Genomy
To podstawowy komplet informacji genetycznej, niezbędny do funkcjonowania organizmu. Może być haploidalny
( komórki prokariotyczne - np.bakterie, sinice oraz nieliczne eukariotyczne -np. wiciowce, sporowce, eugleniny, pokolenie gametofitu roślin, trutnie) oraz diploidalny u zdecydowanej większości Eucaryota. W tym wypadku instrukcja genetyczna w komórce występuje w dwóch kopiach.
Genom Procaryota = genofor + plazmid(y)
Genom Eucaryota = DNA jądrowe + mt.DNA (mitochondrialne), a w komórkach roślinnych + chl.DNA (chloroplastów).
Tab. 1 Wielkości genomów liczona w parach nukleotydów * jednoniciowy RNA, ** jednoniciowy DNA (Lewiński,
Genetyka str. 53 )
organizm |
Wielkość genomu |
|
organizm |
Wielkość genomu |
Wiroid PSTV |
359* |
|
Owady: muszka owocowa |
1, 4 ×108 |
Wirusy: Fag φX174 |
5500** |
|
Płazy: żaba szponiasta |
9,7 ×10 8 4,5 ×10 9 |
Fag λ |
4, 5× 10 4 |
|
Ssaki :mysz domowa |
3 ×10 9 |
Bakterie E. coli |
4, 2 × 106 |
|
człowiek |
3×10 9 |
Bacillus subtilis |
2,0× 106 |
|
Rośliny: wyka |
1 ×10 10 |
Grzyby: drożdże |
2,0 × 107 |
|
Groch zwyczajny |
1 ×10 9 - 9,9 ×10 9 |
Skorupiaki : krab |
1,4 × 109 |
|
trzykrotka |
3,0 ×10 10 |
Genomy wiroidów są za małe, aby zakodować jakiekolwiek białko, najmniejsze wirusy kodują 4 polipeptydy, a największe kilkaset, ale i tak nie są w stanie funkcjonować samodzielnie , jedynie jako bezwzględne pasożyty.
Model pojemności informacyjnej genomów
IN INFOR INFORMACJA INFORMACJA O RÓŻNYCH
KOMÓRKACH
wiroid wirus komórka prokariotyczna komórka eukariotyczna organizmu
Tab. 2 Liczba chromosomów u wybranych gatunków Tab. 3 Liczba genów u wybranych organizmów
Organizm |
Liczba chromosomów 2n |
||||
Muszka owocowa |
8 |
||||
Kukurydza |
20 |
||||
Salamandra |
24 |
||||
Człowiek |
46 |
||||
Nietoperz |
46 |
||||
Nosorożec |
84 |
||||
Jeden z gatunków kraba |
254 |
||||
Organizm |
Dł DNA |
Liczba genów |
Rok ustalenia |
||
Drożdże |
6,8 cm |
6 034 |
1996 |
||
Escherichia coli |
1,4 cm |
4 288 |
1997 |
||
Nicień |
|
19 099 |
1998 |
||
Rzodkiewnik |
|
25 000 |
|
||
Muszka owocowa |
58 cm |
13 061 |
|
||
Człowiek |
190 cm |
30 000 - 40 000 |
2001 |
||
mysz |
166 cm |
30 000 |
2002 |
||
1 gen = 1 łańcuch polipeptydowy Definicja zawodzi jedynie przy opisach genów nie ulegających translacji , tj. kodujących t RNA i rRNA oraz genów regulatorowych u Procaryota.
Odczytywanie informacji genetycznej czyli ekspresja informacji .
Gen to fragment cząsteczki DNA, zawierający informację wystarczającą do wytworzenia jednego rodzaju białka (taka definicja jest prawdziwa w przypadku większości genów, ale trzeba pamiętać, że niektóre geny nie kodują białek, tylko cząsteczki RNA, np. rRNA, tRNA albo zawierają introny lub geny regulacyjne, np. promotor ). Tylko odczytywane geny ulegają ekspresji, czyli wpływają na życie komórki (procesy metaboliczne). Białka kodowane przez geny mogą być:
- enzymami, które kontrolują procesy życiowe komórki;
mogą pełnić funkcje strukturalne, np. tworzyć elementy cytoszkieletu;
mogą budować organella komórkowe... itp., itd.
odpornościowe
regulacyjne (hormony)
motoryczne…………..
transportowe ………………
Ekspresja genów kodujących białka składa się z dwóch głównych etapów. Pierwszym jest TRANSKRYPCJA, czyli przepisanie odpowiedniego fragmentu DNA na mRNA zgodnie z regułą komplementarności nukleotydów (adenina w DNA pasuje do uracylu w RNA; tymina w DNA do adeniny w RNA; guanina w DNA do cytozyny w RNA i cytozyna w DNA do guaniny w RNA;
Drugi etap ekspresji genu to TRANSLACJA - wytworzenie cząsteczki białka na podstawie informacji przenoszonej przez cząsteczkę mRNA.
Jak wygląda transkrypcja?
Transkrypcja zachodzi w jądrze komórkowym (oczywiście tylko w tych komórkach, które mają klasyczne jądro komórkowe, czyli w komórkach eukariotycznych; w komórkach prokariotycznych transkrypcja przebiega w obrębie nukleoidu na terenie cytozolu). Odczytywanie genu zaczyna się od przyłączenia specjalnych białek (czynników transkrypcyjnych) do pierwszej części genu, zwanej promotorem. Czynniki transkrypcyjne pozwalają przyczepić do genu główny enzym transkrypcji (polimerazę RNA) i rozplatają kawałek podwójnej helisy DNA, umożliwiając polimerazie RNA rozpoczęcie transkrypcji. Polimeraza RNA przesuwa się wzdłuż jednego łańcucha podwójnej helisy (- nić sensowna, kodująca, matrycowa), wytwarzając cząsteczkę RNA, która jest komplementarna do tego łańcucha. (drugi łańcuch DNA najczęściej nic nie koduje). Po przejściu polimerazy RNA fragment DNA odzyskuje strukturę podwójnej helisy. W miarę przesuwania się polimerazy RNA po genie powstająca cząsteczka RNA staje się coraz dłuższa i wysuwa się z enzymu.
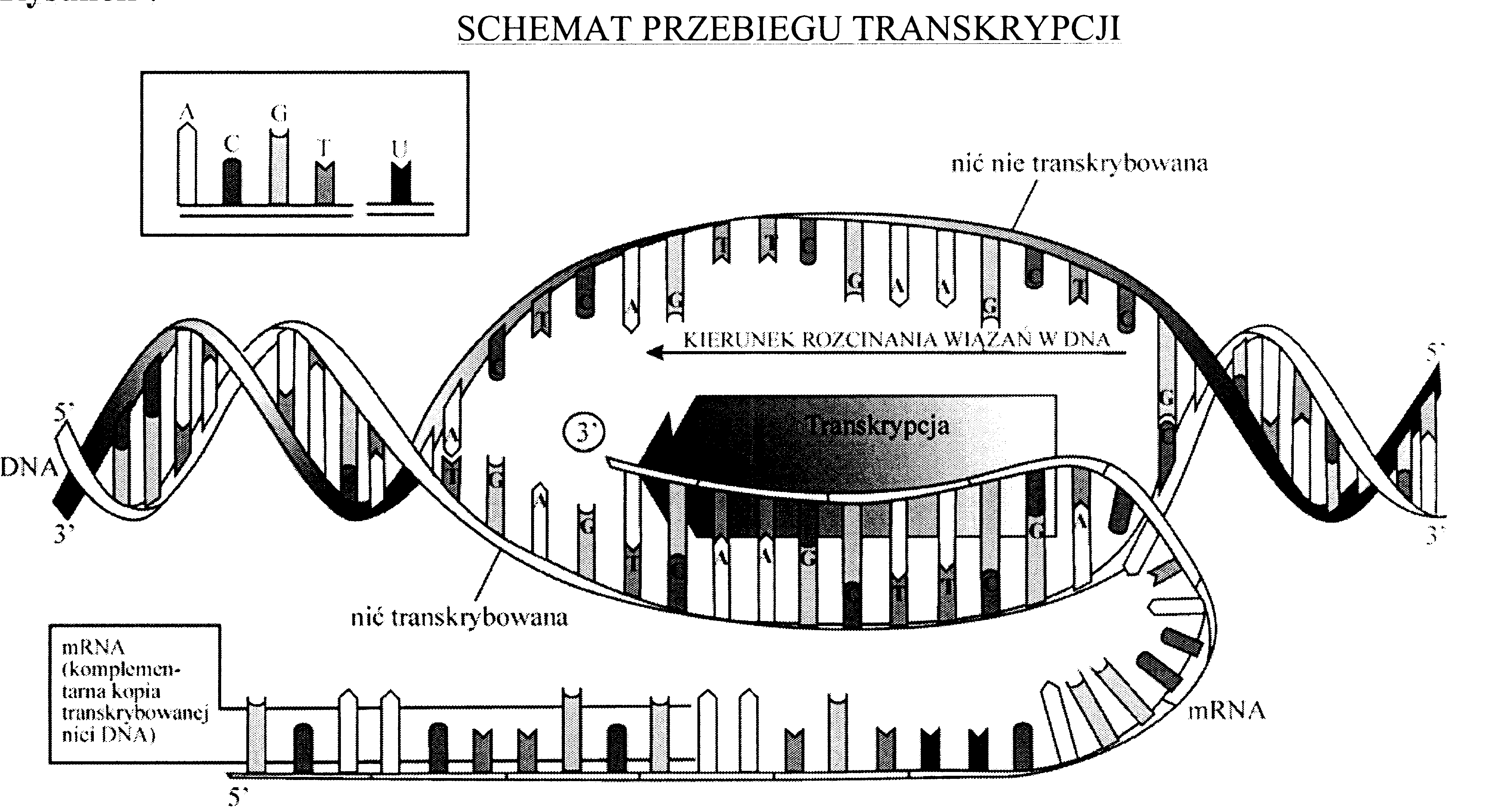
Jak enzymy przeprowadzające transkrypcję rozpoznają koniec genu?
W komórkach prokariotycznych (np. u bakterii) transkrypcja kończy się, gdy polimeraza RNA osiągnie odpowiednie miejsce na cząsteczce DNA. Na prośbę wielbicieli filmów akcji, naukowcy nazwali to miejsce terminatorem. Po osiągnięciu terminatora polimeraza RNA po prostu odpada od DNA i uwalnia świeżo zrobioną cząsteczkę RNA.
W komórkach eukariotycznych rosnąca cząsteczka RNA jest przecinana przez specjalne enzymy, które rozpoznają sekwencje nukleotydów AAUAAA albo AUUAAA. Pojawienie się tej sekwencji w RNA jest sygnałem do odcięcia RNA od polimerazy RNA, zakończenia transkrypcji i rozpoczęcia poliadenylacji (patrz dalej).
Regulacja ekspresji genu na etapie rozpoczynania transkrypcji jest stosunkowo najlepiej poznana przez naukowców (wiadomo, że inicjacja transkrypcji może się zmieniać w zależności od czynników transkrypcyjnych obecnych w komórce albo od rożnych modyfikacji tych czynników, np. fosforylacji).
U Prokaryota cząsteczka RNA wytworzona w procesie transkrypcji jest od razu wykorzystana do translacji.
W komórkach eukariotycznych jest to trochę bardziej skomplikowane: potrzebne jest jeszcze... ...składanie genu (splicing), a po polsku dojrzewanie.
Geny eukariotyczne najczęściej zawierają sekwencje kodujące (eksony - exons) i sekwencje, które nie kodują żadnego fragmentu białka (introny - introns), dlatego kopia jednej nici DNA nosi nazwę pre- mRNA (prekursor) lub hn-mRNA, (heterogenny). Polimeraza RNA nie odróżnia eksonów od intronów - przepisuje cały gen na cząsteczkę RNA. Podczas składania genu niepotrzebne introny są wycinane z dojrzewającej cząsteczki RNA ,a końce eksonów łączą się ze sobą. Gotowa cząsteczka mRNA zawiera tylko sekwencje kodujące.
W składaniu wielu genów uczestniczą kompleksy złożone z białek i małych cząsteczek sn-RNA. (wyjątkowy RNA o właściwościach enzymu, odkryty pod koniec XX w.)
Te kompleksy rybonukleoproteinowe noszą nazwę spliceosomów; (informosomy) można je zobaczyć na zdjęciach z mikroskopu elektronowego.
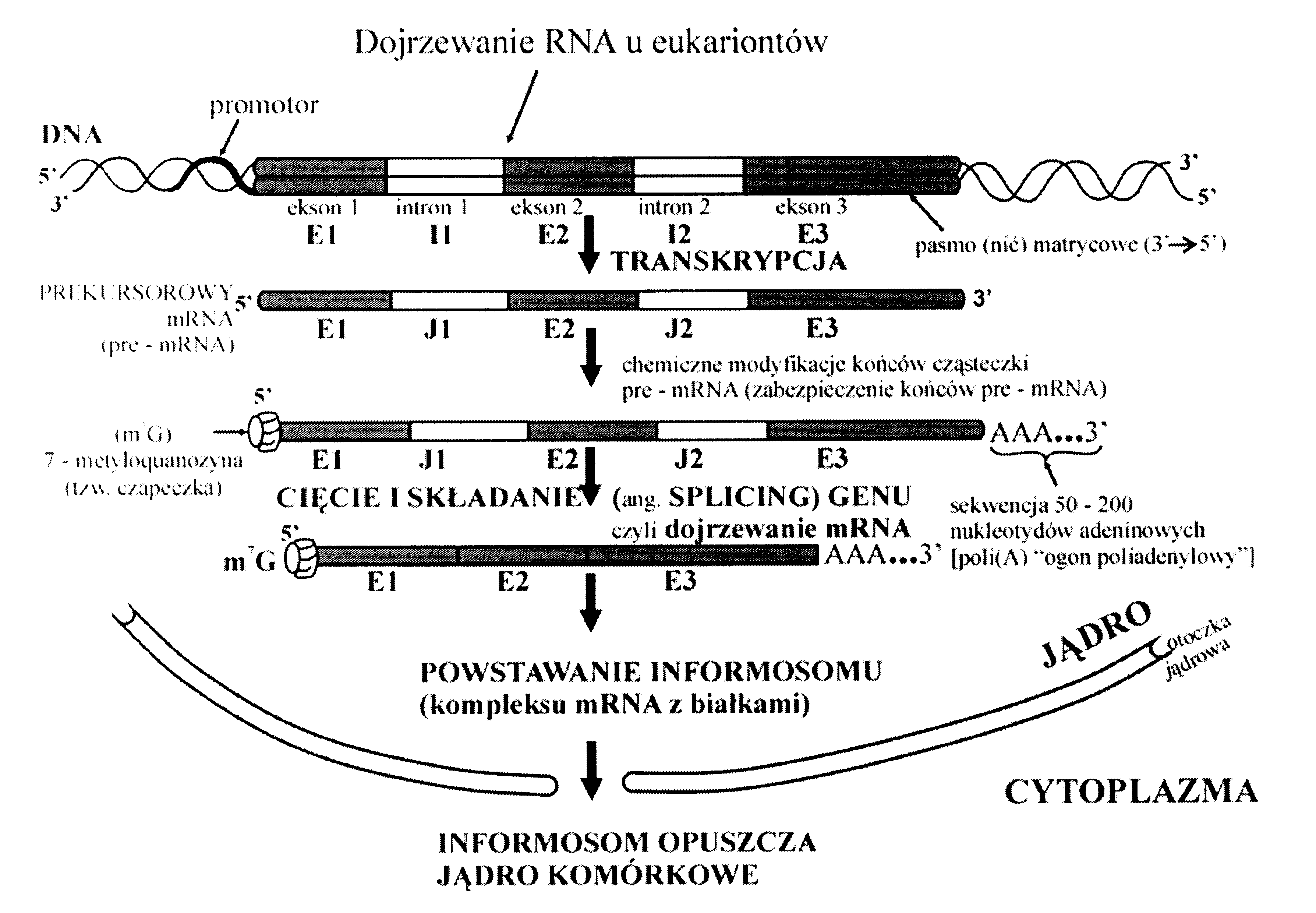
Eukariotyczne cząsteczki mRNA są poddawane jeszcze innym modyfikacjom przed rozpoczęciem translacji. Jeszcze podczas transkrypcji do początku 5' RNA przyłącza się nukleotyd guaninowy (wiąże się z poprzednim nukleotydem nietypowym wiązaniem 5'-5'), który nosi nazwę czapeczki (cap). Do drugiego końca mRNA przyłącza się ogonek kilkuset nukleotydów adeninowych; ten proces nosi nazwę poliadenylacji. Takie modyfikacje mRNA ułatwiają translację, chronią cząsteczki mRNA przed działaniem nukleaz (enzymów rozkładających kwasy nukleinowe) i umożliwiają transport mRNA z jądra komórkowego do cytoplazmy (cząsteczki mRNA pozbawione czapeczki nie opuszczają jądra komórkowego). Zaopatrzony w czapeczkę i ogonek teraz już mRNA nosi nazwę informosomu
Organizmy prokariotyczne nie muszą męczyć się wycinaniem intronów, bo geny prokariotyczne prawie nigdy nie zawierają sekwencji niekodujących (wyjątek Archebacterie). Dlatego komórki prokariotyczne nie potrzebują składać swoich genów. U Prokaryota nie występuje także poliadenylacja i przyłączanie czapeczki.
Czym jeszcze różnią się prokariotyczne cząsteczki mRNA od eukariotycznych? Eukariotyczny mRNA jest najczęściej bardziej stabilny. Poza tym prokariotyczne cząsteczki mRNA często kodują strukturę kilku rożnych białek (powstają w wyniku transkrypcji kilku genów położonych jeden za drugim). Eukariotyczne cząsteczki mRNA zawsze kodują tylko jeden rodzaj białka. No i w komórkach prokariotycznych nie ma otoczki jądrowej, która oddzielałaby geny od cytoplazmy, więc w takich komórkach translacja może się zaczynać jeszcze przed zakończeniem transkrypcji (koniec 3' cząsteczki mRNA nie jest jeszcze gotowy, ale od początku 5' tej samej cząsteczki już rozpoczyna się translacja).
W komórkach eukariotycznych po zakończeniu splicingu i poliadenylacji gotowe cząsteczki mRNA wiążą się ze specjalnymi białkami, które transportują mRNA przez pory jądrowe z jądra do cytoplazmy. Dopiero wtedy zaczyna się... ...translacja.
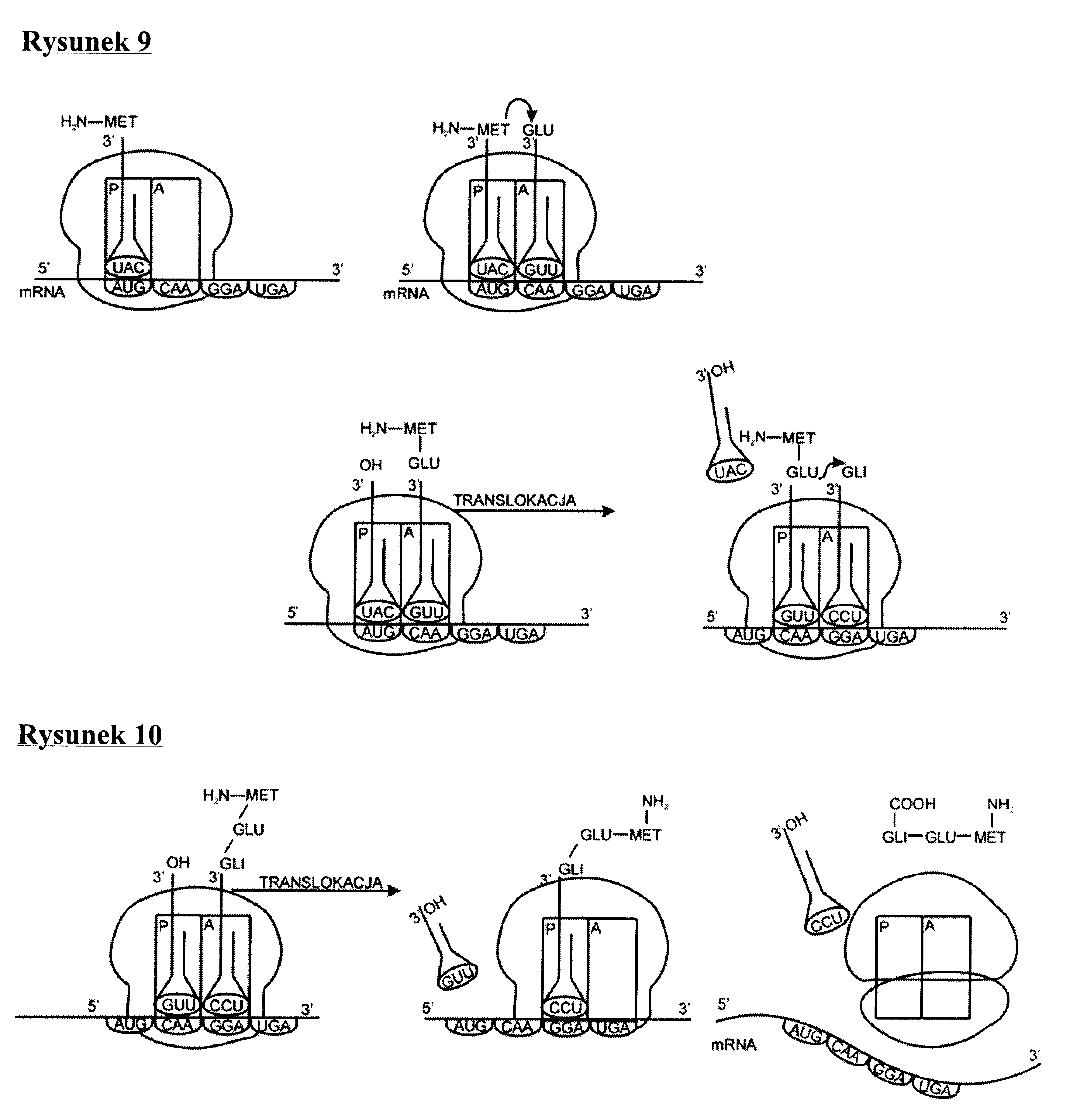
Podczas translacji cząsteczka mRNA wiąże się z rybosomami - małymi organellami zbudowanymi z cząsteczek rRNA (rybosomalnego RNA) i białek.
Każdy rybosom składa się z dwóch podjednostek (mniejszej i większej). Przed rozpoczęciem translacji obie podjednostki pływają osobno w cytoplazmie; łączą się dopiero na cząsteczce mRNA w obecności jonów magnezu.
Rybosomy prokariotyczne i eukariotyczne są podobne, ale nie takie same: prokariotyczne są trochę mniejsze i lżejsze. (W dużej podjednostce rybosomu eukariotycznego są trzy cząsteczki rRNA i ok. 49 białek, a w dużej podjednostce rybosomu prokariotycznego są dwie cząsteczki rRNA i 34 białka. Mała podjednostka eukariotycznego rybosomu składa się z ok. 33 białek i jednej cząsteczki rRNA, a mała podjednostka rybosomu prokariotycznego - z jednej, mniejszej cząsteczki rRNA i 21 białek.) Masę rybosomów określa się w tzw. jednostkach sedymentacji [S](Theodora Svedberga), które opisują zachowanie rybosomów podczas szybkiego wirowania w ultrawirówce. Rybosom prokariotyczny 'waży' 70S, a eukariotyczny - 80S.
Podczas translacji rybosom przesuwa się po cząsteczce mRNA. Każdemu kodonowi (trójce nukleotydów) w RNA odpowiada jeden aminokwas w powstającym białku. Nowe aminokwasy (cegiełki służące do budowy białek) są dostarczane do rybosomu przez cząsteczki transportowego RNA (aa- tRNA, czyli aminoacylo-tRNA). Odpowiednie trójki (antykodony) tRNA pasują do kodonów mRNA i w ten sposób język nukleotydów jest tłumaczony na język białka. W rybosomie znajdują się dwa miejsca, w które wchodzą cząsteczki tRNA: miejsce aminoacylowe, do którego trafia aa- tRNA niosące nowy aminokwas, i miejsce peptydylowe, w którym znajduje się aa - tRNA wiążące wydłużający się łańcuch polipeptydowy.
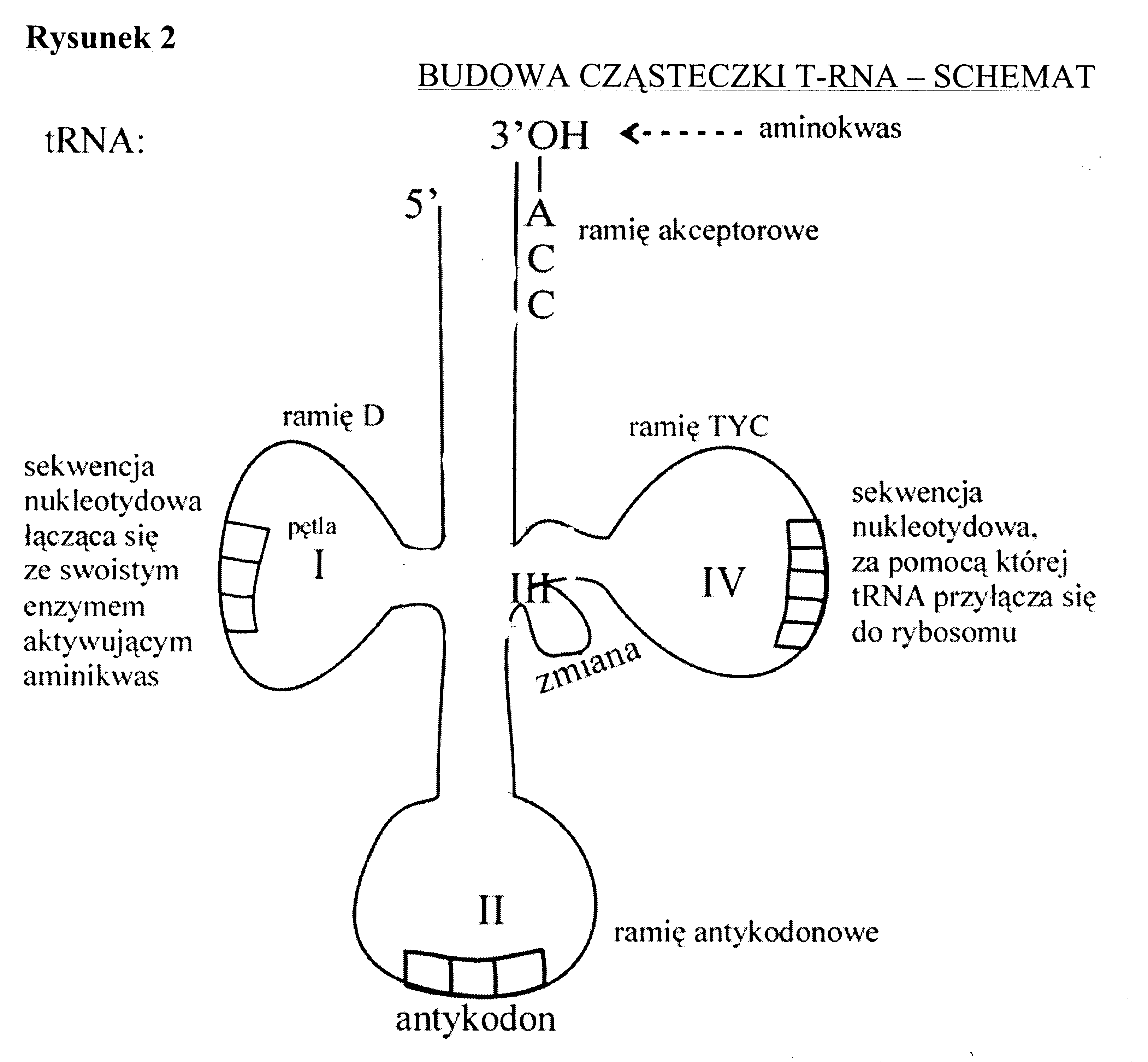
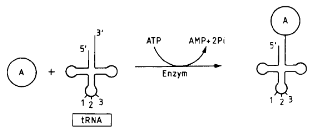
Translacja rozpoczyna się od kodonu AUG kodującego metioninę, a kończy na jednym z terminatorów, kodonów STOP czyli UAA lub UAG lub UGA. Długość peptydu zależy od ilości nukleotydów genu kodującego dane białko. Jeżeli miał ich 300 to białko ma 100 aminokwasów, bo kod genetyczny ma charakter trójkowy. Dzięki badaniom Nienberga, Ochoa i Korany powstała tabela kodu genetycznego zawierająca 64 kodony. (4 rodzaje nukleotydów A,G,U,C, do potęgi 3= 64 KOMBINACJE). Kod genetyczny posiada kilka istotnych cech. Jak wspomniałam wyżej jest trójkowy, poza tym bezprzecinkowy - kodony nie są oddzielone żadnymi elementami, niezachodzący - kodony nie mają wspólnych nukleotydów (wyjątek niektóre wirusy), kolinearny - kolejność kodonów odpowiada kolejności kodowanych przez nie aminokwasów, jednoznaczny( zdeterminowany)-dana trójka koduje jeden aminokwas, zdegenerowany-jeden aminokwas jest kodowany przez więcej niż jeden kodon (wynika to z nadmiaru kodonów - 61 do 20 aminokwasów białkowych),
Pośredni charakter kodu genetycznego podkreśla rolę RNA jak wykonawcy informacji zapisanej w DNA,
zamrożony kod genetyczny tzn. , że jest tak doskonały, że nie podlega już ewolucji.
uniwersalny - kodony są jednakowe u wszystkich, żywych organizmów (przykłady wyjątków : odstępstwa od uniwersalności kodu genetycznego znaleziono w mitochondriach np. UGA sygnał STOP u Eucaryota w organellach półautonomicznych koduje tryptofan, a AGA kodująca argininę jest sygnałem STOP dla mitochondriów).
Podwójną funkcję pełni kodon AUG będący inicjatorem translacji i kodujący metioninę. Kodony STOP nic nie kodują , sygnalizują koniec translacji.
Po zakończeniu biosyntezy białka nowa cząsteczka białka jest uwalniana z rybosomu; rybosom rozpada się na dwie podjednostki, a cząsteczka mRNA służy do produkcji nowej cząsteczki białka albo jest rozkładana na pojedyncze nukleotydy przez specjalne enzymy. Białka często podlegają obróbce posttranslacyjnej polegającej na fosforylacji, czy glikolizacji…. ale to już zachodzi w aparacie Golgiego.
Cała ekspresja jest anaboliczna, czyli wymaga nakładu energii. W transkrypcji jej źródłem są trifosfonukleozydy, a w translacji ATP i każdy z tych procesów dzieli się na etapy, które nazywają się : inicjacja elongacja i terminacja
Podstawowe różnice w replikacji i ekspresji genów u Procaryota i Eucaryota
Organizmy Procarytota |
Organizmy Eucaryota |
Genofor kolisty w cytoplazmie |
Chromatyna w kariolimfie z wolnymi końcami -liniowa |
Brak histonów (DNA nagi - striptisowy ) Brak sekwencji powtarzających się w DNA |
Obecne histony H1, H2A, H2B, H3. H4 -białka reliktowe Obecność sekwencji powtarzających się w DNA |
Jedno miejsce replikacji - origin 50 000 nukl/min - szybkość replikacji brak telomerazy mniejsza szybkość replikacji całego genomu |
Wiele miejsc replikacji 500 - 3600 nukl/min - szybkość replikacji większa szybkość replikacji całego genomu obecna telomeraza |
Biosynteza białka (transkrypcja, translacja) zachodzi w cytoplazmie komórki. Jednocześnie zachodzi transkrypcja i translacja. |
Etapy biosyntezy białka rozdzielone w przestrzeni i czasowo: transkrypcja - w jądrze, translacja - w cytoplazmie |
Geny o budowie ciągłej - brak intronów (wyjątek Archebakterie) |
Geny podzielone: eksony i introny |
Genom jest jednym replikonem, nie posiada nadmiarowego DNA |
- składa się z wielu replikonów - posiada „nadmiarowy” (nie kodujący białek) DNA |
Obecna tylko jedna uniwersalna polimeraza RNA |
Obecne trzy polimerazy RNA Polimeraza I RNA- syntetyzuje rRNA Polimeraza II RNA- syntetyzuje m RNA Polimeraza III RNA- syntetyzuje tRNA |
Brak obróbki mRNA, transkrypt pierwotny ulega bezpośrednio translacji i jest policistronowy -(wielogenowy), jeden promotor do kilku genów Okres półtrwania (życia) mRNA - kilka minut |
Potranskrypcyjna obróbka mRNA (hn mRNA → mRNA=dojrzewanie, składanie, spicing. Monocistronowy, a każdy gen ma własny promotor. Okres półtrwania - kilkanaście godzin do kilku dni |
Brak informosomów, nie ma czapeczki i ogonka |
Obecne informosomy (czapeczka , ogonek)
|
Brak takich powtórzeń |
Odcinki repetytywne |
Kodony inicjacyjne: AUG, GUG wyznaczają N- formylometioninę |
Kodon inicjujący AUG wyznacza metioninę |
Geny kodujące enzymy danego szlaku metabolicznego często zorganizowane są w operony- systemy kontrolujące ekspresję |
Nie ma typowych operonów, a regulacja ekspresji jest bardziej złożona, oparta na enzymach i hormonach. |
Systemy kontrolne w komórkach eukariotycznych
sterowanie szybkością transkrypcji ( wyciszacze i wzmacniacze), wpływaja na aktywność polimerazy RNA,
zmienne składanie eksonów,
amplifikacja genu np. liczba kopii dla każdego histonu sięga 500 na jedną komórkę, a rDNA 450 . Amplifikacja czyli zwiększenie liczby kopii odpowiedniego genu dotyczy zapotrzebowania komórki na pewien rodzaj białek szczególnie w okresie jej intensywnych podziałów.
sterowanie stabilnością mRNA. Trwałość mRNA uzyskana przez jego opakowanie (czapeczka i ogonek) pozwala na wielokrotne wykorzystanie do produkcji cząsteczek białkowych na przykład w uczulonym limfocycie B,
hormony regulują aktywnością komórki
enzymy wpływają na szybkość reakcji metabolicznych.
Budowa i rodzaje genów
Gen = termin wprowadzony przez W. L. Johannsena w 1909 r. Może być zlokalizowany w chromosomie lub cytolazmie
( gen pozachromosomowy). Obecnie traktowany jest jako jednostka funkcji - cistron, może podlegać mutacjom, może być podzielony w wyniku crossing-over na mniejsze odcinki- rekony (najmniejsze jednostki genu, które mogą ulec wymianie w wyniku wewnątrzgenowego crossing-over)
Wyróżniamy różne geny , np.
*geny struktury- kodują określone enzymy np. w operonach
*geny regulatorowe- kontrolują działanie genów struktury (operon)
*geny podzielone- eksony i introny (Eucaryota i Archebacterie, wirusy niektóre)
*geny nakładające się - wirusy wykorzystujące tą samą sekwencję nukleotydów do kodowania różnych polipeptydów
*geny dominujące - „silne” , ujawniajace się w homo- i heterozygocie (współdziałanie alleliczne )
*geny recesywne- „słabe”jedynie w stanie homozygotycznym ujawniaja się (współdziałanie alleliczne)
*geny letalne- śmiercionośne
*geny subletalne - upośledzające czynności życiowe
* geny plejotropowe - wpływające na więcej niż jedna cechę
* geny kumulatywne - (wielokrotne) o niewielkich efektach indywidualnych, współdziałające ze sobą w wytwarzaniu
jednej cechy ilościowej,
* geny zmutowane
*geny „wędrujące”- transpozony
*onkogeny - geny odpowiedzialne za powstawanie raka
5
Wyszukiwarka
Podobne podstrony:
MAKROELEMENTY, Matura, Biologia Matura, składniki chemiczne komórek
Informatyka poziom rozszerzony Matura 2013 id 214067
Informatyka poziom podstawowy c Matura 2013 id 214065
Składniki pokarmowe, Matura, BIOLOGIA, BIOLOGIA MATURA
Mikroelementy i makroelementy, Matura, Biologia Matura, składniki chemiczne komórek
21.Budowa i znaczenie chromosomów jako nośników informacji, studia-biologia, Opracowane pytania do l
skurcz miesni, Biologia maturalna
WIRUSY powtórka, Biologia maturalna
wirusy zadania maturalne, Biologia maturalna
Ciąża2, BIOLOGIA MATURA, Ukł.rozrodczy
hormonalny zad, BIOLOGIA MATURA
U.Poik. 1 cz.(1)(1), BIOLOGIA MATURA
Badanie układów wpisywania i odczytu informacji cyfrowej, Klasa
3. PROTISTY, MATURA, Biologia matura, notatki, 1. INNE ORGANIZMY (Notatki-BIOLOGIA)
więcej podobnych podstron