynie pozakomórkowym, w którym połowa łączy się z białkiem, a druga połowa jest w postaci wolnej jako Ca2+. Biologicznie aktywny jest tylko wapń zjonizowany. W homeostazie wapniowej uczestniczą głównie 2 hormony: parathormon wydzielany przez przytarczyce i kalcytriol będący pochodną witaminy D3 powstający w nerkach. Nagły spadek Ca2+powoduje zwiększoną syntezę parathormonu, zaś wysoki poziom hamuje jego wydzielanie. Parathormon powoduje wzrost poziomu wapnia w surowicy krwi w wyniku uwalniania wapnia z kości. Dodatkowo pobudzając syntezę kalcytriolu w nerkach pobudza wchłanianie Ca2+ w jelitach., a także zmniejsza wydalanie wapnia przez nerki przez co wzrasta stężenie wapnia w płynie pozakomórkowym. Gdy dieta jest uboga w wapń, szczególnie obficie w nerkach powstaje kalcytriol, zaś wysoki poziom hamuje jego syntezę. Kalcytriol działa głownie na geny uczestniczące w regulacji transportu jonów wapnia do i z komórek kosmków jelitowych.
Fosfor - fityniany - fitazy - zwierzęta poli- i mono-gastryczne.
Fosfor to składnik kości, zębów, ATP, kwasów nukleinowych. Jego stężenie w surowicy jest regulowane resorpcją zwrotną w kanalikach nerkowych. Fityniany to sole kwasów fitynowych, które osłabiają wchłanianie minerałów (np. wapnia, magnezu, cynku) oraz uniemożliwiają ich wykorzystanie przez organizm. Fitazy to enzymy rozkładające wiązania fitynowe (organiczne połączenia kwasu fosforowego i alkoholu inozytolu), które są źle trawionym składnikiem pasz. Zwierzęta monogastryczne nie mając flory bakteryjnej nie mogą trawić włókna pokarmowego, fitynianów, tanin i substancji antyodżywczych. Należą do nich trzoda chlewna oraz drób. Zwierzęta poligastryczne mają żołądek składający się z czterech komór. Należą do nich bydło, kozy, owce.
Magnez - tężyczka pastwiskowa.
Za normę referencyjną przyjmuje się stężenie magnezu w surowicy krwi 0,78 - 1,23 mmol/l. Przy obniżeniu poziomu do 0,3 mmol/l może wystąpić lżejsza postać tężyczki, przy której obserwuje się zmniejszenie mleczności, spadek apetytu. Występują na przemian napady podniecenia, lękliwości z napadami depresji, którą cechują senność, a nawet śpiączka. Ostra postać tężyczki pastwiskowej występuje w przypadku stężenia magnezu we krwi do 0,2 a nawet do 0,1 mmol/l. Tutaj objawy są wyraźne w postaci drgawek i charakterystycznych ruchów wiosłowych kończyn.
Metabolizm żelaza.
Transferryna jest glikoproteiną syntetyzowana przez wątrobę. Odgrywa ona główną rolę w gospodarce ustrojowej żelazem, ponieważ transportuje je w układzie krążenia do miejsc, w których jest ono potrzebne, m. in. z jelita do szpiku i innych narządów. Wolne żelazo jest toksyczne, ale w połączeniu z transferryną jego toksyczność zostaje zmniejszona. Ferrytyna to kolejne ważne białko. W prawidłowych warunkach gromadzi żelazo, skąd może być ono pobrane w razie potrzeby. W przypadku nadmiaru żelaza więcej ferrytyny stwierdza się głównie w wątrobie i śledzionie. Synteza receptorów dla ferryty i transferryny jest zwrotnie uzależniona od zawartości żelaza w komórce.
Funkcja błon komórkowych
Stanowią granicę pomiędzy światem zewnętrznym a światem wewnętrznym komórki lub organellum.
Organizują komórkę i jej wnętrze: budują organella komórkowe i tworzą przedziały subkomórkowe.
Umożliwiają kontakt ze środowiskiem: odbieranie bodźców, pobieranie i wydalenia rozmaitych substancji i cząstek.
Przez błony odbywa się transport: na drodze dyfuzji, dyfuzji ułatwionej, transportu aktywnego oraz na drodze endocytoz i egzocytoz.
Odbierają i przewodzą bodźce.
W błonach odbywają się niektóre procesy biochemiczne jak: fosforylacja w fotosyntezie, łańcuch oddechowy w oddychaniu tlenowym.
Rodzaje transportu przez błony komórkowe. - transport bierny: filtracja, dyfuzja prosta, dyfuzja ułatwiona, osmoza; transport aktywny; endocytoza: pinocytoza, fagocytoza; egzocytoza - krótka charakterystyka.
Transport bierny- zachodzi zgodnie z gradientem stężeń, ładunków i ciśnień
Filtracja- polega na przedostawaniu się przez błonę wody i substancji w niej rozpuszczonych, których średnica jest mniejsza od średnicy porów w błonie; proces ten zachodzi dzięki różnicy ciśnień hydrostatycznych po obu stronach błony.
Dyfuzja prosta- przebiega zgodnie z zasadą dążności do wyrównania stężeń; cząstki przechodzą przez błonę z obszaru o większym stężeniu do obszaru o mniejszym stężeniu; dyfuzja ta zachodzi tym szybciej, im większa różnica stężeń.
Dyfuzja ułatwiona- biorą w niej udział białka zwane nośnikami; substancja przenikająca łączy się z nośnikiem i powstaje kompleks substrat-nośnik; po przeniesieniu substancji na drugą stronę nośnik zostaje uwolniony i może przenieść kolejną cząsteczkę; dyfuzja ułatwiona może zachodzić również za sprawą białek błonowych tworzących kanały przez które przechodzą jony nieorganiczne; kanały te wykazują selektywność jonową i nie SA ustawicznie otwarte.
Osmoza- dyfuzja rozpuszczalnika przez błonę półprzepuszczalną rozdzielającą dwa roztwory o różnym stężeniu. Zachodzi od roztworu o niższym stężeniu do roztworu o wyższym, czyli prowadzi do wyrównania stężeń obu roztworów. Roztwór z którego ubywa rozpuszczalnika nazywa się hipotonicznym, tego w którym przybywa nazywa się hipertonicznym. Gdy roztwory pozostają w równowadze osmotycznej, są wzajemnie izotoniczne względem siebie.
Transport aktywny- zachodzi wbrew gradientowi stężeń, ładunków i ciśnień; wymaga nakładu energii pochodzącej z rozkładu ATP.
Endocytoza- proces przenikania dużych cząsteczek (np. cholesterolu, polinukleotydów) do wnętrza komórki w wyniku powstawania wakuol, wraz z fragmentami błony komórkowej; dzięki niej odbywa się transport cieczy i cząsteczek; wyróżniamy w niej pino- i fagocytozę
Pinocytoza- podczas tego procesu pobierane są drobiny białek lub inne wielkocząsteczkowe substancje, które są rozpuszczalne w wodzie; ten sposób odżywiania polega na tworzeniu kanalików zakończonych banieczkami, w których znajduje się pobierana substancja - są to pęcherzyki pinocytarne; odrywają się one od błony komórkowej i poruszają się w cytoplazmie; pęcherzyki zostają wtedy w całości rozłożone enzymatycznie, przy udziale lizosomów, a następnie rozproszone w cytoplazmie; podczas pinocytozy transportowane są substancje płynne.
Fagocytoza- jest związana z wchłanianiem dużych cząsteczek (np. wirusów, bakterii); polega na pobraniu ze środowiska pokarmów stałych, odizolowaniu od cytozolu poprzez utworzenie wodniczki pokarmowej i trawieniu z udziałem lizosomów; w tym procesie nie następuje utrata błony komórkowej.
Egzocytoza- proces uwalniania metabolitów powstających wewnątrz komórki, a także produktów ubocznych metabolizmu i wydalin do przestrzeni pozakomórkowej; zachodzi na drodze fuzji pęcherzyków transportujących metabolity z błoną komórkową od wewnątrz, co skutkuje wyrzuceniem ich zawartości do środowiska pozakomórkowego.
Działanie i znaczenie pompy sodowa-potasowej.
Pompa sodowo potasowa jest białkiem wbudowanym asymetrycznie w błonę komórkową. Ma ona za zadanie utrzymywanie odpowiedniego stężenia jonów potasu i sodu po obu stronach błony. Uczestniczy w aktywnym transporcie substancji (wbrew gradientowi stężeń), dlatego wymaga nakładu energii pochodzącej z hydrolizy ATP. Pompa sodowo-potasowa wypompowuje z komórki sód, a przenosi do wnętrza potas. W każdym cyklu działania pompy na każde 3 wypompowane kationy sodu przypadają 2 wprowadzone kationy potasu. Za każdym razem zużywana jest jedna cząsteczka ATP. Aby nastąpiła hydroliza ATP wymagana jest obecność kationów Mg2+. Znaczenie pompy: transport jonów przeciwko gradientowi stężeń, utrzymanie różnicy stężeń jonów po obu stronach błony, utrzymanie potencjału błonowego, utrzymanie stałej osmolarności wnętrza komórki i jej objętości, napęd do wtórnego transportu czynnego innych związków.
Zasady klasyfikacji enzymów.
Enzymy dzielimy na:
- enzymy proste zbudowane wyłącznie z białka
- enzymy złożone zbudowane z części białkowej (APOENZYMU) i części niebiałkowej (KOENZYMU), które razem tworzą HOLOENZYM
Podział enzymów ze względu na działanie
1) hydrolazy powodują rozkład substancji na prostsze, przy czym zostaje przyłączona woda; proteazy, lipazy, ureazy
2) dehydrazy odszczepiające wodór
3) desmolazy powodują przerwanie łańcuchów węglowych
Podział enzymów ze względu na typ związków pokarmowych na które działają.
1) proteolityczne rozkładające białko
2) amylolityczne rozkładające skrobię i inne węglowodany
3) lipolityczne rozkładające tłuszcz
Podział enzymów ze względu na rozmieszczenie enzymów w przewodzie pokarmowym
• ptyalina w ślinie- rozkłada skrobię na glukozę; środowisko zasadowe
• amylaza ślinowa- podejmuje działanie ptialiny
• pepsyna w soku żołądkowym- rozkłada białko na albumozy i peptozy; środowisko kwaśnie (HCl)
• trypsyna w dwunastnicy- rozkłada cząsteczki białek na aminokwasy; środowisko zasadowe
• lipaza- rozszczepia tłuszcze na glicerynę i kwasy tłuszczowe
• enzym białkowy- erypsyna w jelicie cienkim- rozkłada albumozy i peptozy
Nomenklatura enzymów (zgodna z zaleceniami Międzynarodowej Unii Biochemicznej oraz potoczna).
Według MUB:
1. Oksydoreduktazy- przenoszą elektrony, protony i tlen (np. dehydrogenazy, oksydazy). AH2+B A+BH2 2.Transferazy- przenoszą grupy funkcyjne (np. aminotransferazy, kinazy). AB+C A+BC 3.Hydrolazy- reakcje hydrolizy (np. proteazy, celulaza). AB+ H2O A+B 4. Liazy- rozszczepiają wiązania (C-C, C-O-, C-N i inne) (np. dekarboksylazy aminokwasów). AB A+B
5. Izomerazy- przenoszą grupy w obrębie cząsteczki (np. izomeraza maleinianowa) ABBA
6. Ligazy- tworza wiązania sprzężone z hydrolizą ATP (np. syntetazy) A+BAB
Funkcje biologiczne enzymów.
- przyspieszają reakcje, jednak same nie ulegają przekształceniom w inne związki i nie ulegają szybko zużyciu w przeprowadzanych przez siebie reakcjach
- przeprowadzają reakcje z dużą szybkością w optymalnych dla siebie warunkach temperatury, pH i ciśnienia
- charakteryzują się dużą specyficznością substratową
- w czasie reakcji enzymatycznej nie tworzą się zbędne produkty uboczne
- nie wpływają na równowagę reakcji
Budowa chemiczna enzymów (holoenzym, apoenzym, koenzym, grupa prostetyczna).
HOLOEZNZYM- składa się z części białkowej (apoenzym) oraz niebiałkowej (koenzym)
APOENZYM- część enzymu, która po połączeniu z koenzymem stanowi holoenzym; apoenzym decyduje o swoistości enzymu oraz często o rodzaju reakcji jaką enzym jest zdolny katalizować
KOENZYM- niebiałkowa część enzymu; często może odłączać się od jednego enzymu i przyłączać się do 2.
GRUPA PROSTETYCZNA- koenzym, który jest ściśle związany z enzymem za pomocą wiązań kowalencyjnych lub niekowalencyjnych
Budowa centrum aktywnego enzymu oraz jej wpływ na aktywność katalityczną białka enzymatycznego.
Miejsce aktywne to miejsce wiązania substratu i przemieniania go w produkt. Względnie niewielka część enzymu, stanowiąca określoną trójwymiarową przestrzeń. Jest często szczeliną lub zagłębieniem, które tworzy środowisko w znacznym stopniu niepolarne. Substrat jest tutaj wiązany przez liczne słabe siły. Cząsteczki, które różnią się od określonego substratu, nie mogą zbliżyć się do centrum aktywnego na odległość wiązania i nie ulegają reakcji. Tak więc jeden enzym katalizuje jedną reakcję.
Mechanizm działania enzymu (wpływ substratu na strukturę enzymu, enzymy jako biokatalizatory obniżające energię aktywacji, rodzaje wiązań chemicznych pomiędzy enzymem i substratem).
Wpływ substratu na strukturę enzymu:
1.Utworzenie kompleksu enzym-substrat [ES], S+E [ES]
2. Przekształcenie kompleksu [ES] w kompleks enzym-produkt [EP], [ES] [EP]
3. Rozpad kompleksu [EP] na enzym i produkt [EP] E+P
Enzymy jako biokatalizatory obniżające energię aktywacji charakteryzuja się: sprawnością katalityczną - zdolnością przyspieszania rzędu 106 - 1012 razy; swoistością - może być względem substratu, względem reakcji lub względem reakcji i substratu; działaniem w łagodnych warunkach - niskie ciśnienie, temperatura i zakres łagodnych wartości pH.
Rodzaje wiązań chemicznych między enzymem i substratem: oddziaływania elektrostatyczne, wiązania wodorowe, siły van der Waalsa, oddziaływania hydrofobowe.
Czynniki wpływające na aktywność enzymu i ich mechanizm działania (pH, temperatura, stężenie substratu lub koenzymu, metabolity, aniony, kationy, inhibitory, aktywatory, leki).
1) pH: większość enzymów najszybciej działa w środowisku zbliżonym do obojętnego, czyli pH=7, ale każdy enzym ma optymalne pH działania, w którym szybkość katalizowanej przez niego rekcji jest maksymalna; małe odchylenia od optimum powodują spadek aktywności enzymu wywołany zmianami jonizacji grup w miejscu aktywnym, zaś większe prowadzą do denaturacji białka enzymatycznego w wyniku zakłócenia licznych oddziaływań niekowalencyjnych; laktaza 1-2, amylaza 6, lipaza 7, pepsyna +trypsyna 7.
2)temperatura: wzrost zwiększa energię termiczną cząsteczek substratu, co z kolei zwiększa szybkość reakcji; optimum dla większości białek to 37*C; nadmierny wzrost temp. (pow. 37*C) prowadzi do denaturacji enzymu
3)stężenie substratu lub koenzymu: przy małych stężeniach substratu jego podwojenie powoduje podwojenie początkowej szybkości; przy większych stężeniach substratu enzym ulega wysyceniu i dalszy wzrost substratu powoduje tylko małą zmianę wartości szybkości początkowej
4)inhibitory: hamują bądź spowalniają aktywność enzymów. Jeżeli inhibitor występuje w dostatecznie dużym stężeniu, to może całkowicie zablokować reakcję (przyłączenie substratu); z kolei zwiększenie stężenia substratu może spowodować wyparcie inhibitora
5) aktywatory: zwiększają lub poprawiają działanie enzymów przez odszczepienie od nieaktywnych proenzymów blokujących grup funkcyjnych lub na ochronie enzymów przed działaniem różnych czynników chemicznych
Co to jest aktywność właściwa enzymu i jak się ją wyraża?
Jednostka aktywności enzymatycznej odpowiada takiej ilości enzymu, która w ciągu 1 minuty katalizuje przemianę 1 μmola substratu w temp. 30*C w optymalnych warunkach reakcji.
Koenzymy oksydoreduktaz (dinukleotyd nikotynamido-adeninowy (NAD), fosforan dinukleotydu nikotynamido-adeninowego (NADPH), dinukleotyd flawino-adeninowy (FAD), ubichinon (koenzym Q, CoQ), kwas limonowy), koenzymy transferaz: biotyna, koenzym A, fosforan pirydoksalu (PLP), nukleotydy adenozowe (ATP, GTP)- budowa i rola fizjologiczna.
KOENZYMY OKSYDOREDUKTAZ
1)DINUKLEOTYD NIKOTYNOAMIDOADENINOWY NAD: składa się z adenozyno-5'-monofosforanu i nukleotydu nikotynoamidowego połączonych ze sobą wiązaniem bezwodnikowym; ROLA: uczestnictwo w przenoszeniu jonów H+ od utlenionego substratu do głównego akceptora, gł. w procesach utleniania komórkowego; odbywa się to za pośrednictwem łańcucha oddychania wewnątrzkomórkowego
2)FOSFORAN DINUKLEOTYDU NIKOTYNOAMIDOADENINOWEGO NADP: składa się z adenozyno-5'-monofosforanu i nukleotydu nikotynoamidowego połączonych ze sobą wiązaniem bezwodnikowym oraz reszty fosforanowej; ROLA: akceptor protonów i elektronów
3)DINUKLEOTYD PFLAWINOADENINOWY FAD: organiczny związek chemiczny złożony z mononukleotydu flawinowego i adenozynomonofosforanu: ROLA:uczestniczy w każdych reakcjach oksydoredukcyjnych związanych z przenoszeniem jonów H+ z utleniania substratów na łancuchach oddychania wewnątrzkomórkowego
4)UBICHINON (KOENZYM Q): organiczny związek chemiczny z grupy chinonów, występujący w mitochondriach komórek roślinnych i zwierzęcych; odpowiedzialny za przenoszenie elektronów w łańcuchu oddechowym; po przyłączeniu elektronów swobodnie porusza się w wewnętrznej błonie mitochondrialnej umożliwiając transport elektronów między kompleksami białek łańcucha oddechowego, które wbudowane są w wewnętrzną błonę mitochondrialną
5)KWAS LIMONOWY: kwas alfa-liponowy; koenzym rozpuszczalny w wodzie i w tłuszczach; silny przeciwutleniacz, bierze udział w metabolizmie pokarmów
KOENZYMY TRANSFERAZ
1)BIOTYNA: organiczny związek chemiczny o budowie heterocyklicznej występujący w organizmach zwierzęcych i roślinnych; koenzym kilku różnych enzymów; niezbędny składnik karboksylaz biotyno zależnych; uczestniczy w przenoszeniu grupy karboksylanowej (-COO-) z anionu wodorowęglanu na różne związki organiczne, zależnie od rodzaju danej karboksylazy
2)KOENZYM A: organiczny związek chemiczny powstający w organizmie z adenozynotrifosforanu, pantotenianu oraz cysteaminy, służący jako przenośnik grup acylowych
3)FOSFORAN PIRYDOKSALU PLP: organiczny związek chemiczny, aktywna forma witaminy B6; pełni funkcję koenzymu niezbędnego do działania enzymów odpowiedzialnych za metabolizm aminokwasów (aminotransferaz i dekarboksylaz); podczas transaminacji jest przekształcany w fosforan pirydoksaminy
4)ATP: organiczny związek chemiczny, nukleotydadeninowy zbudowany z grupy trójfosforanowej przyłączonej w pozycji 5' cząsteczki adenozyny, tworząc bezwodnik kwasu fosforowego; wchodzi w reakcje tylko w obecności kationów metali dwuwartościowych tworząc z nimi kompleksy; powst. w wyniku fosforylacji substratowej i w procesie chemiosmozy; jest czynnikiem fosofrylującym- łatwo odczepia 1 resztę kwasu ortofosforowego przekształcając się w ADP; wydzielana jest przy tym znaczna ilość energii; ATP berze udział w biosyntezie KT, reagując z aktyną i miozyną pow. skurcze mięśni
5)GTP: organiczny związek chemiczny, rybonukleotyd purynowy pełniący funkcję przenośnika energii w komórce; zawiera dwa wiązania wysokoenergetyczne; GTP bierze udział w reakcjach fosforylacji, a także dostarcza energię w procesie translacji i transkrypcji; powstaje w cyklu Krebsa
Związki wysokoenergetyczne ich budowa i znacznie.
Związki wysokoenergetyczne- związki chemiczne, które w komórkach służą jako przenośniki energii chemicznej, tj. wodoru i elektronów; związki zawierające "wiązanie wysokoenergetyczne" łatwo ulegające rozerwaniu co wiąże się z przenoszeniem grup fosforanowych lub innych; należą do nich: ATP, NADH, NADPH2, FADH2, acetylo-CoA.
Lokalizacja przemian cukrowych w różnych tkankach i narządach i ich powiązanie
Trawienie węglowodanów rozpoczyna się w jamie ustnej. W ślinie występuje enzym amylaza, który rozszczepia łańcuchy skrobi do maltozy. Następnie pokarm wymieszany ze śliną przechodzi do żołądka gdzie w środowisku kwaśnym pod wpływem jonów H+, rozkładane są dwucukrowce (sacharoza, laktoza). W wyniku tego oddziaływania powstaje glukoza, fruktoza i inne cukry proste.Cukry proste następnie przenikają do krwi i z krwią dostają się do wątroby. W wątrobie w wyniku różnych procesów przekształcają się w glikogen. Glikogen gromadzi się w wątrobie i różnych innych narządach, stanowiąc rezerwę cukrową dla całego organizmu. Gdy nastąpi niedobór glukozy we krwi, wątroba natychmiast oddaje potrzebną ilość glikogenu przerobionego na glukozę.
Acetylo CoA- jego rola w przemianach metabolicznych.
Acetylo-CoA odgrywa kluczową rolę w metabolizmie. Składa się z grupy acetylowej (-COCH3) związanej kowalencyjnie z koenzymem A. Jest on na przykład bezpośrednio wykorzystywany przez połączenie z kwasem szczawiooctowym do syntezy kwasu cytrynowego, który rozpoczyna cykl kwasu cytrynowego (kwasów trójkarboksylowych). W postaci aktywnego octanu kwas octowy łączy się także z choliną, tworząc acetylocholinę, lub z sulfonamidami, które przed wydaleniem ulegają acetylacji. Acetylo-CoA odgrywa również ważną rolę w metabolizmie lipidów - jest prekursorem cholesterolu, a tym samym hormonów steroidowych.
Znaczenie łańcucha oddechowego.
Elektrony w łańcuchu oddechowym wędrują przez białkowe kompleksy dzięki wzrastającej wartości potencjału oksydoredukcyjnego. 3 z tych kompleksów to pompy protonowe, które przekazując elektrony na kolejny akceptor jednocześnie powodują przejście protonów z matrix mitochondrialnego, do przestrzeni międzybłonowej. Tworzy się gradient protonowy (w przestrzeni międzybłonowej jest więcej H+ niż w matrix). Ta różnica pH po obu stronach błony staje się siłą napędową dla tworzenia ATP, ponieważ protony wracają do matrix, jedyną możliwą drogą - przez syntazę ATP, czyli transport elektronów jest niezbędny do wywołania gradientu protonowego, a ten konieczny do syntezy ATP.
Wyjaśnij różnice w przebiegu procesów fosforylacji oksydacyjnej i substratowej - podaj przykłady.
Źródłem energii w fosforylacji substratowej są zmiany wewnątrz substratu a w przypadku oksydacyjnej energia wydziela się w czasie transportu wodorów na tlen. Fosforylacja oksydacyjna zachodzi w obecności tlenu ,a substratowa nie. Fosforylacja oksydacyjna jest młodsza ewolucyjnie i najbardziej produktywna (powstaje więcej ATP). Przykład: substratowa synteza ATP przez przenoszenie reszty fosforowej z bogatego w energie pośrednika na ADP.
Wyjaśnij różnice w budowie i funkcjach NAD+ i NADP+.
Cząsteczka NAD +jest dinukleotydem składającym się z adenozyno-5'-monofosforanu i nukleotydu nikotynoamidowego połączonych ze sobą wiązaniem bezwodnikowym. Cząsteczka NAD+ wiąże jeden proton i dwa elektrony. Miejscem ich działania jest amid kwasu nikotynowego. Drugi proton pozostaje w środowisku reakcji. Cząsteczka NADP+ różni się do NAD+ obecnością reszty fosforanowej przy węglu 2' rybozy nukleotydu adeninowego. NADP+ jest także akceptorem protonu i elektronów w reakcjach redukcji. Rola NAD+ : przeważnie oddaje swój wodór enzymom łańcucha oddechowego. Reakcja z tlenem prowadząca do powstania H2O jest przy tym wykorzystywana do syntezy ATP. Rola NADH+ : służy do tego, aby dostarczyć przy biosyntezach potrzebnego wodoru lub ogólnie „równoważników redukcyjnych”.
Zasady dotyczące funkcjonowania łańcucha oddechowego.
1.Składniki łańcucha oddechowego różnią się powinowactwem do elektronów- wzrasta ono w miarę przebiegu łańcucha
2.Powinowactwo do elektronów = potencjał oksydacyjno-redukcyjny = energia swobodna
3. Elektrony ( z NADH i FADH2 )wchodzą w łańcuch oddechowy z wysoką energią, którą w trakcie transportu tracą
4. W miejscach, w których uwalniana jest wystarczająca ilość energii dochodzi do pompowania protonów
5. O powinowactwie do elektronów, a więc ilości uwalnianej energii, decydują głównie elementy niebiałkowe, tkwiące w kompleksach łańcucha oddechowego
Trawienie i wchłanianie węglowodanów.
Trawienie skrobi i glikogenu rozpoczyna amylaza ślinowa. Jej działanie jest najsilniejsze przy pH 6-7. W ślinie występuje przede wszystkim α-amylaza. Pod jej wpływem następuje dekstrynizacja skrobi połączona z odrywaniem cząsteczek maltozy i niewielkiej liczby cząsteczek glukozy. W żołądku następuje zahamowanie trawienia, gdyż wzrost stężenia kwasu solnego powoduje denaturację amylazy. Dalsze etapy trawienia skrobi zachodzą w jelicie cienkim. Do dwunastnicy wydzielana jest z sokiem trzustkowym amylaza trzustkowa, działająca podobnie do amylazy ślinowej jednak w wyższym pH. Enzym ten kończy trawienie skrobi i glikogenu, przekształcając niestrawione dotychczas fragmenty tych cukrów w maltozę. W soku jelitowym występują liczne glikozydazy, takie jak maltaza, laktaza i sacharaza, które powodują rozszczepienie dwucukrów do cukrów prostych. Laktaza umożliwia hydrolizę laktozy do cukrów prostych: glukozy i galaktozy. Maltaza powoduje hydrolizę maltozy do glukozy. Sacharoza powoduje hydrolizę sacharozy do glukozy oraz fruktozy. Cukry proste są wchłaniane do krwiobiegu głównie w początkowym odcinku jelita cienkiego. Cukry proste nie podlegają biernej dyfuzji, lecz transportowane są za pomocą przenośnika, znajdującego się wewnątrz błony i w sposób specyficzny transportującego cząsteczki monosacharydów do wnętrza komórek nabłonka jelitowego.
Cukry proste wchłaniane są przez kosmki jelitowe, dostają się do naczyń krwionośnych a układ zwrotny prowadzi je do wątroby, gdzie glukoza ulega zamianie na glikogen i w tej postaci jest tam magazynowana.
Przebieg glikolizy - szlak EMP (glikoliza tlenowa i beztlenowa), bilans energetyczny.
Glikoliza to proces enzymatycznego rozkładu cukrów przekształcający 1 cząsteczkę glukozy w 2 cząsteczki pirogronianu z jednoczesnym wytworzeniem ATP; dostarcza substratów do cyklu Krebsa i fosforylacji oksydacyjnej.
Pierwszy etap glikolizy polega na fosforylacji glukozy lub innych heksoz i przekształceniu do fruktozo-1,6 bisfosforanu. Następne reakcje polegają na rozszczepieniu fruktozo-1,6 bisfosforanu do 2 cząsteczek trójwęglowych : aldehydu 3- fosfoglicerynowego i fosfodihydroksyacetonu. Triozy te łatwo przechodzą jedna w drugą. W fazie tej zużyte jest ATP.
Drugi etap glikolizy dostarcza użytecznej energii w postaci NADH i ATP. Polega on na utlenieniu aldehydu 3- fosfoglicerynowego do kwasu 3- fosfoglicerynowego, który jest źródłem fosforu.
Ostatni etap glikolizy polega na odłączeniu fosforanu od 3- fosfoglicerynianu. Powstaje 2- fosfoglicerynian, który zostaje przekształcony w 3- fosfoenolopirogronian. Od 3- fosfoenolopirogronianu odłączana jest reszta fosforanowa i powstaje pirogronian - ostatni produkt glikolizy.
W warunkach tlenowych cząsteczka pirogronianu ulega dekarboksylacji i powstaje acetylo-CoA, który zostaje włączony do cyklu Krebsa, po czym pośredniki trafiają do układu oddechowego.
W warunkach beztlenowych pirogronian zostaje zredukowany do produktów fermentacji, takich jak kwas mlekowy.
W tkankach występuje heksokinaza, która ma duże powinowactwo do glukozy i dzięki temu zapewnia dostarczanie cukru nawet przy niskim poziomie we krwi. W wątrobie natomiast występuje glukokinaza. Ma ona mniejsze powinowactwo do glukozy. Służy do wyciągania nadmiaru cukru z krwi po posiłkach w celu zmagazynowania go w postaci glikogenu lub zamiany na tłuszcz.
Bilans glikolizy w warunkach beztlenowych to 2 mole ATP z jednego mola glukozy, a w warunkach tlenowych 8 moli ATP.
Losy końcowych produktów glikolizy w warunkach tlenowych i beztlenowych.
W warunkach tlenowych pirogronian, otrzymany w wyniku glikolizy pobierany jest przez mitochondria , w których ulega dekarboksylacji do acetylo-CoA i zostaje utleniony do dwutlenku węgla w cyklu Krebsa . Powstałe w glikolizie równoważniki redukujące są przenoszone z NADH + H+ do wnętrza mitochondriów.
W warunkach beztlenowych uniemożliwiona staje się reoksydacja NADH w łańcuchu oddechowym przez przeniesienie równoważników redukujących na tlen. Pirogronian ulega redukcji przez NADH do mleczanu w reakcji katalizowanej przez dehydrogenazę mleczanową . Reoksydacja NADH w reakcji powstawania mleczanu przez odtworzenie NAD potrzebnego w następnym cyklu reakcji umożliwia dalszy przebieg glikolizy w przypadku nieobecności tlenu.
Reakcja utleniania w glikolizie.
W glikolizie reakcji utleniania podlega aldehyd 3-fosfoglicerynowy . Reakcja katalizowana przez dehydrogenazę aldehydu 3 fosfoglicerynowego jest sumą dwóch procesów utleniania aldehydu z udziałem NAD do kwasu karboksylowego oraz połączenia kwasu karboksylowego i ortofosforanu z utworzeniem acetylofosforanu.
Enzymatyczna regulacja procesu glikolizy.
1) Aktywność heksokinazy jest hamowana przez produkt glukozo-6-fosforan
2) reakcja katalizowana przez fosfofruktokinazę, enzym hamowany jest allosterycznie przez wzrost stężenia ATP, cytrynianu, H+; aktywowany przem AMP
3) Kinaza pirogronianowa kontroluje wpływ metabolitów glikolizy, enzym jest hamowany przez wysokie stężenie ATP oraz pośrednio przez niskie stęż. glukozy; aktywowany przez fruktozo-1,6-bifosforan.
Hormonalna regulacja procesu glikolizy.
Insulina- hormon wydzielany przez komórki β trzustki
1) Zwiększa wychwyt glukozy przez mięśnie i tkankę tłuszczową
2) Nasila syntezę glikogenu
3) Hamuje proces glikogeogenezy
4) Nasila syntezę kw. tłuszczowych i tryacylogriceroli
5) zwiększa wychwyt aminokwasów przez mięśnie i aktywuje syntezę białek mięśniowych
6) Hamuje degradację bialek
Glukagon- hormon wydzielany przez komórki α trzustki w odpowiedzi na niskie stężenie cukru we krwi
1) Stymuluje rozkład glikogenu wątrobowego (za pośrednictwem cAMP) i glukoneogenezy
2) Hamuje glikolizę oraz syntezę kw. tłuszczowych
Znaczenia glikolizy w funkcjonowaniu organizmu.
- podstawowa droga katabolizmu glukozy do AcCoA
- główny szlak metabolizmu pokarmowego fruktozy i galaktozy
- dostarcza dużo cząst. ATP- glikoliza tlenowa
- dostarcza produktów do syntezy biomolekuł- glicerol
-może zachodzić w warunkach beztlenowych energia dla tkanki mięśniowej w stanie niedotlenienia
- źródło energii dla komórek pozbawionych możliwości oddychania tlenowego pozbawionych mitochondriów (np. erytrocyty)
W jakim procesie całkowitemu utlenieniu ulega Acetylo-CoA? Omówić ten proces.
Acetylo-CoA ulega całkowitemu utlenieniu w procesie cyklu Krebsa=cyklu kwasu cytrynowego składającego się z 8 etapów, przebiegającego w matrix mitochondrium:
1. Acetylo-CoA ulega reakcji kondensacji ze szczawiooctanem do cytrynianu pod wpływem syntazy cytrynianowej
2.Cytrynian przekształca się w izocytrynian podczas izomeryzacji katalizowanej przez akonitazę
3.Izocytrynian utlenia się do alfa-ketoglutaranu pod wpływem dehydrogenazy izocytrynianowej
4.Alfa-ketoglutaran utlenia się do bursztynylo-CoA i CO2 przez kompleks dehydrogenazy alfa-ketoglutaronowej
5.Bursztynylo-CoA przekształca się w bursztynian przez syntetazę bursztynylo-CoA
6.Utlenienie bursztynianu do fumaranu przez dehydrogenazę bursztynianowi
7.Fumaran przekształca się w jabłczan przez fumarazę
8. Jabłczan utlenia się do szczawiooctanu przez dehydrogenazę jabłczanową
Pochodzenie grupy acetylowej potrzebnej do powstania Acetylko CoA
Reszty te mogą się brać z metabolizmu cukrowców - kwas pirogronowy ulega oksydacyjnej dekarboksylacji w wyniku czego powstaje acetylo-CoA. Również szkielety węglowe szeregu aminokwasów mogą ulegać przekształceniu do acetylo-CoA. Grupa acylowa pochodzi też z kwasu palmitynowego, acetooctanu.
Kierunki przemian kwasu pirogronowego.
Kwas pirogronowy jest punktem węzłowym w przemianie cukrów. Jego dalszy metabolizm. może przebiegać w wielu kierunkach. Podstawowe to: różnorodne fermentacje, dekarboksylacja oksydacyjna prowadząca do aktywnego octanu, przekształcenie w kwas mlekowy. Kwas pirogronowy może stać sie również substratem do syntezy glukozy.
Dekarboksylacja oksydacyjna kwasu pirogronowego.
W wyniku oksydacyjnej dekarboksylacji pirogronian jest utleniany przez dehydrogenazę pirogronianową złożoną z trzech enzymów i pięciu koenzymów wykorzystując NAD+ zredukowany do NADH, wskutek czego powstaje acetylo-CoA i CO2.
Dlaczego w komórkach mięśniowych powstaje niekiedy kwas mlekowy.
Zużyty podczas glikolizy NAD+ musi zostać zregenerowany jeżeli glikoliza ma przebiegać w dalszym ciągu. Przy ograniczonej ilości tlenu, np. w mięśniach podczas energicznego skurczu reoksydacja NADH do NAD+ przez łańcuch transportu elektronów staje się niewystarczająca, aby podtrzymać glikolizę. W tych warunkach NAD+ jest regenerowany podczas przemiany pirogronianu w mleczan działaniem dehydrogenazy pirogronianowej.
Cykl Krebsa - przebieg, rola biologiczna, bilans energetyczny.
PRZEBIEG:
1. Acetylo-CoA ulega reakcji kondensacji ze szczawiooctanem do cytrynianu pod wpływem syntazy cytrynianowej
2.Cytrynian przekształca się w izocytrynian podczas izomeryzacji katalizowanej przez akonitazę
3.Izocytrynian utlenia się do alfa-ketoglutaranu pod wpływem dehydrogenazy izocytrynianowej
4.Alfa-ketoglutaran utlenia się do bursztynylo-CoA i CO2 przez kompleks dehydrogenazy alfa-ketoglutaronowej
5.Bursztynylo-CoA przekształca się w bursztynian przez syntetazę bursztynylo-CoA
6.Utlenienie bursztynianu do fumaranu przez dehydrogenazę bursztynianowi
7.Fumaran przekształca się w jabłczan przez fumarazę
8. Jabłczan utlenia się do szczawiooctanu przez dehydrogenazę jabłczanową
ROLA:
Główną funkcją jest utlenianie pirogronianu (wytwarzanego podczas rozkładu glukozy w procesie glikolizy) do CO2 i H2O z jednoczesnym uzyskiwaniem energii. Cykl ten również odgrywa rolę w wytwarzaniu prekursorów dla szlaków biosyntez
BILANS:
Podczas każdego obotu powstają 3 cząst. NADH, 1 cząst. GTP i 1 cząst. FADH2, czyli razem 12 cząsteczek glukozy pomnożone przez 2 ( bo powstają 2 cząst. acetylo-CoA).
Wykazać powiązanie cyklu Krebsa z łańcuchem oddechowym.
Cykl Krebsa dostarcza substratu niezbędnego do przeprowadzenia łańcucha oddechowego, tj. NADH2 i FADH2.
Regulacja przebiegu cyklu Krebsa
* Ogólna: dostępność ADP, szybkość zużywania ATP
* Wewnetrzna:
- dehydrogenaza α-heteroglutanowa
- syntaza cytrynianowa- hamowana przez ATP i długołańcuchowe KT
- dehydrogenaza izocytrynianowa- hamowana przez ATP i NADH
- dehydrogenaza bursztynianowa- hamowana przez szczawiooctan
- jabłczanowa dehydrogenaza- stosunek [NADH]/[NAD+]
* Hormony:
- hormon wzrostu (GH)
- hormon kortykotropowy (ACTH)
Etapy pobierania energii z pożywienia.
Duże cząsteczki z pożywienia małe cząsteczki
Małe cząsteczki kilka prostych jednostek (wytworzenie energii użytecznej w postaci ATP)
AcetyloCoA 2x CO2 (cykl Krebsa) (utworzenie zredukowanej formy NADH/FADH, które ulegają reoksydacji)
Biosyntetyczna rola cyklu Krebsa
- AcetyloCoA, który jest włączany do cyklu powstaje tez przy rozpadzie białek, cukrów i tłuszczów cykl Krebsa jest ostatecznym wspólnym etapem rozpadu tych związków
- metabolity pośrednie wiążą cykl Krebsa z różnymi przemianami innych związkow
* synteza układu porfirynowego
* przemiany aminokwasów
* proces glikolizy
- CO2 wykorzystany do syntezy zasad azotowych, biosyntezy mocznika i procesów karboksylacji
- dostarczenie komórce zredukowanych koenzymów (NADH, FADH)
Syntezy i rozkład glikogenu.
SYNTEZA: przeprowadzana przez 3 enzymy
1. Pirofosforylaza UDP-glukozy katalizuje syntezę UDP-glukozy z UTP i glukozo-1-fosforanu
2.Syntaza glikogenowa używa UDP-glukozy jako substratu, dodając pojedyncze reszty do nieredukującego końca cząsteczki glikogenu i tworząc wiązanie alfa-1,4-glikozydowe między sąsiadującymi resztami. Syntaza może tylko wydłużać łańcuch, do rozpoczęcia syntezy potrzebny jest inicjator- glikogenina (białko mające 8 reszt glukozy połączonych wiązaniami alfa-1,4.
3. Enzym rozgałęziający zrywa w łańcuchu glikogenowym wiązania alfa-1,4 i przenosi fragmenty zbudowane z około 7 reszt ku wnętrzu cząsteczki, wiążąc je z głównym łańcuchem wiązaniem alfa-1,6, tym samym tworzy rozgałęzienia.
ROZKŁAD: przeprowadzany przez 2 enzymy
1.Fosforylaza glikogenowa rozbija wiązania alfa-1,4 i usuwa reszty glukozy z nieredukującego końca cząsteczki glikogenu, uwalniając jako produkt glukozo-1-fosforan
2.Enzym usuwający rozgałęzienia glikogenu usuwa wiązania alfa-1,6-glikozydowe tworząc również glukozo-1-fosforan, który zostaje przekształcony przez fosfoglukomutazę fo glukozo-6-fosforanu
Glikoneogeneza - przebieg, znaczenie.
SCHEMAT
Glukoneogeneza:
- proces syntezy glukozy z mleczanu i zw. nie cukrowych
- zachodzi w wątrobie, w niewielkim stopniu w korze nerek
- 3 etapy glikolizy nie mogą zostać odwrócone, potrzebne są do tego inne enzymy
- zachodzi w cytoplazmie (poza karboksylacją pirogronianu)
- zużyte zostaje 6 ATP
Wskaż reakcje różnicujące przebieg glikolizy i resyntezy glukozy.
W glikolizie są trzy nieodwracalne reakcje:
1) pirogronianszczawiooctanfosfoenolopirogronian
2) gruktozo-1,6-difosforanfruktozo-6-fosforan
3) glukozo-6-fosforanglukoza
Reakcje dekarboksylacji, odwodorowania w cyklu Krebsa.
1) izocytrynian(dekarbosylacja/dehydrogenacja) α-heteroglutan
2) α-hetoglutaran(dekarboksylacja/dehydrogenacja) bursztynylo CoA
3) bursztynian(dechydrogenacja) furman
4) jabłczan(dechydrogenacja) szczawiooctan
Cykl pentozanofosforowy - schemat, znaczenie.
Podstawowym zadaniem szlaku jest utlenianie glukozo-6-fosforanu do rybozo-5-fosforanu potrzebnego do syntezy DNA, RNA, FAD, ATP i in., a więc przekształcanie pentoz w heksozy, a także wytwarzanie NADPH. Przebiega w 3 etapach:
1)Reakcje utleniania przekształcające glukozo-6-fosforan w rybulozo-5-fosforan z wytworzeniem 2-óch cząsteczek NADPH. Glukozo-6-fosforan jest utleniany przez dehydrogenazę glukozo-6-fosforanową do 6-fosfoglutano-sigma-laktonu, a ten jest hydrolizowany przez laktonazę do 6-fosfoglukanianu. Degydrogenaza 6-fosfoglukanianowa przekształca 6-fosfoglukanion w rybulozo-5-fosforan.
2)Izomeryzacja rybulozo-5—fosforanu do rybozo-5-fosforanu pod wpływem izomerazy pentozofosforanowej.
3)Powiązanie szlaku pentozo fosforanowego z glikolizą przez działanie transketolazy i trans aldolazy. Transketolaza i trans aldolaza przekształcają rybozo-5-fosforan do fruktozo-6-fosforanu i aldehydu 3-glicerynowego.
Hormonalna regulacja przemiany węglowodanowej.
Za duży poziom cukru wywołuje wydzielanie insuliny, stymulującej wykorzystywanie szybkie glukozy poprzez komórki. Ten hormon, poprzez przyspieszenie transportu cukru do różnych komórek, intensyfikuje procesy utleniania cukru a także pobudza wytwarzanie glikogenu w mięśniach i wątrobie oraz przemianę cukrów w tłuszcze (tzw. litogeneza). Glukagon działa antagonistycznie, wydzielany jest po zmniejszeniu ilości cukru we krwi. Związek ten uruchamia węglowodanowe rezerwy wątroby, tym sposobem zwiększając stężenie cukru którego poziom powraca do normalnej wartości. Horomon wzrostu, hormon kortykotropowy zwiększają stężenie glukozy we krwi poprzez zmniejszanie pobierania glukozy przez niektóre tkanki. Glikokortykosteroidy powoduje wzmożoną glukoneogenezę, a także hamują zużywanie glukozy w tkankach pozawątrobowych. Adrenalina prowadzi do glikogenolizy w wątrobie i mięśniach. Tyroksyna ma działanie diabetogenne.
Schemat blokowy całkowitego utleniania glukozy.
Harper, strona 219 Jeśli teraz podliczymy wszystkie cząsteczki ATP, jakie mogą powstać przy spalaniu 1 cząsteczki glukozy, to uzyskamy: 8 ATP (glikoliza) + 2*3 ATP (pirogronianacetylo-CoA) + 2*12 ATP (cykl Krebsa). Razem daje to 38 cząsteczek ATP.
Glikogeneza i glikoliza- przebieg, znaczenie biologiczne.
GLIKOGENOGENEZA: proces syntezy glikogenu, w którym cząsteczki glukozy są dodawane kolejno do łańcuchów glikogenu w celach gromadzenia zapasów do spożytkowania w przyszłości; składa się z szeregu etapów:
1)glukoza pod wpływem gluko- lub heksokinazy przechodzi w glukozo-6-fosforan
2)glukozo-6-fosforan pod wpływem fosfoglukomutazy przechodzi w glukozo-1-fosforan
reszta j.w. w pkt. 54
GLIKOLIZA: j.w. w pkt. 43
Trawienie i wchłanianie tłuszczowców w przewodzie pokarmowym
Trawienie: polega na emulgowaniu tłuszczów i ich hydrolizie na wolne kwasy tłuszczowe i glicerol. Rozpoczyna się w jamie ustnej. Lipaza ślinowa działająca w pH 4-4,5 odczepia krótko łańcuchowe kwasy tłuszczowe, kontynuuje swoje działanie w żołądku, po dostaniu się do niego kęsa pokarmowego. Działa tak długo aż pH kęsa obniży się do ok. 1. W żołądku lipaza żołądkowa atakuje wiązanie estrowe naturalnie zemulgowanych tłuszczów. Zasadnicze trawienie rozpoczyna się w dwunastnicy, gdzie lipaza trzustkowa w połączeniu z kolipazą działające w pH 8 hydrolizuja wiązania triacylogliceroli, dzięki czemu powstają wolne kwasy tłuszczowe i monoacyloglicerole. Fosfolipaza aktywowana pzez trypsynę i jony Ca2+ odczepia kwasy tłuszczowe od fosfolipidów; esteraza karboksylowa aktywowana przez żółć i jony Ca2+ odczepia kwasy tłuszczowe połączone z cholesterolem. Sole żółciowe wydzielane przez wątrobę emulgują tłuszcze i tworzą z fosfolipidami micele. Natomiast HCO3- zapewniają optymalne pH środowiska dla enzymów lipolitycznych. Micele stanowią rodzaj transportera nierozpuszczalnych w wodzie produktów lipolitycznych (kw. tłuszczowe i monoacyloglicerole) z miejsca ich powstawania na powierzchni kropelek tłuszczowych do powierzchni błony śluzowej jelita. Monoacyliglicerole, cholesterol, kwasy tłuszczowe z miceli wchodzą drogą biernej dyfuzji do komórek błony śluzowej-enterocytów. Wchłanianie zachodzi w jelicie cienkim, największe w jelicie czczym i krętym.
Transport lipidów w organizmie i pomiędzy tkankami.
Z racji tego, że lipidy są nierozpuszczalne w wodzie, aby mogły być transportowane między tkankami w środowisku wodnym osocza, niepolarne lipidy muszą połączyć się z amfipatycznymi lipidami i białkami - powstają lipoproteiny mieszające się z wodą. Są 4 główne grupy lipoprotein:
chylomikrony transportują lipidy będące produktem trawienia i wchłaniania
lipoproteiny o bardzo małej gęstości (VLDL) transportują triacyloglicerol z wątroby
lipoproteiny o małej gęstości (LDL) mają dużą zawartość cholesterolu, powstają w wyniku metabolizmu VLDL
lipoproteiny o dużej gęstości (HDL) też bogate w cholesterol, ale zaangażowane w usuwanie cholesterolu z tkanek i uczestniczące w metabolizmie innych lipoprotein
Katabolizm triacyloglicerolu w komórce.
1. Lipoliza triacyloglicerolu do glicerolu i kwasów tłuszczowych
2. Fosforylacja glicerolu i utlenienie do fosfodihydroksyacetonu
3.Aktywacja kwasów tłuszczowych do acylo-CoA
Katabolizm kwasów tłuszczowych w komórce.
Kwasy tłuszczowe w komórce rozpadają się w procesie b-oksydacji polegającej na utlenieniu długołańcuchowych kwasów tłuszczowych, czemu towarzyszy wytarzanie ATP. KT są przekształcane w pochodne w postaci acylo-CoA, z których następnie od końca łańcucha acylowego są usuwane dwuwęglowe jednostki acetylo-CoA, powst. NADH i FADH2.
1.Utlenienie acylo-CoA do enoilo-CoA katalizowane przez dehydrogenazę acylo-CoA
2.Uwodnienie do 3-hydroksyacylo-CoA katalizowane prze hydratazę enoilo-CoA
3.Utlenienie3-hydroksyacylo-CoA do 3-ketoacylo-CoA katalizowane przez dehydrogenazę hydroksyacylo-CoA
4.Tioliza 3-ketoacylo-CoA do acetylo-CoA i acylo-CoA katalizowane przez b-ketotiolazę
Aktywacja kwasów tłuszczowych.
Rozpad KT zachodzi w cytozolu komórek prokariotycznych i w matrix mitochondrialnej komórek eukariotycznych. Zanim KT dotrą do matrix ulegają aktywacji przez utworzenie wiązania tioestrowego z CoA. Reakcja ta zużywa 1 cząst. ATP i jest katalizowana przez syntezę acylo-CoA. Nieodwracalna.
Bilans energetyczny β-oksydacji.
Jest to cykl następujących po sobie 4 reakcji, w wyniku których nasycony łańcuch węglowodorowy grupy acetylowej jest skracany o 2 atomy węgla oraz powstaje FADH2, NADH i acetylo-CoA. W każdym cyklu reakcji acetylo-CoA ulega skróceniu o 2 atomy węgla.
Obliczanie bilansu energetycznego:
Cn~acetylo-CoA + FAD + NAD +H2O + CoA Cn~acetylo~CoA + FADH2 + NADH + acetylo~CoA + H+
Degradacja palmitoilo-CoA wymaga 7 cykli reakcji:
Palmitoilo-CoA + 7FAD +7NAD+ + 7H2O 7CoA 8acetylo-CoA+ 7FADH2 +2NADH+ + 7H+
1)czasie 1 obrotu:
FADFADH+ 2 ATP w łańcuchu
NADNADH+H+ 3 ATP oddechowym
ZYSK: 5 ATP
2)aktywacja kw. tłuszczowego
Zachodzi w błonie mitochondrialnej
STRATA: 2ATP
3)końcowe utlenianie acetylo-CoA w Cyklu Krebsa:
3NADH+ H+ 3x3 ATP=9 ATP
1FADH2 1x2 ATP=2 ATP
11 ATP w ł. oddech.
1GTP 1 ATP
ZYSK: 12 ATP
4)utlenienie kw. tłuszczowego
Parzystowęglowego (np. kw. palmitynowy)
16 atomów węgla- 7 obrotów spirali Lynena
5 ATPx7 obrotów= 35 ATP
8 cząst. Acetylo-CoAx12 ATP= 96 ATP
131 ATP
131 ATP-2 ATP (aktywacja)= 129 ATP
całkowity zysk utlenienia kw. palmitynowego
Co to jest spirala Lynena ?
Spirala Lynena to inna nazwa b-oksydacji.
β oksydacja nienasyconych kwasów tłuszczowych.
*) podwójne wiązanie przy nienasyconym atomie węgla
- 3 cykle są podobne do β-oksydacji nasyconych kwasów tłuszczowych
- w 3 cyklu pojawia się cis-Δ3-enoilo-CoA(izomeracja) trans-Δ2-enoilo-CoA
- trans-Δ2-enoilo-CoA ulega dalszym reakcjom
*) podwójne wiązanie przy parzystym atomie węgla
-w jednym z etapów β-oksydacji powstaje 2,4-dienolany związek pośredni
- 2,4-dienolany związek pośredni(reduktaza 2,4-diemolo-CoA) cis-Δ3-enoilo-CoA
- cis-Δ3-enoilo-CoA(izomeraza) trans-Δ2-enoilo-CoA
- trans-Δ2-enoilo-CoA ulega dalszym reakcjom
Regulacja metabolizmu kwasów tłuszczowych.
Szybkość rozpadu KT jest regulowana przez dostępność wolnych KT we krwi, które z kolei powstają w wyniku lipolizy triagliceroli.
Biosynteza kwasów tłuszczowych.
Zachodzi w cytozolu. Składa się z kilku etapów:
1)Karboksylacja acetylo-CoA do malonylo-CoA katalizowana przez karboksylazę acetylo-CoA zawierającą biotynę przy wykorzystaniu HCO3-
2)Acetylo-CoA i malonylo-CoA zostają przekształcone w ich ACP-pochodne: acetylo-ACP i malonylo-ACP przy udziale transacylazyzcetylowej i transacylazy malonylowej
3)Następnie następuje cykl elongacji: kondensacja->redukcja->odwodnienie->redukcja aż do momentu powstania palmitynianu; dalsza elongacja zachodzi na powierzchni gładkiego retikulum endoplazmatycznego.
Etapy elongacji w syntezie kwasów tłuszczowych
1. KONDENSACJA acetylo-ACP i malonylo-ACP do acetoacetylo-ACP, czemu towarzyszy uwolnienie wolnego ACP i CO2, katalizowana przez enzym kondensujący
2. REDUKCJA acetoacetylo-ACP do D-3-hydroksybutyrylo-ACP przy użyciu NADPH katalizowana przez beta-ketoacylo-ACP
3. ODWODNIENIE D-3-hydroksybutyrylo-ACP do krotonylo-ACP, katalizowane przez dehydratazę 3-hydroksyacylo-ACP
4. REDUKCJA krotonylo-ACP przez kolejną cząsteczkę NADPH do butyrylo-ACP, katalizowana przez reduktazę enoilo-ACP
Bilans energetyczny utleniania kwasów tłuszczowych.
W każdym cyklu, kiedy acylo-CoA jest skracany o dwa atomy węgla, powstaje po jednej cząsteczce,
odpowiednio FADH2,NADH i acetylo-CoA. Rozkład palmitylo-CoA przebiega w siedmiu cyklach. W procesie odszczepienia acetylo-CoA zachodzą dwie reakcje utlenienia, w których biorą udział FAD i NAD ( powstają w sprzężeniu z tlenową fosforyzacją odpowiednio 2 i 3 cząsteczki ATP) Acetylo-CoA spalając się w cyklu Krebsa dostarcza 12 cząsteczek, zaś straty na aktywację kwasu tłuszczowego wynoszą 1 cząsteczkę. 1 reszta acetylo-CoA=12ATP, NADH=3ATP, FADH2=2ATP.ZYSK 17ATP
Biosynteza triacyloglicroli.
Triacyloglicerole są syntetyzowane z cząsteczek acylo-CoA i 3-fosfoglicerolu powstającego z fosfodihydroksyacetonu, będącego związkiem przejściowym glikolizy. W wyniku połączenia cząsteczek acylo-CoA z 3-fosfoglicerolem powstaje (w wyniku działania acetylotransferazy 3-fosfoglicerolowej) najpierw kwas lizofosfatydowy, który następnie reaguje z kolejną cząsteczką acylo-CoA, co prowadzi do wytworzenia kwasu fosfatydowego. Usunięcie grupy fosforanowej z kwasu fosfatydowego powoduje utworzenie diacyloglicerolu (DAG), ulegającego dalszej acylacji do triacyloglicerolu. Siłę napędową reakcji stanowi hydroliza wysokoenergetycznych wiązań tioestrowych łączących część acylową z CoA.
Regulacja metabolizmu triacylogliceroli.
Na etapie, w którym bierze udział lipaza kontrolowana przez cAMP.
*) glukagon, adrenalina, noradrenalina
- wiążą się z receptorem na powierzchni komórki tłuszczowej aktywacja cyklazy adrenylowej wzrost stopnia cAMP (w kwasach tłuszczowych)
- cAMP aktywator kinazy białkowej, która fosforyluje lipazę aktywacja lipazy ulonienie WKT do krwi
*) insulina
- działa odwrotnie
- obniża poziom cAMP defosforylacja zahamowanie lipazy
- stymuluje tworzenie triacylogliceroli
Porównanie syntezy i degradacji kwasów tłuszczowych.
- synteza zachodzi w cytozolu komórek prokariotycznych i eukariotycznych, zaś rozpad w mitochondriach komórek eukariotycznych
- w syntezie związkiem redukującym jest NADPH, zaś podczas rozpadu KT wytwarzany jest NADH
- podczas syntezy KT są kowalencyjnie związane z białkowym nośnikiem grup acylowych (ACP), zaś podczas rozpadu wiążą się z CoA
- poszczególne aktywności enzymatyczne przeprowadzające syntezę KT występują w pojedynczym łańcuchu polipeptydowym, którego dimer nazywa się syntezą kwasów tłuszczowych, a w cyklu beta-oksydacji poszczególne aktywności enzymatyczne są związane z odrębnymi enzymami
Biosynteza cholesterolu.
SCHEMAT
Opisz proces tworzenia ciał ketonowych (Ketogeneza).
Nadmiar acetylo-CoA powstającego w wyniku beta-oksydacji kwasów tłuszczowych jest przekształcany w acetooctan i D-3-hydroksymaślan, które wraz z acetonem są nazywane ciałami ketonowymi. Powstawanie acetooctanu z acetylo-CoA obejmuje trzy etapy. Dwie cząsteczki acetylo-CoA łączą się tworząc acetoacetylo-CoA w reakcji katalizowanej przez 3-ketotiolazę. Następnie acetoacetylo-CoA reaguje z kolejną cząsteczką acetylo-CoA i z wodą, tworząc 3-hydroksy-3-metyloglutarylo-CoA, który jest rozszczepiany na acetylo-CoA i acetooctan. W matriks mitochondrialnej acetooctan w reakcji katalizowanej przez dehydrogenazę 3-hydroksymaślanu, ulega redukcji do 3-hydroksymaślanu.
Ketogeneza w tkankach pozawątrobowych.
- ciało ketonowe to alternatywny produkt utleniania WKT w wątrobie
- u zdrowego człowieka stężenie ciał ketonowych nie przekracza 0,2 mmol/l
- mózg, serce, nerki wykorzystują ciała ketonowe jako materiał energetyczny 9ale głownie wykorzystują glukozę)
- najczęstsza produkcja ciał ketonowych przy cukrzycy (niedobór insuliny), może to doprowadzić do ketonemii i kwasicy ketonowej, a nawet śpiączki cukrzycowej
- nadmiar ciał ketonowych we krwi spowodowany jest zwiększeniem ich produkcji
Znaczenia związków ketonowych w organizmie.
- materiał energetyczny w tkankach pozawątrobowych (utleniane prze glukozę i KT)
- ich utlenianie może być czynnikiem oszczędzającym glukozę dla tkanek zależnych od glukozy
Wykazać schematycznie możliwości powstawania cukrów z tłuszczów.
*Formy wydalania azotu u zwierząt.
w formie amoniaku zwierzęta amonioteliczne: larwy płazów, ryby kostnoszkieletowe, krokodyle, żółwie słodkowodne
w formie mocznika zwierzęta ureoteliczne: ryby chrzęstnoszkieletowe, dorosłe płazy, ssaki, lądowe skąposzczety
w formie kwasu moczowego zwierzęta urikoteliczne: gady (oprócz krokodyla), ptaki, pajęczaki, owady
Synteza aminokwasów z tłuszczów.
acetylo-CoA+ 7 malonylo-CoA+ 14 NADPH+ 14H+ palmitylo-CoA+ HCO3- + 7 CoA-SH+ 14NADP+
Rola L-karnityny w transporcie kwasów tłuszczowych.
Aby cząsteczki acylo-CoA o łańcuchach dłuższych niż 10 atomów węgla mogły przejść przez we. błonę mitochondrialną muszą ulec sprzężeniu z polarną cząsteczką karnityny. Reakcja sprzęgania jest katalizowana przez enzym umieszczony po zew. stronie błony -acylotransferazę karnitynową I i polega na usunięciu CoA i zastąpieniu go cząsteczką karnityny. Następnie translokaza karnityna/acylokarnityna transportuje acylokarnitynę przez wew. błonę mitochondrialną. Translokaza będąc przenośnikiem antyportowym przenosi cząsteczki acylokarnityny do matrix mitochondrium, gdzie cząsteczki karnityny są uwalniane, czemu towarzyszy przeniesienie grupy acylowej z powrotem na CoA. Reakcję tę katalizuje acylotransferaza karnitynowa II znajdująca się po wew. stronie błony od strony matrix.
Ketoza
- stan, w którym dochodzi do nadmiernego wytwarzania ciał ketonowych w organizmie
- ciała ketonowe obecne w surowicy i moczu chorego kwasica
- kwasica ketonowa wywołana przez: zatrucia alkoholowe, głodzenie, śpiączkę wątrobową i mózgową
- związana ze znaczną hiperglikemią
- brak insuliny
Ujemny bilans energii - przyczyny.
- wartość energii wydalanej z organizmu> energia dostarczana do organizmu
- taki bilans grozi osłabieniem i zmniejszeniem masy ciała (korzystny u osób otyłych)
Ogólny SCHEMAT degradacji aminokwasów.
Procesy dekarboksylacji i dezaminacji i transaminacji.
DEKARBOKSYLACJA
Polega na rozerwaniu wiązania między gr. karboksylową -COOH i resztą cząsteczki aminokwasu, w wyniku czego wydziela się CO2 i powstaje odpowiednia amina. Reakcję katalizują dekarboksylazy aminokwasowe. Dekarboksylacja aminokwasowi jest źródłem amin biogennych- substancji o dużej aktywności fizjologicznej, np. histamina (po dekarboksylacji histydyny)
DEZAMINACJA
Zachodzi dzięki obecności gr. aminowej. U kręgowców reakcje dezaminacji zachodzą poprzez transminację i oksydację, która prowadzi do powstania ketokwasów. W procesie transminacji akceptorem amoniaku z aminokwasu jest ketokwas, który jest też donorem tlenu na rzecz dezaminowanego aminokwasu. Tak alanina, kw. asparaginowy i kw. glutaminowy mogą poswatać z kw. pirogronowego, kw. szczawiowooctowego i kw. 2-oksoglutarowego. Podczas dezaminacji oksydacyjnej najpierw zachodzi odwodorowanie aminokwasu przy udziale enzymu dehydrogenazy do aminokwasu, potem dołączana jest woda i odłączany amoniak.
TRANSAMINACJA
Reakcja transaminowania, reakcja przeniesienia gr. aminowej (np. z aminokwasu) na cząsteczkę ketokwasu (np. kw. pirogronowego), w wyniku której powstaje nowy ketokwas i nowy aminokwas. W ukł. biologicznych reakcja ta katalizowana jest przez aminotransferazy. Umożliwia ona w ukł. biologicznych biosyntezę różnych aminokwasów z np. kw. glutaminowego.
Dekarboksylacja (aminokwasów zasadowych, kwaśnych, obojętnych) przebieg enzymatyczny procesu.
Produktami dekarboksylacji aminokwasów są aminy I-rzędowe. Reakcję te katalizują enzymy klasy liaz, dekarboksylazy aminokwsowe, które współdziałają z fosforanem pirydoksalu. PLP łączy się z aminokwasem, tworząc przejściowo zasadę Schaffa. Od tego połączenia zostaje odszczepiony CO2, a po kilku dalszych przekształceniach zostaje odtworzony PLP oraz uwolniona amina.
Na skutek dekarboksylacji aminokwasów obojętnych powstają monoaminy I-rzędowe.
Dekarboksylacja aminokwasów zasadowych prowadzi do wytworzenia tiamin I-rzędowych.
Produktami dekarboksylacji aminokwasów kwaśnych są obojętne β- lub γ-aminokwasy (np. z kw. asparaginowego powstaje β-alanina).
Istnieją także specyficzne dekarboksylazy aminokwasów kwaśnych, działające na końcowe gr. karboksylowe. Ich produktami są odpowiednie α-aminokwasy obojętne.
Przebieg i znaczenie deaminacji.
Deaminacja
- polega na eliminacji z cząsteczek zw. chemicznego grupy aminowej
- dwa etapy: dehydratacja i właściwa deaminacja- odłączenie gr. NH2
- w środowisku naturalnym jest to pierwszy proces degradacji aminokwasów
- umożliwia wykorzystanie aminokwasu jako substratu
Losy grupy aminowej po dezaminacji.
NH2 zostają przekształcone do amoniaku lub mocznika i wydalone z ustroju wraz z potem i moczem.
Los aminokwasowych szkieletów węglowych w czasie degradacji aminokwasów.
Mogą być przekształcane i wprowadzone do przemian cyklu Krebsa jako jego następne metabolity
* acetylo-CoA
*kw. szczawiooctanowy
*kw. 2-oksyglutanowy
*kw. bursztynowy
*kw. fumarowy
Mogą posłużyć do syntezy cukrów i tłuszczów, dlatego wyróżniamy aminowkwasy:
* cukrotworcze
*tłuszczotwórcze
Przebieg transaminacji.
- przeniesienie gr. aminowej z aminokwasu na część ketokwasu
- powstaje nowy ketokwas i nowy aminokwas
- katalizator: aminotransferaza
- umożliwia biosyntezę aminokwasów np. kw glutaminowego
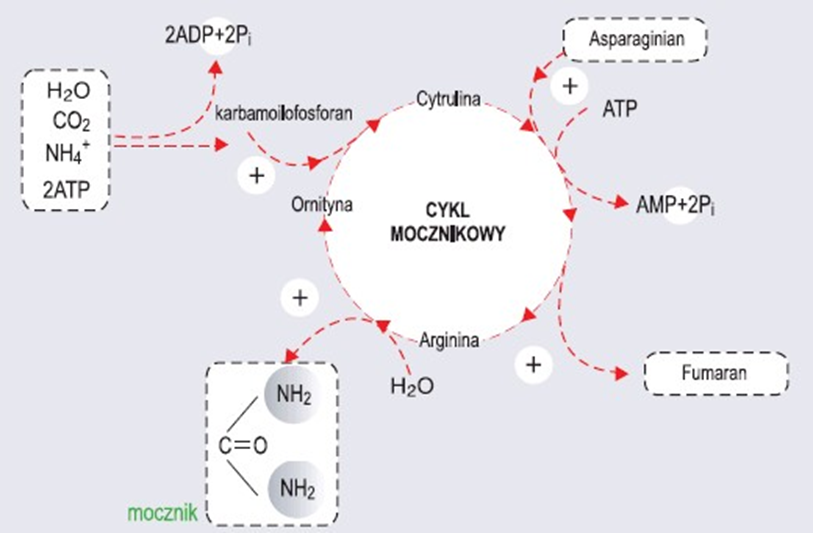
Cykl mocznikowy-przebieg, funkcja biologiczna.
Mocznik jest syntetyzowany w wątrobie w cyklu mocznikowym. Następnie jest wydzielany do krwioobiegu, filtrowany przez nerki i wydalany z moczem. Oto sumaryczna reakcja cyklu mocznikowego: NH4+ + HCO3- + H2O + 3ATP + asparaginian mocznik + 2ATP + AMP + 2Pi + PPi + fumaran
Trawienie białek i absorpcja aminokwasów w przewodzie pokarmowym.
Polega na stopniowym ich rozkładzie aż do uzyskania wolnych aminokwasów, które są zw. dobrze rozpuszczalnymi i łatwo wchłanianymi. Trawienie rozpoczyna się w żołądku. Gruczoły znajdujące się w ścianach żołądka, których wydzielina tworzy sok żołądkowy, produkują enzymy powodujące rozkład cząsteczek białka na mniejsze i już dobrze rozpuszczalne cząsteczki. Są to pepsyna i kalepsyna działające w silnie kwaśnym środowisku wytworzonym przez kw. solny. Następny etap trawienia białek odbywa się w dwunastnicy pod wpływem enzymów znajdujących się w soku trzustkowym: trypsyny i chymotrypsyny. Strawność białka zależy od zawartości celulozy, chemiceluliz, lignin i sub. Hamujących działanie proteaz trawiennych, jak np. inhibitor trypsyny i chymotrypsyny. Ujemny wpływ na wykorzystanie białek zapasowych ma też niekorzystny ich skład aminokwasowi- mała zawartość aminokwasów niezbędnych.
Bilans azotowy.
Różnica pomiędzy dzienną ilością azotu przyjętego wraz z białkiem (N-absorbowany) a iloscią azotu wydalanego. (N-wydalany)- gł. Mocznik, jony, kreatyniny, kw moczowy
*) osoby zdrowe Nabs-Nwyd= 0 - równowaga azotowa
*) u dzieci, młodzieży, podczas ciąży Nabs-Nwyd > 0 - dodtani bilans azotowy
*) u osób niedożywionych, na diecie wysokobiałkowej, w ciężkiej chorobie
Nabs-Nwyd < 0 - ujemny bilans azotowy
Regulacja cyklu mocznikowego.
1.Enzym regulatorowy - syntetaza karbamoilofosforanowa
2.Allosteryczny aktywator - N-acetyloglutaminian
3.Zmiany w pożywieniu - podczas głodzenia zwiększa się stężenie enzymów cyklu mocznikowego
Regulacja hormonalna przemian białek i aminokwasów.
Niemal wszystkie hormony wpływają na przemiany białek i aminokwasów
- istnieje różnica w działaniu hormonów na przemiany białek i aminokwasów
- hormony anaboliczne (hormon wzrostu, insulina, androgeny), budowa białek w mięśniach, rozpad białek w trzewiach (wątroba i przewód pokarmowy)
- hormony kataboliczne (glikokortykosteroidy, glukagon), rozpad Białek w mięśniach, powodują rozpad białek i syntezę węglowodanów z aminokwasów, budowa białek w trzewiach
Połączenie cyklu mocznikowego z cyklem Krebsa.

Aminy biogenne -postawanie rola.
- związki powstałe w wyniku dekarboksylacji aminokwasów obojętnych i zasadowych
- zwężają naczynia krwionośne i podwyższają ciśnienie krwi (histamina działa odwrotnie)
- pełnia role hormonów tkankowych, stabilizatorów rybosomów, przenośników w przenoszeniu impulsów nerwowych przez zakończenia nerwowe
Aminokwasy gluko i ketogenne.
Glukogenne: dają jako końcowy produkt depolaryzaji bursztynylo-CoA, pirogronian, alfa-ketoglutaran, furman, szczawiooctan
Ala, Arg, Gly, Gys, His, Pro-OH, Asp, Glu, Ser, Thr, Val
Ketogenne rozkładajace się do acetylo-CoA lub acetylooctanu Lys, Lev
Gluko-ketogenne: daja produkty rozkładu mogące służyć jako substraty glukoneogenezy i syntezy kw. tłuszczowych Phe, Lys, Tyr
Witaminy rozpuszczalne w wodzie i ich rola w procesach metabolicznych.
Tiamina (B1)
- niezbędna w metabolizmie węglowodanów
- niezbędna w prawidłowym funkcjonowaniu ukł. Nerwowego
- wspomaga proces gojenia się ran, uśmierza ból
Ryboflawina (B2)
- udział w końcowych etapach niektórych reakcji metabolicznych
- ważna rola w utrzymaniu zdrowej skóry, błon śluzowych, osłonek nerwów, rogówki
- ważna do prawidłowego funkcjonowania narządu wzroku
Niacyna (B3)
- odpowiada za prawidłowy metabolizm pokarmów
- pomaga w utrzymaniu zdrowej skóry
- wpływa na funkcjonowanie ukł. nerwowego i pokarmowego
Pirydoksyna (B6)
- niezbędna w metabolizmie aminokwasów, kwasów tłuszczowych oraz glukozy
- bierze udział w wytwarzaniu erytrocytów
- odp. Za prawidłowe ciśnienie tętnicze
Cyjanokobalamina (B12)
- działa we wszystkich komórkach organizmu przede wszystkim w jelitach, szpiku, ukł nerwowym
- odpowiada za wytwarzanie zdrowych komórek krwi
- utrzymuje prawidłową jakość osłonek nerwowych
- bierze udział w syntezie kw nukleinowych
Kwas foliowy (B11)
- z wit. B12 w syntezie kw nukleinowych
- w produkcji krwinek czerwonych
- prawidłowy rozwój płodu
- zapobiega uszkodzeniom ukł nerwowego
- reguluje podział komórek dziedzicznych
Kwas pantolenowy i biotyna
- prawidłowe funkcjonowanie tarczycy, skóry i włosów
- ważna rola w wielu procesach metabolicznych
Kwas askorbinowy (C)
- tworzenie kolagenu i utrzymywanie go w optymalnym stanie
- silny przeciwutleniacz (chroni kom przed wolnymi rodnikami)
- ułatwia przyswajanie żelaza
- udział w metabolizmie pokarmów
Wyszukiwarka
Podobne podstrony:
pytania na egzamin z Biochemi jaki miala FIZJO
baza pytań na egzamin z biochemii 2010 wersja I (1)
egzamin biochemia
Egzamin z biochemii pytania id 153179
Egzamin Biochemia 13 opracowanie Ewy B
Egzamin biochemia
egzamin biochemia 2012, BIOCHEMIA D, egzamin biochemia
EGZAMIN BIOCHEMIA
pytania egzamin biochemia, STUDIA, WSR - Fizjoterpia, Rok I, Semestr 1, Semestr I, Biochemia, Wykład
Egzamin biochemia1 05 12 odpowiedzi
Egzamin z biochemii 4, Farmacja
Egzamin z biochemii, Farmacja
Egzamin z biochemii-, far, II rok III sem, biochemia, egzamin
Egzamin z biochem, Medycyna, Biochemia (HogwartZabrze)
Egzamin luty 09, biochemia I, zadania z egzaminu z biochemii
Biochemia 3, EGZAMIN BIOCHEMIA 06-2003 II, (1) Jakie funkcje MOŻNA przypisać albuminie krwi:
Wersja A, (Sylwia) studia semestr 3, Biochemia, EGZAMIN, EGZAMIN, biochemia
więcej podobnych podstron