Wykład
Ogólna budowa komórki
Wszystkie żywe organizmy są zbudowane z komórek - małych zbiorniczków z różnymi związkami chemicznymi.
Najprostszy organizm (na przykład bakteria albo pierwotniak) to pojedyncza, samodzielna komórka. Taka komórka pobiera wybrane cząsteczki chemiczne z otoczenia, produkuje z nich energię albo inne związki chemiczne, dostosowuje swoje zachowanie do warunków środowiska, czasem potrafi pływać... Co jakiś czas dla rozrywki (i zdobycia nowego źródła pokarmu) może zaatakować inny organizm, na przykład mnie. W końcu się dzieli i powstają z niej dwie komórki - dwa nowe organizmy, które wyglądają i zachowują się bardzo podobnie. Życie jednokomórkowców jest chyba dość monotonne.
Większe organizmy składają się z większej liczby komórek, które mają różne poziomy specjalizacji, kształty i zadania. W organizmie wielokomórkowym - na przykład u człowieka - komórki łączą się tworząc tkanki (zespoły komórkowe wyspecjalizowane w podobnych funkcjach) i narządy (zespoły kilku połączonych ze sobą tkanek, które spełniają jakieś zadanie w organizmie). Komórki należące do jednego organizmu porozumiewają się ze sobą, wzajemnie kontrolują swoje zachowanie i przekazują sobie najróżniejsze informacje.
Jak jest zbudowana pojedyncza komórka?
Najogólniej komórki dzieli się na prokariotyczne (pozbawione typowego jądra komórkowego) i eukariotyczne, zawierające jądro komórkowe. Człowiek (tak jak inne wielokomórkowe organizmy wyższe) składa się z kilkuset miliardów komórek eukariotycznych.
Wnętrze komórki jest oddzielone od środowiska zewnętrznego przez błonę komórkową, zbudowaną z białek i substancji tłuszczowych. Niektóre białka umieszczone w tej błonie transportują cząsteczki chemiczne do wnętrza albo na zewnątrz komórki.
Komórka jest wypełniona przez cytoplazmę - półpłynną masę, w której są zawieszone różne części komórki, czyli organella.
W środku komórki eukariotycznej znajduje się jądro komórkowe. Otoczka jądrowa (zbudowana z dwóch błon białkowo-lipidowych) oddziela wnętrze jądra od cytoplazmy. Jądro komórkowe zawiera materiał genetyczny, czyli cząsteczki DNA, w których znajdują się geny kontrolujące życie komórki.
W cytoplazmie pływają mitochondria - organella otoczone podwójną błoną białkowo-lipidową. Mitochondria wytwarzają większość energii, której komórka potrzebuje do swoich procesów życiowych. Mitochondria przypominają bakterie - mają własne cząsteczki DNA, wytwarzają własne białka, rozmnażają się przez podział; ich kształt i rozmiar jest podobny do komórek bakteryjnych. Prawdopodobnie kiedyś były bakteriami, które na pewnym etapie ewolucji 'zgodziły się' współpracować z komórkami eukariotycznymi. Teraz ich współpraca z resztą komórki jest tak bliska, że mitochondria nie mogłyby żyć samodzielnie (między innymi dlatego, że do prawidłowego działania mitochondriów są potrzebne geny jądra komórkowego).
Cytoszkielet jest wewnętrznym, elastycznym rusztowaniem komórki, które utrzymuje organella na swoich miejscach, nadaje komórce kształt i potrafi się bardzo szybko przebudowywać w odpowiedzi na sygnały otrzymywane przez komórkę.
W cytoplazmie jest dużo rybosomów - maleńkich organelli wytwarzających nowe białka. Rybosomy mogą swobodnie pływać w cytoplazmie albo przyczepiać się do układu cystern i kanalików określanego jako szorstka siateczka śródplazmatyczna ('szorstka' od doczepionych do niej rybosomów). Większość białek powstających na wolnych rybosomach jest przeznaczona na wewnętrzne potrzeby komórki, a białka produkowane na rybosomach siateczki są eksportowane, czyli wydzielane poza komórkę. Eksportowane białka przechodzą przez aparat Golgiego - stos płaskich cystern i pęcherzyków - a potem pęcherzyki niosą je w stronę błony komórkowej i wylewają na zewnątrz komórki.
Oprócz szorstkiej siateczki śródplazmatycznej w komórce jest też gładka siateczka śródplazmatyczna, do której rybosomy się nie przyczepiają. Zadaniem gładkiej siateczki jest wytwarzanie lipidów i niszczenie cząsteczek trujących dla komórki.
Inne pęcherzyki pływające w cytoplazmie to lizosomy, które zawierają enzymy trawiące różne związki chemiczne, i peroksysomy - organella związane z produkcja i rozkładem nadtlenku wodoru.
W komórkach roślinnych są jeszcze chloroplasty (otoczone podwójną błoną organella, które przeprowadzają fotosyntezę, czyli reakcje robienia czegoś z niczego, a konkretnie - złożonych cząsteczek chemicznych z dwutlenku węgla i wody pod wpływem energii światła słonecznego); ściana komórkowa, która otacza komórki roślinne z zewnątrz i nadaje im sztywność; oraz wakuole (wodniczki) - duże, otoczone pojedyncza błoną pęcherzyki, do których komórka wydziela różne substancje.
Nawet z tego pobieżnego wyliczenia widać, że błony białkowo-lipidowe mają dla komórki szczególne znaczenie. Oddzielają od siebie poszczególne przedziały komórki, co pozwala jej przeprowadzać w tym samym czasie przeciwstawne reakcje biochemiczne (na przykład jedna część komórki wytwarza jakieś cząsteczki, a kilka mikrometrów dalej identyczne cząsteczki są spalane w mitochondriach). Komórka potrafi przesyłać cząsteczki z jednego przedziału (na przykład z cytoplazmy) do drugiego (na przykład do szorstkiej siateczki śródplazmatycznej). Podział komórki na różne, otoczone błonami przedziały (inaczej: kompartmenty) to jedno z bardzo użytecznych rozwiązań, które pozwoliły komórkom eukariotycznym stworzyć organizmy wielokomórkowe z prawdziwego zdarzenia.
W komórkach prokariotycznych nie ma tak rozbudowanego labiryntu błon białkowo-lipidowych: cała bakteria to jeden kompartment. Nie ma w niej mitochondriów, siateczek śródplazmatycznych, chloroplastów, lizosomów ani peroksysomów, a cząsteczka DNA będąca materiałem genetycznym (tzw. genofor) nie jest otoczona żadną błoną - zajmuje miejsce zwane nukleoidem, które nie jest oddzielone żadną granicą od innych części komórki.
Błony biologiczne.
Błony biologiczne (inaczej: błony białkowo-lipidowe) są niezwykle ważną częścią każdej komórki .
Bez szczelnej błony komórkowej, czyli tej błony biologicznej, która oddziela wnętrze komórki od środowiska zewnętrznego, komórka w ogóle nie mogłaby istnieć. Nie tylko dlatego, że jej zawartość rozlałaby się na wszystkie strony - zadania błony komórkowej są dużo bardziej wyrafinowane i nie ograniczają się do mechanicznego utrzymywania wszystkich ważnych cząsteczek w jednym miejscu Wszechświata. Warunki, w których żyją komórki, często są bardzo zdradliwe. Temperatura, pH, dostępność rożnych pierwiastków i związków chemicznych... to wszystko może zmienić się dosłownie w każdej chwili. Ale skomplikowane procesy życiowe, które zachodzą w komórkach, mogą się odbywać tylko pod warunkiem bardzo precyzyjnego zachowania warunków panujących wewnątrz komórki. Zmiana pH o pół jednostki może spowodować, że enzymy odczytujące informacje zakodowane w genach się zbuntują, produkcja energii się nie uda, a z kopiowania cząsteczek DNA też nic nie wyjdzie. Na szczęście błona komórkowa zapewnia względną stałość warunków panujących w komórce, bo jest selektywnie przepuszczalna - mogą przez nią przechodzić tylko niektóre cząsteczki chemiczne.
Inne błony biologiczne tworzą rożne organella komórkowe: szorstką i gładką siateczkę śródplazmatyczną, aparat Golgiego, mitochondria, otoczkę jądra komórkowego, lizosomy, peroksysomy, chloroplasty... Te wszystkie błony również są selektywnie (wybiórczo) przepuszczalne. Dzięki temu poszczególne części komórki mogą mięć rożny skład chemiczny i sprawnie przeprowadzać przeciwstawne reakcje chemiczne.
Błony biologiczne komórki pełnia rożne funkcje (na przykład błona oddzielająca mitochondrium od cytoplazmy zachowuje się inaczej niż błona komórkowa), ale struktura tych wszystkich błon jest zaskakująco podobna. I to nie tylko w obrębie jednej komórki! Praktycznie wszystkie błony biologiczne żywych komórek mają zbliżoną budowę. Mówiąc ogólnie, składają się z dwóch rodzajów cząsteczek chemicznych: lipidów (tłuszczy) i białek.
Głównymi substancjami błon biologicznych są fosfolipidy - specjalne lipidy z przyłączonymi grupami fosforanowymi. W cząsteczce każdego fosfolipidu można wyróżnić hydrofilową główkę i hydrofobowe ogonki - a może nóżki? Spójrzcie na rysunek. (Cząsteczki hydrofobowe to takie, które odpychają cząsteczki wody i wola kontaktować się z innymi cząsteczkami hydrofobowymi; cząsteczki hydrofilowe zachowują się odwrotnie - z rozkoszą otaczają się molekułami wody).
Teraz trochę chemii: główka jest zbudowana z reszty glicerolu, grupy fosforanowej i np. choliny (wtedy fosfolipid nosi nazwę fosfatydylocholiny). Nóżki to kwasy tłuszczowe. Na pewno zauważyliście, że prawa nóżka jest w jednym miejscu załamana - ma coś w rodzaju kolana. Prawa nóżka jest nienasyconym kwasem tłuszczowym, to znaczy - zawiera podwójne wiązania w łańcuchu węglowym. Kolanko znajduje się w miejscu podwójnego wiązania. Im więcej takich podwójnych wiązań w kwasie tłuszczowym, tym nóżka staje się bardziej giętka i elastyczna - sprawnie rozdaje kopniaki innym cząsteczkom fosfolipidów.
Skoro główka fosfolipidu jest hydrofilowa, a nóżki - hydrofilowe, to cała cząsteczka ma schizofreniczny stosunek do wody: główka chętnie się nią otacza, a nóżki przed nią uciekają. Chemicy określają takie cząsteczki mianem 'amfipatyczne' (częściowo hydrofobowe, częściowo hydrofilowe).
Dzięki takim właściwościom fosfolipidów powstały błony biologiczne (jedna wstawiona w odpowiednie miejsce grupa fosforanowa zadecydowała o tym, że Ziemia nie jest martwą planetą). Kiedy cząsteczki fosfolipidów wrzuci się do wody i porządnie pomiesza, to po chwili na powierzchni wody cząsteczki fosfolipidów same stworzą najprostsza błonę - DWUWARSTWĘ LIPIDOWĄ:
Obie warstwy fosfolipidów są odwrócone do siebie nóżkami. W ten sposób hydrofobowe nóżki, które stykają się tylko ze sobą, unikają kontaktu z cząsteczkami wody, a hydrofilowe główki bez przeszkód mogą robić to, co najbardziej lubią, czyli hm... otaczać się cząsteczkami wody. Wszystkie fragmenty fosfolipidów są wtedy usatysfakcjonowane pod względem energetycznym i dlatego dwuwarstwa lipidowa jest trwała i sama się odtwarza, jeśli jakiś złośliwiec wsadzi łapę do naczynia, w którym przeprowadzamy nasze wiekopomne doświadczenie, i zburzy delikatną strukturę dwuwarstwy.
Taka dwuwarstwa lipidowa jest delikatna i płynna - przypomina cienką warstewkę oleju rozlana na powierzchni wody. James Watson nazwał dwuwarstwę lipidową 'dwuwymiarowym płynem'... Płynność dwuwarstwy zależy od jej składu chemicznego: oprócz fosfolipidów są w niej też inne tłuszcze, na przykład (w komórkach zwierzęcych) cząsteczki cholesterolu, które przyczepiają się do nóżek fosfolipidów i usztywniają błony. W komórkach dwuwarstwa lipidowa jest asymetryczna, to znaczy, że jej górna warstwa rożni się składem chemicznym od dolnej. Kiedy komórka popełnia samobójstwo na drodze apoptozy, można ją rozpoznać miedzy innymi po tym, że lipidy z cytoplazmatycznej warstwy przeskakują na powierzchnie komórki...
Błony biologicznie są selektywnie przepuszczalne, bo prawie wszystkie duże, obdarzone ładunkiem elektrycznym cząsteczki chemiczne odbijają się od dwuwarstwy lipidowej jak piłeczki pingpongowe. Dotyczy to między innymi białek i kwasów nukleinowych. Te, które są w komórce, nie mogą swobodnie z niej zwiać, a te, które pukają do komórki z zewnątrz, muszą się strasznie namęczyć, żeby się do niej przedostać. Tylko cząsteczki rozpuszczalne w tłuszczach mogą bez trudu przenikać przez dwuwarstwę lipidową, która nie stanowi dla nich najmniejszej przeszkody (takie cząsteczki po prostu jej 'nie widza'). W ten sposób do komórki wchodzą np. cząsteczki hormonów steroidowych (np. hormonów płciowych) i hormony tarczycy. No i cząsteczki alkoholu...
Druga grupa cząsteczek tworzących błony biologiczne to białka. Białka błonowe mogą być na stale wbudowane w błonę i przechodzić na wylot przez dwuwarstwę lipidowa; mogą też przyczepiać się do wewnętrznej albo zewnętrznej powierzchni błony i nie trzymać się zbyt mocno dwuwarstwy lipidowej. To właśnie rożne rodzaje białek błonowych w największym stopniu decydują o tym, jaką funkcję w komórce może pełnić dana błona biologiczna. Inne białka są w błonie komórkowej, inne w błonie mitochondrialnej, jeszcze inne w błonie lizosomów...
W błonie przez cały czas coś się dzieje. Tak jak wiele innych części komórki, błony biologiczne mają bardzo dynamiczną strukturę i ciągle się przebudowują.
Co mogą robić białka błonowe? Bardzo wiele. Niektóre z nich są pompami wyrzucającymi z komórki niepotrzebne cząsteczki czy jony albo wychwytującymi cenne związki chemiczne z płynu otaczającego komórkę. Do tego procesu potrzebna jest energia wytwarzana w mitochondriach. Oto druga tajemnica selektywnej przepuszczalności błony biologicznej: komórka nie przepuszcza przez nią niechcianych cząsteczek, które odbijają się od elastycznej dwuwarstwy lipidowej, a jednocześnie zmusza inne cząsteczki do przejścia przez błonę, wychwytując je z roztworów i bezlitośnie przepychając na druga stronę błony.
Inne białka błony komórkowej są np. receptorami dla hormonów białkowych, które (w przeciwieństwie do hormonów lipidowych) nie mogą swobodnie przechodzić na drugą stronę błony. Taki receptor po związaniu hormonu uruchamia kaskadę innych białek, które przekazują sygnał do odpowiedniego miejsca komórki. Zresztą receptory błonowe wiążą się także z innymi cząsteczkami, nie tylko z hormonami. Prawie wszystkie metody wykorzystywane przez komórki do rozmowy ze sobą mają coś wspólnego z białkami błonowymi. Innymi białkami błonowymi komórka trzyma się włókien substancji międzykomórkowej... Do białek błonowych od wewnątrz przyczepia się cytoszkielet; białka błony nadają komórkom charakterystyczny kształt (np. formując erytrocyty w dwuwklęsłe biszkopty)... Niektóre białka przytwierdzone do błony są enzymami przeprowadzającymi jakieś reakcje chemiczne (wykorzystując do tych reakcji np. niektóre lipidy błony - komórki w ten sposób przekazują sobie pewne rodzaje sygnałów)... Limfocyty przy pomocy białek błonowych rozpoznają uszkodzone albo zakażone komórki, które trzeba zniszczyć... Chyba nie da się jednym tchem wymienić wszystkich procesów związanych z białkami błon biologicznych.
Krótkie podsumowanie... Wszystkie błony biologiczne są zbudowane z podwójnej warstwy cząsteczek fosfolipidów i innych pochodnych tłuszczów oraz z białek błonowych, które kontrolują rożne procesy życiowe komórki. Taka częściowo płynna błona przez cały czas się zmienia, przebudowuje i reaguje na zmieniające się warunki środowiska, a to umożliwia prawidłowe działanie i komórki jako całości, i jej poszczególnych organelli.
Jądro komórkowe.
Trudno jest wybierać ważniejsze i mniej ważne organella komórki - w końcu wszystkie są potrzebne, żeby to skomplikowane laboratorium biochemiczne, jakim jest komórka, działało zgodnie z planem. Ale jednak jądro komórkowe ma dość szczególna rolę. W końcu właśnie w nim mieści się zakodowana informacja genetyczna, która steruje całym życiem komórki. Współczesna biologia molekularna, która przede wszystkim zajmuje się działaniem genów, jest szczególnie zainteresowana jądrem komórkowym.
Oglądając nie dzielącą się komórkę pod mikroskopem optycznym nie dowiemy się za dużo o budowie jądra komórkowego. W najlepszym wypadku, gdy preparat jest odpowiednio zabarwiony, zobaczymy w środku komórki kółko wypełnione mozaiką ciemnych ziarnistości. To kółko (które w niektórych komórkach może mieć eliptyczny albo zupełnie nieregularny kształt) to właśnie przekrojone jądro komórkowe.
Budowę jądra można dużo dokładniej poznać przy pomocy mikroskopu elektronowego.
Zawieszone w cytoplazmie jądro komórkowe jest od niej odgraniczone przez otoczkę jądrowa, która jest zbudowana z dwóch błon białkowo-lipidowych. Błony tworzące otoczkę jądrową są - tak jak inne błony biologiczne - selektywnie przepuszczalne, więc skład chemiczny kariolimfy (płynu wypełniającego wnętrze jądra komórkowego) różni się od składu chemicznego cytoplazmy.
Dwie błony białkowo-lipidowe otoczki jądrowej w niektórych miejscach łącza się ze sobą. W tych punktach znajdują się pory jądrowe - bramki, przez które przechodzą cząsteczki transportowane z jądra do cytoplazmy i w przeciwnym kierunku. Pory jądrowe są zbudowane z białek i mają ciekawy kształt przestrzenny: wyglądają jak dwa połączone szprychami koncentryczne pierścienie. Przez wewnętrzny pierścień przepływa większość cząsteczek podróżujących z albo do jądra komórkowego. Oczywiście w tym wewnętrznym pierścieniu siedzą specjalne białka - 'bramkarze', którzy nie pozwalają niektórym cząsteczkom przekraczać granicy otoczki jądrowej, a inne cząsteczki przepychają na druga stronę błony.
Nitkowate zagęszczenia tworzące delikatna siateczkę w jądrze komórkowym to chromatyna. Chromatyna jest zbudowana z kwasu deoksyrybonukleinowego (DNA), w którym zakodowane są geny, oraz z różnych białek, które przyczepiają się do DNA i pozwalają zmieścić długie cząsteczki tego kwasu w jądrze komórkowym oraz uczestniczą w odczytywaniu informacji genetycznych. W chromatynie można też znaleźć trochę cząsteczek kwasu rybonukleinowego (RNA).
Chromatyna jądra nie dzielącej się komórki może być bardziej albo mniej skondensowana. Postać bardziej rozluźniona, w której nukleosomy nie leżą tak blisko siebie, nosi nazwę euchromatyny; geny zlokalizowane we fragmentach DNA znajdujących się w euchromatynie często są aktywne - białka odczytujące informacje genetyczna mają do nich łatwiejszy dostęp. W mikroskopie euchromatyna jest widoczna jako delikatne włókienka.
Heterochromatyna to bardziej zbity rodzaj chromatyny. W obrębie heterochromatyny najczęściej leża nieaktywne geny albo cząsteczki DNA, które nic nie kodują. Na preparatach mikroskopowych heterochromatynę widać jako ciemne grudki często zbierające się na obrzeżach jądra, tuż pod otoczką jądrowa.
Wewnątrz jądra komórkowego są jąderka - owalne, nie otoczone żadną dodatkową błoną miejsca produkcji rybosomalnego RNA (rRNA), kwasu nukleinowego, który wchodzi w skład rybosomów (organelli, które biorą udział w biosyntezie białek). Łatwo zgadnąć, że komórki produkujące dużo białek (na przykład komórki trzustki wytwarzające enzymy trawienne) często mają większe i wyraźniejsze jąderka.
Wszystkie elementy jądra komórkowego utrzymują się we właściwych miejscach, bo są przyczepione do macierzy jądrowej - skomplikowanej sieci białkowej, która tworzy wewnętrzny szkielet jądra komórki. Szczególnie ciekawymi białkami macierzy jądrowej są laminy - cząsteczki wyściełające wewnętrzną powierzchnie otoczki jądrowej. Laminy odpadają od otoczki jądrowej podczas podziału komórki (wtedy są fosforylowane przez specjalne enzymy, a otoczka jądrowa się rozpada, żeby segregacja chromosomów mogła przebiegać prawidłowo), a podczas apoptozy są trawione przez kaspazy - enzymy, które biorą udział w komórkowym samobójstwie.
Czy mogą istnieć komórki bez jądra komórkowego? Tak. Takimi komórkami są erytrocyty, czyli czerwone ciałka krwi. Dojrzewające erytrocyty wypluwają z siebie jądra komórkowe i gotowe czerwone ciałko krwi jest po brzegi napchane cząsteczkami hemoglobiny. Może bez reszty poświęcić się przenoszeniu tlenu we krwi i robi to bardzo dobrze, chociaż nie ma już swoich własnych genów. Ale nie potrafi robić już nic innego. Nie może uruchomić nowych genów i reagować na zmieniające się warunki środowiska. Staje się takim ogłupiałym, komórkowym zombie, który razem z genami traci zdolność produkcji wielu rożnych białek zakodowanych w DNA jądra komórkowego.
Mitochondrium.
Komórka rozkłada skomplikowane związki chemiczne na drobne cząsteczki, z innych małych cząsteczek robi większe i bardziej złożone, transportuje rożne substancje przez swoje błony biologiczne. Do tych wszystkich procesów życiowych potrzebna jest energia.
Skąd komórka bierze energię?
Z takich reakcji rozkładu złożonych cząsteczek, które mają charakter egzoenergiczny, czyli wiążą się z uwalnianiem energii. Komórka nie pozwala, żeby cała energia pochodząca z takich reakcji rozpraszała się w postaci ciepła; specjalne enzymy magazynują duża cześć tej energii w wysokoenergetycznych wiązaniach rożnych cząsteczek chemicznych. Taka energia chemiczna może być stopniowo uwalniana z wiązań wysokoenergetycznych wtedy, gdy komórka jej potrzebuje, np. do przeprowadzenia jakiejś endoergicznej reakcji chemicznej, która nie mogłaby zajść bez impulsu energii doprowadzonej z zewnątrz.
Główna cząsteczka magazynująca energię jest adenozynotrójfosforan (ATP).
Cząsteczka ATP składa się z adeniny, rybozy i trzech grup fosforanowych.
Jak wyglądają mitochondria?
Są trochę podobne do bakterii pływających w cytoplazmie komórek eukariotycznych (dlaczego? - pisałem o tym w pierwszym odcinku naszego kursu). Są dość ruchliwe; kręcą się po komórce (często przyczepiają się do mikrotubuli cytoszkieletu i ślizgają się po nich jak małpy po lianach), a czasem nawet łączą się ze sobą, ale w niektórych komórkach twardo tkwią w jednym miejscu. Kształt mitochondriów bywa rożny i zmienia się między innymi w zależności od zawartości tlenu w komórce; najczęściej mitochondria wyglądają jak owalne pałeczki, ale mogą też wyglądać jak kulki albo długie wstęgi.
Mitochondrium jest zbudowane z dwóch błon białkowo-lipidowych. Zewnętrzna błona mitochondrialna nadaje kształt mitochondrium i dość łatwo przepuszcza rożne cząsteczki pływające w cytoplazmie. Za to wewnętrzna błona jest bardzo szczelna. Charakterystyczne uwypuklenia wewnętrznej błony są określane jako grzebienie mitochondrialne (cristae). Wąska szparka położona pomiędzy wewnętrzną i zewnętrzną błoną mitochondrium to przestrzeń międzybłonowa.
Wnętrze mitochondrium (to, co znajduje się pod wewnętrzną błoną) nosi nazwę macierzy (matriks) mitochondrialnej. W macierzy jest pełno enzymów przeprowadzających reakcje biochemiczne. Oprócz tego pływają w niej koliste cząsteczki mitochondrialnego DNA - mtDNA (mitochondrium ma własne geny - to taka pamiątka po pięknych czasach, kiedy mitochondria były jeszcze wolnymi jak ptaki bakteriami). Każda cząsteczka mtDNA zawiera pełny genom mitochondrialny, ale występuje w macierzy w kilku kopiach. Poza tym w macierzy można też znaleźć rybosomy mitochondrialne, na których powstają cząsteczki białek kodowanych przez geny mitochondrialne (te rybosomy są mniejsze niż rybosomy pływające w cytoplazmie i przypominają raczej rybosomy bakteryjne. Znajdują się tam również cząsteczki tRNA przenoszące aminokwasy do mitochondrialnych rybosomów.
Jakie reakcje biochemiczne przebiegają w mitochondriach?
W macierzy mitochondrialnej:
- cykl kwasu cytrynowego (cykl kwasów trójkarboksylowych, cykl Krebsa) - utlenianie dwuwęglowych fragmentów rożnych związków chemicznych (miedzy innymi glukozy) do dwutlenku węgla z jednoczesnym wytworzeniem energii chemicznej; elektrony i protony uwalniane w tym cyklu są przekazywane enzymom łańcucha oddechowego, które mieszczą się w wewnętrznej błonie mitochondrialnej;
- beta-oksydacja kwasów tłuszczowych - rozkład cząsteczek kwasów tłuszczowych na dwuwęglowe fragmenty, które następnie wchodzą do cyklu kwasu cytrynowego; uwalniane przy okazji elektrony i protony przechodzą do łańcucha oddechowego;
- cykl mocznikowy (głównie w mitochondriach wątroby) - wytwarzanie mocznika z grup aminowych uwolnionych z rozkładanych aminokwasów;
- transkrypcja genów mitochondrialnych i produkcja białek mitochondrialnych.
Cykl kwasu cytrynowego i beta-oksydacja są bezpośrednio związane z produkcją energii; cykl mocznikowy pozwala wyeliminować z organizmu szkodliwe produkty przemiany materii, a transkrypcja i translacja genów mtDNA pozwalają wytwarzać nowe białka mitochondrialne i w ten sposób kontrolować reakcje przeprowadzane przez mitochondrium. Nie będę teraz dokładnie omawiał tych reakcji... wrócimy do nich w którymś z kolejnych odcinków naszego kursu.
W wewnętrznej błonie mitochondrialnej siedzą enzymy łańcucha oddechowego. Łańcuch oddechowy to szeregowo ustawiona grupa enzymów, przez które przechodzą kationy wodorowe i elektrony. Energia związana z przechodzeniem elektronów przez enzymy łańcucha oddechowego jest wykorzystywana do pompowania protonów do przestrzeni międzybłonowej. W przestrzeni międzybłonowej gromadzi się wiele protonów, dlatego pH panujące w tej przestrzeni jest niskie i tworzy się gradient (różnica stężeń) protonów, zwany gradientem chemiosmotycznym. Wypompowane protony wracają do macierzy mitochondrialnej przez kanały cząsteczek enzymu zwanego syntazą ATP. Kiedy protony przechodzą przez syntazę ATP, uwalnia się energia wykorzystywana do produkcji nowych wiązań wysokoenergetycznych w cząsteczkach ATP. Jednocześnie elektrony przechodzą na atomy tlenu, które przygotowują się na przyjęcie wracających do macierzy protonów. W ten sposób tworzą się cząsteczki wody.
Duża cześć energii powstającej w procesie biologicznego utleniania ulega rozproszeniu w postaci ciepła, ale i tak komórki są bardziej sprawne niż maszyny stworzone przez człowieka. Ocenia się, że ok. 40% energii powstającej ze spalania złożonych cząsteczek ulega zmagazynowaniu w wysokoenergetycznych wiązaniach ATP.
Cytoszkielet.
Cytoszkielet to sieć białkowych włókien i rurek, która tworzy rusztowanie komórek eukariotycznych. Szkielet komórki wcale nie jest sztywny, tylko elastyczny - potrafi się szybko przebudowywać zgodnie z potrzebami komórki.
Włókna i rurki cytoszkieletu dzielimy na: filamenty i mikrotubule.
Cytoszkielet bierze udział:
- w nadawaniu kształtu komórkom i utrzymywaniu tego kształtu;
- w utrzymywaniu prawidłowej struktury tkanek (jeśli naciśnie się na skórę, to komórki naskórka nie rozlatują się na wszystkie strony miedzy innymi dlatego, że cytoszkielet je usztywnia i łączy ze sobą);
- w poruszaniu się komórek (np. w pełzaniu komórek układu odpornościowego i w fagocytozie, czyli pożeraniu takich 'ciał obcych', jak bakterie i wirusy);
- w skurczu mięsni;
- w podziałach komórek (niektóre leki przeciwnowotworowe, np. taksol, niszczą szybko dzielące się komórki nowotworowe uniemożliwiając prawidłowe działanie cytoszkieletu);
- w transporcie pęcherzyków i organelli przez cytoplazmę;
- w przekazywaniu informacji miedzy komórkami;
- w utrzymywaniu polarności komórek (czyli wyznaczaniu, gdzie są poszczególne strony komórki - góra, dol., lewo, prawo...);
- w machaniu rzęskami i wiciami;
- w apoptozie - genetycznie programowanym samobójstwie komórek.
Szorstka siateczka śródplazmatyczna i transport białek.
Szorstka siateczka śródplazmatyczna (rough endoplasmic reticulum, RER) to układ poplątanych kanalików i spłaszczonych woreczków, obecny w cytoplazmie komórek eukariotycznych. Światło tych wszystkich kanalików łączy się ze sobą, więc białka podróżujące przez szorstka siateczkę mogą bez problemów trafić do wszystkich zakamarków RER. Ale białko musi najpierw dostać się z cytoplazmy do wnętrza siateczki.
Transport nowych cząsteczek białek do wnętrza siateczki śródplazmatycznej zaczyna się już na początku translacji, kiedy białko nie jest jeszcze gotowe. Wysuwający się z rybosomu koniec takiego białka zawiera specjalna etykietkę - sekwencje sygnałową, która kieruje cząsteczkę białka do RER. Ta sekwencja sygnałowa jest rozpoznawana przez specjalne białka, które pływają w cytoplazmie. Po przyłączeniu tych białek do rybosomu translacja jest zatrzymywana, a cały rybosom podpływa do siateczki i przyczepia się do niej (szorstka siateczka śródplazmatyczna nosi nazwę 'szorstkiej' właśnie z powodu rybosomów, które chętnie się do niej przyczepiają i na niektórych zdjęciach z mikroskopu elektronowego wyglądają jak kropki przyłączone do powierzchni RER).
Kiedy rybosom przyczepi się do RER, translacja może być kontynuowana. Wydłużający się łańcuch polipeptydowy od razu wpada do kanału błonowego, który tworzy się w błonie siateczki śródplazmatycznej. Kiedy translacja się skończy, całe białko wpada do wnętrza RER, kanał błonowy się zamyka, a rybosom odpada od siateczki i rusza do cytoplazmy w poszukiwaniu nowej cząsteczki mRNA. (Sekwencja sygnałowa jest najczęściej odcinana ze świeżego białka przez enzymy obecne w siateczce).
Niektóre białka są przeznaczone do wbudowania w błonę RER. Takie białka nie przechodzą do końca przez błonę siateczki śródplazmatycznej i pozostają przyczepione do tej błony. Naukowcy znaleźli w tych białkach drugi rodzaj sekwencji sygnałowych, który oznacza, że białko powinno zostać zatrzymane w błonie siateczki.
A co dzieje się z białkami, które wpadną do wnętrza szorstkiej siateczki śródplazmatycznej? Natychmiast rzuca się na nie cała banda różnych enzymów. Niektóre z nich to białka opiekuńcze (chaperones), które pomagają młodym cząsteczkom białek przyjąć i utrzymać właściwy kształt przestrzenny. Inne enzymy katalizują tworzenie wiązań dwusiarczkowych pomiędzy resztami cysteiny nowej cząsteczki białka i w ten sposób decydują o tym, jak powinno wyglądać nowe białko.
Potem niektóre cząsteczki białek są poddawane jeszcze innym modyfikacjom. Wreszcie nowe białka trafiają do pęcherzyków transportowych, które niosą cząsteczki białek do aparatu Golgiego (tam do wielu białek przyłączają się reszty cukrowe, czyli - mówiąc językiem naukowym - dochodzi do glikozylacji białek). Inne pęcherzyki odrywają się od aparatu Golgiego i płyną w stronę błony komórkowej; po połączeniu się z błona wylewają swoja zawartość na zewnątrz komórki i w taki sposób cząsteczki białek są wydzielane poza cytoplazmę. Jeszcze inne pęcherzyki aparatu Golgiego wrzucają swoje białka do lizosomów.
Peroksysomy.
Wyglądają jak wielokształtne pęcherzyki o średnicy od 0.1 do około 1 mikrometra. Ściana każdego pęcherzyka jest zbudowana z pojedynczej błony białkowo-lipidowej. Pomiędzy peroksysomami istnieją dość liczne połączenia; dlatego czasem mówi się nawet o siateczce peroksysomalnej. Wnętrze tej niby-siateczki jest po brzegi wypchane różnymi enzymami. W niektórych peroksysomach stężenie enzymów jest tak wysokie, że cząsteczki białek enzymatycznych łączą się ze sobą i tworzą duże kompleksy, które na zdjęciach z mikroskopu elektronowego są widoczne jako ciemne 'kupki'.
W organizmie człowieka najwięcej peroksysomów można znaleźć w komórkach wątroby i kanalików nerkowych.
Peroksysomy uczestniczą miedzy innymi w niszczeniu substancji szkodliwych dla organizmu, czyli detoksykacji trujących związków chemicznych. Jedna z metod detoksykacji wykorzystuje cząsteczki nadtlenku wodoru (H2O2) wytwarzane w peroksysomach. Specjalny enzym - katalaza - odłącza atomy wodoru od cząsteczek unieszkodliwianych substancji i przyłącza je do nadtlenku wodoru, produkując wodę:
XH2 + H2O2 -------------> utleniony X + 2H2O
[katalaza]
W ten sposób komórki naszego organizmu mogą rozkładać m.in. trujące cząsteczki kwasu mrówkowego, aldehydu mrówkowego i - jakżeby inaczej! - etanolu. Katalaza może również przekształcać cząsteczki nadtlenku wodoru w wodę i tlen (nadmiar nadtlenku wodoru też jest szkodliwy dla komórek):
2H2O2 ------------> 2H2O + O2
[katalaza]
Nadtlenek wodoru to po angielsku 'hydrogen peroxide', stąd wzięła się nazwa 'peroksysomy'.
W ogóle enzymy peroksysomów przeprowadzają dużo reakcji metabolicznych wykorzystujących tlen (podobno peroksysomy zużywają aż 20% tlenu wytwarzanego przez komórkę). Co jeszcze ciekawego dzieje się w peroksysomach? Zachodzi w nich na przykład szczególny rodzaj beta-oksydacji kwasów tłuszczowych o długich łańcuchach. Peroksysomalna beta-oksydacja różni się od beta-oksydacji zachodzącej w mitochondriach głownie tym, że w jej pierwszym etapie uwalniana energia rozprasza się w postaci ciepła (jednocześnie powstaje nadtlenek wodoru). Taka odmiana beta-oksydacji wydaje się mniej korzystna energetycznie (protony i elektrony uwalniane w pierwszym etapie szlaku reakcji nie trafiają do łańcucha oddechowego), ale być może w ten sposób komórka trochę się 'podgrzewa'. Po kilku rundkach beta-oksydacji cząsteczka kwasu tłuszczowego ulega skróceniu i jest transportowana do mitochondrium, gdzie ulega 'normalnej' beta-oksydacji.
Peroksysomy biorą też udział:
- w produkcji kwasów żółciowych;
- w wytwarzaniu cholesterolu;
- w rozkładzie cząsteczek puryn;
- w rozkładzie niektórych aminokwasów.
W komórkach roślin peroksysomy pełnią trochę inne funkcje; uczestniczą m.in. w fotooddychaniu albo w metabolizmie cząsteczek tłuszczów do sacharydów w dojrzewających nasionach.
Niektóre leki oraz substancje toksyczne powodują 'mnożenie się' peroksysomów w komórkach - w ten sposób komórki próbują bardziej wydajnie usuwać trujące cząsteczki. Podejrzewa się, że nadmierne zwiększenie liczby peroksysomów może ułatwiać przemianę prawidłowej komórki w komórkę rakową, bo w peroksysomach powstaje dużo chemicznie aktywnych pochodnych tlenu, a takie wolne rodniki tlenowe mogą łatwo uszkadzać DNA i prowadzić do mutacji w genach komórki.
Jak powstają peroksysomy? Prawdopodobnie przez pączkowanie z innych peroksysomów. Znajdujące się w nich enzymy są wytwarzane na wolnych rybosomach cytoplazmatycznych. Potem są importowane do wnętrza peroksysomów, bo zawierają specjalne sekwencje sygnałowe, które pozwalają im trafić do właściwych organelli.
Cykl komórkowy.
Narodziny, życie, rozmnażanie, starość i w końcu śmierć - to etapy, przez które przechodzi każda żywa istota. W przypadku komórek sytuacja jest trochę bardziej skomplikowana: życie komórek można podzielić na wiele powtarzających się okresów. Każdy taki okres to jeden cykl komórkowy (inaczej: cykl życiowy komórki).
Pojedynczy cykl komórkowy można podzielić na kilka głównych faz, które reguluje tykający zegar biologiczny:
- faza G1 - wysoka aktywność metaboliczna, wzrost komórki - komórka wykonuje zadania, do których została powołana, i przygotowuje się do skopiowania swojego materiału genetycznego, czyli replikacji DNA;
- faza S (synteza) - replikacja DNA;
- faza G2 - końcowe przygotowania do podziału;
- faza M - mitoza - podział, w którym powstają dwie komórki potomne o tej samej liczbie chromosomów.
Obie komórki powstające podczas mitozy wchodzą w fazę G1 i cykl komórkowy zaczyna się od nowa.
W cyklu życiowym komórki wyróżnia się jeszcze fazę G0. Komórka też musi odpocząć... Faza G0 jest właśnie taką fazą spoczynkową, do której komórka 'wychodzi' najczęściej z fazy G1 albo G2. Komórki znajdujące się w fazie G0 też pełnią funkcje, do których zostały powołane przez organizm, ale przestają się dzielić (takimi komórkami są np. dojrzałe neurony). Niektóre komórki mogą wracać z fazy G0 do cyklu komórkowego i dalej się dzielić, jeśli zostaną odpowiednio pobudzone na przykład przez hormony albo czynniki wzrostowe.
Jednak komórka nie może niezależnie podejmować decyzji o podziałach. Stwarzałoby to niebezpieczeństwo nadmiernego wzrostu albo powstania komórki nowotworowej. Dlatego w podejmowaniu decyzji o mitozie pomagają jej 'sąsiadki', czyli położone obok niej inne komórki, oraz zależność od zakotwiczenia komórki do podłoża (większość komórek swobodnie zawieszonych w roztworze nie chce się dzielić, w przeciwieństwie do komórek rakowych, które trąca kontrolę nad cyklem komórkowym oraz zależność od zakotwiczenia i mogą bez trudu dzielić się pływając w roztworze). 'Sąsiadki' wydzielają specjalne białka - czynniki wzrostowe, które łączą się ze swoimi receptorami w błonie komórkowej tej komórki, dla której przeznaczony jest sygnał, i w ten sposób uruchamiają kaskadę sygnałową. Taka kaskada może polegać np. na przyłączaniu grup fosforanowych do kolejnych cząsteczek białek, które po uaktywnieniu wędrują w dół, aż do jądra komórkowego, gdzie uaktywniane są czynniki transkrypcyjne. Aktywne czynniki transkrypcyjne pobudzają wybrane geny, które pozwalają komórce przejść do kolejnych faz cyklu komórkowego.
Przechodzenie komórki przez poszczególne fazy cyklu jest napędzane przez ewolucyjnie konserwatywny mechanizm regulujący. Ten mechanizm składa się z dwóch głównych rodzajów białek - cyklin i kinaz cyklinozależnych.
Cykliny to białka, których poziom w komórce zmienia się w różnych fazach cyklu komórkowego. W miarę przechodzenia komórki przez fazy G1, G2 i S gwałtownie wzrasta poziom cyklin typu D, A, E i w końcu B. Te cykliny łączą się ze swoimi kinazami cyklinozależnymi (kinazy to enzymy, które przyłączają grupy fosforanowe do innych cząsteczek, czyli przeprowadzają fosforylację tych cząsteczek; słowo 'cyklinozależne' wskazuje na to, że aktywność tych kinaz jest regulowana przez cykliny).
A skąd komórka wie, kiedy się zestarzeć i przestać się dzielić?
Niektórzy twierdzą, że duże znaczenie mają segmenty DNA zwane telomerami. Telomery znajdują się na końcach chromosomów i niczym 'kapturki' chronią chromosomy przed sklejaniem podczas podziałów i innymi uszkodzeniami. Jednak w fazie S, czyli podczas replikacji DNA, telomery nieznacznie się skracają... aż po (najczęściej) kilkudziesięciu podziałach osiągają wartość progową. To jest dla komórki sygnałem, że należy wejść w etap starzenia się, a potem śmierci.
Niektóre komórki wytwarzają enzym zwany telomerazą, który potrafi wydłużać telomery i 'oszukiwać' ten zegar biologiczny. Takie komórki mogą dzielić się dłużej niż inne. Telomerazę produkują m.in. komórki zarodkowe, komórki macierzyste szpiku kostnego, komórki jelit i inne komórki, które szybko się dzielą. Wiele komórek nowotworowych też wytwarza telomerazę; uważa się, że między innymi z tego powodu komórki raka mogą dzielić się tak szybko i w niekontrolowany sposób. Jednak rola telomerazy w powstawaniu nowotworów nie jest jeszcze do końca wyjaśniona: być może sama obecność telomerazy w komórce utrudnia mutacje (bo telomeraza przyczepia się do telomerow i zabezpiecza chromosomy przed uszkodzeniem), a długość telomerów nie ma tak dużego znaczenia. Istnieją też inne teorie tłumaczące starzenie się komórek. Niektóre z nich uwzględniają duże znaczenie uszkodzeń DNA (zwłaszcza mitochondrialnego) przez wolne rodniki.
Niektóre komórki, na przykład te, w których doszło do poważnych uszkodzeń DNA, opuszczają cykl komórkowy i umierają na drodze programowanej śmierci, czyli apoptozy.
APOPTOZA
Apoptoza to zaprogramowana śmierć komórki - dzięki temu mechanizmowi usuwane są zużyte lub uszkodzone komórki. Można ją przyrównać do zaplanowanego samobójstwa komórki w organizmie wielokomórkowym mające na względzie dobro całego organizmu. W odróżnieniu od martwicy, gdzie dochodzi do uszkodzenia jakimś zewnętrznym czynnikiem, apoptoza jest zjawiskiem naturalnym w rozwoju i życiu organizmów. Termin apoptoza wprowadzono w 1972 roku i z greckiego oznacza nie śmierć komórki ale opadanie liści i kwiatów.
Apoptoza, w odróżnieniu od nekrozy, polega na kurczeniu się komórki poprzez utratę wody. Szybkie zmiany w jądrze komórkowym mają charakter zorganizowany - chromatyna jądrowa ulega kondensacji, a DNA zostaje pocięte przez endonukleazy. Następuje dezintegracja cytoszkieletu. W procesie apoptozy organelle komórkowe pozostają jednak nienaruszone. Są one usuwane z komórki wraz z fragmentami chromatyny w tzw. ciałkach apoptycznych, pęcherzykach powstałych w wyniku zmian w strukturze błony komórkowej. W większości przypadków są one następnie fagocytowane przez komórki żerne. Wyjątkiem są np. ciałka apoptyczne soczewki oka, które zawierają zamiast cytoplazmy z organellami białko krystalinę. Apoptoza nie wywołuje stanu zapalnego.
Czynnikami powodującymi apoptozę mogą być:
oddziaływania międzykomórkowe - na skutek przekazywania błędnych informacji o podziałach komórkowych.
Odzwierciedleniem rosnącego zainteresowania badaniami nad apoptozą było przyznanie Nagrody Nobla z fizjologii lub medycyny w roku 2002. Otrzymali ją Sydney Brenner, H. Robert Horvitz i John E. Sulston za ich odkrycia z dziedziny genetycznej regulacji organogenezy i zaprogramowanej śmierci komórki.
DNA wszystkich komórek posiada informację niezbędną do przeprowadzenia procesu apoptozy czyli tzw. samodestrukcji komórki.
Takie akty samozniszczenia są czymś całkowicie normalnym. Dochodzi do nich w czasie rozwoju zarodkowego np. podczas zanikania błony pławnej między palcami człowieka, czy też podczas eliminacji limfocytów T rozpoznających własne antygeny. Apoptoza ma też miejsce w dorosłych organizmach umożliwiając np. utrzymanie stałej liczby komórek w poszczególnych narządach. W ten sposób mogą też być usuwane potencjalnie niebezpiecznie komórki jak np. te zakażone wirusami, posiadające uszkodzone DNA lub nawet komórki nowotworowe.
Podczas apoptozy następuje kurczenie się komórek, zagęszczenie chromatyny a w efekcie do fragmentacji komórki - powstają tzw. ciałka apoptyczne zawierające fragmenty jądra komórkowego. Takie pofragmentowane elementy komórki zostają wchłonięte przez komórki sąsiednie na drodze fagocytozy.
W procesie apoptozy udział biorą enzymy z rodziny ICE (interleukin -1-beta-converting enzyme) nazywane też kaspazami. Występują one w komórce jako proenzymy, które aktywują wzajemnie same siebie oraz inne enzymy, a to z kolei prowadzi do uszkodzenia białek cytoszkieletu, enzymów jądrowych oraz do cięcia DNA między poszczególnymi nukleosomami. Aktywacja kaspaz zależna jest z kolei od innych białek z rodziny Bcl-2. W skład tych ostatnich wchodzą zarówno enzymy aktywujące jak i hamujące działanie kaspaz, a tym samym regulujące występowanie apoptozy. I to właśnie równowaga między tymi grupami białek decyduje o wystąpieniu apoptozy.
Podczas apoptozy zachodzą różne, nie do końca poznane zmiany w metabolizmie komórki i ekspresji niektórych jej genów. Materiał genetyczny komórki ulega zniszczeniu: specjalne enzymy tną DNA jądra komórkowego na kawałki o długości kilkuset par nukleotydów. Zmienia się struktura błony komórkowej i cytoszkieletu. Komórka ulega obkurczeniu, bo z cytoplazmy uciekają cząsteczki wody. Błona komórkowa tworzy ciałka apoptotyczne, czyli uwypuklenia, do których przechodzą różne organella komórkowe. Ciałka apoptotyczne są fagocytowane i trawione przez komórki otaczające komórkę popełniającą samobójstwo. 
{kind=link}
Apoptoza często występuje w komórkach rosnącego zarodka człowieka i innych zwierząt, bo niektóre komórki zarodka muszą umrzeć, żeby embrion mógł się prawidłowo rozwijać. Pewne dojrzałe komórki, na przykład komórki wyścielające jelita albo komórki naskórka, również umierają w wyniku apoptozy, kiedy ich śmierć jest potrzebna dla prawidłowego funkcjonowania organizmu.
Komórki wchodzą na drogę apoptozy, kiedy nie potrafią naprawić uszkodzeń swojego DNA, spowodowanych działaniem różnych czynników mutagennych. W ten sposób organizm broni się przed powstaniem nowotworu, bo komórka zawierająca pewne mutacje DNA może zmienić się w komórkę nowotworową.
Apoptoza chroni przed rakiem
Apoptoza - nazywana inaczej programowaną lub bardziej popularnie "samobójczą" śmiercią komórki, to bardzo ważny proces autodestrukcji, regulowany genetycznie. Umożliwia on organizmom wyższym kontrolowanie liczby komórek w trakcie dojrzewania, a także pozwala usunąć komórki uszkodzone lub zmienione nowotworowo.
Badania nad apoptozą dają nadzieję na rozwiązanie wielu problemów medycznych. Zaburzenia w procesie apoptozy zwiększa bowiem ryzyko chorób nowotworowych.
Wywoływanie apoptozy to jeden z pożądanych efektów terapii przeciwnowotworowej. W procesie apoptozy komórek ważną rolę odgrywają pewne enzymy - nukleazy. Są to enzymy trawiące kwasy nukleinowe (DNA trawiony jest przez DNazy, a RNA - przez RNazy).
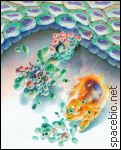
Apoptoza komórki
Na początku komórka kurczy się oddzielając się od sąsiadnich. Następnie na powierzchni pojawiają się pęcherzyki ("komórka wrze"), chromatyna (czarna na obrazku) - kompleks jądrowego DNA i białek - kondensuje się na krawędzi jądra. Dalej jądro i cała komórka fragmentują się, a powstałe "szczątki komórki" są szybko wchłaniane przez sąsiednie komórki.
Wyszukiwarka