
N
EUROPHYSIOLOGIC
E
VALUATION OF
E
ARLY
C
OGNITIVE
D
EVELOPMENT IN
H
IGH
-R
ISK
I
NFANTS AND
T
ODDLERS
Raye-Ann deRegnier*
Department of Pediatrics, Northwestern University Feinberg School of Medicine, Prentice Women’s Hospital, Chicago, Illinois
New knowledge of the perceptual, discriminative, and memory ca-
pabilities of very young infants has opened the door to further evaluation of
these abilities in infants who have risk factors for cognitive impairments. A
neurophysiologic technique that has been very useful in this regard is the
recording of event-related potentials (ERPs). The event-related potential
(ERP) technique is widely used by cognitive neuroscientists to study cogni-
tive abilities such as discrimination, attention, and memory. This method
has many attractive attributes for use in infants and children as it is relatively
inexpensive, does not require sedation, has excellent temporal resolution,
and can be used to evaluate early cognitive development in preverbal in-
fants with limited behavioral repertories. In healthy infants and children,
ERPs have been used to gain a further understanding of early cognitive
development and the effect of experience on brain function. Recently, ERPs
have been used to elucidate atypical memory development in infants of
diabetic mothers, difficulties with perception and discrimination of speech
sounds in infants at risk for dyslexia, and multiple areas of cognitive differ-
ences in extremely premature infants. Atypical findings seen in high-risk
infants have correlated with later cognitive outcomes, but the sensitivity
and specificity of the technique has not been studied, and thus evaluation
of individual infants is not possible at this time. With further research, this
technique may be very useful in identifying children with cognitive deficits
during infancy. Because even young infants can be examined with ERPs, this
technique is likely to be helpful in the development of focused early inter-
vention programs used to improve cognitive function in high-risk infants
and toddlers.
©
2005 Wiley-Liss, Inc.
MRDD Research Reviews 2005;11:317–324.
Key Words: neurophysiologic assessment; cognition; infants; high risk
M
any infants and children are at increased risk for cog-
nitive impairments due to genetic or hereditary con-
ditions, perinatal illnesses, brain injuries, and brain
malformations. Typically, impaired cognitive function is diag-
nosed at preschool or school age, long after the presumed
pathogenic process has occurred. Earlier diagnoses of specific
cognitive abnormalities could enable the development of fo-
cused intervention programs to improve outcomes. This future
goal requires a solid understanding of early cognitive develop-
ment in infancy and early childhood.
Early theories of infant development focused primarily on
sensorimotor development [Ginsberg and Opper, 1979], and
infants were not considered to show evidence of true cognitive
function until the emergence of object permanence and lan-
guage abilities during the toddler years. In more recent times,
sophisticated neuroimaging, behavioral, and neurophysiologic
studies have revealed that, although cognitive processes in very
young infants are rudimentary, there is evidence of memory and
early language processing in healthy infants from the time of
birth [DeCasper and Fifer, 1980; DeCasper and Spence, 1986;
Rose et al., 1988; Fagan, 1990; Geva et al., 1999; deRegnier et
al., 2000]. Application of these techniques to the study of
high-risk infants has revealed infant origins of differences in
cognitive processing that were formerly thought to surface in
childhood. This article will review the use of one technique,
event-related potentials (ERPs), in the study of early cognitive
development in high-risk infants and toddlers.
TECHNICAL ASPECTS OF ERPS
ERPs have been used for many years in adults to evaluate
cognitive processes such as attention, memory, and language
[Coles and Rugg, 1995; Naatanen, 1995]. The technique nat-
urally lends itself to the study of developing cognitive processes
in infants as it is noninvasive, relatively inexpensive, and can be
performed in unsedated, preverbal infants with limited behav-
ioral responses. Although it is not yet possible to determine the
precise neural structures that generate each of the ERP compo-
nents, the temporal resolution of ERPs is superior to all other
currently available neuroimaging techniques [Nelson and
Monk, 2001; DeBoer et al., 2005a]. Functional information
gained through use of the ERP can be complementary to
anatomical information gained through current neuroimaging
techniques such as magnetic resonance imaging (MRI). Because
of these characteristics, ERPs have been used in a growing
number of studies of high-risk and healthy infants and toddlers.
In an ERP, brain activity is recorded from the scalp while
the participant is repeatedly presented with stimuli that vary
along perceptual or cognitive dimensions, such as memory.
Brain activity recorded in response to the stimuli is thought to
arise from postsynaptic potentials of pools of cortical neurons
that are physically oriented to yield dipolar fields that are volume
conducted to the surface of the scalp. [Nelson, 1994; Coles and
*Correspondence to: Raye-Ann deRegnier, MD, Department of Pediatrics,
Northwestern University Feinberg School of Medicine, Prentice Women’s Hos-
pital, Room 404B, 333 East Superior Street, Chicago, IL 60611. E-mail:
r-deregnier@northwestern.edu
Received 19 August 2005; Accepted 29 August 2005
Published online in Wiley InterScience (www.interscience.wiley.com).
DOI: 10.1002/mrdd.20085
MENTAL RETARDATION AND DEVELOPMENTAL DISABILITIES
RESEARCH REVIEWS 11: 317–324 (2005)
©
2005 Wiley-Liss, Inc.
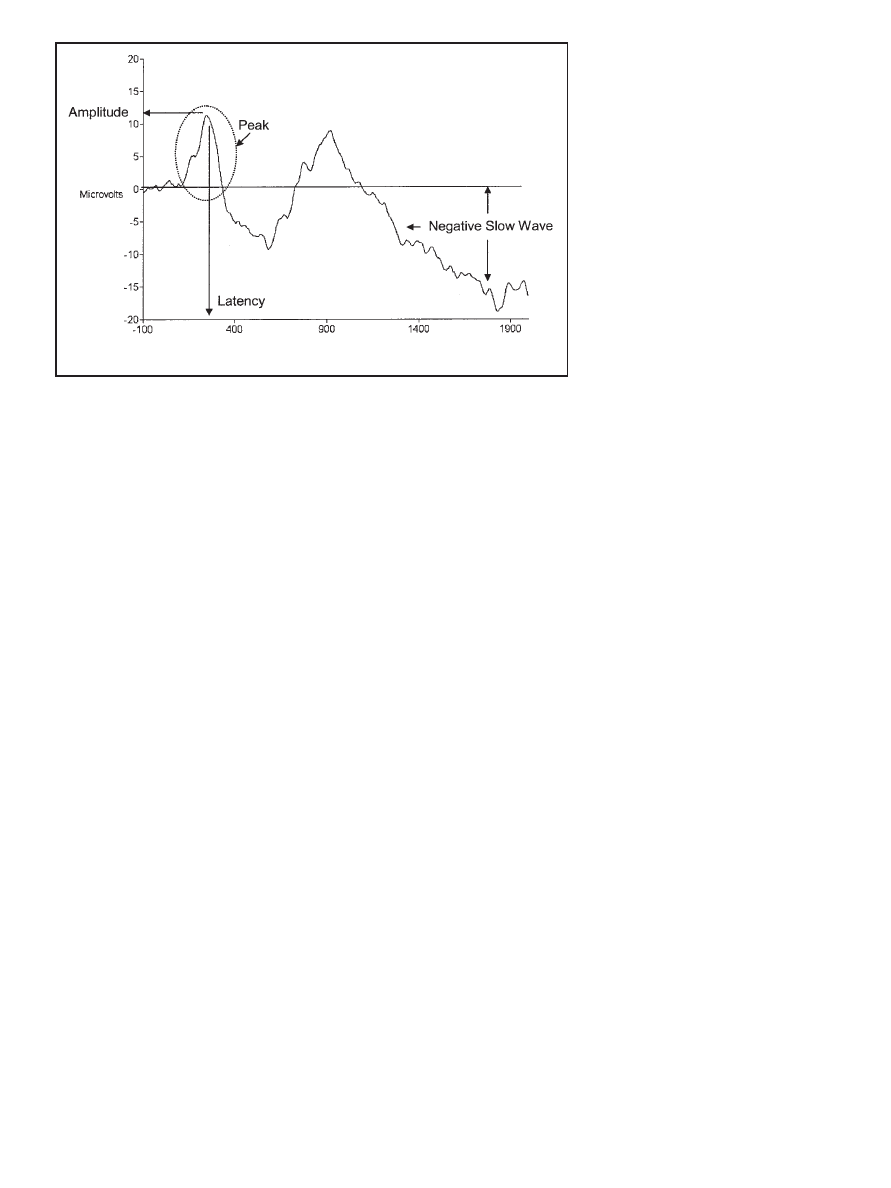
Rugg, 1995]. The raw recordings consist
of the total brain activity including activ-
ity that is directly related to the cognitive
process under investigation plus simulta-
neous unrelated brain activity. Brain ac-
tivity that is not related to the cognitive
process being studied is assumed to be
random and, when the electroencepha-
logram is averaged over many trials, this
random activity will average to zero and
the activity related to the cognitive pro-
cess being studied will emerge as a series
of components or slow wave activity
[Regan, 1989], Figure 1. Components
are typically referred to as peaks that are
measured in terms of the peak amplitudes
(the
maximal
voltage
measurement
above the baseline) and latencies (the
time from the onset of the stimulus to the
point of maximal deflection). Slow waves
are slow drifts in the voltage recordings
of the ERP that are seen with sustained
mental processing. Typically, there is no
discrete peak to measure and slow waves
are assessed by integrating the area under
the voltage–time curve or by computing
the average amplitude across specified
time units [Nelson, 1994].
The most common use of ERPs in
infants and toddlers is to evaluate the
development of specific cognitive func-
tions, such as auditory discrimination
[Cheour et al., 1998, 2002] or memory
[Nelson and Collins, 1991; de Haan and
Nelson, 1997; deRegnier et al., 2000].
This does not require localization of the
neural generators of the ERP activity.
Furthermore, it should be recognized
that neural activity recorded at the scalp
surface represents only that portion of the
total brain activity that has a dipolar ori-
entation and can be recorded at the
scalp’s surface. Significant brain activity
may occur in response to the testing but
may not be recorded at the surface of the
scalp. This means that failure to find ev-
idence of cognitive differences between
stimuli in an ERP study is not decisive
evidence of lack of function.
General techniques and methods
for using ERPs in adults and children
have been reviewed extensively by sev-
eral authors [Regan, 1989; Coles and
Rugg, 1995; Nelson and Monk, 2001;
DeBoer et al., 2005a, 2005b] and readers
are referred to these sources for addi-
tional background information. There
are however, several specific consider-
ations relevant to the study of infants.
It should be noted that infant ERPs
tend to show more variable responses
than older children and adults [Kushne-
renko et al., 2002a, 2002b; Leppanen et
al., 2004]. The reasons for this variability
are not completely understood but likely
are due to a combination of individual
differences in brain maturation, state, or
attentional differences and to the fact that
it is not possible to directly instruct in-
fants on the task to be performed in the
study.
Individual differences in matura-
tion are to be expected in infants and
young children due to ongoing develop-
ment of the brain. General developmen-
tal changes seen in ERPs are the emer-
gence
of
components,
increases
in
amplitudes, and shortening of latencies
[Weitzman and Graziani, 1968; Novak et
al., 1989; Kushnerenko et al., 2002a].
These changes correlate with synapto-
genesis and myelination of the brain
[Eggermont, 1988], and a number of
ERP studies have shown developmental
effects that will be discussed below. Un-
fortunately, there are still relatively few
true developmental ERP studies in
which development of attention, mem-
ory, or language discrimination is studied
at several different ages and the range of
normal has not been well defined for
scalp distribution, amplitudes, and laten-
cies of specific ERP components. These
studies are needed to understand normal
developmental processes and individual
differences.
Understanding
individual
differences in normally developing chil-
dren is vitally important in interpreting
differences that are seen in studies of low-
and high-risk groups of infants.
Due to the ease of recording and
low rate of artifacts, many ERP studies
have evaluated very young infants during
sleep. However, differences in state, at-
tention, or arousal are known to affect
cognition in infants and must be con-
trolled in ERP studies as much as possi-
ble. For very young infants, sleep state
has been shown to have both qualitative
and quantitative effects upon ERPs. For
example, auditory cortical potentials re-
corded during quiet sleep in neonates
tend to be of larger amplitude than po-
tentials recorded during active sleep or
wakefulness (states that are similar to each
other) [Novak et al., 1989; Duclaux et
al., 1991]. Duclaux et al. [1991] also ob-
served that components recorded during
active sleep appeared to have shorter la-
tencies than those recorded during quiet
sleep. Hemispheric asymmetry appears to
be affected by sleep state in adults and
may also be affected in infants as well,
with more hemispheric asymmetry noted
during quiet sleep [Duclaux et al., 1991].
On the other hand, studies of auditory
discrimination in the newborn infant
have shown that infants are able to detect
changes in speech sounds in both quiet
and active sleep [Hirasawa et al., 2002;
Martynova et al., 2003]. An additional
issue surrounding sleep state in young
infants is that behavioral and electroen-
cephalographic assessments of sleep state
may be discordant. Although behavioral
assessments are considered to be the gold
standard for evaluation of sleep [Tho-
man, 1990], there have been no studies
that attempt to describe differences in
ERPs using different methods of assess-
ment of sleep states. It should be noted
that exhaustive studies of the effects of
sleep state have not been performed and
most studies have controlled for state or
excluded infants based upon their sleep
states.
In behavioral studies, differences in
arousal or attention during wakefulness
may also affect the assessed cognitive
Fig. 1.
Individual newborn infant’s ERP demonstrating positive peak (circled) and negative slow
wave. Arrows indicate peak amplitude and latency measurements on respective axes.
318
MRDD R
ESEARCH
R
EVIEWS
●
C
OGNITIVE
E
VALUATION
U
SING
ERP
S
●
DE
R
EGNIER

abilities. Geva et al. [1999] have demon-
strated that differences in arousal level in
awake infants after feedings predict vari-
ability of behavioral responses in tasks of
infant visual memory in the newborn and
1-month-old infant, whereas, in older
infants, responses are less variable and do
not differ with feeding-related arousal
states. Richards and Casey [1991] have
extensively studied visual attention in in-
fants and toddlers have described three
phases of attention that correlate with
heart rate and predict distractibility (ori-
enting, sustained attention, and inatten-
tiveness). Richards has recently demon-
strated that ERPs obtained during these
three phases of attention differ, with the
largest attentional component [negative
component (NC)] described during the
sustained attentional phase [Richards,
2003]. It is therefore apparent that ERPs
are affected by sleep state in sleeping in-
fants and attentional and arousal states in
awake infants.
Thus, even in low-risk infants
studied in a single laboratory setting, in-
dividual differences in ERP findings can
be due to both individual differences in
brain maturation as well as state differ-
ences between infants that may be diffi-
cult to control. When subtly different
paradigms are used by different investi-
gators, it can be very difficult to directly
compare data between studies and derive
normative data for the finer aspects of the
recordings such as peak latencies. All of
this means that ERP differences between
groups of high- and low-risk infants may
be difficult to interpret unless state is
routinely assessed and the study is driven
by a hypothesis that is based upon known
neurobiologic differences between the
studied groups. It is also very important
to study adequate numbers of participants
so that individual differences that are seen
within groups of high- or low-risk in-
fants will not skew the group findings,
potentially masking or falsely amplifying
differences between groups.
TYPES OF COGNITIVE
FUNCTIONS STUDIED USING
ERPS
Although the preceding discussion
has focused on methodological caveats
and sources of variability, ERPs have
been very useful in the evaluation of in-
fants’ auditory perception, auditory dis-
crimination, and auditory and visual
memory. In the adult, auditory cortical
responses (indexing auditory perception)
show a well-ordered pattern of positive
and negative components that are labeled
as P1, N1, P2, and N2 [Regan, 1989].
Although it is possible to record long
latency auditory responses from ex-
tremely premature infants, these wave-
forms undergo a prolonged maturational
period and the adult waveforms are not
fully realized until 20 years of age [Pon-
ton et al., 2000; Kushnerenko et al.,
2002a]. During the first year of life, dra-
matic changes are seen in the morphol-
ogy of the responses, coinciding with
rapid development of the cerebral cortex
at this age. In the extremely preterm in-
fant, the predominant ERP response to
an auditory stimulus is a simple negative
wave with a very long peak latency
[Weitzman and Graziani, 1968]. By term
gestation, a positive peak is seen over the
midline electrode sites in the majority of
infants [Novak et al., 1989; deRegnier et
al., 2002] but, by the end of the first year,
a more stable series of positive and neg-
ative peaks emerges [Kushnerenko et al.,
2002a]. With further development over
the first year of life, the amplitudes of
these components increase in amplitude
and
their
latencies
decrease.
These
changes are thought to be indicative of
synaptogenesis, increased synaptic effi-
ciency and myelination of the brain
[Eggermont, 1988].
Auditory Perception
Although immature in morphol-
ogy compared with adults and older chil-
dren, auditory cortical responses from in-
fants and young children nonetheless
reflect the acoustic properties of the
evoking stimuli [Molfese and Molfese,
1997]. Studies evaluating differences in
ERPs evoked by speech sounds with
phonetical differences provide an impor-
tant method of evaluating the develop-
ment of speech perception. For example,
Molfese et al. have evaluated the devel-
opment of cortical responses to speech
and/or nonspeech stimuli in several stud-
ies [Molfese, 1973; Molfese and Molfese,
1997]. In one of the earliest infant ERP
studies, Molfese evaluated children be-
tween 1 week and 11 years of age com-
pared with young adults, using consonant
syllables, monosyllabic words, a piano
chord, and white noise. At all ages, left
hemisphere amplitudes were larger than
right hemisphere amplitudes for the
speech sounds, with a reversed pattern
for the nonspeech sounds [Molfese,
1973]. Further study of auditory percep-
tion has also shown differences in ERPs
that are elicited by individual speech
sounds, such as /ba/, /da/, and /ga/ in
newborn infants [Molfese and Molfese,
1997; Guttorm et al., 2001]. Differential
responding to acoustic differences in
speech sounds is clearly important for the
further development of receptive lan-
guage and later speech. It would be ex-
pected that infants who have difficulties
with this task may show difficulties with
language and reading and indeed these
early auditory perceptual abilities have
been exploited in ERP studies of high-
risk infants. Some of these studies will be
described below.
The use of ERPs also has been
beneficial in evaluating the development
of the word learning. Several investiga-
tors have evaluated toddler’s brain re-
sponses to known and unknown words
[Molfese, 1989, 1990; Mills et al., 1997,
2004]. In these studies, parents provided
information regarding words that were
thought to be understood by the toddler
and these words were presented in ERP
tasks along with words that were not
known. In all studies, known words elic-
ited different ERP patterns from un-
known words, with differences being
noted in the amplitude and scalp distri-
bution of the ERP activity. With con-
tinued development, Mills et al. [1997,
2004] described a change in brain activity
from being generalized over both hemi-
spheres (at 13–17 months) to becoming a
more localized pattern over the temporal
and parietal areas of the left hemisphere
(at 20 months). The more localized pat-
tern was associated with a larger vocab-
ulary rather than age per se.
Auditory Discrimination
Beyond auditory perception and
word learning, auditory discrimination is
another important ability that can be
studied in young children using ERPs.
Here,
the
perception
of
individual
sounds or words themselves is not the
focus of the study, but rather the brain’s
responses to a change in a stream of
sounds. In a typical auditory change par-
adigm, a frequent sound is occasionally
interrupted by a novel, deviant sound. In
the theory put forth by Naatanen [1995],
when a sound is repeated frequently, a
short-lived, sensory memory trace is es-
tablished. The neural representation of
the novel sound is compared with the
memory trace, and a mismatch response
is generated. In adults, this mismatch re-
sponse is a negative wave (called a mis-
match negativity) that is recorded over
the frontal and central areas of the scalp
[Naatanen, 1995]. Studies of infants and
children have shown evidence of similar
waveform morphologies [Alho et al.,
1990; Cheour-Luhtanen et al., 1996;
Cheour et al., 1998; Hirasawa et al.,
2002], even from preterm infants; ini-
tially it was suspected that this type of
discriminative response might be the on-
togenetically earliest discriminative re-
319
MRDD R
ESEARCH
R
EVIEWS
●
C
OGNITIVE
E
VALUATION
U
SING
ERP
S
●
DE
R
EGNIER

sponse of the human brain [Cheour-Lu-
htanen et al., 1996]. However, with
wider application of the auditory change
paradigm, it has become apparent that
the mismatch response in very young
infants (
⬍3 months of age) is more vari-
able than previously recognized, with
some authors finding no discriminative
responses and other authors finding pos-
itive rather than negative responses [Lep-
panen et al., 1997; Therien et al., 2004].
Some variability in research findings can
be attributed to differences in the meth-
odology of the studies, but Leppanen et
al. [2004] have demonstrated that some
of this variability is due to different levels
of maturity, particularly in newborn in-
fants. With this greater appreciation of
the variability in auditory change re-
sponses, it is no longer clear whether
findings generated in infants are analo-
gous to those found in adults.
However, this qualification has not
limited the usefulness of the auditory
change paradigm. This paradigm has
been used to study the development of
auditory discrimination in infants. If
acoustic differences are too small to be
perceived, no mismatch response will be
noted. This offers the ability to evaluate
infant development of perception of
acoustic differences between sounds, and
to evaluate the role of experience in
learning language. For example, it has
been shown that 2.5–5 h of auditory
training during sleep enabled newborn
infants to discriminate acoustically similar
Finnish vowel sounds that were not dis-
criminated prior to the training [Cheour
et al., 2002]. The auditory change para-
digm has also been used in a number of
studies to evaluate the discriminative
abilities of high-risk children compared
with control children, as will be discussed
later.
Infant Memory
The development of auditory and
visual recognition memory has also been
studied using ERPs. Recognition mem-
ory is a type of explicit memory that is
dependent on the hippocampus and
other medial temporal lobe structures
[Broadbent et al., 2002]. It is the earliest
type of memory to develop and many
behavioral studies have described the rec-
ognition abilities of infants [Rose et al.,
1988, 2001; Fagan, 1990]. It is therefore
possible to create ERP tasks that are
based on behavioral information about
the constraints of early memory develop-
ment.
ERP waveforms that have been
observed in memory studies include the
negative component (NC), the negative
slow wave (NSW), and the positive slow
wave (PSW). The NC is a component
that is thought to index obligatory or
automatic attention to a stimulus [Nel-
son, 1994; Nelson and Monk, 2001;
Richards, 2003]. The NSW is a wave
that emerges when a familiar stimulus is
paired with an entirely novel stimulus.
The wave is thought to represent the
detection of novelty, meaning that the
infant has detected that a stimulus is new
or different, but the stimulus itself has not
been encoded. As encoding abilities im-
prove and infants are able to begin to
encode a novel stimulus during the
course of the ERP study, a PSW is de-
tected in response to a repeated novel
stimulus paired with a familiar stimulus.
It is presumed that repeated presentations
of a novel stimulus result in encoding of
the novel stimulus so that during the
course of the study, the novel stimulus
itself becomes “partially familiar.” It is
thought the PSW represents updating of
memory for the “partially familiar” stim-
ulus [Nelson and Collins, 1991; Nelson
and deRegnier, 1992; Nelson and Monk,
2001].
Although the number of infant and
toddler ERPs is still relatively small, it is
apparent that this technique can be used
to describe the early perceptual, discrim-
inative, and memory abilities of very
young, preverbal infants with limited be-
havioral responses. Though a great deal
of normative data is still required, knowl-
edge of cognitive function in these very
young infants opens up the possibility of
early assessment of cognitive function in
the high-risk infant with congenital, fa-
milial, or perinatal risk factors.
STUDIES OF HIGH-RISK
INFANTS
Timely identification of infants
with congenital, hereditary, or perina-
tally acquired cognitive difficulties may
improve our understanding of cognitive
development in high-risk children and
facilitate the development of focused
early intervention programs for specific
disabilities, similar to the situation that
now exists for early diagnosis of congen-
ital hearing loss. A specific approach is
likely to be most accurate, e.g., early
testing of memory deficits would be most
useful in prediction of later memory abil-
ities.
Toward this end, ERPs have been
used to evaluate neural correlates of cog-
nitive function in a number of groups of
high-risk infants and children. It should
be noted that studies to this date have
focused on evaluating differences in brain
activity between high-and low-risk in-
fants, to further understand cognitive
processes in high-risk children. Some
studies have performed statistical correla-
tions between early ERP activity and
later development, but no studies have
evaluated the reliability, sensitivity, or
specificity of specific ERP patterns, and,
at this time, the usefulness of ERPs in
prediction of developmental disorders in
individual children is untested. Large
longitudinal studies of healthy and high-
risk infants and children are needed be-
fore this technique can be regarded as
clinically useful. Despite this caveat, ERP
studies of high-risk children studied to
date have offered new information on
the neural underpinnings of later cogni-
tive impairments. The most intensively
studied groups include infants of diabetic
mothers, children with familial risk for
dyslexia, and premature infants, and these
groups will be discussed in turn.
Infants of Diabetic Mothers (IDMs)
Infants of poorly controlled dia-
betic mothers are known to have diffi-
culties with cognitive development that
originate in the fetal period [Stehbens et
al., 1977; Rizzo et al., 1997]. Poorly
controlled diabetes may lead to fetal met-
abolic derangements that can be delete-
rious to fetal brain development, includ-
ing hypoglycemia, ketonemia, acidosis,
chronic hypoxia, and brain iron deple-
tion [Georgieff and Rao, 2001]. In ani-
mal models, both brain iron depletion
and hypoxia result in aberrant develop-
ment of neural structures important for
explicit or declarative memory [deUngria
et al., 2000]. Thus it would be predicted
that children of poorly controlled dia-
betic mothers may have difficulties with
explicit memory and this hypothesis has
recently been tested using ERPs.
Memory development in a cohort
of IDMs from Minnesota has been stud-
ied from birth using ERPs and behavioral
testing (deRegnier et al., 2000; Nelson et
al., 2000, 2003; Sidappa et al., 2004; De-
Boer et al., 2005b]. It is known that
newborn infants are capable of recogniz-
ing the maternal voice [DeCasper and
Fifer, 1980]. For the first test, ERPs were
recorded at term gestation (40 weeks;
mean postnatal age of 10 days) while
infants listened to the maternal voice
compared with a stranger’s voice [de-
Regnier et al., 2000]. The word “baby”
was chosen as a stimulus. The stranger’s
voice was the voice of the previous
mother and therefore varied for each in-
fant. In the control group’s ERPs, the
maternal voice elicited a positive wave
peaking at approximately 290 ms. This
same peak was noted in response to the
320
MRDD R
ESEARCH
R
EVIEWS
●
C
OGNITIVE
E
VALUATION
U
SING
ERP
S
●
DE
R
EGNIER

stranger’s voice, but the ERP elicited by
the stranger’s voice also elicited a nega-
tive slow wave (consistent with novelty
detection). In the IDM group, these
findings were attenuated and no signifi-
cant differences were noted between the
ERPs elicited by the maternal and strang-
er’s voice. Follow-up through 1 year of
age showed that the presence of the neg-
ative slow wave for the stranger’s voice
was associated with higher scores on the
Bayley Scales of Infant Development in
both the control and IDM groups [de-
Regnier et al., 2000]. Since no children
in this study had delayed development,
the negative slow wave was interpreted as
a
marker
of
better
developmental
progress.
The authors speculated that the
differences in neonatal memory in the
IDM group may have been due to peri-
natal brain iron deficiency. IDMs are
known to be at risk for perinatal iron
deficiency, and, in animal models, this
causes iron depletion in the explicit
memory network that is important for
recognition memory [deUngria et al.,
2000]. To evaluate whether the neonatal
findings in the IDM group could be re-
lated to perinatal brain iron depletion, a
second group of IDMs were tested and
grouped according to their iron status at
birth [Sidappa et al., 2004]. Ferritin is an
iron storage protein in the body, and very
low ferritin values at birth (
⬍35 g/dL)
have been associated with depletion of
brain iron (see Sidappa et al., 2004 for a
discussion). The next newborn study
stratified IDMs into two groups, one
with suspected brain iron deficiency (fer-
ritin
⬍ 35 g/dL) and another presumed
to have brain iron sufficiency (ferritin
⬎
35
g/dL). The infants were tested at 40
weeks postmenstrual age and a mean
postnatal age of 2 weeks. The IDMs
thought to be brain iron sufficient
showed findings similar to control infants
from the previous study (a negative slow
wave for the stranger’s voice), whereas
the IDMs suspected to be brain iron de-
ficient showed markedly attenuated find-
ings, with no differences seen between
the ERPs derived from the maternal and
stranger’s voices. In this group of infants,
there was a correlation between the se-
verity of iron deficiency (measured by
the ferritin concentration) and the ERP
patterns, with infants having higher fer-
ritin values showing ERP patterns typical
of healthy control infants. Although it is
necessary to more fully evaluate the role
of other factors such as chronic hypoxia
in these results, the authors have specu-
lated that perinatal brain iron deficiency
affects the development of human in-
fants’ memory networks in a similar
manner as in animal models.
Not only have infants with diabetic
mothers shown neurophysiologic evi-
dence of differences in auditory recogni-
tion memory, these findings have per-
sisted in visual recognition memory at 6
months [Nelson et al., 2000]. Healthy
6-month-old infants show behavioral
and ERP evidence of recognition of the
maternal face [de Haan and Nelson,
1997; Nelson et al., 2000], and a fol-
low-up study evaluated this ability in
6-month-old IDMs compared with con-
trol infants tested at the same age [Nelson
et al., 2000]. ERPs were recorded while
each infant was shown digital photo-
graphs of the maternal face and a strang-
er’s face. As expected from previous
studies, the control infants showed a
larger NC to the maternal face that was
followed in the recording by a positive
slow wave for the stranger’s face. In con-
trast, the IDMs showed no significant
differences between the maternal and
stranger’s
ERPs.
Consequently,
this
study showed that the presumed deficits
in auditory recognition memory seen in
the full-term newborn were again found
in the visual modality at 6 months of age.
Cross-modal recognition memory
refers to the ability to recognize objects
in different sensory modalities than the
original presentation. In infant studies,
infants are typically tested for visual rec-
ognition of objects that previously were
palpated but not seen. Behavioral studies
have shown when 8-month-old infants
palpate a simple but unique shape in the
dark, they show visual recognition mem-
ory for the palpated object [Rose et al.,
1983].
Neurophysiologic
studies
of
8-month-old infants have shown that,
similar to the maternal face, the familiar
palpated object elicits a larger NC com-
ponent than does the novel object and,
similar to the situation with the stranger’s
face, there is a positive slow wave for the
ERP from the novel object [Nelson et
al., 1993]. However, in a study of infants
of diabetic mothers, the slow wave was
not seen, indicating that alterations in
memory development seen in the new-
born infant were persistent through 8
months of age [Nelson et al., 2003] and
noted in multiple modalities.
With this series of ERP studies,
deficits of auditory, visual, and cross-
modal recognition memory were de-
scribed in infants of diabetic mothers.
Furthermore, specific ERP patterns re-
corded from the neonate were associated
with mental developmental test scores at
1 year of age. However, these findings
would not be of concern if there were no
behavioral manifestations of memory
deficits. A recent study [DeBoer et al.,
2005b] evaluated IDMs at 12 months of
age for behavioral evidence of memory
using an elicited imitation paradigm. In
an elicited imitation paradigm, sequences
of events leading to the creation of a toy
are modeled for the infant. Infants are
then provided with the supplies needed
to recreate the toy, either immediately or
after a delay. In this study, infants of
diabetic mothers were found to have dif-
ficulties with elicited imitation after a
delay, providing behavioral substantia-
tion of the previous ERP findings.
Though long-term follow up of this co-
hort continues, this series of ERP and
behavioral studies was useful in translat-
ing information gleaned from animal
models of iron deficiency into informa-
tion about memory development in hu-
man infants. These findings have led the
Minnesota IDM research group to initi-
ate a trial of early iron therapy for infants
of diabetic mothers with suspected brain
iron deficiency to try to improve mem-
ory development in these infants.
Infants at Risk for Dyslexia
The relationship between early
speech perception and later reading skills
or dyslexia has been the source of a num-
ber of studies correlating early ERPs with
later cognitive function. Dyslexia, or
reading disorder, is a learning disability
that is known to have a familial basis and
therefore may be amenable to early diag-
nosis [Lyytinen et al., 2004]. The under-
lying neuropsychological deficit that un-
derlies dyslexia is not known, but
difficulties with speech perception have
been suspected. Children with dyslexia
may have very early deficits with speech
perception that may be useful in the early
diagnosis of dyslexia that is formally di-
agnosed much later. Given that ERPs
have been used to evaluate early neural
underpinnings of speech perceptions,
several authors have used ERPs to test
infants at risk for dyslexia and have de-
tected early differences in auditory corti-
cal responses to speech sounds. In the
ongoing Jyvaskyla Longitudinal Study of
Dyslexia [Lyytinen et al., 2004], children
with a strong family history for dyslexia
have been followed since birth with a
combination of ERP and behavioral
tests. ERPs were conducted in the new-
born period in response to consonant-
vowel speech sounds (/ba/, /da/, and
/ga/). Those newborns with a family his-
tory of dyslexia showed a difference in
the hemispheric patterns of activation for
the speech sound /ga/, with larger and
more positive responses recorded in the
321
MRDD R
ESEARCH
R
EVIEWS
●
C
OGNITIVE
E
VALUATION
U
SING
ERP
S
●
DE
R
EGNIER

high-risk group over the right hemi-
sphere compared with children at low
risk for dyslexia, possibly indicating poor
hemispheric specialization for language
in the high-risk children. Five-year out-
comes of these children have been re-
ported. The ERP findings that were
characteristic of the risk group showed a
significant association with poorer recep-
tive language at 2.5 years of age. Further-
more, poor verbal memory skills at age 5
years were predicted by neonatal ERP
findings [Guttorm et al., 2001].
These same authors also studied in-
fants at risk for dyslexia at 6 months of
age in comparison with low-risk infants.
For this study, they utilized an auditory
change paradigm in which a long vowel
served as the frequent stimulus and a
short vowel served as the deviant stimu-
lus.
They
demonstrated
that
larger
change detection responses over the left
hemisphere predicted better receptive
language skills at 2.5 years and better
verbal memory skills at 3.5 years. Larger
change detection response over the right
hemisphere was associated with poorer
verbal memory at 5 years [Lyytinen et al.,
2004]
Molfese et al. also reported data
consistent with these findings in a group
of children studied from birth through
age 8 using ERPs and behavioral testing
[Molfese and Molfese, 1985, 1997;
Molfese, 2000; Molfese et al., 2001].
Neonatal ERPs from children diagnosed
with dyslexia at 8 years differed from
ERPs obtained from neonates who were
later found to be normal readers. Several
differences were found between normal
readers, poor readers, and dyslexic chil-
dren in their neonatal ERP responses to
speech sounds over both right and left
hemispheres. The investigators used a
discriminant function analysis to identify
areas of difference between the control
and dyslexic children’s neonatal ERPs.
They found that measures of peak ampli-
tudes and latencies in the neonatal period
correctly classified 13 of 17 dyslexic chil-
dren, a finding well above chance levels
of prediction [Molfese, 2000].
This same group also used ERPs
longitudinally, every 6 months from
birth through 3 years of age in 16 infants
in response to synthetic speech stimuli
that had been previously found to pro-
duce hemispheric difference effects as
well as discrimination effects. The best
predictor of later development was found
to be lateralized speech sound discrimi-
nation ability. Children with left hemi-
sphere– generated discrimination of con-
sonant sounds showed better language
skills at 3 years [Molfese and Molfese,
1985].
Molfese et al. [2001] also evaluated
the predictive value of newborn ERPs
coupled with 3-year evaluations of the
home environment, language develop-
ment, and early intelligence testing in
prediction of 8-year reading scores, using
the
WRAT-R.
Multiple
regression
models were created to evaluate these
early factors in prediction of the 8-year
reading scores. Newborn ERP peak la-
tency measurements over the right tem-
poral region and the left frontal area in
response to the speech sound /gi/ con-
tributed significantly to the regression
model and were correlated with the
8-year reading scores. The authors hy-
pothesized that the results of their mul-
tiple studies indicate the children who
process
language
information
more
quickly from birth have advantages in
learning language and, later, in learning
to read.
Overall these studies suggest that
very early brain responses to speech sounds
are important in language acquisition. The
speed of these responses and hemispheric
lateralization both appear to be important,
though a consensus has not yet been
reached regarding the basic underlying al-
terations in the neural pathways leading to
dyslexia and poor reading skills.
Premature Infants
Premature infants, particularly those
born at less than 32 weeks gestation, are at
high risk for long-term cognitive problems
including lowered intelligence, memory
impairments, attention deficit disorders,
learning disabilities, and difficulties with
verbal processing [Rose et al., 1988, 2001;
Aylward, 2002]. Premature infants are a
heterogeneous group of infants who may
experience multiple risk factors, including
intracranial hemorrhage and white matter
damage, perinatal nutritional deficits, an al-
tered environment, and physiological insta-
bility with recurrent episodes of hypoxia,
hypotension, or acidosis [Mattia and de-
Regnier, 1998; Maalouf et al., 1999; Hack
and Taylor, 2000]. Because some of the
factors associated with premature infants’
development may be modifiable, it is very
important to develop techniques to under-
stand early brain function in these infants,
so that interventions can be tested without
the long time lag associated with traditional
assessment of cognitive function. For this
group of infants, ERPs are likely to be a
useful adjunct to anatomic imaging proce-
dures that are currently very popular.
Early language acquisition may be
delayed and later complex verbal process-
ing may be problematic in preterm chil-
dren [Aylward, 2002]. In contrast to
IDMs or infants at risk for dyslexia, no
longitudinal studies have followed pre-
term children from birth to school age
using ERPs and behavioral or standard-
ized testing. However, some shorter fol-
low-up studies and cross-sectional studies
have evaluated auditory perception and
discrimination in premature infants.
Kurtzberg et al. [1984] tested pre-
mature, very low birth weight infants’
responses to speech sounds (/da/ and
/ta/) and a 800-Hz tone at term (40
weeks postmenstrual age) compared with
full-term infants. Monthly ERPs were
conducted until 3 months corrected age.
The premature infants showed immature
ERP patterns for the speech sounds at 40
weeks with progressive normalization of
the ERPs by 3 months corrected age.
Similar differences were seen in the pre-
term infants’ responses to the 800-Hz
tone, but these findings did not achieve
statistical significance. The authors theo-
rized that relative immaturity of auditory
cortical processing of speech might con-
tribute to subsequent difficulties with
speech and language development.
An auditory change paradigm was
used in a more recent study of preterm
infants, including a group of infants with
intrauterine growth restriction [Fellman
et al., 2004]. In this study, ERPs were
recorded from preterm infants and con-
trol infants while infants were tested for
discrimination of a change in harmonic
tone frequency. Infants were tested at
term and 6 and 12 months of age, cor-
rected for prematurity. In the infants
tested at term, the control group showed
evidence of auditory change detection at
several time points in the ERP, whereas
the responses were atypical or absent in
the preterm infants with and without in-
trauterine growth restriction. Some of
the ERP findings at each age (term, 6,
and 12 months corrected age) were
found to correlate with 2-year tests of
mental development (the Bayley Scales
of Infant Development).
Older preterm children have also
been the subject of some ERP studies.
Jansson-Verkasalo et al. [2004] in Finland
evaluated 4-year-old former preterm
children using ERPs and the auditory
change paradigm using syllables and fol-
lowed these children until 6 years of age,
at which time an object naming test and
additional ERP was performed. The pre-
term infants showed reduced amplitudes
of the auditory change response com-
pared with full-term control infants. The
amplitude of the 4-year auditory change
response correlated with the 6-year re-
sponse, and the absence of the auditory
322
MRDD R
ESEARCH
R
EVIEWS
●
C
OGNITIVE
E
VALUATION
U
SING
ERP
S
●
DE
R
EGNIER

change response at 4 years predicted
naming difficulties at 6 years. This study
showed stability of the ERP findings
over 2 years and revealed that difficulties
with auditory change detection were as-
sociated with language difficulties.
These studies show evidence of
persistent alteration of the neural path-
ways for perception and discrimination
of speech sounds in preterm infants from
term gestation through the later pre-
school years. Alterations in perception
and discrimination of speech sounds may
be indicative of later difficulties with lan-
guage and cognitive development. The
sensitivity and specificity of the ERP
findings has not been tested to determine
whether ERPs could be used as predic-
tors of language development and verbal
processing for individual infants.
In addition to difficulties with lan-
guage and verbal development, preterm
infants also have been shown to have
deficits in visual recognition memory
that persist over the first year of life and
correlate with later intelligence [Rose et
al., 1988, 2001]. Additionally, a syn-
drome of developmental amnesia as well
as a growing spectrum of clinical mem-
ory deficits have been described in ado-
lescents or young adults who were born
prematurely [Isaacs et al., 2000, 2003;
Vargha-Khadem et al., 2001]. MRI stud-
ies have shown that these findings are
associated with decreased hippocampal
volumes [Isaacs et al., 2003]. These be-
havioral and MRI studies indicate that
preterm infants are at increased risk for
hippocampal injury that can be associated
with behavioral deficits if severe.
ERPs have recently been used to
evaluate memory development at term
(40 weeks) postmenstrual age in ex-
tremely premature infants by Therien et
al. [2004]. Preterm infants with normal
cranial ultrasound tests at 1 week and 1
month of age were compared with full-
term control infants for ERP responses to
the maternal voice compared with a
stranger’s voice. For the preterm infant
group, no significant differences were
found between the maternal and strang-
er’s voice ERPs. Lack of difference be-
tween the maternal and stranger’s voice
ERPs persisted even when an additional
familiarization period was provided to
increase the infants’ experience with the
maternal voice (though 80% of the in-
fants had been discharged for at least 6
days before the ERP test was conducted).
An auditory change paradigm was also
used in this study to determine whether
patterns of auditory discrimination cor-
related with memory abilities. Interest-
ingly, the control infants and preterm
infants both showed evidence of auditory
change detection, but the ERP patterns
were opposite from each other. For the
control group, the auditory change re-
sponse was a positive wave over the fron-
tal scalp, whereas the preterm infants’
ERPs showed a negative auditory change
response over frontal and lateral scalp
sites. There was no correlation between
the results of the auditory change para-
digm and the maternal voice recognition
paradigm. The findings were consistent
with altered development of multiple
neural pathways, including those in-
volved with auditory recognition mem-
ory and auditory discrimination.
These studies indicate that cogni-
tive impairments in preterm infants al-
most certainly have their origins in the
perinatal time period. These data and re-
cent MRI studies [Maalouf et al., 1999]
suggest that many preterm infants expe-
rience brain injury or disruption of the
normal developmental trajectory. For the
children who ultimately have normal
outcomes, it seems possible that families
are providing a nurturing environment
that facilitates positive changes in brain
development and function. Several of the
studies described here have shown effects
of the environment or experience on
ERP manifestations of auditory percep-
tion or recognition memory [Cheour et
al., 1998,;2002; Molfese et al., 2001; de-
Regnier et al., 2002]. The ERP tech-
nique has the potential to evaluate
changes in brain development and func-
tion that are associated with interventions
to improve developmental outcomes,
even in the preverbal infant. Although
underutilized for this purpose currently,
this may be the most promising use of
this technique in our efforts to help im-
prove the outcomes of high-risk infants.
CONCLUSION
The results of the ERP studies pre-
sented here indicate that memory abilities
and speech perception and discrimination
are present from birth and are important in
the development of both specific and gen-
eral cognitive abilities. These studies have
shown that ERPs can be used to evaluate
cognitive function and neural plasticity in a
longitudinal fashion beginning prior to the
onset of language and sophisticated behav-
ior. It is clear that early perceptual abilities
are tied to later cognitive skills. Further-
more, although cognitive function is very
immature in infants and is not likely to be
immutably fixed from birth, the specific
cognitive abilities seen in even the young-
est infants do appear to germinate into
more mature abilities in both healthy and
high-risk children. Very early differences in
perceptual and cognitive function are seen
between low-risk infants and infants with a
variety of risk factors, including prematu-
rity, uncontrolled maternal diabetes, and
familial disorders such as dyslexia. In a small
number of studies thus far, these differences
in very early brain function have been pre-
dictive of later difficulties. It should be
noted that the sensitivity, specificity, and
predictive value of individual infants’ ERPs
have not yet been tested. Furthermore, no
systematic study of individual differences
has been performed in low-risk infants.
This will be important because there is a
great deal of variability seen in infant ERPs
that may or may not be indicative of cog-
nitive impairments. A great deal of further
research is needed before ERPs can be used
to evaluate individual infants, but the tech-
nique shows promise in early diagnosis and
assessing new therapies for infants at risk for
later cognitive impairments.
f
REFERENCES
Alho K, Sainio K, Sajaniemi N, et al. 1990. Event-
related brain potential of human newborns to
pitch change of an acoustic stimulus. Electro-
encephal Clin Neurophysiol 77:151–155.
Aylward GP. 2002. Cognitive and neuropsycho-
logical outcomes: More than intelligence
quotient (IQ) scores. Ment Retard Dev Dis-
abil Res Rev 8:234 –240.
Broadbent NJ, Clark RE, Zola S, et al. 2002. The
medial temporal lobe and memory. In: Squire
LR, Schacter DL, editor. Neuropsychology
of memory. 3rd edition. New York: Guilford
Press. p 3–23.
Cheour-Luhtanen M, Alho K, Sainio K, et al.
1996. The ontogenetically earliest discrimina-
tive response of the human brain. Psycho-
physiology 33:478 – 481.
Cheour M, Ceponiene R, Lehtokoski A, et al.
1998. Development of language-specific pho-
neme representations in the infant brain. Nat
Neurosci 1:351–353.
Cheour M, Martynova O, Naatanen R, et al. 2002.
Speech sounds learned by sleeping newborns.
Nature 415:599 – 600.
Coles MGH, Rugg MD. 1995. Event-related brain
potentials: an introduction. In: Rugg MD,
Coles MGH, editors. Electrophysiology of
mind: event-related potentials and cognition.
Volume 25. Oxford: Oxford University
Press. p 1–26.
de Haan M, Nelson CA. 1997. Recognition of the
mother’s face by 6-month-old infants: a neu-
robehavioral study. Child Dev 73:187–210.
DeBoer T, Scott LS, Nelson CA. 2005. ERPs in
developmental populations. In: Handy TC,
editor. Event-related potentials: a methods
handbook. Cambridge, MA: The MIT Press.
p 264 –297.
DeBoer T, Wewerka S, Bauer PJ. 2005. Explicit
memory performance in infants of diabetic
mothers at 1 year of age. Dev Med Child Neu-
rol 47:525–531.
DeCasper AJ, Fifer WP. 1980. Of human bonding:
newborns prefer their mothers’ voices. Sci-
ence 208:1174 –1176.
DeCasper AJ, Spence MJ. 1986. Prenatal maternal
speech influences newborns’ perception of
speech sounds. Infant Behav Dev 6:19 –25.
deRegnier R, Nelson CA, Thomas K, et al. 2000.
Neurophysiologic evaluation of auditory rec-
323
MRDD R
ESEARCH
R
EVIEWS
●
C
OGNITIVE
E
VALUATION
U
SING
ERP
S
●
DE
R
EGNIER

ognition memory in healthy newborn infants
and infants of diabetic mothers. Disabil Re-
habil 137:777–784.
deRegnier R, Wewerka S, Georgieff MK, et al.
2002. Influences of post-conceptional age and
postnatal experience on the development of
auditory recognition memory in the newborn
infant. Dev Psychobiol 41:216 –225.
deUngria M, Rao R, Wobken JD, et al. 2000.
Perinatal iron deficiency decreases cyto-
chrome c oxidase activity in selective regions
of the brain. Pediatr Res 48:169 –176.
Duclaux R, Challamel MJ, Collet L, et al. 1991.
Hemispheric asymmetry of late auditory
evoked response induced by pitch changes in
infants: influence of sleep stages. Brain Res
566:152–158.
Eggermont JJ. 1988. On the rate of maturation of
sensory evoked potentials. Electroencephal
Clin Neurophysiol 70:293–305.
Fagan JFI. 1990. The paired-comparison paradigm
and infant intelligence. Ann NY Acad Sci
608:337–364.
Fellman V, Kushnerenko E, Mikkola K, et al. 2004.
Atypical auditory event-related potentials in
preterm infants during the first year of life: a
possible sign of cognitive dysfunction? Pediatr
Res 56:291–297.
Georgieff MK, Rao R. 2001. The role of nutrition
in cognitive development. In: Nelson CA,
Luciana M, editors. Handbook of develop-
mental cognitive neuroscience. Cambridge,
MA: MIT Press. p 45–58.
Geva R, Gardner JM, Karmel BZ. 1999. Feeding-
based arousal effects on visual recognition
memory in early infancy. Dev Psychol 35:
640 – 650.
Ginsberg H, Opper S. 1979. Piaget’s theory of
intellectual development. Englewood Cliffs,
NJ: Prentice-Hall.
Guttorm TK, Leppanen PHT, Richardson U, et al.
2001. Event-related potentials and consonant
differentiation in newborns with familial risk
for dyslexia. J Learn Disabil 34:534 –544.
Hack M, Taylor HG. 2000. Perinatal brain injury
in preterm infants and later neurobehavioral
function. JAMA 284:1973–1974.
Hirasawa K, Kurihara M, Konishi Y. 2002. The
relationship between mismatch negativity and
arousal level: can mismatch negativity be an
index for evaluating the arousal level in in-
fants? Sleep Med Suppl 2:S45– 48.
Isaacs EB, Lucas A, Chong WK, et al. 2000. Hip-
pocampal volumes and everyday memory in
children of very low birthweight. Pediatr Res
47:713–720.
Isaacs EB, Vargha-Khadem F, Watkins KE, et al.
2003. Developmental amnesia and its rela-
tionship to degree of hippocampal atrophy.
Proc Acad Natl Sci USA 100:13060 –13063.
Jansson-Verkasalo E, Korpilahti P, Jantti V, et al.
2004. Neurophysiologic correlates of defi-
cient phonological representations and object
naming in prematurely born children. Clin
Neurophysiol 115:179 –187.
Kurtzberg D, Hilpert PL, Kreuzer JA, et al. 1984.
Differential maturation of cortical auditory
evoked potentials to speech sounds in normal
full term and very low-birthweight infants.
Dev Med Child Neurol 26:466 – 475.
Kushnerenko E, A
˜ eponiene¨ R, Balan P, et al.
2002a Maturation of the auditory event-re-
lated potentials during the first year of life.
NeuroReport 13:47–51.
Kushnerenko E, A
˜ eponiene¨ R, Balan P, et al.
2002b. Maturation of the auditory change
detection response in infants: a longitudinal
ERP study. NeuroReport 13:1843–1848.
Leppanen PHT, Eklund KM, Lyytinen H. 1997.
Event-related brain potentials to change in
rapidly presented acoustic stimuli in new-
borns. Dev Neuropsychol 13:175–204.
Leppanen, PHT, Guttorm TK, Pihko E, et al.
2004. Maturational effects on newborn ERPs
measured in the mismatch negativity para-
digm. Exp Neurol 190:S91–S101.
Lyytinen H, Ahonen T, Eklund K, et al. 2004.
Early development of children at familial risk
for dyslexia-follow-up from birth to school
age. Dyslexia 10:146 –178.
Maalouf EF, Duggan PJ, Rutherford MA, et al.
1999. Magnetic resonance imaging of the
brain in a cohort of extremely preterm infants.
Disabil Rehabil 135:351–357.
Martynova O, Kirjavainen J, Cheour M. 2003.
Mismatch negativity and late discriminative
negativity in sleeping human newborns. Neu-
rosci Lett 340:75–78.
Mattia FR, deRegnier RAO. Chronic physiologic
instability is associated with neurodevelop-
mental morbidity at one and two years in
extremely premature infants. Pediatrics elec-
tronic pages 102:e35, 1998.
Mills DL, Coffey-Corina SA, Neville HJ. 1997.
Language comprehension and cerebral spe-
cialization from 13–20 months. Dev Neuro-
psychol 13:397– 446.
Mills DL, Prat C, Zangl R, et al. 2004. Language
experience and the organization of brain ac-
tivity to phonetically similar words: ERP ev-
idence from 14- and 20-month olds. J Cogn
Neurosci 16:1452–1464.
Molfese DL. 1973. Cerebral asymmetry in infants,
children and adults: auditory evoked re-
sponses to speech and noise stimuli. Diss Abstr
Int 34:1298.
Molfese DL. 1989. Electrophysiological correlates
of word meanings in 14-month-old human
infants. Dev Neuropsychol 5:79 –103.
Molfese DL. 1990. Auditory evoked responses re-
corded from 16-month-old human infants to
words they did and did not know. Brain Lang
38:345–363.
Molfese DL. 2000. Predicting dyslexia at 8 years of
age using neonatal brain responses. Brain
Lang 72:238 –245.
Molfese DL, Molfese VJ. 1985. Electrophysiological
indices of auditory discrimination in newborn
infants: the basis for predicting later language
performance? Infant Behav Dev 8:197–211.
Molfese DL, Molfese VJ. 1997. Discrimination of
language skills at five years of age using event-
related potentials recorded at birth. Dev Neu-
ropsychol 13:135–156.
Molfese VJ, Molfese DL, Modglin AA. 2001. New-
born and preschool predictors of second-
grade reading scores: an evaluation of cate-
gorical and continuous scores. J Learn Disabil
34:545–554.
Naatanen R. 1995. The mismatch negativity: a
powerful tool for cognitive neuroscience. Ear
Hearing 16:6 –18.
Nelson CA. 1994. Neural correlates of recognition
memory in the first postnatal year of life. In:
Dawson G, Fischer K, editors. Human behav-
ior and the developing brain New York:
Guilford Press. p 269 –313.
Nelson CA, Collins PF. 1991. An event-related
potential and looking time analysis of infants’
responses to familiar and novel events: impli-
cations for visual recognition memory. Dev
Psychol 27:50 –58.
Nelson CA, deRegnier R. 1992. Neural correlates
of attention and memory in the first year of
life. Dev Neuropsychol 8:119 –134.
Nelson CA, Henschel M, Collins PF. 1993. Neural
correlates of cross-modal recognition mem-
ory by 8-month-old infants. Brain Cogn 29:
411– 420.
Nelson CA, Monk CS. 2001. The use of event-
related potentials in the study of cognitive
development. In: Nelson CA, Luciana M.
editors. Handbook of developmental cogni-
tive neuroscience. Cambridge, MA: MIT
Press. p 125–136.
Nelson CA, Wewerka S, Borscheid AJ, et al. 2003.
Electrophysiologic
evidence
of
impaired
cross-modal
recognition
memory
in
8-month-old infants of diabetic mothers. J
Pediatrics 142:575–582.
Nelson CA, Wewerka S, Thomas KM, et al. 2000.
Neurocognitive sequelae of infants of diabetic
mothers. Behav Neurosci 114:950 –956.
Novak GP, Kurtzberg D, Kreuzer JA, et al. 1989.
Cortical responses to speech sounds and their
formants in normal infants: maturational se-
quence and spatiotemporal analysis. Electro-
encephal Clin Neurophysiol 73:295–305.
Ponton CW, Eggermont JJ, Kwong B, et al. 2000.
Maturation of human central auditory system
activity: evidence from multi-channel evoked
potentials. Clin Neurophysiol 111:220 –236.
Regan D. 1989. Human brain electrophysiology:
evoked potentials and evoked magnetic fields
in science and medicine. New York: Elsevier.
Richards JE. 2003. Attention affects the recognition
of briefly presented visual stimuli in infants: an
ERP study. Dev Sci 6:312–328.
Richards JE, Casey BJ. 1991. Heart rate variability
during attention phases in young infants. Psy-
chophysiology 28:43–53.
Rizzo TA, Metzger BE, Dooley SL, et al. 1997. Early
malnutrition and child neurobehavioral devel-
opment: insights from the study of children of
diabetic mothers. Child Dev 68:26 –38.
Rose SA, Feldman JF, Jankowski JJ. 2001. Atten-
tion and recognition memory in the 1st year
of life: a longitudinal study of preterm and
full-term infants. Dev Psychol 37:135–151.
Rose SA, Feldman JF, McCarton CM, et al. 1988.
Information processing in seven-month-old
infants as a function of risk status. Child Dev
59:589 – 603.
Rose SA, Gottfried AW, Bridger WH. 1983. In-
fants’ cross-modal transfer from solid objects
to their graphic representations. Child Dev
54:686 – 694.
Sidappa A, Georgieff MK, Wewerka S, et al. 2004.
Iron deficiency alters auditory recognition
memory in newborn infants of diabetic moth-
ers. Pediatr Res 55:1034 –1041.
Stehbens JA, Baker GL, Kitchell M. 1977. Out-
come at ages 1, 3, and 5 years of children born
to diabetic women. Am J Obstet Gynecol
127:408 – 413.
Therien JM, Worwa CT, Mattia FR, et al. 2004.
Altered pathways for auditory discrimination
and recognition memory in premature new-
borns. Dev Med Child Neurol 46:816 – 824.
Thoman EB. 1990. Sleeping and waking states in
infants: a functional perspective. Neurosci
Biobehav Rev 14:93–107.
Vargha-Khadem F, Gadian DG, Mishkin M. 2001.
Dissociations in cognitive memory: the syn-
drome of developmental amnesia. Philos Trans
R Soc Lond Ser B Biol Sci 356:1435–1440.
Weitzman ED, Graziani LJ. 1968. Maturation and
topography of the auditory evoked response
of the prematurely born infant. Dev Psycho-
biol 1:79 – 89.
324
MRDD R
ESEARCH
R
EVIEWS
●
C
OGNITIVE
E
VALUATION
U
SING
ERP
S
●
DE
R
EGNIER

Wyszukiwarka
Podobne podstrony:
więcej podobnych podstron