FITOCHROM?
Fitochrom jest to barwnik, który uczestniczy w procesie związanym z pochłanianiem widma słonecznego w zakresie światła czerwonego i dalekiej czerwieni.
Obecność fitochromu stwierdzono u wszystkich roślin. Barwnik ten funkcjonuje w dwóch formach molekularnych : Pr - wykazuje maksimum absorbcji w zakresie światła czerwonego - 660 nm , aPfr - pochłania światło dalekiej czewieni - 730 nm.(Może dochodzić do przekształcenia jednej formy w drugą). Izolowany fitochrom jest rozpuszczalną w wodzie chromoproteiną o barwie błękitnej. Fitochrom jest zbudowany z polipeptydu połączonego z grupą chromoforową. Wyróżniono dwie fizjologiczne populacje fitochromu : labilną (PI) i stabilną (PII). Fitochrom labilny oznacza się jako fitochrom A , a stabilny jako fitochrom B. Przyjmuje się, że tylko Pfr jest aktywną formą fitochromu . Pfr i Pr absorbują światło czerwone ale daleką czerwień już tylko Pfr. Pochłonięciu światła przez Pr towarzyszy uruchomienie łańcucha przemian metabolicznych.
Przypuszcza się,że fitochrom labilny i stabilny działają niezależnie od siebie lub tez współdziałają w kontroli wzrostu i rozwoju roślin. Fitochrom kontroluje także deetiolację (przejście od heterotroficznej do autotroficznej formy odżywiania roślin).
Procesy w których pośredniczy fitochrom to :
stymulacja kiełkowania
hamowanie wydłużania się łodygi
stymulacja wzrostu powierzchni liści
stymulacja kwitnienia
FAZA CIEMNA FOTOSYNTEZY PRZEBIEG I ZNACZENIE.
Stroma to miejsce fazy ciemnej fotosyntezy w której ATP i NADPH wytworzone w fazie świetlnej są używane do redukcji CO2 i wbudowania go w związki organiczne. Oprócz enzymów zaangażowanych w asymilację CO2 ( cykl Calvina Bensona ) w stromie znajdują się DNA ( genom) chloroplastowy oraz plastytowy aparat biosyntezy białka. Reakcje ciemnej fazy zachodzą w stromie gdzie znajdują się wszystkie enzymy uczestniczące w tym procesie. W stromie zachodzi proces karboksylacji katalizowany przez karboksylazę 1,5-bifosforybulozy. Siła asmilacyjna redukuje grupę karboksylową wytworzoną po związaniu CO2 do grupy aldehydowej czyli do poziomu cukrów. Przemiany fazy ciemnej mają charakter cykliczny i noszą nazwę cyklu Calvina. W cyklu Calvina rozróżnia się 3 fazy :
1)karboksylacja - przyłączenie CO2 do odpowiednich związków organicznych. Karboksylacja związana z aktywnością 1,5 bifosforybulozy i utworzeniem 2 cząsteczek kwasu 3-fosfoglicerynowego (PGA). Reakcja ta przebiega ze spadkiem energii swobodnej co powoduje jej nieodwracalność nawet gdy stężenie CO2 jest małe.
PGA - trwały produkt wiązania CO2 ulega przemianom w związki pośrednie służące do syntezy aminokwasów, białek, tłuszczów. Zawiera grupę COOH.
2)Redukcja - do zredukowania związków organicznych potrzeba wodoru i energii. Czynników tych dostarcza NADPH ( wodoru) ATP ( energii). Faza redukcji polega na wytworzeniu fosfotriozy (aldehyd 3 fosfoglicerynowy). Uczestniczą takie enzymy jak kinaza fosfoglicerynianowa i dehydrogenaza aldehydu 3- fosfoglicerynowego.
3)Regeneracja- odtwarza się związek wiążący CO2 ( 1,5-bifosforybulozy). Wymaga to udziału enzymów: aldolazy, transketolazy, fosfatazy, izomerazy, epimerazy i kinazy.
GLIOKSYSOMY - mikrociała charakterystyczne tylko dla roślin, nie wiadomo w jaki sposób powstają. Mogą przekształcać się w peroksysomy. Glioksysomy przekształcają się w peroksysomy przez zanik enzymów cyklu glioksalanowego i wzrost aktywności enzymów szlaku glikolanowego. Obecność glioksysomów stwierdza się wyłącznie w liścieniach oraz komórkach bielma oraz warstwie aleuronowej nasion wyspecjalizowanych w gromadzeniu tłuszczów jako materiału zapasowego. Zawarte w glioksysomach umożliwiają przekształcenie lipidów w cukry wykorzystywane podczas kiełkowania nasion przez rozwijający się zarodek.
Peroksysomy : srednica 0,1-1,5 um otoczone pojedyńczą błoną we wnętrzu obserwuje się amorficzne i włókienkowate wkręty tzw. nukleoidy.
Glioksysomy zawierają enzymy cyklu glioksalowego umożliwiają szybki rozkład kwasów tłuszczowych i zamianę na cukier. Proces ten odbywa się przy udziale mitochondriów i cytoplazmy podstawowej.
OPIS CYKLU KALVINA BENSONA W FOTOSYNTEZIE.
W cyklu Calvina-Bensona wyróżnia się 3 fazy :
- karboksylacyjną (utworzenie 2 cząsteczek kwasu 3-fosfoglicerynowego)
- redukcyjną (polega na wytworzeniu fosfotriozy)
- regeneracyjną (polega na odtworzeniu 1,5-bisfosforybulozy)
przebieg: Karboksylaza 1,5-bisfosforybulozy przyłącza CO2 do 1,5-bisfosforybulozy dając nietrwały związek 6-węglowy, który rozpada się na dwie cząsteczki kwasu 3- fosfoglicerynowego. Kwas 3-fosfoglicerynowy jest następnie fosforylowany do kwasu 1,3-bisfosfoglicerynowego, który z udziałem NADPH ulega redukcji do poziomu cukrów , tj.do aldehydu 3-fosfoglicerynowego. Na 6 związanych cząsteczek CO2 można wyprowadzić zcyklu 2 cząsteczki trioz lub 1 cząsteczkę heksozy, które mogą być użyte do syntezy sacharozy lub skrobi, zaś pozostałe związki służą do odtworzenia akceptora dwutlenku węgla.
CO TO JEST GLIKOLIZA I JAKIE JEST JEJ ZNACZENIE W PROCESACH KATABOLICZNYCH ROŚLIN ?
Glikoliza odbywa się w cytozolu z rozkładu skrobi powstaje glukoza . Następne reakcje to utworzenie fruktozo -1, 6 - bifosforanu i jego rozłożenie, utlenienie do pirogronianu. Najważniejszym momentem glikolizy jest utlenienie aldehydu 3- fosfoglicerynowego do 3- fosfoglicerynianu. Uwalnia się energia użyta do fosforylacji i ADP , następuje redukcja NAD+. Przy przemianie 3- fosfoglicerynianu w pirogronian następuje druga fosforylacja substratowa . W sumie podczas glikolizy powstają z jednej glukozy 4 ATP ( dwie zużywane przy fosforylacji cukrów) 2NADH i dwa pirogroniany. Pirogronian przechodzi z cytozolu do mitochondrium i tam ulega dalszym przemianom- ostatecznemu utlenieniu do CO2 i H2O .
CYKL KREBSA - PRZEBIEG I ZNACZENIE
Cykl Krebsa znany też jako cykl kwasu cytrynowego to proces w którym pochodzący od pirogronianu octan zostaje utleniony do CO2 i H2O. Powstają przy tym kolejne cząsteczki NADH i jedna FADH 2 . Enzymy tego cyklu znajdują się w macierzy mitochondrialnej ( matrix) i tam odbywają się reakcje cyklu. Głównym substratem cyklu Krebsa jest pirogronian , który najpierw jest przetransportowany z cytoplazmy do wnętrza mitochondrium - w matrix pirogronian ulega karboksylacji do acetylokoenzymu A. Acetylokoenzym A w poszczególnych ogniwach cyklu Krebsa ulega :
-dwukrotnej dekarboksylacji z wydzieleniem dwóch cząsteczek CO2 oraz czterokrotnej dehydrogenacji z wydzieleniem 3 cząsteczek NADH i jednej FADH2
Poszczególne etapy cyklu Krebsa:
- przyłączenie grupy acetylo-CoA do szczawiooctanu i powstanie cytrynianu.
-Cytrynian przechodzi w izocytrynian który w procesie dekarboksylacji daje oksoglutaron, NADH i CO2
-Oksoglutaron podlega także procesowi dekarboksylacji, przyłącza się znów acetylo- CoA i powstaje sukcynylo- CoA, NADH i CO2
-Sukcynylo-CoA przekształca się w bursztynian, uwalnia się wtedy energia umożliwiająca syntezę ATP na drodze fosforylacji substratowej
-Bursztynian jest utleniany na fumaron. Powstaje wtedy FADH2
-Fumaron przekształca się w jabłczan.
-Jabłczan utleniany jest w szczawiooctan i jednocześnie powstaje cząsteczka NADH.
Zregenerowany szczawiooctan zdolny jest do reakcji z kolejnym acetylo-CoA. W ten sposób cykl przemian zostaje zamknięty i może rozpocząć się od nowa.
Znaczenie cyklu:
- w cyklu tym następuje utlenienie cukrów a także tłuszczów i białek przekształconych na czynny octan.
- jest też ważnym dostawcą metabolitów do różnych syntez
-stanowi pulę różnych łańcuchów węglowych substratów do różnorodnych procesów biochemicznych w komórkach roślinnych
-w tkankach niefotosyntetyzujących jest jedynym źródłem ATP i zredukowanych nukleotydów do syntezy nowych związków.
FOTOSYNTEZA- TRANSPORT ELEKTRONÓW I ZNACZENIE W METABOLIŹMIE
Elektrony wędrują z H2O na NADP+ przez łańcuch przenośników. Jest to szereg układów oksydoredukcyjnych , których większość została wyizolowana i scharakteryzowana. Transport elektronów w fazie świetlnej fotosyntezy odbywa się z udziałem 3 dużych kompleksów białkowych. Są to fotosystem II, kompleks cytochromowy b6-f i fotosystem I . PSII po utracie elektronów nabiera silnych właściwości utleniających pozwalających na utlenienie wody , a pochodne chlrofilu a 680 (P680) elektrony przenoszone są za pośrednictwem plastochinionu na kompleks cytochromowy b6-f . Z kompleksu cytochromów przechodzą na PS I za pośrednictwem plastocyjaniny. Wybicie elektronu z chlorofilu a700 ( P700) powoduje pojawienie się właściwości utleniających pozwalających na utlenienie cytochromów oraz silnych zdolności redukcyjnych prowadzących do redukcji NADP+. Podczas wędrówki elektronu następują stopniowe zmiany potencjałów oksydoredukcyjnych kolejnych przenośników. Występuje tu paradoksalna sytuacja, gdzie układ o wysokim potencjale dodatnim redukuje układ o wysokim potencjale ujemnych. Redukcja taka wymaga dużego nakładu energii. Fotosysem II (PS II) działający z maksymalną efektywnością przy 680 nm długości fali światła zlokalizowany jest głównie w tylakoidach gran. Wybicie elektronów z centrum reakcji PS II powoduje odciągnięcie elektronu z H2O co wiąże się z wydzieleniem O2 i uwolnieniem protonów ( H+). Przyjęcie elektronu z H2O powoduje powrót P680 do formy zredukowanej, a elektron wybity przedtem przez foton zostaje przekazany na feofitynę ( związek podobny do chlorofilu). Z feofityny elektrony przechodzą na plastochiniony. Ostatni z nich przyjmując elektron pobiera ze stromy protony i przekształca w hydroplastochiniony. W tej postaci zanurza się w błonie i przekazuje elektrony na układ cytochromowy, a protony uwalnia do wnętrza tylakoidu.
Mechanizm wędrówki elektronu przez kompleks cytochromowy nie jest dokładnie poznany. Fotosytem I czyli PSI występuje głównie w tylakoidach stromy. Centrum reakcji PS I po wybiciu z niego elektronu przez kolejny foton staje się słabym utleniaczem odbierającym elektron od plastocyjaniny, która łącznie z plastochinionem pełni funkcję ruchliwego łącznika między PS II a PS I . Równocześnie PS I staje się silnym reduktorem oddającym elektrony na kolejne przenośniki. Pierwsze układy pośredniczące w przekazywaniu elektronu z PS I są bardzo nietrwałe. Znane są natomiast następne, są to żelazo-siarkowe białka które przenoszą elektrony na ferrodoksynę. W końcu flawoproteinowy enzym przenosi elektron na NADP+ i redukuje go do NADPH. NADPH stanowi końcowy produkt transportu elektronu z H2O. Przepływ elektronu od PS II przez PS I do NADP+ jest to tzw. niecykliczny transport elektronu. Może on być uzupełniany przez przepływ cykliczny w którym ferrodoksyna przekazuje elektron z powrotem na układ cytochromowy b6 -f, a następnie przy współudziale plastochinionu na PS I . Przy okazji zostają do kanału tylakoidowego przeniesione dodatkowe protony. Znaczenie tej wędrówki elektronu jest ogromne. Powstaje bowiem nie tylko NADPH ale również ATP. Są to podstawowe związki wysokoenergetyczne wykorzystywane do i redukcji CO2 i wytwarzania substancji organicznych. Bez tego przepływu nie istniała by fotosynteza.
CZYNNIKI WARUNKUJĄCE INTENSYWNOŚĆ FOTOSYNTEZY
Na efektywność fotosyntetyczną wpływają różne czynniki środowiska. Najważniejsze z nich to: woda CO2,światło, temperatura.
WODA - jej wpływ nie jest bezpośredni, ponieważ zawartość H2O w chloroplastach dla potrzeb fotosyntezy jest wystarczająca.
Jednak wpływa ona silnie na asymilację CO2, gdyż w przypadku deficytu wody zamykają się aparaty szparkowe i praktycznie zostaje odcięty dopływ CO2 z powietrza do liści, prowadząc do spadku stężenia CO2 w przestworach międzykomórkowych. Ponadto jej niedobór hamuje wzrost roślin i w wyniku zmniejszenia blaszek liściowych mniej światła i CO2 jest odbierane
ŚWIATŁO-
Krzywa świetlna obrazuje zmiany natężenia fotosyntezy , mierzone ilością pobranego CO2 pod wpływem różnych natężeń światła.
Przy bardzo niskich natężeniach światła, mimo że fotosynteza się odbywa nie obserwujemy pobierania CO2 , a jedynie zmniejszona jego wydalanie.
Równocześnie z fotosyntezą odbywa się oddychanie, które zużywa tlen i wytwarza CO2. Początkowo fotosynteza zużywa CO2 powstały w procesie oddychania, nie pobierając go z atmosfery. Przy zwiększonym natężeniu światła wymiana gazowa ustaje - brak pobierania lub wydalania CO2 orazO2 . takie stężenie światła w którym dochodzi do zrównania fotosyntezy i oddychania oraz ustania wymiany gazowej to świetlny punkt kompensacyjny. Jego wartość zależy od gatunku i warunków rozwoju drzewa.
Przy ilościach światła powyżej punktu kompensacyjnego następuje liniowy wzrost pobierania CO2, co jest miarę natężenia fotosyntezy netto.
Wzrost ustaje gdy ilość światła osiąga określoną wartość. Powyżej określonego stężenia nie następuje wzrost fotosyntezy. Jest to świetlny punkt wysycenia. Jego wartość zależy od gatunku i warunków rozwoju.
Bardzo wysoki stopień natężenia światła przy długim czasie naświetlania może hamować fotosyntezę poprzez uszkodzenie aparatu asymilacyjnego.
CO2- μmol*m-2*s-1 (fotosynteza)
Wpływ stężenia CO2 na fotosyntezę, której miarą jest stężenie CO2 w jednostce czasu na jednostkę powierzchni liścia. Poczynając od bardzo niskich wartości stopniowe zwiększenie CO2 powoduje wpierw dojście fotosyntezy do stanu równowagi między fotosyntezą a oddychaniem, stężenie w którym ten stan się pojawia nazywamy stężeniem kompensacyjnym , a następnie wzrost natężenia fotosyntezy.
Stężenie CO2 o wartości 0,1% jest optymalna dla rośliny, a 1% już trujące. Korzystne jest większe stężenie CO2 przy większej temperaturze i natlenieniu.
TEMPERATURA-
CO2
μmol*m-2*s-1
(fotosynteza)
Wpływ temperatury na asymilację CO2 przy silnym i słabym natężeniu światła.
Przy niskim natężeniu światła temperatura w zakresie do 30°C nie ma wpływu na fotosyntezę -czynnikiem ograniczającym jest wtedy światło. W wyższych temperaturach natężenie fotosyntezy zmienia się (nasila się fotorespiracja + ogólnie hamujący wpływ temperatury na efektywność wielu enzymów)
Temperatura może być czynnikiem ograniczającym, zwłaszcza w warunkach wysycenia aparatu fotosyntetycznego światłem (powyżej świetlnego punktu wysycenia)przy dobrym zaopatrzeniu w CO2 .Przy wysokim stężeniu światła krzywa zależności fotosyntezy od temperatury ma wyraźne optimum około 30°C jest to wartość charakterystyczna dla reakcji enzymatycznych.
MIEJSCE SYNTEZY , DROGI I MECHANIZMY TRANSPORTU AUKSYNY.
Biosynteza auksyny ( kwasu inohilo-3- octowego IAA) odbywa się w rosnących tkankach ( wierzchołki wzrostu, młode liście, pąki , kwiaty, owoce) z aminokwasu aromatycznego ( tryptofanu) w procesach deaninacji , dekarboksylacji i utleniania. Auksyna jest aktywnie transportowana przez kambium bazypetalnie czyli od końca apikalnego w kierunku bazypetalnego. Kierunek transportu jest związany z biegunowym zróżnicowaniem komórek. O polarnym transporcie auksyny świadczy doświadczenie w którym w pędzie wierzbowym wycięto pierścień kory wraz z łykiem przecinając tym samym drogę aktywnego transportu auksyny. Po nałożeniu na koniec apikalny auksyny, po pewnym czasie okazało się ,że auksyna wywarła hamujący wpływ narozwój pączków śpiących tylko do strefy przecięcia drogi transportu. W dolnej części pędu pąki rozwinęły się w pędy- auksyna nie zadziałała. O transporcie auksyny tylko w kierunku bazypetalnym świadczy doświadczenie w którym auksynę nałożono na koniec bazalny. Nie została ona przetransportowana aktywnie, tym samym nie zahamowała rozwoju pąków śpiących które rozwinęły się w pędy. Szybkość transportu polarnego auksyny wynosi około 10cm /h. Transport tego hormonu nie polega na przemieszczaniu się jej od korony wzdłuż całego pnia. Auksyna jest bodźcem indukującym szereg łańcuchowych reakcji w komórkach wrzecionowatych kambium. Pobudza ona komórki do intensywnych podziałów.
ZNACZENIE GIBERYLINY W PROCESIE WZROSTU I RÓŻNICOWANIA DRZEW.
Giberelinę - hormon roślinny odkrył w latach 20-tych naszego wieku japoński badacz Kurosawa. Z grzyba wywołującego chorobę „wściekłego ryżu” wyizolował on krystaliczną, aktywną substancją zwaną gibereliną. Największe stężenie giberelin występuje w młodych, rozwijających się nasionach i owocach, które stanowią miejsce ich syntezy. Gibereliny są tworzone w wierzchołkowych partiach pędu oraz w korzeniu. Główna, aktywna biologicznie giberelina u roślin to giberelina A1.. Podstawowym przejawem działania giberelin na rośliny jest silna stymulacja wzrostu międzywęźli prowadząca do wydłużania się pędów. Przy pomicy giberelin można przywrócić normalny wzrost i pokrój karłowatym mutantom. Hormony te mogą również wywoływać kwitnienie u roślin wymagających do zakwitnięcia długich dni. Gibereliny działają również antagonistycznie do auksyn, ponieważ pobudzają do rozwoju pąki. Wywołują one również partenokarję, czyli rozwój owoców baz zapłodnienia (np. grusza). Odgrywają one również dużą rolę w procesie przerywania stanu głębokiego spoczynku utworów przetrwalnikowych (nasion), mogą one w tym procesie zastąpić niektóre czynniki jak chłód i światło.
POWSTAWANIE I PRZEMIESZCZANIE SIĘ HORMONÓW WZROSTU
Do regulatorów wzrostu zaliczamy:
A)auksyny - powstają w zielonych częściach roślin w formie nie czynnej jako tzw. proauksyny i stąd rozprowadzane są do części wzrostowych, gdzie przetworzone zostają na formy aktywne. Ze stożków wzrostu spływają w dół floemem. W części nadziemnej rośliny auksyna jest transportowana od wierzchołka pędu do podstawy - jest to transport bazypetalny; natomiast w korzeniu auksyna jest transportowana od podstawy rośliny do wierzchołka korzenia-akropetalny.
B)gibereliny- wytwarzane są głównie w liściach, skąd spływają w dół floemem. Z korzeni mogą „wędrować” w górę ksylemem. Transport giberelin nie ma więc charakteru polarnego.
C)cytokininy- wytwarzane są głównie w stożkach wzrostu u korzeni, skąd wędrują w górę z prądem transpiracyjnym (ksylemem).
inhibitory wzrostu - np. kwas obscysynowy, kwas jasmonowy. Kwas obscysynowy (ABA) powstaje w różnych organach, szczególnie zaś w starych liściach - miejscem syntezy są plastydy. Transportowany jest głównie floemem, ale także miękiszem oraz ksylemem.
INHIBITORY WZROSTU.
ETYLEN - jest to związek o bardzo prostej budowie. Substratem w syntezie jest metionina. Etylen hamuje wzrost wydłużeniowy siewek, przyspiesza procesy dojrzewania, starzenia się tkanek i warstwy odcinającej u liści i owoców.
KWAS ABSCYSYNOWY (ABA) - związek o strukturze przypominającej karotenoidy. Jego synteza odbywa się z kwasu mewalonowego lub pośrednio z karotenoidów. Wytwarzany jest w komórkach zawierających plastydy w organach pod działaniem stresu. Indukuje stan spoczynku w pąkach i w nasionach gdzie jest inhibitorem kiełkowania. Hamuje wzrost objętościowy i wydłużeniowy komórek. Hamuje fotosyntezę , syntezę chlorofilu i transport jonów przez błony, indukuje zamykanie się aparatów szparkowych, przyspiesza starzenie
JASMONIANY- kwas jasmonowy przypomina strukturą ABA . Hamują wzrost elongacyjny pędu , kiełkowanie pyłku i nasion oraz tworzenie się pąków pędowych i kwiatowych. Stymuluje zamykanie się aparatów szparkowych , rozwój bulw , dojrzewanie owoców, rozkład chlorofilu, starzenie się liści. Podobnie jak ABA towarzyszą reakcji roślin na stresy.
ZWIĄZKI FENOLOWE- stanowią wielką grupę substancji produkowanych przez rośliny większość syntetyzowana jest z aminokwasu fenyloalaniny. Są to inhibitory wzrostu i kiełkowania mają udział w allelopatii. Ogólnie nie mają cech typowych hormonów
MECHANIZM TRANSPORTU BIERNEGO I CZYNNEGO
W zależności od rodzaju transportowanej substancji oraz czynników warunkujących pojawienie się transportujących te substancje przez błony wyróżniono kilka sposobów transportu :
Transport bierny - polega na przemieszczaniu się substancji w obu kierunkach ale zawsze zgodnie z gradientem stężeń. W ten sposób gazy (tlen, azot, CO2, NH3) i mocznik oraz niektóre jony dyfundują wraz z prądem wody przez błonę do miejsc mniejszym stężeniu. Transport bierny zachodzi do czasu wyrównania się różnicy stężeń po obu stronach błony.
Dla cząsteczek związków organicznych i niektórych jonów (K+, Cl- ) błony stanowią przeszkodę, którą mogą pokonać tylko przy udziale nośników (białek nośnikowych -ruchliwych składników błon). Jeżeli substancja jest transportowana przez nośnik zgodnie z gradientem stężeń to proces taki nie wymaga nakładu energii i nazywamy go transportem ułatwionym lub ułatwioną dyfuzją.
Transport aktywny wymaga obecności nośników i dopływu energii pochodzącej z przemian metabolicznych. W ten sposób następuje przemieszczanie się nie tylko wielu jonów, lecz również związków organicznych, np. cukrów lub aminokwasów, wbrew gradientowi stężeń. Transport przy udziale nośników jest zwykle jednokierunkowy. Ten typ transportu substancji jest niezbędny do życia komórki ponieważ warunkują pobieranie przez nią składników koniecznych do prawidłowego funkcjonowania nawet wtedy, gdy ich stężenie w środowisku jest bardzo niskie, wówczas wymaga nakładu energii.
Transport cieczy i cząsteczek większych od tych, które mogą być przemieszczane przez pory błon lub przy udziale nośników odbywa się na drodze endocytozy - przenikania do wnętrza komórki wyniku tworzenia się wakuol. Pobieranie cieczy i koloidów następuje w procesie pinocytzy a ciał stałych przez fagocytozę. Fagocytoza wymaga dostarczenia energii. Wypieranie różnych substancji z komórki do środowiska zachodzi w procesie egzocytozy. W ten sposób wydzielane są np. hormony i enzymy oraz produkty przemiany materii.
MECHANIZM TRANSPORTU WE FLOEMIE
Podobnie jak w przypadku transportu ksylemowego, podstawą oceny, jakie związki i jony oraz w jakich ilosciach są transportowane przez floem, jest analiza chemiczna soku floemowego( na podstawie soku wypijanego bezpośrednio z rurek sitowych przez mszyce) Skład chemiczny soku floemowego jest cechą gatunkową ,nie zawiera cukrów redukujących). Przez tą tkanke przewodzącą w czasie starzenia się liści następuje export dużych ilości produktów rozkładu białek w chloroplastach i mitochondriach, zaopatrzenie organów generatywnych w organiczne związki azotowe przemieszczane z organów wegetatywnych. Przez rurki nitowe przemieszczają się wszystkie hormony. We floemie występują również kwasy organiczne, witaminy oraz ATP i ADP
Na przekroju poprzecznym łodygi rurki sitowe stanowią ok. 1/5 pow łyka o sprawnosci transportu floemowego decyduje liniowa szybkość przepływu , jak tez stężenie przemieszczanych związków w soku floemowym. Intensywność transportu;
Intensywność transportu floemowego jest różna dla różnych gat i mniejsza od liniowej intensywności transportu ksylemowego.
Przeciętnie wynosi ona około 0,6-5 g*cm-2*h-1
Czynniki wpływające na transport floemowy:
-temperatura i zmienność dobowa :
a)temp w zakresie ok. 50st C słabo wpływa na ten proces
b)temp ma wpływ na intensywność rozładunku, słabo wpływa na przepływ przez rurki sitowe.
„Siłownia” floemu (centrum syntezy ATP) są komórki towarzyszące.
Transport asymilatów odbywa się przez całą dobę z różną intensywnością , co świadczy o braku zależności tego procesu od fotosyntezy.
Mechanizm transportu floemowego na duże odległości:
A)załadunek floemu może się odbywać w dwojaki sposób;
-przez symplastyczny kontakt-plazmodesmy między komórkami miękiszowymi a komórk towarzyszącymi i rurkami sitowymi
-na drodze apoplastycznej
B)rozładunek floemu- odbywa się w komórkach akceptorów do których transportowane są substancje przemieszczane przez floem (przebiega to zgodnie z malejącym gradientem potencjału chemicznego)
Sprawność rozładunku floemu i metabolizm substancji transportowych determinuje wielkość importu substancji pokarmowych do akceptorów
Rozładunek może odbywać się przez symplast i apoplast:
-akceptory intensywnie rosnące (młode liście) zaopatrywane są przez symplast
-akceptory akumulujące (organy spichrzowe) importują zw węglowe przez apoplast
W organach generatywnych, w rozwijajacych się nasionach, u których brak jest kontaktu symplastycznego między kom zarodka a kom rośliny macierzystej , rozładunek jest apoplastyczny.
Siłą motoryczną ruchu w rurkach sitowych jest gradient ciśnienia hydrostatycznego, czyli turgorowego, miedzy punktem załadunku i rozładunku floemu. W teorii przepływu masowego związki i jony przemieszczają się w rurkach sitowych pod ciśnieniem wraz z wodą, w każdej rurce tylko w jednym kierunku. Gradient potencjału wody między rurami sitowymi i ksylemem powoduje stałą cyrkulację wody.
ksylem- transport wody i składników mineralnych. Do transportu służą naczynia i cewki oraz rurki sitowe ( do dalekiego transportu).
W ksylemie następuje przemieszczanie pomiędzy organami rośliny. Woda w ksylemie przemieszcza się do wyżej położonych części roślin. Słupy wody podciągane są do góry dzięki ssącemu działaniu parowania tworząc tzw. prąd transpiracyjny. Szybkość transportu produktów zależy od intensywności transpiracji. Intensywność transpiracji nie ma bezpośredniego wpływu na ilość substancji przetransportowanych do górnej części rośliny.
a)Parcie korzeniowe jest jednym z elementów wpływających na transpirację.
b)Transpiracja oraz teoria kohezyjno - transpiracyjna.
Siły kohezyjne powodują wzajemne przyciąganie się cząsteczek wody. Utrzymują słup pod napięciem. O szybkości przepływu decyduje powierzchnia systemu przewodzącego. Siły adhezyjne (przylegania)- wytwarzanie wiązań wodorowych między cząsteczkami wody i polarnymi grupami - OH, np. celulozy, z której zbudowane są ściany naczyń.
KWITNIENIE
Po zakończeniu okresu młodocianego (juwenilnego) -różn.weget.roś.wchodzi w okr.ROZW.GENERATYWNEGO.Rośl.uzyskuje stan gotowości do KWITNIENIA.U ros.1-rocznych czy wielol. Kwitn.tylko raz(ROS.MONOKARPICZNE)wytw.kwiatów prowadzi do starzenia i obumierania. Podobnie jest z niekt.roś.2-letnimi(np.rozetowymi),wytw. pędy kwiatowe i kwiaty w 2 roku weget. U roś.wielol. kwitnących wielokrotnie(ROŚ.POLIKARPICZNE)wyst. często zjaw. niejednoczesności pochodzenia z fazy młodej do dojrzałej w poszczególnych część. org. Najwcześniej osiąg. fazę dojrzałości wierzchołki najwyż. pędów, zaś pędy bliżej podstawy zachow. dłuz. cechy młodociane(ZJAWISKO TOPOFIZY)
W regulacji pochodz. roś. z f.weget. do generat. uczestniczą sterowane genetycznie CZYNN.WEWNĘTRZNE- reak. endogenne i CZYNN.ŚRODOWISKOWE. (ich udział nie u wszystk.roś jest jednakowy)
GOTOWOSC DO KWITN. Związ. z osiągnięciem przez roś. pewn. minim. wlk. lub decyduje odpowiednio duża licz. podz. kom. w merystemie wierzchołk. pędu. Inna hipoteza utrzym. stanu młodocianego :zbyt bliskie sąsiedztwo korzenia i pędu. Boczne lub szczyt. wierz. pędu przekszt. się w generat. dop.wtedy gdy gdy będę daleko korzeni. Przypuszcza się też ,że roś.posiada zdolność do tw. poza wierz. wzr. okr. impulsów czy czynn.(induktorów)kwitn. kt. przemieszcz. się w roś. powodują przejście do f.generat.(uruchomienie aktywnosci okr. genów kwitn)
Po uzyskaniu gotowości do kwitn.zmienia wzorzec różnicowania się wierz.wzr.W meryst.wierz.zamiast zawiąz.lisc. i pąków kw. różn. się zaw.kwiatowe.U roś.rozetowych-meryst.częsci podwierzch.-poczatek rozw.pedu kw. Rozpocz. się pr.morfogenezy generatywnej, koń. się wytw. gamet i osiągn. dojrzałości płc. Roś. kwiat.wytw.nasiona,a okrytozal.również owoce.
W morfog. gener. uaktywn. się szereg genów odpowiedz.za zjaw. Indukcji i rózn.się poszczególnych elem. kwiatów-genów kwiatowych. Również czyn.zewn. i odpow. Warunki termiczne oraz war. świetlne. Roś. niewrażliwe w niesprzyjaj.warun. pozostają weget. Roś. wrażliwe przechodzą do f. gener. w wyniku indukcji kwitn., kt. Może być powod. Temp.(termoindukcja, wernalizacja, jaryzacja) bądź światłem (indukcja fotoperiodyczna, fotoindukcja generatywna)
ZNACZENIE FOTOINDUKCJI I TERMOINDUKCJI W PROCESIE WZROSTU I ROZWOJU ROŚLIN.
Światło jest czynnikiem silnie wpływającym na morfogenezę rośliny. Pęd siewki rozwijający się w ciemności jest blady i wiotki. Pozbawiony jest chlorofilu, hipokotyl i międzywęźla są wydłużone, blaszki liściowe zredukowane, wierzchołek zgięty w dół. Zjawisko to nosi nazwę etiolacji (wypłowienia). Na świetle pęd zazielenia się, łodyga skraca, blaszki liściowe są dobrze rozwinięte, wierzchołek wyprostowany. Cechy odróżniające rozwijającą się w ciemności od rośliny rozwijającej się w świetle to fotomorfozy. Ogromna rola światła w rozwoju roślin jest związana z fotosyntezą, lecz znaczenie światła nie ogranicza się do fotosyntezy i produkcji asymilatów, ponieważ zmiany w rozwoju siewki pod wpływem światła nie zależą od fotosyntezy. Dowodzi tego bardzo niskie stężenie światła lub bardzo krótki czas naświetlenia jaki wystarcza do wywołania fotomorfozy. W przypadku wypłoweiałych siewek wystarczy pojedynczy błysk słabego światła, aby wywołać po 10 minutach spowolnienie wzrostu łodygi, zapoczątkowanie wyprostowania się wierzchołka pędu i syntezy chlorofilu. Światło działa tu nie jako źródło energii, ale jako sygnał do wywołania zmian rozwojowych. Nosi to nazwę FOTOINDUKCJI. Inne fotomorfozy to: zmiana kształtu komórek szparkowych ( wraz z otwieraniem się szparek), kiełkowanie niektórych nasion. Indukcja biegunowości w komórkach, różnicowanie się przedrośla u paproci, synteza karotenoidów i wzrost sporangioforów u grzybów. Fotomorfozy zachodzą pod wpływem światła czerwonego i niebieskiego.
U wielu roślin konieczne jest działanie niskich temperatur, w celu zainicjowania kwitnienia, lub przynajmniej niska temperatura kwitnienie przyspiesza . Działanie chłodu ma tu charakter bodźcowy i nazywa się TERMOINDUKCJĄ. Indukowany niską temp. Umożliwiający kwitnienie nosi nazwę wernalizacji lub jaryzacji. Termoindukcja zachodzi pod wpływem temp. Od nieco poniżej zera do około 10oC. Wymagany jest przy tym dłuższy czas działania niskiej temp. co najmniej 3-8 tygodni. Gdy w tym czasie proces wernalizacji zostanie przerwany przez podwyższenie temp., efekt termoindukcji nie pojawi się (dewernalizacja). Wrażliwość roślin na okresowe działanie chłodu jest różna: od niewrażliwości ( u roślin jednorocznych ), poprzez przyspieszenie kwitnienia, do pełnej zależności kwitnienia od termoindukcji. Rośliny ozime przechodzą wernalizację w stadium spęczniałego nasienia. Rośliny 2-letnie stają się wrażliwe na wernalizację po osiągnięciu pełnego rozwoju wegetatywnego.
Miejscem percepcji bodżca wernalizacyjnego jest merystem wierzchołkowy pędu. W laboratorium można chłodźić same wierzchołki pędów i doprowadzać rośliny do kwitnienia. U roślin 2-letnich wymagających wernalizacji można ją zastąpić podaną z zewn. gibereliną, która wymagana jest głównie do „wystrzelenia” wysokiego pędu kwiatonośnego ( rośliny rozetkowe).
OSMOZA
Jest to niezależne od nakładu energii, spontaniczne przemieszczenie się wody przez błonę zgodnie z gradientem stężenia wody po obu stronach błony. Błony kom. rośl. Są selektywnie przepuszczalne. W przemieszcz. się subst. biorą udział białka transportowe a w przemieszcz. Się wody przez błony biorą udział akwaporyny.O tym przemieszcz. Decydują 2 czynniki: gradient wody i gradient ciśnienia. Ciśnienie, które trzeba by przyłożyć aby przeciwdziałać przepływowi przez błonę zgodnie z gradientem potencjałów: z ośrodka zawierającego czystą wode do ośrodka zawier. Roztwór wodny jakiejś subst. nazywamy ciśnieniem osmotycznym. Wzrasta ono wraz z temp. I z liczbą rozpuszczonych w wodzie cząsteczek, które dalej dysocjują. Ciśnienie osmotyczne rozcieńczonego roztworu jest równe ciśnieniu, jakie wywierałaby dana subst. w stanie gazowym w tej samej temp. i objętości.
CO TO JEST POTENCJAŁ WODY I JAKIE JEST JEGO ZNACZENIE ?
O przechodzeniu wody z jednej komórki do drugiej decyduje różnica pomiędzy ich siłami ssącymi, co wyraża się równaniem S=W-T, gdzie W-oznacza wartość osmotyczną soku komórkowego, a T ciśnienie turgorowe komórki.
Jednakże w ujęciu ogólnym siła ssąca komórek nie polega wyłącznie na zjawiskach osmotycznych, gdyż w grę wchodzą także: pęcznienie różnych substancji, a zwłaszcza biokoloidów, adchezja itp. W celu całościowego rozumienia stanu uwodnienia komórek, tkanek i organizmów roślinnych należy oprzeć się na termodynamicznym pojęciu potencjału wody.
O wielkości potencjału wody stanowi stosunek różnicy między wartością potencjału chemicznego wody danego roztworu lub danej substancji i potencjału chemicznego czystej wody do cząstkowej objętości molalnej wody w rozpatrywanym układzie:
![]()
μw-potencjał chemiczny wody roztworu
μwo- potencjał chemiczny czystej wody
Vw - cząstkowa objętość molalna wody układu
Chemiczny potencjał wody można rozumieć intuicyjnie jako stan nią w danym układzie. Jest on miarą zdolności danej substancji do wykonywana pracy. Wartość potencjału chemicznego czystej wody pod ciśnieniem 1 atmosfery przyjmuje się za 0; stąd Ψ ma wartość ujemną.
Przepływ wody pomiędzy dwoma układami skierowani jest do bardziej negatywnej wartości Ψ(czyli do układu mniej uwodnionego w stosunku do jego możliwości ).
W układach nieożywionych Ψwykazuje przy normalnym ciśnieniu wartość ujemną, to jednak w organizmach żywych nie zawsze tak musi być. Przykładem mogą być gutujące rośliny, które „aktywnie” wydzielają wodę.
Potencjał wody wyrażamy w jednostkach ciśnienia.
FUNKCJE WODY W ROŚLINIE:
1) jest rozpuszczalnikiem subst. biologicznie czynnych, jest środow. wielu reakcji 2)bierze bezpośr. udział w wielu reakcjach chem. Jako substrat lub produkt. 3)wypełniając wakuolę podtrzymuje turgor całej rośl. 4) jest czynnikiem umożliwiającym szybki wzrost kom. i tkanek 5) bierze udział w przemieszcz. się subst. w organ. roślinnym: zw. miner. i niektórych metabolitów w ksylemie oraz produktów asymilacji i subst. czynnych biolog. we floemie .
CO OZNACZA TERMIN STRES W FIZJOLOGII ROŚLIN.
Stres to stan pewnego „napięcia” w organizmie w którym wskutek działania niesprzyjających czynników środowiska następuje upośledzenie albo ograniczenie metabolizmu lub rozwoju . Czynnikiem stresowym może być : zbyt niska lub wysoka temperatura , brak lub nadmiar wody , nadmierne zasolenie lub brak soli , światła itp. Rośliny wykształciły mechanizmy obronne przed działaniem czynników stresowych. Są to mechanizmy trwałe i wykształcone ( aklimatyzacja)
OD JAKICH CZYNNIKÓW ZALEŻY WRAŻLIWOŚĆ ROŚLIN NA NIEKORZYSTNE WARUNKI ŚRODOWISKA.
Rośliny posiadają trwałe mechanizmy odpornościowe ( budowa liści, kora itp.) i nabyte - pojawiają się w odpowiedzi na czynnik stresowy. Zatem wrażliwość roślin będzie zależała od nabytych cech w toku ewolucji, czasu działania i natężenia czynnika stresowego co w ostateczności może przyczynić się do aklimatyzacji.
-wrażliwości na chłód i mróz
-w błonach roślin odpornych na chłód jest przewaga kwasów tłuszczowych nienasyconych.
-Wyciąganie wody przez ściany komórkowe
-U roślin odpornych na mróz wysokie stężenie soku komórkowego
A)wrażliwość i odporność na wysoką temperaturę
-przewaga w błonach kwasów tłuszczowych nasyconych
-ilość aparatów szparkowych umożliwiających transpirację
A)susza
-wzrasta synteza wosków
-w soku komórkowym związki osmotycznie czynne
A)zasolenie
-odprowadzanie soli do wodniczek
ROŚLINY C3 I C4
Różnice metaboliczne
Rośliny C3
-pierwotny akceptor CO2- RuBP
-wtórny akceptor RuBP- brak
-pierwszy trwały produkt karboksylacji- PGA (3C-fosfoglicerynian)
-zużycie ATP na redukcje 1 cząs. CO2-3
-fotooddychanie- intensywne
-akumulacja skrobi- normalna
-podział funkcji między chloroplastami- brak
Rośliny C4
-pierwotny akceptor CO2- PEP (mezofil)
-wtórny akceptor CO2-RuBP (pochwa)
-pierwszy trwały produkt kondensacji- OAA (HC-szczawiooctan, mezofil)
-zużycie ATP na redukcję 1 cząs. CO2- 4-5
-fotooddychanie- brak lub śladowe
-akumulacja skrobi- wyłącznie w chloroplastach- komórki pochwy okołozalążkowej
-podział funkcji między chloroplastami-istnieje:1. mezofil- cykl Hatcha i Slacka; 2.pochwa okołozalążkowa- wtórna karboksylacja (RuBP), synteza skrobi, cykl Calvina .
Rośliny C4 pochodzą na ogół z klimatów gorących i suchych
-wysokie wymagania natężenia promieniowania
-wyższa optymalna temperatura fotosyntezy
-są bardzo odporne na susze i inne stresy.
Z rodziny C4-trawy-kukurydza, sorgo, proso; z innych-komosowate, amarantowate.
Różnice fizjologiczne:
Rośliny C3
-intensywność fotosyntezy rzeczywistej-przeciętna (15-35%)
-świetlny punkt wysycenia (0,5-3,5%)
-optimum temp. -około 25C
-gradient stężenia CO2 atmosfera/ chloroplast- średni
-punkt kompensacyjny- wysoki (około 50 ppm CO2)
-gospodarka wodna- mało oszczędna
-odprowadzenie asymilatów z liści- średnie
-reakcja fotosyntezy na spadek stężenia CO2( z 21 do 1%)-wzrost intensywności fotosyntezy o około 45%)
Rośliny C4
-intensywność fotosyntezy rzeczywistej- wysoka 40-80
-Świetlny punkt wysycenia- do 7%
-optimum temp. Około 35C
-gradient stężenia CO2 atmosfera/ chloroplast-wysoki
-punkt kompensacyjny-niski (0-10 ppm CO2)
-gospodarka wodna- oszczędna
-odprowadzenie asymilatów z liści- szybkie
-reakcja fotosyntezy na spadek stężenia CO2 (z 21 do 1%) brak reakcji (o około 0-6%)
Fitochrom: barwnik, fotoreceptor odbierający bodziec świetlny. Jego budowa jest podobna do chlorofilu bez magnezu. Jest połączony z białkiem np. plazmalemma. Pochłania światło w zakresie czerwieni bliskiej (b 666) i dalekiej(b730), która jest fizjologicznie aktywna. Działa przez uruchamianie pewnych genów, a nastepnie systemów enzymatycznych. Bez światła (czarnuszka, szarłat).
CO2 na ogół optymalne stężenie około 0,03%; niewielki wzrost stężenia nie wpływa negatywnie na kiełkowanie (nawet lekka stymulacja np. rzepień, sałata, koniczyna), wzrost stężenia CO2 do 15-20% na ogół hamowanie kiełkowania, stężenie 30-35% (rzadko w warunkach naturalnych)-śmierć nasion.
Składniki mineralne: na ogół stężenie przedsiewne w glebie- mały wpływ na kiełkowanie. Azotany: (np. KNO3 przedsiewnie)-stymulacja kiełkowania działanie makro i mikroelementów zleży od składu chemicznego nasion (np. niedobór P w roślinach macierzystych- słabsza zdolność kiełkowania; niedobór K- szybki spadek zdolności kiełkowania), zbyt wysokie stężenie makroelementów w glebie- obniżenie zdolności kiełkowania (zachwiana gospodarka wodna nasion), zbyt wysokie stężenie mikroelementów- działanie toksyczne.
Pęcznienie:
zawartość wody koniecznej dla kiełkowania (wilgotność krytyczna)-I etap (inhibicja): nasiona oleiste i skrobiowe- 20-40%, nasiona białkowe- 50-60%, zarodki nasion oleistych-70%.
Zawartość wody podczas dalszych etapów pęcznienia: II etap: wzrost zawartości wody w stosunku do początkowej suchej masy: groch, fasola- 110%, burak ćwikłowy- 63%
Kiełkowanie: zespół procesów zachodzących w nasieniu, których wynikiem jest aktywacja zarodka prowadząca do wzrostu siewki
Fazy kiełkowania (przemiany metaboliczne):
I Pęcznienie (inhibicja)- faza fizyczna: intensywne pobieranie wody (bielmo, liścienie, wolniej zarodek), *aktywacja zapasowego m-RNA, aktywacja resztkowego M-RNA (forma zapasowa m-RNA), pod wpływem wzrostu uwodnienia o spadku ABA, aktywacja głównego zapasowego m-RNA (pod wpływem zw. wysokoenergetycznych)-trnslacja m-RNA-początek syntezyi aktywacji enzymów hydrolitycznych, brak zmian w zawartości DNA (brak podziałów komórkowych), * gwałtowny wzrost oddychania: początkowo beztlenowego- wzrost aktywn. Enzymów glikolitycznych- wzrost glikolizy- mało ATP, etanol, ograniczona dyfuzja tlenu, nieaktywnych mitochondria; potem tlenowego- wzrost aktyw. Enzymów cyklu Krebsa i fosforylacji oksydacyjnej- dużo ATP, synteza de novo i dojrzewanie mitochondriów, * synteza związków wysokoenergetycznych (AMP, ADP, ATP), *mobilizacja substratów oddechowych: cukry proste (glukoza, fruktoza), początek hydrolizy oligosacharydów (sacharozy, rafinozy, stachiozy), początek hydrolizy polisacharydów ścian komórkowych (celuloza, pektyny).
Szybkośc pęcznienia zależy od: *temp (od 5-50C)-nadmiernie wysoka- hamowanie dostępu tlenu, *zawartości wody w podłożu (dobra pojemność wodna-50%(lub powyżej)ogólnej pojemności wodnej), *zawartość tlenu- konieczny czynnik dla rozpoczęcia pęcznienia (21%- korzystne stężenie), *położenie nasion w podłożu, *uwarunkowana genetycznie: najszybciej pęcznieje groch, fasola, peluszka, mniejsza szybkość pęcznienia np. ziarniaki traw (zbóż), rodziny lnowatych, baldaszkowatych (marchew), niełupki złożonych (słonecznik), komosowatych(buraki), * budowy okryw nasiennych: zaw. Śluzów0 aktywne wiązanie wody (lepsze pęcznienie) np. len, babka lancetowata, półprzepuszczalność lub trudna przepuszczalność dla wody, *zawartości (składu chemicznego) materiałów zapasowych, *wielkości nasion- drobne wchłaniają wodę szybciej niż większe (nawet tego samego gatunku).
II Faza biochemiczna:
1- faza kataboliczna: *transkrypcja m RNA- synteza m RNA (gł. w zarodku), *wzrost aktywności giberelin, *wzrost syntezy i aktywności białek enzymatycznych (np. hydrolaz), brak zmian w zawartości DNA, aktywna hydroliza związków zapasowych 1-tłuszczów zapasowych(lipaz- kw. Tłuszczowe, glicerole- substraty oddechowe i metabolity pośrednie); 2-skrobi (alfa i beta- amylaza- monosacharydy- substraty oddechowe), 3- polisacharydów ścian komórkowych (monosacharydy), 4- białek zapasowych (proteazy= aminokwasy, amidy- substraty oddechowe i metabolity pośrednie np. kw. Glutaminowy i asparaginowy do innych syntez aminokwasów), *wysoka intensywność oddychania tlenowego, *początek transportu pri\ostych zw. organicznych do zarodka.
2- faza anaboliczna: *wzrost biosyntezy m RNA, wzrost biosyntezy białek(gł. strukturalnych), brak zmian w zawart. DNA, *wysoka intensywność odyychania tlenowego (ATP), *wysoka aktywność stymulatorów wzrostu (np. GA), *spadek intensywności hydrolizy zw. zapasowych, *bardzo aktywny transport prostych zw. org. w zarodku (np. nowych białek, węglowodanów, tłuszczów)- aktywacja zarodka, *ujemny bilans energetyczny zarodka często spadek zawartości s. m.).
III faza fizjologiczna (wzrost embrionalny i wydłużeniowy kiełka): *wzrost zawartości DNA w zarodku, *embriogeneza komórek zarodka, *pojawienie się korzonka zarodkowego, wzrost i podziały komórkowe części podliścieniowej (hipokotylu)- kiełkowanie nadziemne- wyniesienie liścieni nad powierzchnię gleby (fasola, rzodkiewka, cebula, sałata, pomidor), *wzrost i podziały komórek części nadliścieniowej (epikotylu)- kiełkowanie podziemne- liścienie pozostają pod ziemią( groch, zboża, dąb, leszczyna, orzech włoski), *wzrost zawartości auksyn (regulacja wzrostu siewki), *początek autotrofizmu siewek (liścieni lub pierwszych liści)- dodatni bilans przemiany materii- wzrost s. m.
Żywotność zależy od gatunku rośliny-jak jest uwarunkowana genetycznie i jakie są warunki przechowywania nasion.
Nasiona roślin motylkowych- gr. o niskiej żywotności(szybko tracą żywotność).
2 x martwych nasion fasoli niż pszenicy. Nasiona zbóż można przechowywać dłużej, nawet po kilku latach nie tracą swej żywotności.
Metoda konduktometryczna- inna metoda oceny żywotności nasion.
Młode-wyższa żywotność. Potraktowanie nasion krótkim szokiem termicznym(uszkodzenie błon kom. i silny wzrost elektroprzewodnictwa-obce grupy nasion), ale większą destrukcję zaobserwowano w przypadku nasion starych.
ODŻYWIANIE MINERALNE ROŚLIN
I grupa-Pierwiastki ruchliwe(N, P, K, Mg)-poruszające się w roślinie, w momencie niedoboru pierw. jest on przemieszczany do najmłodszej rosnącej części rośliny i tam ponownie włączany do struktur(reutylizacja- ponowne wykorzystanie)-pierwsze objawy niedoboru na liściach starszych.
II grupa-Pierwiastki nieruchliwe(Ca, Fe, S)-nie mogą być przemieszczane, tkwią w danej strukturze. Pierwsze objawy niedoboru na liściach młodych, wierzchołkach wzrostu.
Bierne pobieranie pierwiastków na zasadzie wymiany jonowej miedzy glebą a korzeniem. Pobieraniu kationów towarzyszy wydzielanie równoważnej liczby kationów w formie H+ do roztworu
K2SO4
K- chętniej pobierane przez rośliny
H+ -pH - sól fizjologicznie kwaśna (borówka, żurawina, rododendron).
Pobieraniu anionów towarzyszy wydzielanie równoważnej ilości OH- np.
Ca(NO)3, NO3 -chętniej pobierany jest przez rośliny ten anion.
OH—pH - sól fizjologicznie zasadowa (lawenda, tulipan, cebulowe)
Możemy modyfikować pH r-ru w zależności od tego czy chętniej jest pobierany kation czy anion.
Reakcje kilku gatunków roślin na zasolenie
W pożywce zasolonej pomidor i fasola zwiększyły wymiary o bardzo mało( dł., szer. liścia i długość rośliny). Natomiast kukurydza zwiększyła wymiary o trochę więcej- bardziej odporna na zasolenie. Próby kontrolne-normalnie.
Halofity- Salicarnia herbacea, Łoboda(Atriplex), Soliród zielny, muchotrzew solniskowy.
Glikofity- rośliny uprawne:1)stosunkowo odporne: jęczmień, burak cukrowy, soja, pszenica.2)średnio odporne: owies, kukurydza, pomidor, kapusta.3)wrażliwe: sałata, ziemniak, marchew, cebula, ogórek.4): bardzo wrażliwe: rzodkiewka, fasola, drzewa owocowe(jabłoń, czereśnia, brzoskwinia-szczególnie bardzo młode).
Stres solny(nadmierne stężenie soli min. np. NaCl w podłożu)
Efekt stresu
-zaburzenie pobierania wody, (gdy unikanie pobierania NA+ Cl-= deficyt wodny; zbyt niski ΔΨk roztwór glebowy/ kom. korzenia)
-zatrucie jinami Na+ Cl-(gdy zwiekszone pobieranie jonów= toksyczność)
-naruszenie równowagi jonowej
↑kumulacja Na+Cl--dezintegracja membran
↓deficyt K+-blokada kanałów jonowych
↓gwałtowne uwalnianie Ca z membran (plazmalemma, tonoplast,, ER, PSII)-zaburzenia ruchu ap. Szparkowych, inaktywacja ATP-azy,
↓pobieranie azotanów i fosforanów (spadek ich stężenia)
-↓aktywności enzymów regulowanych przez potas (synteza białek, kw. Nukl.)
-↓asymilacji CO2-zamykanie szparek, ↓chlorofilu, ↓ aktywności karboksylazy RuBP, inaktywacja PSII, ↓akt. ATP-azy w chloroplastach
-↑(na ogół) oddychania, większe zapotrzebowanie na energię,
-zmiany w transporcie i dystrybucji asymilatów (akumulacja asymilatów w liściach, zahamowany eksport),
-zahamowanie wzrostu (szczególnie najmłodszych części roślin).
Stres solny - Mechanizmy odporności
Tolerancja na stresy:
Glikofity- wrażliwe,
Halofity- odporne.
1)-obniżenie pobierania jonów
2)-tolerancja na akumulację jonów(akumulacja Na w liściach; akum. K w pędach; wydzielanie nadmiaru przez korzenie; akum. w wakuoli; gruczoły wydzielnicze)
Mechanizmy adaptacyjne:
1)adaptacje osmotyczne(utrzymanie równowagi osmot. wakuola / cytoplazma)synteza osmoprotektantów= ochrona białek enzymatycznych przed inaktywacja. ↑
stęż.(w cytozolu): prolina, sacharoza, glicerol, betaina, inozytol, poliaminy
2)indukcja specyficznego białka OSMOTYNA(ABA---mRNA)
3)zmiany w zawartości hormonów ↑ABA, ↓(na ogół)cytokininy, auksyny, gibereliny(na ogół)-zmiany zależą od fazy reakcji roślin na stres.
4)zmiany w składzie lipidów w membranach(utrzymanie ich struktury) ↑lipidów zawierających kw. linolowy, synteza nowych lipidów, ↑fosfatydylocholiny, sulfo- i glikolipidów.
Stres wodny
Deficyt wody-stres suszy
Błony komórkowe:
-odwodnienie komórek (dehydratacja)
-zmiany w strukturze białek lipidów= denaturacja białek
-↑przepuszczalności błon
-↓aktywności enzymów np. karboksylazy RuBP
Fizjologia komórki (rośliny)
-↓transpiracji ↑ALA-zamykanie aparatów szparkowych
-↓fotosyntezy (↓asymilacja CO2, ↓przewodności szparkowej, ↓aktywności RuBP, ↓zawartości chlorofilu, karotenoidów, i białek w chloroplastach=↓syntezy cukrów)
-niewielkie zmiany intensywności oddychania mitochondrialnego
-↑fotooddychania,
-zmiany metaboliczne: zahamowanie syntezy i stymulacja rozkładu skrobi, ↑syntezy sacharozy
-spadek turgoru= więdnięcie roślin
-ograniczenie wzrostu części nadziemnej rośliny („ zagłodzenie roślin” ↓ syntezy asymilatów, ↓DNA)
-rozbudowa systemu korzeniowego
Reakcja na stres suszy (mechanizmy odporności)
Osmoregulacja komórki:
-↑stężenia soku komórkowego: ↑niskocząsteczkowych subs. Rozpuszczalnych w wodzie (cukry i ich pochodne)
↑aminokwasy, kwasy organiczne
↑sorbitol
↑K+
↑prolina, osmotyna
↑specyficzne białka stresowe (stymulacja przez ABA)
↑inne związki czynne biologicznie (zw. fenolowe: kumaryna, fitoaleksyny)
-zmiany w zawartości regulatorów wzrostu
↓cytokininy ↓gibereliny
↑ABA (zamykanie szparek, zmiany w przepuszczalności membran=zwiekszenie transportu K+ ze szparek do kmórek otaczających)
Rośliny odporne
Kserofity(małe liście, kolce, włoski, ↑systemu korzeniowego)=tolerancja na stres
Sukulenty (duża zawartość wody związanej)= unikanie stresu
Stres- wartość czynnika środowiska, która wywołuje niekorzystny nawet letalny wpływ na roślinę.
Stresor- każdy czynnik środowiska, który ogranicza procesy życiowe tzn. obniża ich intensywność poniżej potencjalnych- uwarunkowanych genetycznie możliwości.
Stresory:
Fizyczne: woda: deficyt, nadmiar; temp.: chłód, mróz; promien.: daleka czerwień, ultrafiolet, promien. Widzialne; inne: pole magn., pole elektr.
Chemiczne: pierwiastki miner.: deficyt, nadmiar; zanieczyszczenia: atmosf., glebowe; pestycydy; środki ochrony
Biotyczne: konkurencja: wewnątrz i zewnątrz gatunkowa; allelopatia- oddziaływanie różnych gatunków; porażenie przez patogeny, owady.
Skutki stresu: dehydratacja komórek-plazmoliza, nieodwracalna zmiana konfiguracji przestrzennej białek (utlenianie grup tiolowych-SH, tworzenie wiązań s-s, trwała deformacja struktury białek enzymatycznych).
Reakcja na stres- ogólna mobilizacja organizmu w celu przeciwdziałania skutkom czynnika stresowego.
Strategia przetrwania- mechanizmy odpornościowe
-unikanie stresów- wytworzenie chemicznych i fizycznych barier zmniejszających prawdopodobieństwo uszkodzeń komórek.
-tolerancja skutków stresów- minimalizacja negatywnych skutków stresu.
Unikanie stresu: adaptacje ewolucyjne: przystosowania morfologiczne:1przystosowania rozwojowe(np. efemerydy, tkanki spoczynkowe) 2.adaptacje fizjologiczne
Tolerancja: 1.unikanie odkształceń (np. denaturacji białek)-zapobieganie zmianom(nieodwracalnym i odwracalnym), 2. tolerancja odkształceń: regeneracja uszkodzeń, uruchomienie specyficznych procesów metabolicznych.
Mechanizmy zwiększające odporność roślin na niekorzystne czynniki środowiska (przekazywane dziedzicznie potomstwu)-ADAPTACJE
-zmiany strukturalne
-zmiany procesów fizjologicznych
-zmiany procesów biochemicznych
Zabezpieczenie struktur komórkowych przed deformacją.
Stres termiczny-temp. Ujemna.
Objawy stresu:
- zamarzanie (krystalizacja) wody w komórce i przestworach komórkowych. Skutki: obniżenie potencjału osmotycznego w przestrzeniach komórkowych. Osmotyczny transport wody do przestrzeni- odwodnienie komórki.
-zmiany fazy lipidów i zawartości białek w błonach komórkowych
-denaturacja i dysocjacja białek-zmiany w przepuszczalności błon.
-zahamowanie fotosyntezy
-wzrost intensywności oddychania.
Mechanizm odporności na zamarzanie:
-zabezpieczenie komórek przed krystalizacją wody w jej wnętrzu- minimalizacja ośrodków krystalizacji lodu.
Mechanizmy adaptacyjne (hartowanie-wzrost odporności)
-wzrost przepuszczalności błon dla wody,
-wzrost zawartości wody związanej (koloidy hydrofobowe), obniżenie temp. Zamarzania,
-zmiana zawartości cukrowców hydroliza skrobi=podwyzszenie zawartości cukrów prostych=podwyż zawart. Sacharozy= podwyż. Potencjału osmotycznego
-hydroliza białek= podwyż. Zawartości wolnych aminokwasów (zabezpieczenie grup tiolowych i białek przed tworzeniem mostków S-S
-podwyż zawartości kompleksów cukry+ białka (zabezpieczenie przed koagulacją)
-zmiany zawartości regulatorów wzrostu- podwyż. ABA, obniżenie auksyn, giberelin, cytokinin.
Stres suszy: spadek turgoru- więdnięcie roślin, zwiększenie substancji osmotycznie czynnych (proste cukry, sorbitol)- aby zahamować stres suszy.
Wśród naszych roślin uprawnych stosunkowo odporne są zboża. Najwrażliwsza na zasolenie jest fasola. Pomidor- średnio odporny, najbardziej odporny jęczmień, burak cukrowy, kukurydza.
TRANSPIRACJA
Parowanie wody przez żywą roślinę w warunkach niedosytu wilgotności w powietrzu.
Prężność pary wody w przestworach komórkowych jest większy niż prężność pary wody w atmosferze.
Rodzaje transpiracji: -kutikularna (5-20% transpiracji ogólnej), - szparkowa i przetchlinkowa (ponad 80% transpiracji ogólnej).
Znaczenie: *uruchomienie głównego (biernego) mechanizmu pobierania i transportu wody, *ułatwienie pobierania składników mineralnych z gleby, *ochrona przed przegrzaniem rośliny, *zbyt wysoka intensywność-więdnięcie roślin.
Susza fizjologiczna: *zbyt niska temperatura, *niedobór (brak) tlenu w glebie, *zasolenie w glebie.
Intensywność transpiracji - ilość wytranspirowanej wody / jednostkę powierzchni (masy) liścia/ jednostkę czasu.
Jednostki transpiracji:
-Intensywność transpiracji: [mol H2O*m-2*s-1], [mg H2O*dm-2*h-1]
-Współczynnik transpiracji: [mol H2*mol-1 CO2], [gH2O*g-1 s.m.]
WUE- współczynnik wykorzystania wody (produktywność transpiracji): [mol CO2*mol-1 H2O], [g s.m.*kg-1H2O]
-transpiracja względna: T/E gdzie E- intensywność parowania, T- Intensywność transpiracji
Współczynnik transpiracji względnej- stosunek wody (ilości) wytranspirowanej przez jednostkę powierzchni liścia do ilości wody wyparowanej przez jednostkę wolnej powierzchni cieczy (0,01-0,80- wysoka sprawność dyfuzyjna aparatów szparkowych).
Prawo Stephana (intensywność dyfuzji gazu przez małe otworki jest proporcjonalna do ich średnicy a nie powierzchni.
Efekt brzeżny: zaginanie torów dyfuzyjnych pary wodnej na krawędzi otworków co usprawnia odprowadzanie pary wodnej znad powierzchni liścia.
Czynniki wpływające na intensywność transpiracji: *ilość wody w glebie, *dostępność wody dla roślin, *temperatura gleby, *światło, *temperatura, *niedosyt wilgotności powietrza, *wiatr, *stężenie CO2, *cechy anatomiczno- morfologiczne, *aparaty szparkowe: ilość, rozmieszczenie, budowa, zdolność otwierania się, *kutikula, *kutner, *redukcja blaszek liściowych, *zwijanie blaszek.
sścięciu.
FAZA ŚWIETLNA- cząsteczki chlorofilu po zaabsorbowaniu kwantów światła przechodzą w stan wzbudzenia, co powoduje wybicie z nich elektronów. Wybite elektrony mają zapas energii wskutek pochłonięcia kwantów światła. Elektrony te są wyłapywane i transportowane przez przenośniki elektronowe, które tworzą łańcuch przenoszenia elektronów, wbudowany w błony gran. Podczas przepływu przez kolejne ogniwa łańcucha elektrony tracą swą energię, która jest przechwytywana i wiązana chemicznie w postaci fosforanowych wiązań ATP. Jeżeli wybity z chlorofilu elektron wraca przez system przenośników na cząsteczkę chlorofilu, z którego został wybity, to - przechodząc na swój wyjściowy poziom energetyczny- oddaje energię, która jest akumulowana w ATP. Jest to fotofosforylacja cykliczna, bowiem towarzyszy ona cyklicznemu transportowi elektronów. Gdy wybite z chlorofilu elektrony zostaną przetransportowane na przenośnik wodorowy NADP, to do wytworzonej „ dziury” w cząsteczce chlorofilu są przekazywane elektrony wybite z innej cząsteczki chlorofilu, a ta z kolei wysysa elektrony z wody, powodując jej fotodysocjację. Zmiana poziomu energetycznego przenoszonych elektronów umożliwia zakumulowanie energii w ATP. Jest to fosforylacja fotosyntetyczna niecykliczna, ponieważ zachodzi proces niecyklicznego transportu elektronów. W przenoszeniu elektronów uczestniczą dwa systemy barwników, wodór z wody zostaje przyłączony do NADP, a tlen jest uwalniany do atm. Obie drogi foftosyntetycznego transportu elektronów działają jednocześnie, a ich efektem jest wytworzenie siły asymilacyjnej: ATP i NADPH2. U roślin wyższych wykryto dwa fotosystemy. Fotosystem I (PSI) zawiera gł. chlorofil a i karotenoidy, a jego centrum aktywne wykazuje max absorpcji światła o długości 700nm. Fotosystem II (PS2) zawiera chlorofil a,b i ksantofil, a jego max absorpcji światła wynosi 680nm.
CZYNNIKI WPŁYWAJĄCE NA POBIERANIE WODY PRZEZ KORZENIE
Czynniki środowiskowe wpływają :
-pośrednio; czynniki zmieniające transpirację- część, działania liścia, wzrost korzeni.
-Bezpośrednio czynniki glebowe- zawartość wody w glebie, stężenie roztworu glebowego, areacja, temperatura.
ZAWARTOŚĆ WODY W GLEBIE; postacie wody w glebie: higroskopowa, błonkowata ( niedostępna dla roślin) , kapilarna grawitacyjna ( dostępna dla roślin). Rośliny czerpią wodę z gleby gdy jest ona w stanie polowej pojemności wodnej( max. Ilość wody jaką gleba może utrzymać po wpłynięciu nadmiaru wody wskutek sił ciążenia wyrażona w procentach suchej masy i pełnej pojemności wodnej ( dochodzi woda grawitacyjna. Zawartość wody w glebie gdy rośliny zaczynają w niej więdnąć wyrażoną w procentach suchej masy gleby nazywamy wilgotnością stałego więdnięcia. Temperatura niska silnie zmniejsza pobranie wody , zwiększa się lepkość cytoplazmy i zmniejsza się jej przepuszczalność dla wody, zmniejsza się wzrost korzenia i jego aktywność metaboliczna, zwiększa się lepkość wody i zmniejsza się natężenie procesów dyfuzji. Areacja gleby- w glebach dobrze natlenionych jest szybsze pobieranie wody niż w glebach o niskiej zawartości tlenu- zmniejsza się oddychanie korzeni, przepuszczalność cytoplazmy i zachwianie wzrostu korzeni. Stężenie roztworu glebowego - wzrost stężenia roztworu glebowego może być wywołany nadmierną ilością nawozów sztucznych lub zraszaniem wodą bogatą w składniki mineralne. Może to spowodować suszę fizjologiczną- brak możliwości pobierania wody mimo jej dostatku w środowisku, a nawet może być przyczyną oddawania wody przez roślinę do roztworu glebowego - zgodnie z gradientem stężeń.
ROLA TRANSPIRACJI W TRANSPORCIE WODY
Mechanizm pobierania i przewodzenia wody jest inny w przypadku silnej transpiracji a inny u roślin , które transpirują słabo. Podczas silnej transpiracji działają : mechanizm pasywny, który jest umiejscowiony w liściach. Natomiast w przypadku słabej transpiracji widoczny staje się mechanizm aktywny, umiejscowiony w korzeniu i przejawia się jako tzw. ciśnienie korzeniowe.
MECHANIZM PASYWNY: Jest wynikiem ssącego działania transpiracji . W procesie transpiracji woda wyparowuje ze ścian mezofilu, a wysychające ściany czerpią wodę z wnętrza komórek wskutek działania sił inhibicyjnych .Obniża się wtedy ciśnienie turgorowe (cT) wzrasta natomiast potencjał osmotyczny (Po) wskutek jej odparowania. Obniżenie turgoru i wzrost potencjału osmotycznego prowadzi do wzrostu siły ssącej komórek (S=Po-cT) . Wzrost siły ssącej komórek powoduje przenikanie do nich wody z sąsiednich komórek, otrzymujących z kolei wodę z cewek i naczyń w nerwach liścia . Ubytek wody w naczyniach wywołuje w nich podciśnienia hydrostatyczne ( sięga przez łodygę do korzeni).
W poszczególnych elementach naczyniowych woda tworzy słupy sięgające od cewek w liściu do korzeni . Ciągłość słupów wodnych jest warunkiem niezbędnym pobierania jak i przewodzenia pasywnego. Każdy słup wody podnoszony jest w górę za pomocą sił ssących wywołanych dużą transpiracją. Ruch wody odbywa się na drodze najmniejszego oporu poprzez ściany komórkowe albo przez cytoplazmę i plazmodesmy lub od wakuoli do wakuoli na zasadzie osmozy. Korzenie w tym procesie odgrywają jedynie bierną rolę tworząc system rurociągów. Roślina nie zużywa więc własnej energii do pobierania wody , lecz wykorzystuje różnicę pomiędzy siłą ssącą gleby i atmosfery.
TEORIA KOHEZYJNA. Podnoszenie się słupa wody jest uwarunkowane napięciem w górnej części słupa wytworzonym przez transpirację i spójnością cząstek wody.
MECHANIZM AKTYWNY . Tkwi w korzeniu i uwidacznia się w przypadku słabej transpiracji, wymaga dopływu energii. W wyniku tego procesu pojawia się ciśnienie korzeniowe, którego widocznymi objawami są guttacja i płacz roślin.
Istota tego procesu:- wg. hipotezy osmotycznej ( woda transportowana jest na zasadzie osmozy) - wg . hipotezy nieosmotycznej ( komórki miękiszu wydzielają wodę do naczyń czynnie lecz z wykorzystaniem energii uwalnianej w procesie oddychania)
ZNACZENIE TRANSPIRACJI
Dzięki transpiracji następuje obniżenie temperatury rośliny , co zapobiega przegrzaniu. Transpiracja warunkuje jednokierunkowy przepływ wody i rozpuszczonych w niej substancji mineralnych, a w pewnych przypadkach również transport związków organicznych ( alkaloidów produkowanych w korzeniach, cukrów transportowanych z korzenia do wierzchołków pędów). Występowanie zjawiska transpiracji jest nieuniknione gdyż liście mogą spełniać swe funkcje jedynie przy otwartych szparkach przez które musi wnikać dwutlenek węgla. Nadmierna jest niepożądana gdyż przez otwarte szparki zachodzi utrata wody , której niedostatek hamuje wzrost i rozwój roślin, a w skrajnych przypadkach może doprowadzić do śmierci.
CZYNNIKI WEWNĘTRZNE FOTOSYNTEZY.
ZAWARTOŚĆ CHLOROFILU- wahania zawartości chlorofilu mają niewielki wpływ na fotosyntezę gdyż nie cały chlorofil bierze aktualny udział w tym procesie. Zahamowanie syntezy chlorofilu powoduje zahamowanie procesu fotosyntezy.
ANATOMIA LIŚCIA- od wielkości i rozmieszczenia aparatów szparkowych, od wielkości przestworów międzykomórkowych ( szybkość wnikania CO2) od grubości kutykuli, od budowy miękiszu gąbczastego i palisadowego ( ilość światłą jaka przenika do liścia). ROZMIESZCZENIE CHLOROPLASTÓW- wpływa na natężenie fotosyntezy jedynie przy niezbyt intensywnym świetle ( wtedy gdy światło jest czynnikiem ograniczającym). Chloroplasty zmieniają swe położenie w komórce pod wpływem zmian oświetlenia. EPISTROFIA chloroplasty ustawione są wzdłuż górnej powierzchni komórki ( przy niskich intensywnościach światła) PARASTROFIA -chloroplasty przemieszczają się na boczne ściany komórki i w ten sposób się zasłaniają ( przy wyższych intensywnościach światła)
GUTACJA
Jest to wydzielanie przez liście wody w stanie ciekłym ( przy transpiracji wydalana jest para wodna). Odbywa to się przez szparki wodne, wypotniki czyli hydatody. Wydzielana ciecz oprócz wody zawiera ślady cukrów, soli mineralnych, aminokwasów. Gutacja występuje wtedy kiedy istnieją warunki korzystne dla pobierania wody przez korzenie a niekorzystne dla transpiracji i występuje to w naszym klimacie późnym latem przy bardzo dużej wilgotności powietrza ( niedosyt wilgotności powietrza jest bliski zera) . Zwykle wieczorem lub wczesnym rankiem i gdy po ciepłym dniu następuje chłodna noc. Nad ranem można zaobserwować na brzegach liści kropelki wody i są one często brane za rosę. GUTACJA BIERNA- wykraplany płyn nie jest czynną wodą lecz jest mniej lub bardziej wykroplonym sokiem komórkowym . GUTACJA CZYNNA- przyczyną gutacji jest dodatnie ciśnienie panujące ciśnienie panujące w naczyniach zwane ciśnieniem korzeniowym. Wywołane jest ono aktywnym pobieraniem wody przez korzenie
ODPORNOŚĆ ROŚLIN NA NISKIE TEMPERATURY.
Mrozoodporoność - jest to odporność na niskie temperatury mogące spowodować śmierć mrozową komórek przez wykrystalizowanie wody w przestworach międzykomórkowych co powoduje odwodnienie i koagulację cytoplazmy.
Mrozoodporność polega na zwiększaniu stężenia osmotycznego soku komórkowego i obniżenia punktu zamarzania, głównie drogą przemiany skrobi w cukier częściowym odwodnieniu i zagęszczeniu cytoplazmy , dzięki czemu pozostała woda związana jest znacznymi siłami z koloidami plazmatycznymi.
Zimotrwałość w obniżonej temperaturze jest utrudnione pobieranie wody i wysychanie , ochroną przed tym jest zrzucanie liści na zimę ( u cypryśnika zrzucanie całych gałęzi) Zamarzanie wody i powstawanie lodu nie zachodzi w nasionach gdyż są one wysuszone. Skutki wytworzenia lodu zależą od miejsca gdy w protoplaście następuje zniszczenie organelli i śmierć komórki) . Gdy ochładzanie jest bardzo szybkie nie dochodzi do trwałego ochłodzenia. Lód w przestworach międzykomórkowych prowadzi do odwodnienia komórki.
Teoria napięć mechanicznych ( z lat 30 tych) - odwodnienie poszczególnych elementów protoplastu zachodzi z niejednakową prędkością co może spowodować duże różnice naprężeń.
Odwodnienie komórek w trakcie przemrożenia prowadzi do zniszczenia struktury oraz denaturacji białek - zbliżenie łańcuchów polipeptydowych i powstanie mostków dwusiarczkowych . Obie teorie nie uzyskały akceptacji. W czasie łagodnych i śnieżnych zim wysoka temperatura może sprzyjać intensywności oddychania.
Wymakanie roślin . Woda z topniejącego śniegu nie może przenikać głębiej - stwarza warunki anaerobowe , może to powodować uszkadzanie korzeni , tkanek w czasie rozrywania gleby i korzeni.
Wysuszające działanie wiatru
Oblodzenie
FOTOPERIODYZM jest reakcją roślin na czas trwania i periodyczne następstwo okresów światła i ciemności w cyklach dobowych. Zjawisko to opisali Garner i Allard w 1920 roku. Długość dnia i nocy zmienia się na całej kuli ziemskiej sezonowo (z wyłączeniem strefy równikowej ) dlatego też rośliny przystosowały się w znacznym stopniu do następstw występowania pór roku. Rośliny kwitną zależnie od faz jasnej i ciemnej w 24 godzinnym cyklu dobowym. Wyróżniamy następujące grupy fotoperiodyczne:
rośliny dnia krótkiego SDP kwitnące gdy w dziennym fotoperiodzie jest przewaga fazy ciemnej ( powyżej 12 godzin ciemności).
Rośliny dnia długiego LDP kwitnące gdy w dziennym fotoperiodyźmnie jest przewaga fazy jasnej ( powyżej 12 godzin jasnośći)
Rośliny dnia krótkiego- długiego. SLDP i długiego - krótkiego LSDP . Ich inicjacja zależy od ekspozycji na dzień krótki i długi w odpowiedniej kolejności
Rośliny neutralne - DNP . Niewrażliwe na długość dnia. W przypadku tej grupy roślin sygnałem inicjacji kwitnienia jest osiągnięcie przez roślinę odpowiedniego wieku bądź wielkości.
Reakcje fotoperiodyczne:
ilościowa reakcja fotoperiodyczna tzw. wrażliwość względna. Ekspozycja na indywidualny fotoperiod tylko przyspiesza lub hamuje kwitnienie.
Jakościowa reakcja fotoperiodyczna -wrażliwość bezwględna- ekspozycja na odpowiedni fotoperiod bezwzględnie warunkuje indukcję fotoperiodyczną . Niektóre rośliny wymagają tylko jednego indukcyjnego fotoperiodu. Inne wymagają kilku stosownych fotoperiodów.
OSMOREGULACJA
Gdy w wyniku działania różnych czynników stresowych komórki zaczynają tracić wodę i maleje turgor, u roślin odpornych na ten stres włączają się mechanizmy osmoregulacji, które umożliwiają regulację potencjału osmotycznego. Zjawisko to, zwane dostosowaniem osmotycznym polega na przyspieszonej syntezie i akumulacji w wakuoli osmolitów: pewnych aminokwasów i 4-rzędowych zw. amoniowych i innych związków organicznych. Procesów tych nie można traktować jako zakłóceń metabolicznych wywołanych stresem, gdyż wiadomo, że dostosowanie osmotyczne odgrywa kluczową rolę w aklimatyzcji roślin do suszy i zasolenia. W tym ostatnim przypadku w wakuoli akumulowane są również jony nieorganiczne- obniża to potencjał wody w komórce. Dzięki dostosowaniu osmotycznemu komórki mezofilu i szparkowe tracą turgor co pozwala utrzymać rozwarcie szparek- czyli jest więcej czasu na asymilację CO2.
FOTOMORFOGENEZA
Jest to niezależny od fotosyntezy wpływ światła na wzrost, rozwój i różnicowanie się komórek roślin ze względu na to, że światło jest nieodzownym czynnikiem towarzyszącym roślinie w takcie ontogenezy. Procesy te zachodzą również w okresie młodocianym. Światło wpływa na sposób funkcjonowania genów oraz metabolizmu rośliny. Fotomorfogeneza polega na uruchamianiu lub hamowaniu przez światło specyficznych genów, kontrolujących metabolizm. Wpływ światła jest niezwykle istotny w przypadku młodych siewek. Siewka rosnąca w ciemności ma charakterystyczny rośliny etiolowanej- szybki wzrost wydłużeniowy oraz niedorozwój liści. Gdy naświetlimy roślinę etiolowaną rozpoczyna się proces deetiolacji- gwałtowne zahamowanie wzrostu wydłużeniowego łodygi, stopniowy rozwój liści. Głównym fotoreceptorem światła w tym procesie jest fitochrom.
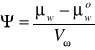
Wyszukiwarka