
http://bioslawek.wordpress.com/2011/07/17/13
7/#more-137
‘Marzenie nanotechnologów: silnik protonowy
bakterii Escherichia coli’
Pozwoliłem sobie wzbogacić artykuł p. Michała Ostrowskiego kilkoma dodatkowymi
ilustracjami poglądowymi, komentarzami do nich ,linkami oraz filmami. W tym poniższym
cytatem B.Webera.
Przeciwnik koncepcji Inteligentnego Projektu oraz nieredukowalnej złożoności, biochemik
Bruce Weber napisał :
www.nauka-a-religia.uz.zgora.pl/index.php?action=tekst&id=89
„(……)Udzielono licznych odpowiedzi Behe’emu – mieli w tym udział
również biochemicy, łącznie ze mną – które odnosiły się do tego, jak
naprawdę powinien on przedstawić obecny stan literatury przy
uwzględnieniu podanych przez niego konkretnych przykładów. 2 W
rzeczywistości opublikowano próby wyjaśnienia zagadnień, takich jak
powstanie wici, układu krzepnięcia krwi czy biochemicznej podstawy
procesu widzenia.
Należy przyznać, że przedstawienie tego, w jaki sposób aktualne dane biologii molekularnej
sugerują procesy duplikacji genu, tasowania domen i eksonów oraz ewolucji dywergentnej nie
wyjaśnia, jak NIEKOMPLETNE SYSTEMY (przyp. moja: nie w pełni ukształtowane pewne
systemy biochemiczne-jak szlaki metaboliczne , czy organy , jak np. : „ćwierć-witka czy pół-witka
bakteryjna”) zyskują przewagę selekcyjną.
Obecnie jesteśmy jednak na etapie gromadzenia wystarczającej ilości danych na podstawie
sekwencji DNA i trójwymiarowych struktur białek, by w niedalekiej przyszłości
przeprowadzać liczne testy przypuszczalnych wyjaśnień ewolucyjnych, co do których można
by się spodziewać, że nie będą „takimi sobie bajeczkami”.”
Nie trudno jest zauważyć , że tym samym Bruce Weber przyznaje iż ewolucjoniści nie mają
pojęcia , w jaki możliwy do zaakceptowania sposób mogły wyewoluować układy
nieredukowalnie złożone. Przy okazji sugerując zachowanie optymizmu i nadziei na

przyszłość Bruce Weber dopuszcza się w swojej argumentacji błędów logicznych :
1) „Argument to the future”- ‘Błąd ten popełnia się, gdy się twierdzi, że coś w przyszłości uda
się wykazać, choć teraz jest to niemożliwe’.
2) „Hypothesis contrary to fact”-Błąd tego typu powstaje wtedy, gdy ktoś argumentuje na
podstawie tego, że coś mogło się zdarzyć, choć jednocześnie nic nie wskazuje na to, że tak się
zdarzyło, lub nawet coś temu przeczy.’
Czy koncepcja oparta na braku jakiegokolwiek modelu teoretycznego, poza kilkoma
„TAKIMI SOBIE BAJECZKAMI”, jak np. przestarzały i przereklamowany model Matzkego
(będzie w przyszłości na temat tego „modelu” osobny artykuł), i na błędach logicznych
zasługuje na miano ” teorii naukowej ” ?
(‘The Evolution of the Flagellum’
And the Climbing of „Mt. Improbable „
Sean D. Pitman, M.D.
http://www.detectingdesign.com/flagellum.html )
http://creationism.org.pl/artykuly/MOstrowski2
‘Marzenie nanotechnologów: silnik protonowy bakterii Escherichia coli’
Michał Ostrowski
Silnik bakteryjny (Wić bakteryjna).
Wprowadzenie :
Silnik protonowy bakterii Escherichia coli robi ostatnio oszałamiającą karierę często
pojawiając się w literaturze kreacjonistycznej jako przykład systemu nieredukowalnie
złożonego. Przykład ten spopularyzował amerykański biolog molekularny, Michael Behe w
swojej bestsellerowej książce Darwin’s Black Box. [1] Dzieje się tak ze względu na intuicyjne
narzucającą się analogię tego systemu z wytworami ludzkiej technologii oraz na niezwykłe
wyrafinowanie jego funkcjonowania i budowy, dzięki czemu niektórzy naukowcy nazywają
bakteryjny silnik protonowy „najbardziej wydajną maszyną świata”. Nic dziwnego, że ta
molekularna maszyna stała się wręcz maskotką ruchu teorii inteligentnego projektu. [2]

Nanotechnologia w praktyce
Escherichia coli (pałeczka okrężnicy) jest jednokomórkowym cylindrycznym organizmem
żyjącym powszechnie w naszych jelitach. Swoją nazwę zawdzięcza austriackiemu lekarzowi
— Theodorowi Escherichowi — który po raz pierwszy zaobserwował ją i opisał. W
normalnych warunkach jest to organizm symbiotyczny, ponieważ wnosi pewien wkład w
nasze odżywianie się: rozkłada w jelitach niektóre białka (m.in. kolagenowe) do stanu, w
którym mogą być one wchłonięte przez ścianki jelit (oczywiście samemu się przy tym
posilając).
Twór ten waży tylko około 1 pikograma, czyli jedną bilionową cześć grama (1 pikogram (pg) =
10-15 kilograma) z czego około 70 procent stanowi woda i przypomina krótki walec średnicy
jednego mikrometra (jedna tysięczna milimetra, 1 mikrometr (μm) = 10-6 m) i długości około
dwóch mikrometrów.
Niektóre szczepy posiadają narządy lokomocyjne i potrafią się poruszać, niektóre zaś nie.
Kiedy bakterie wyposażone w systemy lokomocyjne rozmnażają się w bogatym w pokarm
środowisku (takim jak roztwór soli z dodatkiem mieszanki aminokwasów i cukrów) potrafią
one w czasie jednej sekundy przepłynąć odległość równą około 30-40 długościom swojego
ciała (nie licząc długości wici). [3] Ruch ten odbywa się zasadniczo w osi długiej ciała bakterii,
ale odcinki ruchu prostolinijnego są przerywane nagłymi zwrotami, zwykle o ponad 60-90
stopni. Przerwy w ruchy prostolinijnym zdarzają się u E. coli średnio co 10 sekund.
Ruch umożliwia bakterii skorelowana praca zestawu silników, każdy o średnicy tylko 45
nanometrów (cztery milionowe części milimetra) i wysokości 60 nanometrów (1 nanometr
(nm) = 10-9 m). Każdy silnik porusza długą, spiralną wić, kilkakrotnie dłuższą od ciała
bakterii. Bakteria posiada zespół czujników biochemicznych (chemoreceptorów),
rejestrujących różnice stężenia pokarmu w otoczeniu. Kierując się wskazaniami tych
sensorów, bakteria może modulować pracę silników kierując się dzięki temu w obszar
bogatszy w odżywcze składniki. Oprócz chemoreceptorów, silników i wici, standardowe
wyposażenie E. coli stanowią „prędkościomierz” i „skrzynia biegów”, pozwalająca na różne
tryby pracy silnika.
Układ napędowy (flagellum) bakterii składa się z 3 elementów:
rotacyjnego silnika protonowego o odwracalnym ciągu osadzonego w ścianie komórkowej
(jego wewnętrzna część umieszczona jest w cytoplazmie, zewnętrzna w zewnętrznej błonie
komórkowej);

krótkiego haczyka (kątnika), który pełni funkcję elastycznego łącza;
długiej spiralnej wici.
Obserwacje nad pałeczką jelitową Salmonella typhimurium, należącą do tej samej rodziny co
bakteria Escherichia coli, wykazały, że kątnik jest strukturą elastyczną, zbudowaną z
jednakowych elementów białkowych o masie cząsteczkowej 42 000, natomiast wić jest
praktycznie sztywnym tworem przypominającym, pod względem kształtu, spiralę korkociągu.
Jeden zwój sprali ma długość dwóch mikrometrów, czyli długość ciała bakterii, a długość całej
wici jest 8-10 razy większa od długości ciała bakterii. [4] Średnica zwoju wynosi około 0,2
mikrometra, zaś grubość wici 0,02 mikrometra (20 nm). Wić zbudowana jest z białka zwanego
flagelliną. Na jeden zwój spirali wici przypada około pięciu tysięcy cząsteczek flagelliny.
Napęd do wici dostarczają każdej bakterii molekularne silniki w liczbie od 4 do 6, bo tyle ich
zazwyczaj posiada E. coli, napędzane strumieniem protonów. Obroty generowane są
pomiędzy statorem sztywno osadzonym w ścianie komórkowej (peptydoglikanowej) a rotorem
połączonym z wicią. Silnik jest napędzany przez strumień protonów (jonów wodoru)
płynących z zewnątrz do wnętrza bakterii (z wyjątkiem bakterii morskich i bakterii żyjących
w wysokim pH, które zamiast protonów używają jonów sodu). Źródłem energii jest
międzymembranowy potencjał elektryczny — gradient elektro-dodatni, tzw. gradient
chemiosmotyczny ΔμH+. [5]
Jak każdy techniczny silnik elektryczny, tak i silnik bakteryjny posiada rotor, stator i łożyska
(zob. rys. 2). Ośka (wał napędowy), ustawiona pionowo do powierzchni błony komórkowej,
zabudowana jest pomiędzy dwie sąsiadujące membrany w błonie komórkowej. Środkowa
membrana spełnia rolę dialektyka (izolatora) kondensatora, który naładowany jest na
zewnątrz dodatnio, a wewnątrz — ujemnie. Powstające przy tym napięcie elektryczne wynosi
0,2 wolta. W odróżnieniu od innych molekularnych maszyn napędzanych hydrolizą ATP
(adenozynotrójfosforanu), bakteryjny silnik napędzany jest bezpośrednio przez gradient
elektrochemiczny. Specjalny system molekularnych pomp, wykorzystując złożone
metaboliczne procesy, wypompowuje protony z wnętrza komórki „pod prąd” ciśnienia
osmotycznego, wytwarzając w ten sposób wspomniany gradient elektro-dodatni. Powstająca
w ten sposób różnica potencjałów z powrotem przyciąga protony do wnętrza komórki, co
wykorzystuje bakteria do napędzania nimi silników. [6]
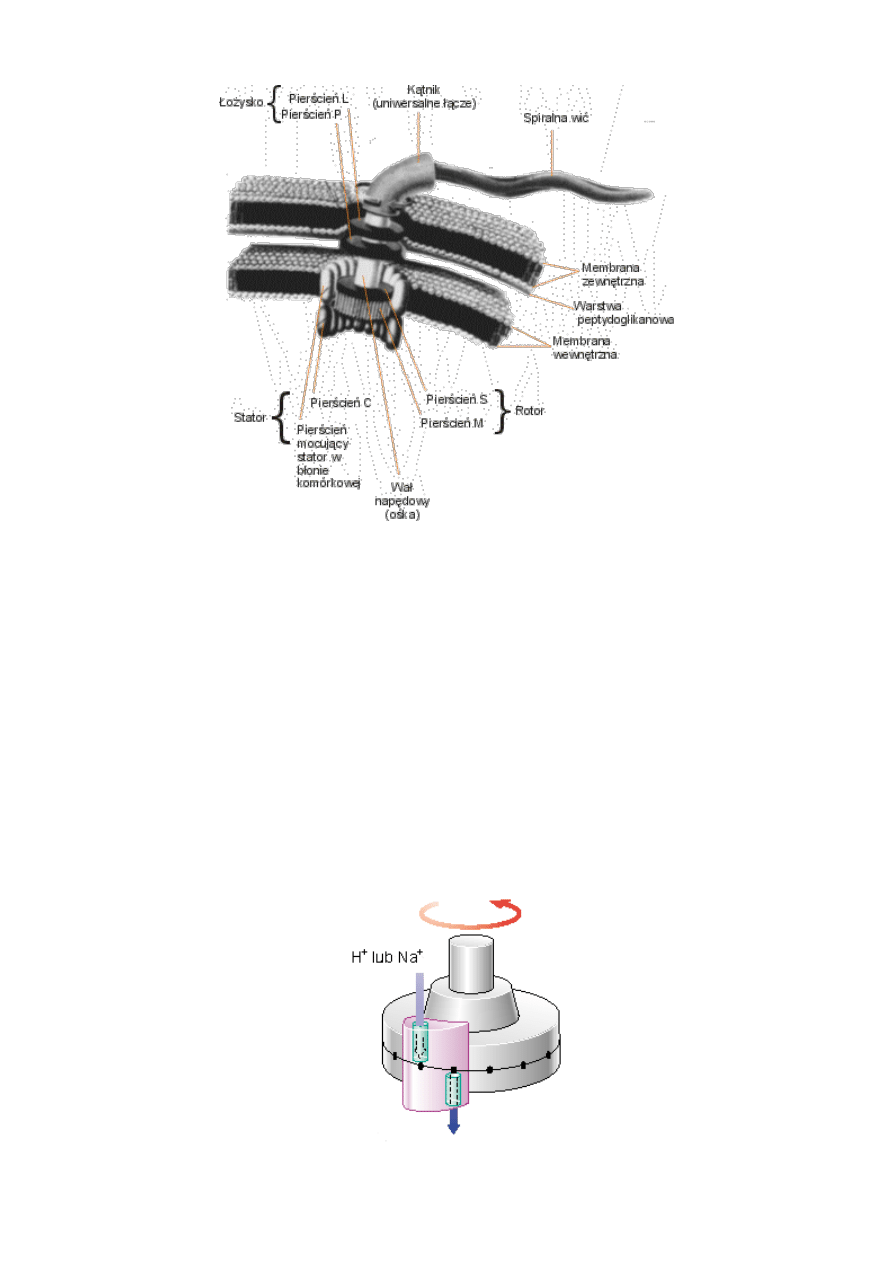
Rys. 2. Schemat bakteryjnego silnika protonowego.
Mechanizm generowania obrotów rotora jest słabo poznany. Wiadomo jednak, że
odpowiadają za to złożone interakcje pomiędzy nim a statorem. [7] Istnieją trzy modele
tłumaczące ten machanizm: model turbiny wodnej, model „kołowrotka” i model
elektrostatycznej turbiny protonowej.
Według modelu turbiny wodnej, protony lub jony sodu płynące na powierzchni białek statora
MotA i MotB wywołują kierunkowy ruch cząsteczek wody, który zmienia sferyczny kształt
tych białek, dzięki czemu wywierają one nacisk na rotor w ten sposób go napędzając. [8]
Model „kołowrotka” zakłada, że protony/jony sodu wpływają do specjalnego tunelu w
białkach kompleksu statora MotA i MotB i kierowane są do specyficznych komponentów
rotora. Wymusza to ruch rotora, który dalej przekazuje te protony/jony sodu do następnego
tunelu w statorze, skąd płyną one już do cytoplazmy (zob. rys. 3).
Rys. 3. Model „kołowrotka”. Zakłada on, że strumień protonów lub jonów sodu płynie przez
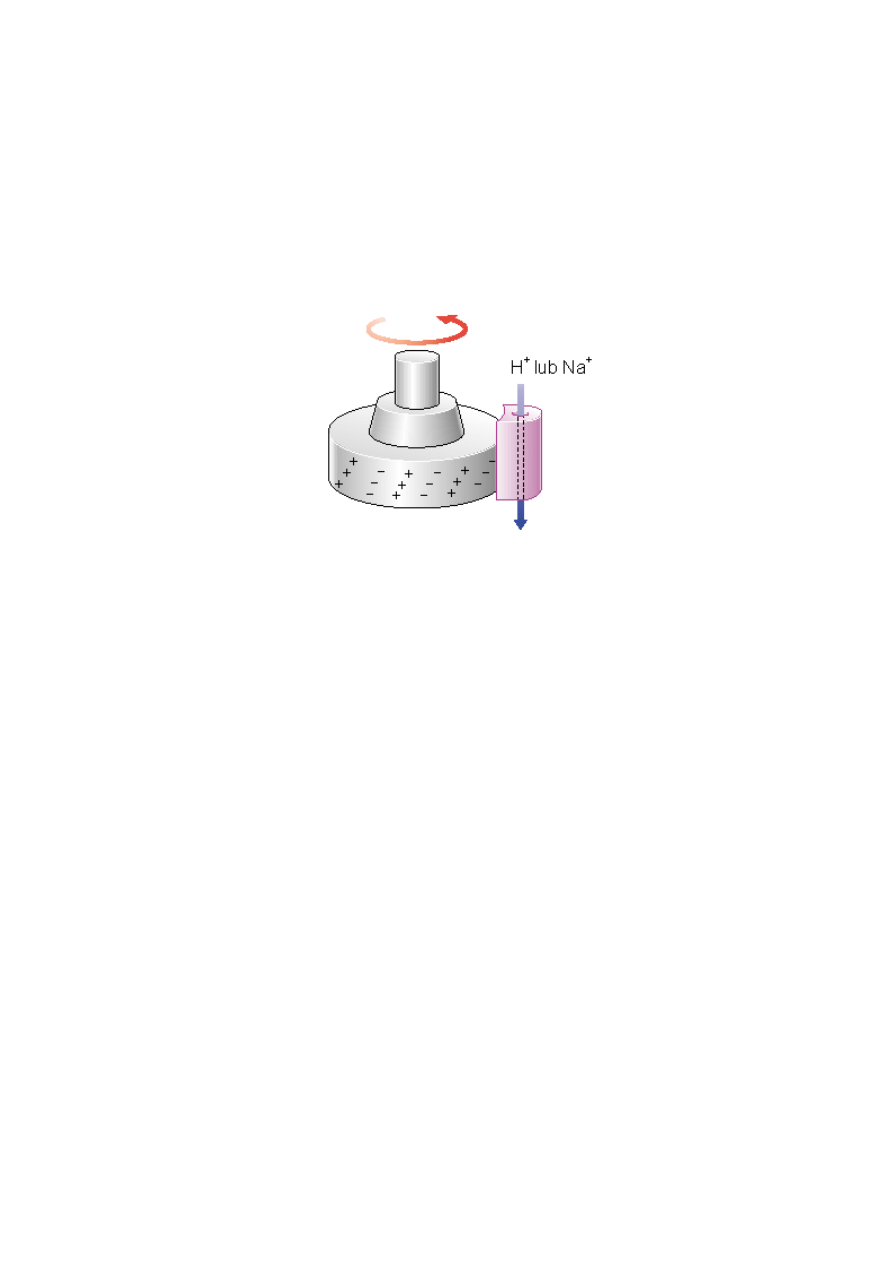
specjalny tunel w białku statora, nastpnie przepływa przez część rotora napędzając go w ten
sposób i sąsiednim tunelem w statorze spływa do wnętrza komórki.
Model elektrostatycznej turbiny protonowej przewiduje, że protony/jony sodu przepływające
przez tunel w białkach statora MotA i MotB oddziałują z ładunkami elektrostatycznymi
precyzyjnie rozmieszczonymi na tworzących pierścień rotora cząsteczkach białka FliG,
stwarzając w ten sposób dynamiczne pole elektrostatyczne, które napędza rotor (zob. rys. 4).
[9] Wydaje się, że na dzień dzisiejszy ten właśnie model jest wśród badaczy najbardziej
popularny.
Rys. 4. Przypuszczalny model generowania obrotów rotora w wyniku elektrostatycznego
oddziaływania pomiędzy strumieniem protonów lub jonów sodu przepływających przez tunel
w białku statora MotA i MotB a ładunkami na powierzchni białkia rotora FliG.
Niezależnie od tego, który model jest poprawny, obliczono, że jeden obrót silnika wymaga
przepływu około tysiąca protonów. Obroty rotora napędzają ośkę, która z kolei przenosi
obroty przez błonę komórkową i poprzez kątnik (uniwersalne łącze) nadaje ruch wici.
Silniki bakterii dają ciąg zarówno do przodu jak i wstecz na kilku trybach pracy z prędkością
od 6000 do 17 000 obrotów na minutę, ale u niektórych morskich bakterii szacuje się, że
prędkość ta wynosi ponad 100 000 obrotów na minutę. [10] Silnik ten potrafi niezwykle
szybko, w czasie ćwiartki swojego obrotu, odwrócić kierunek swojej rotacji. Działa on z
niezwykłą wydajnością — zużycie przez niego energii jest stosunkowo niewielkie. Dla Bacillus
subtilis, pałeczkowatej bakterii wyposażonej w pojedynczą wić, obliczono, że gdy porusza się
ona z prędkością dziesięciu mikrometrów na sekundę, wydatek energii odpowiada hydrolizie
około 100 cząsteczek ATP na sekundę. Straty wolnej energii muszą być w tym procesie —
zwłaszcza w obrębie wnętrza komórki — bardzo małe, bowiem energia rozpraszana
musiałaby niekorzystnie wpływać na przebieg innych procesów chemicznych i na trwałość
delikatnych struktur molekularnych.
Nanotechnologia w praktyce — ciąg dalszy
Nie od rzeczy będzie wspomnieć o nadzwyczajnych właściwościach białka flagelliny, z którego
zbudowana jest wić. Cząsteczki tego białka (oznaczonego jako FliC) o masie cząsteczkowej 54
000 mają zdolność do łączenia się razem w regularne struktury podobne do nici i spirali o
różnym promieniu krzywizny. Jest to proces przypominający krystalizację. Bakteria buduje z
flagelliny zawsze ściśle określoną formę sztywnej, skręconej w kształt korkociągu rureczki.
Podczas montażu wici powstaje pytanie w jaki sposób tysiące cząsteczek flagelliny
dostarczane są do nowo powstającej i wydłużającej się wici (a musi być to mechanizm
niezwykle wydajny, ponieważ jeden skręt spirali wici składa się z około 5000 cząsteczek tego

białka).
Otóż flagellina transportowana jest na koniec aktualnie montowanego odcinka poprzez
specjalny kanał wewnątrz budowanej wici w tempie około 50 cząsteczek flagelliny na sekundę.
Proces budowy wici wieńczy umieszczenie na jej końcu specjalnej nasadki. Utworzona
ostatecznie wić jest strukturą prawo- lub lewoskrętną, przy czym zmiana „skrętności” wici
zachodzi przy każdej zmianie kierunku obrotów silnika obracającego wić. Przy obrotach
silniczka zgodnego z ruchem wskazówek zegara wić wykazuje prawoskrętność i odwrotnie,
przy obrotach w przeciwnym kierunku wykazuje lewoskrętność. Wić nadająca bakterii ruch
postępowy w płynnym środowisku jest zatem odpowiednikiem śruby okrętowej lub śmigła.
Wszystkie te właściwości wici narzucają szczególne wymagania wobec materiału, z którego
jest ona zbudowana. [11]
Proces budowy silnika jest przykładem zdumiewającej precyzji wymagającym skorelowanej,
wieloaspektowej dynamiki. Do zmontowania całego funkcjonalnego silnika bakteria
potrzebuje około 30-40 genów i ich produktów. Są to grupy genów oznaczone jako fla (np.: flg,
flh, fli lub flj) i mot, ponieważ kodują one białka potrzebne do konstrukcji silnika i
poruszania się bakterii — flagella i motility. Defekt jakiegokolwiek z tych genów powoduje
zawsze kompletne załamanie się podstawowej funkcji silnika. Produkty tych genów — białka
o zróżnicowanej masie cząsteczkowej od 8000 do 76 000 — oznaczane są analogicznie do
kodujących je genów jako np.: FlgE, FlgH, FliI, lub MotA, MotB (zob. rys. 5).
We właściwych momentach włączane są odpowiednie geny syntetyzujące właściwe białka, po
czym białka te są odpowiednio transportowane i montowane. Budowa tego precyzyjnie
zintegrowanego zespołu napędowego rozpoczyna się od zmontowania najbardziej
wewnętrznego (względem błony komórkowej) kręgu rotora, następnie powstaje ośka, dalej
kolejne kręgi rotora, składający się przypuszczalnie z ośmiu elementów stator, następnie
kątnik (uniwersalne elastyczne łącze) przez który z kolei budowana jest z tysięcy cząsteczek
flagelliny wić. Materiał do budowy wici transportowany jest poprzez kanał przebiegający
przez centrum kątnika, ośki i dalej na zewnątrz, środkiem rurkowatej wici. U wejścia do tego
kanału znajduje się specjalne urządzenie, regulujące dopływ tego materiału (zob. rys. 5). [12]
Aby uświadomić sobie szybkość z jaką powstaje ta molekularna maszyna, wystarczy
powiedzieć, że cała bakteria E. coli powstaje w czasie nie dłuższym niż 1200 sekund.
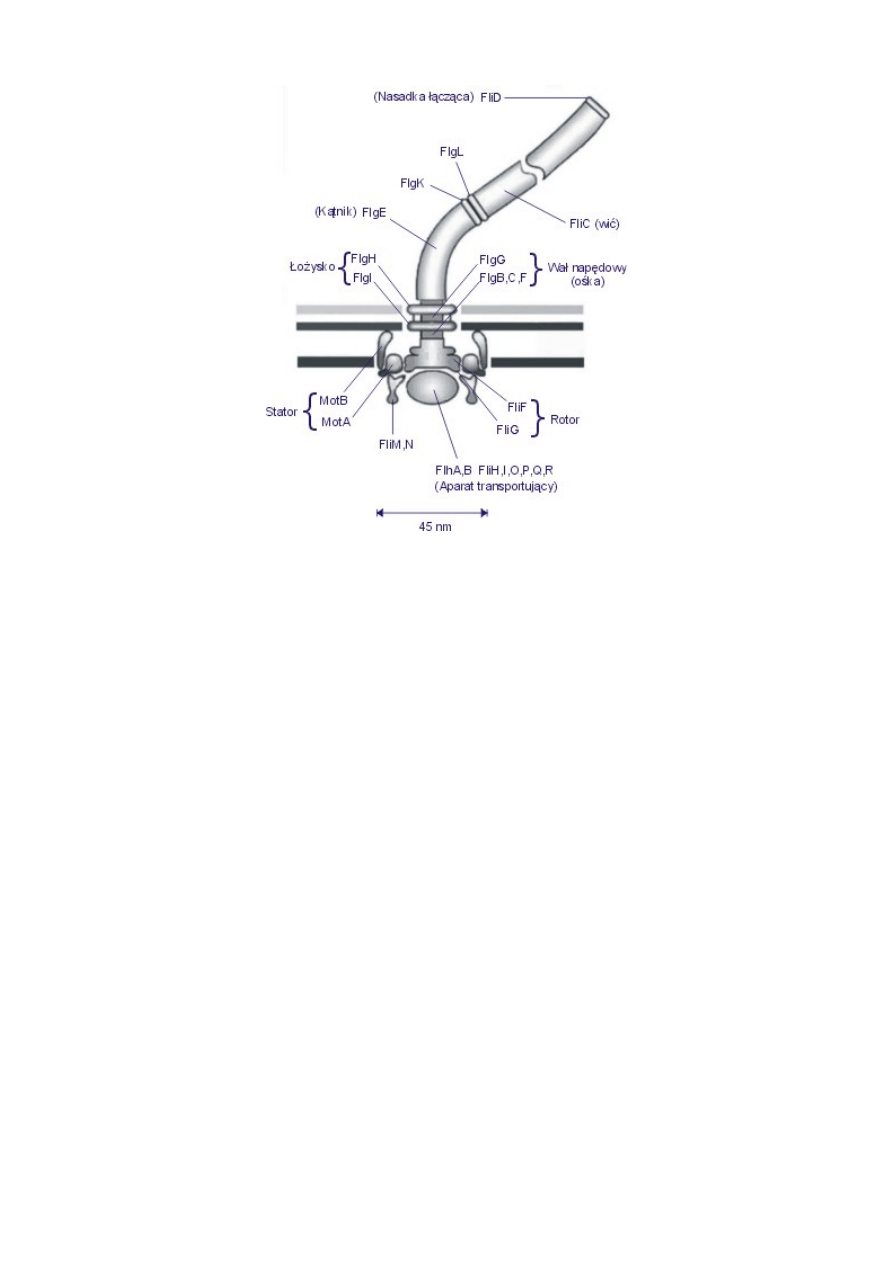
Rys. 5. Koncepcyjny schemat bakteryjnego silnika protonowego E. coli z oznaczeniem
białkowych kompleksów składających się na ten układ. Białka FliF i FliG tworzą —
składające się na rotor — pierścienie S i M z rysunku 2. Pierścienie statora tworzą białka
MotA, MotB i FliM,N, a pierścienie łożyska L i P — białka FlgH i FlgI. Inne białka z grupy
Flg tworzą ośkę (wał napędowy), kątnik i specjalną nasadkę mocującą do elastycznego
kątnika długą spiralną wić. Aparat transportujący to specjalne urządzenie regulujące dopływ
poszczególnych podzespołów koniecznych do zmontowania silnika i wici.
Natomiast cały genom E. coli składa się z podwójnej nici polimeru DNA około 700 razy
dłuższego od ciała bakterii. Bakteryjne DNA zawiera 4 639 221 par zasad, które składają się
na 4 288 genów, z których większość koduje białka. Dotychczas poznano funkcję tylko około
60 procent z tych białek. Około 50 różnego rodzaju białek jest wymaganych do montowania
układu nawigacyjnego i lokomocyjnego bakterii pozwalających jej poruszać się w kierunku
chemicznie atrakcyjnego środowiska i oddalać się od nieatrakcyjnego (chemotaxis).
Tak długa — w stosunku do ciała bakterii — helisa DNA narzuca specjalne wymagania co do
jej czysto mechanicznego upakowania wewnątrz bakteryjnej komórki, przy konieczności
szybkiego pobierania z niej potrzebnej informacji. Odcinki DNA, na których zakodowane są
informacje niezbędne do prawidłowego skonstruowania elementów silnika, wici i systemu
nawigacji rozrzucone są w różnych regionach nici DNA. Odnalezienie tych informacji
wewnątrz podwójnej helisy DNA jest problemem niezwykle skomplikowanym i choć komórka
radzi sobie z tym doskonale, to jednak mechanizm wyszukiwania i selekcjonowania tych
informacji pozostaje nadal w dużej mierze zagadką. Wystarczy wspomnieć, że chemiczna
struktura DNA pozwala odczytywać szyfr molekularny na sześć różnych sposobów, z których
zwykle tylko jeden jest prawidłowy.
Wykryto dotąd wiele elementów kilkupiętrowego systemu sygnałów chemicznych koniecznych
do precyzyjnego odczytywania informacji, ukrytej wewnątrz DNA. Równocześnie wykryto
wiele modyfikacji, jakim wstępna, „surowa” informacja — wydobyta z DNA — jest
poddawana, zanim osiągnie poziom wystarczający do sterowania pewnymi etapami rozwoju.

[13]
Na osobne potraktowanie zasługuje układ nawigacyjny wyznaczający kierunek ruchu
bakterii. Układ ten reaguje na różne na różnego rodzaju bodźce chemiczne dzięki specjalnym
cząsteczkom białka odgrywającym rolę chemoreceptorów. Wyróżniono trzy zasadnicze
sposoby lokomocji bakterii:
1. Lokomocja chaotyczna. W jednorodnym środowisku bogatym w odżywcze składniki,
lokomocja bakterii przypomina chaotyczne ruchy w trójwymiarowej przestrzeni. Bakteria co
kilka sekund osobno włącza swoje silniki (wić porusza się wtedy zgodnie z ruchem wskazówek
zegara), ale tylko na jedną dziesiątą część sekundy, dzięki czemu bakteria zmienia kierunek
poruszania się o 60-90 stopni („koziołkuje”). Wykorzystuje do tego jednak nie siłę
bezwładności. Przy rozmiarach i wadze E. coli jej lokomocja zdominowana jest nie przez siłę
bezwładności, ale lepkość cieczy, w której się porusza. Bakteria wykorzystuje fakt, że na
poprzecznie ułożoną względem dłuższej osi ciała bakterii wić działa około dwukrotnie większa
siła lepkości niż na tę ułożoną wzdłuż ciała, co pozwala wykorzystać tę pierwszą jako swoisty
„hamulec” odkształcający kierunek poruszania się komórki. [14]
Za następnych kilka sekund bakteria ponownie na krótką chwilę uruchamia silniki, ale tym
razem na wstecznym ciągu, czyli wici obracają się przeciwnie do wskazówek zegara, co znowu
pozwala bakterii „przekoziołkować” i zupełnie zmienić kierunek poruszania się.
2. Lokomocja przeszukująca. Jeśli jednak sensory E. coli wykryją rosnące w określonym
kierunku stężenie odżywczych składników, wtedy jej częste „koziołkowanie” jest natychmiast
wygaszane. Silniki przełączają się na inny, bardziej jednostajny tryb pracy (rotacja przeciwna
do ruchów wskazówek zegara) kierując w ten sposób bakteryjną komórkę w kierunku
bogatszego źródła odżywczych składników. Chaotyczny „wypas” zamienia się w zygzakowaty
kurs w określonym kierunku (bakteria korektuje go dzięki możliwości dwukierunkowej pracy
silników — patrz niżej).
3. Lokomocja ucieczki. Ten sposób lokomocji jest obserwowany, gdy bakteria wykryje w
otoczeniu pojawienie się szkodliwych substancji. W tej sytuacji bakteria szybko oddala się od
obszaru koncentracji takich składników.
Rys. 6. „Ten mikroorganizm jest marzeniem nanotechnologów” — Howard Berg, profesor
biologii molekularnej i komórkowej oraz fizyki na Uniwersytecie w Harvardzie.
Molekularna powielarka w praktyce
Ale Escherichia coli to nie tylko układy nawigacyjne i lokomocyjne. To także zdumiewająca
powielarka potrafiąca kopiować samą siebie z szokującą wydajnością. Bakteria ta
umieszczona w środowisku bogatym w pokarm i w temperaturze ludzkiego jelita (37 stopni C)
może syntetyzować i replikować wszystko co potrzebuje do zrobienia swojej nowej kopii w
czasie 1200 sekund. Tak więc, jeśli zaczniemy dziś o północy z jedną bakterią (i dużą ilością
pokarmu) jutro o północy będzie 272, albo inaczej 4.7×1021 bakterii — co wystarczy do
zapełnienia nimi kwadratowego pudła o długości boku 17 metrów!
Ta szybkość replikacji wyjaśnia dlaczego potomstwo jednej bakterii umieszczonej na twardej
odżywce natychmiast rozprasza się na jej powierzchni — w przeciwnym razie wkrótce na
powierzchni odżywki wyrósłby kopiec bakterii wielkości jednego milimetra.
Pojedyncza komórka E. coli buduje w sobie drugą, identyczną z pierwszą, z identycznym
wyposażeniem. Potem dochodzi do rozdzielenia się w połowie tych dwóch kompletnych
komórek. Trudno powiedzieć, która jest „matką” a która „córką”. Obie są jednakowo
„młode”. Ich cząsteczki DNA są identyczne (za wyjątkiem mutacji, które spontanicznie
zdarzają się dla danego genu w tempie około 10-7 na generację) po połowie (wzdłuż) stare, a
po połowie nowo wybudowane. Zatem proces rozmnażania produkuje dwie komórki o tym
samym „wieku”. Gdy one się dzielą to znowu powstają cztery komórki o tym samym „wieku”.

W pewnym więc sensie E. coli jest więc „nieśmiertelna” ponieważ macierzysta komórka dzieli
się na dwie potomne, doskonale identyczne z macierzystą, a te na kolejne, także identyczne.
Dopóki ten szczep bakterii zachowa się w jednym choćby osobniku, to ten osobnik jest tak
„stary”, jak pierwsza komórka tego gatunku.
W świetle powyższego zrozumiała jest opinia profesora biologii molekularnej i fizyki na
Uniwersytecie w Harvardzie, Howarda Berga:
Ten mikroorganizm jest marzeniem nanotechnologów. [15]
Projekt bez projektanta?
Maszyną, zgodnie z definicją jaką podaje sześciotomowa Encyklopedia PWN, jest:
„Urządzenie techniczne zawierające mechanizm lub zespół połączonych ze sobą
mechanizmów, służące do przetwarzania energii lub wykonywania określonej pracy
mechanicznej; z energetycznego punktu widzenia maszyna jest przetwornicą energii
przekształcającą w energię mechaniczną inny jej rodzaj.” [16]
Czy biomolekularne systemy w świetle powyższej definicji są maszynami? Oczywiście tak.
Otwarcie potwierdzają to biochemicy. Przykładowo w lutym 1998 r. ukazało się specjalne
wydanie czołowego biochemiczngo czasopisma Cell poświęcone „makromolekularnym
maszynom”. Dowiedzieć się z niego możemy, że wszystkie komórki używają kompleksowych
molekularnych maszyn do obróbki informacji, przemiany energii, przyswajania składników
odżywczych i transportowania materiałow przez błony komórkowe. Bruce Alberts, prezydent
amerykańskiej National Academy of Sciences napisał wprowadzenie do tego specjalnego
wydania zatytułowane „The Cell as a Collection of Protein Machines: Preparing the Next
Generation of Molecular Biologists” (Komórka jako zbiór białkowych maszyn:
przygotowując następną generację biologów molekularnych). Stwierdził w nim m.in.:
„Zawsze niedocenialiśmy komórki. (…) Cała komórka może być przedstawiona jako fabryka
zawierającą skomplikowaną sieć połączonych linii montażowych, każda z nich złożona z
układu dużych białkowych maszyn. (…) Dlaczego możemy nazywać duże białka montażowe
tkwiące u podstaw komórkowych funkcji maszynami? Ponieważ dokładnie tak, jak maszyny
wynalezione przez ludzi do efektywnego działania w makroskopowym świecie, te montażowe
białka zawierają precyzyjnie zintegrowane ruchome części. „[17]
Prawie sześć lat później, w grudniu 2003 r., inny fachowy periodyk, BioEssays, opublikował
własne wydanie dotyczące „molekularnych maszyn”. We wprowadzeniu do niego Adam
Wilkins, redaktor BioEssays wyjaśnia:
„Artykuły w tym wydaniu ukazują niektóre pewne uderzające podobieństwa pomiędzy
sztucznymi a biologicznymi/molekularnymi maszynami. Przede wszystkim molekularne
maszyny, tak jak maszyny robione przez człowieka, wykonują dokładnie określone funkcje.
Po drugie, makromolekularne maszyny składają się z wielu części, oddziałujących ze sobą w
odrębny i precyzyjny sposób. Po trzecie, wiele z tych maszyn posiada cześci, które mogą być
użyte w innych molekularnych maszynach (przynajmniej z niewielkimi modyfikacjami), co
jest porównywalne z wymienialnymi częściami w sztucznych maszynach. W końcu, co
najważniejsze, posiadają one podstawową cechę maszyn: zamieniają energię w jakąś formę
„pracy”. „[18]
W przypadku protonowego silnika Escherichii coli truizmem jest stwierdzenie, że jest on
molekularną maszyną. Ten truizm niesie jednak bardzo poważne implikacje. Wszystko
bowiem co wiemy o technicznych maszynach mówi nam, że w ich powstanie musi być
zaangażowana inteligencja. Ateistyczny filozof Paul Ricci w swojej książce Fundamentals of
Critical Thinking napisał, że: „prawdą jest, że każdy projekt ma projektanta”, co więcej
twierdzenie „każdy projekt ma projektanta” jest analitycznie twierdzeniem prawdziwym i nie
wymaga formalnego dowodu. [19] Gdzie jest projekt, tam musi być — z definicji —

projektant. Ewolucjoniści doskonale zdają sobie sprawę z tej niebezpiecznej dla nich sytuacji.
Jak wyjaśnia Francisco Ayala, znany biolog ewolucyjny, były przewodniczący American
Association for the Advancement of Science:
„Funkcjonalny projekt organizmów i ich cech może zatem zdawać się być argumentem za
istnieniem projektanta.” [20]
Jednak postulowanie rozumnego projektu przy wyjaśnianiu genezy bakteryjnego silnika, czy
innych molekularnych maszyn jest dla większości biologów ciężką herezją. Ze względu na
przyjęte wcześniej filozoficzne założenia zmuszeni są oni sięgać lewą ręką do prawego ucha,
czyli a priori wykluczyć rozumny projekt i próbować wyjaśniać powstanie układu
lokomocyjnego E. coli w wyniku naturalistycznej ewolucji. Rozwijają w ten sposób
zaskakującą swoją głębią ewolucyjną myśl — że projekt nie musi mieć projektanta. Podkreśla
to cytowany powyżej Ayala:
„Największym dokonaniem Darwina było pokazanie, że organizacja życia może zostać
wyjaśniona jako rezultat naturalnych procesów, doboru naturalnego, bez jakiejkolwiek
potrzeby odwoływania się do Stwórcy lub innych zewnętrznych czynników.” [21]
Według ewolucjonistów są więc dwa rodzaje projektów: projekt z projektantem i projekt bez
projektanta. Biologiczne maszyny mają się plasować w tej drugiej kategorii — projektów bez
inteligentnego projektanta.
„Takie sobie bajeczki” w praktyce
W uzasadnianiu tej wiekopomnej idei służy ewolucjonistom specyficzna logika rozumowania,
zwana ewolucjonistyczną logiką ślepej wiary. William Dembski wyjaśnia, co kryje się pod tym
tajemniczym pojęciem:
Ewolucjonistyczna logika jest formą sprowadzania do absurdu (reductio ad absurdum).
Niedorzecznością jest sugerowanie inteligentnego projektu, czy w ogóle jakiejkolwiek formy
teleologii. Dla ewolucyjnych biologów potraktowanie projektu lub teleologii jako
fundamentalnego sposobu wyjaśniania, zdolnego do wytłumaczenia powstania biologicznych
struktur jest absolutnie nie do zaakceptowania. Przy takim założeniu każdy przekonujący
argument wskazujący na projekt musi wywodzić się z błędnych przesłanek. Tak więc każde
twierdzenie, które pociąga za sobą, czyni prawdopodobnym lub w jakikolwiek sposób przy
wyjaśnianiu powstania biologicznych struktur implikuje projekt, musi zostać odrzucone.


„Ale ewolucjonistyczna logika nie poprzestaje na tym. Nie tylko każde twierdzenie
wskazujące na projekt musi zostać odrzucone, ale — skutkiem tego — każde twierdzenie,
które wyklucza projekt nakazane jest do wierzenia i wymaga uznania. To jest właśnie
ewolucjonistyczna logika ślepej wiary — wiara w ewolucjonistyczne wyjaśnienia jest
nakazana nie dlatego, że wiara ta ma oparcie w aktualnym materiale dowodowym, ale po
prostu dlatego, że pozwala ona wykluczyć projekt.” [22]
Kiedy Michael Behe spopularyzował argument z nieredukowalnej złożoności, niektórzy
ewolucjoniści przyznali, że ta koncepcja przysłużyła się nauce przez to, że zmusza ich do
zmierzenia się z następującym problemem: w jaki sposób układy nieredukowalnie złożone
mogły powstać w wyniku stopniowej ewolucji, skoro do ich funkcjonowania konieczne są
wszystkie podzespoły? Potwierdzają to ewolucyjni biologowie Thornhill i Ussery:
„Jednak dokładnie nie przebadano bardziej teoretycznej kwestii, czy darwinowska ewolucja
jest w stanie zbudować nieredukowalnie złożone struktury, posiadające funkcjonalnie
niepodzielne składniki (o ile takie struktury istnieją). (…) Czynnikiem hamującym badanie
możliwości skonstruowania takich biologicznych struktur przez darwinowską ewolucję jest
brak klasyfikacji możliwych dróg takiej ewolucji. „[23]
Ewolucjoniści mają tu poważny problem. Jak stwierdził w 1949 r. znany brytyjski
ewolucjonista (i komunista) J.S.B. Haldane, ewolucja nie może nigdy wytwarzać „różnych
mechanizmów takich jak tryby i magnesy, które byłyby bezużyteczne aż do momentu, gdy
osiągną one pełną funkcjonalność”. [24] Jeśli dany biologiczny system, aby mógł
funkcjonować potrzebuje wszystkich podzespołów, to jak taki system powstawał w wyniku
stopniowej ewolucji, skoro zarówno 20 procent takiego systemu jest równie niefunkcjonalne
jak 80 procent? Bajeczka o stopniowym budowaniu takiego systemu przez dodawanie
kolejnych części w tym przypadku odpada — dobór naturalny musiałby wtedy rozwijać
niefunkcjonalne narządy, aż do momentu ich pełnego skompletowania, co by znaczyło, że
dobór ten posiada tajemniczą zdolność przewidywania przyszłości.
Jak wyżej wspomniano, silnik protonowy E. coli do złożenia i funkcjonowania potrzebuje
około 30-40 precyzyjnie zintegrowanych białek oraz dodatkowych 20 asystujących przy jego
budowaniu. Brak któregokolwiek białka wchodzącego w skład silnika, uniemożliwia
zmontowanie sprawnego systemu lokomocji. Oznacza to, że nieredukowalna złożoność
bakteryjnego silnika protonowego jest empirycznie potwierdzonym faktem i system ten
doskonale spełnia kryteria oryginalnej definicji nieredukowalnej złożoności zaproponowanej
przez Behe’ego:
„System nieredukowalnie złożony to pojedynczy system składający się z wielu precyzyjnie
zintegrowanych i oddziałujących ze sobą podzespołów, tworzących podstawową funkcję

systemu, w którym usunięcie jakiegokolwiek z tych podzespołów skutkuje faktycznym
załamaniem się funkcjonowania takiego systemu.” [25]
Pompa rotacyjna do produkcji ATP.

W przypadku bakteryjnego silnika jego podstawową funkcją jest napędzanie bakteryjnej
komórki w wodnym środowisku przez szybkoobrotową wić o odwracalnych obrotach. Warto
tutaj zauważyć, że długa wić i odwracalna rotacja silnika nie są jakimiś nadmiarowymi,
luksusowymi dodatkami, ale koniecznością. Bakteria aby móc poruszać się w cieczy musi
pokonać ruchy Browna cząsteczek zawiesiny w jakiej pływa. Bardziej przypomina to
„przepychanie się” niż pływanie, jakie znamy z własnego doświadczenia. Powód, dla którego
silniki bakterii mają odwracalne obroty jest taki, że ruchy Browna cząsteczek roztworu często
spychają bakterię z właściwego kursu, kiedy płynie ona w kierunku koncentracji odżywczych
składników albo oddala się od szkodliwych. Odwracalny ciąg silników pozwala bakterii w tej
sytuacji szybko zatrzymać się i próbować powrócić na właściwy kurs. Analogicznie w
przypadku „wypasu” odwracalne rotacje pozwalają na chaotyczne przeczesywanie obszaru

bogatego w odżywcze składniki. Aby bakteryjny silnik spełniał swoją podstawową funkcję
musi napędzać wić bardzo szybko i dwukierunkowo.
Ewolucyjni biologowie upierają się jednak, że bakteryjny silnik protonowy musiał pojawić się
wskutek działania naturalistycznych, ewolucyjnych procesów. Świetnie, tylko jak? Bogata
wyobraźnia poszła w ruch, jej produktem jest zwykle następująca historyjka: silnik ten
składa się z części, które wcześniej pełniły inne funkcje i dobór naturalny adaptował te
elementy do kooperacji w nowej funkcji i nowym mechanizmie — bakteryjnym silniku (idea
preadaptacji lub egzaptacji). Fantazje takie snuje John Rennie w Świecie Nauki:
„Sedno w tym, że elementy składające się na wić bakteryjną, którym Behe przypisuje
przydatność wyłącznie jako podzespołom napędowym, mogą pełnić różne funkcje, sprzyjające
jej ewolucji. Ostateczne powstanie wici mogło polegać tylko na nowej kombinacji
skomplikowanych podzespołów.” [26]
Spekulacja ta zakłada, że stopniowo ewoluuje nie tylko sama struktura, ale równolegle z nią
ewoluuje funkcja tej struktury. Ten scenariusz brzmi sensownie tylko do momentu, gdy
próbujemy poznać jego szczegóły. Jedynie dobrze udokumentowane przykłady tego typu
„koewolucji” pochodzą z inżynierii. Przykładowo inżynier-elektronik może, używając
elementów kuchenki mikrofalowej, radia i monitora komputera, zmontować coś w rodzaju
telewizora. Lecz w tym przypadku mamy inteligentny projekt — kogoś działającego według
pewnego planu i ze sporą wiedzą na temat elektrycznych gadżetów. Jednak dobór naturalny
nie wie nic o silniku bakteryjnym, który ma montować. Jak więc taki proces może adaptować
białka pełniące wcześniej zupełnie inne funkcje do wytworzenia bakteryjnego silnika?
Problemem jest tu, że dobór naturalny może selekcjonować funkcje wcześniej istniejące. Dla
przykładu może selekcjonować większe dzioby, które są lepsze do otwierania twardszych
ziaren. Większe dzioby są cały czas są obecne w populacji, a w danych warunkach dobór
naturalny po prostu zaczyna faworyzować ich funkcjonalność. Ciśnienie doboru może nawet
nieco rozszerzyć oryginalną funkcję do wykonywania podobnych zadań, np. przekształcić
dziób przystosowany do otwierania ziaren i wyjadania nasion do zjadania owadów.
Jednak w przypadku genezy bakteryjnego silnika nie możemy mówić o faworyzowaniu już
istniejącej funkcji — bo jej nie ma, rozszerzeniu działania tej funkcji, czy o dostosowaniu się
jakiejś istniejącej pojedynczej struktury do nowej funkcji, ale o skorelowanym przestawieniu
się całego zespołu struktur wcześniej pełniących inne funkcje do nowej struktury pełniącej
nową funkcję.
Ale to jeszcze nie wszystko. Aby neodarwinowski mechanizm wytworzył jakąś nową funkcję
nie wystarczy, by po prostu istniała hipotetyczna sekwencja form pośrednich łącząca starą i
nową funkcję. Każda drobna modyfikacja wiodąca od starej do nowej funkcji musi jeszcze
dawać jakąś korzyść przystosowawczą. W języku biologów, każda taka forma pośrednia
wymaga zwiększenia się dostosowania mierzonego kryteriami przeżywalności i reprodukcji.
Aby dobór naturalny faworyzował takie zmiany, każdy krok w takiej serii drobnych zmian
musi dawać jakiś wzrost przystosowania, dodawać jakąś nową jakość zwiększającą
przeżywalność i zdolność reprodukcyjną.
Dodatkowym warunkiem takiej ewolucji jest niedopuszczalność chwilowego nawet
pogorszenia stopnia przystosowania organizmu. Jest tak nawet wtedy, gdy dzięki temu
możliwe byłoby osiągnięcie korzystnych rezultatów w nieco dalszej perspektywie. Ewolucja
nie planuje długoterminowo. Dobór naturalny zawsze preferuje to, co w danej chwili, tu i
teraz, jest lepsze. To jest główne przesłanie jednej z książek Richarda Dawkinsa Wspinaczka
na szczyt nieprawdopodobieństwa. [27] Roztoczył on w niej wzniosłą wizję ewolucji
wspinającej się na różne szczyty nieprawdopodobieństwa, czyli biologiczne systemy złożone w
rodzaju oka, nie od strony stromizn, ale drobnymi kroczkami poprzez łagodne serpentyny.
W przypadku ewolucji jakiegoś hipotetycznego molekularnego mechanizmu do postaci silnika

bakteryjnego oznacza to, że każda drobna modyfikacja tego mechanizmu nie tylko musiałaby
zachowywać ciągłą funkcjonalność, ale również dawać jakąś korzyść przystosowawczą. Jak to
ujął Karol Darwin, takie ewolucyjne zmiany musiały być „niezmiernie drobne i stopniowe”.
[28] Jedyną więc możliwą drogą ewolucji tego systemu było stopniowe przekształcanie
pierwotnych białkowych struktur, przy czym funkcje tych struktur musiały ewoluować
(koewoluować) wraz z tymi strukturami. Jakie empirycznie testowanle świadectwo
przedstawiają ewolucjoniści na poparcie takiej koewolucji?
Nie ma takiego świadectwa. Według ewolucjonistów ta koewolucja zniknęła w pomrokach
dziejów, dziś możemy co najwyżej snuć bardziej lub mniej fantastyczne jej scenariusze. Cała
moc ewolucyjnej argumentacji sprowadza się do analogii z projektu i do wskazywania
podobieństwa. W pierwszym przypadku chodzi o to, że jedyną solidną analogią takiej
„kooperacyjnej ewolucji” jest świadectwo inżynierii — a więc dziedziny opartej o rozumny
projekt. Jeśli model ma być w tym przypadku dobrym odwzorowaniem rzeczywistości, to
jakim sposobem dobrym odwzorowaniem procesu bezcelowego i bezrozumnego jest proces
oparty na projekcie i z założonym wcześniej celem? Ewolucjoniści opierają się więc na
inteligentnym projekcie, aby krytykować koncepcje oparte o inteligentny projekt. Ot, kolejna
perełka ewolucjonistycznej logiki.[29]
Ćwiczy się w niej biochemik Kenneth Miller z Brown University regularnie praktykując
„ewolucję” pułapki na myszy — aby zilustrować ewolucję układów nieredukowalnie
złożonych. Według niego trzy z pięciu części pułapki na myszy tworzą spinkę do krawata.
Próbując budować ze spinki od krawata pułapkę na myszy, twierdzi on, że podobny proces
może mieć miejsce również w przyrodzie, gdzie te same elementy mogą pojawiać się w
zupełnie różnych funkcjach. Co więcej, uważa on swoją pracę nad zastosowaniem spinek do
krawatów do budowy pułapek na myszy za ogromnie doniosłą dla ewolucyjnej biologii. [30]
Ale niektórzy ewolucjonistyczni bajarze nie kłopoczą się nawet fantazjowaniem na temat
hipotetycznych dróg ewolucji takich mechanizmów. Im wystarczy tylko wskazanie podobnych
elementów pełniących różne funkcje w różnych strukturach i stwierdzenie „a więc to musiało
wyewoluować”. Ewolucja pełni w tym przypadku rolę magicznego, uniwersalnego
wypełniacza, którym zalepia się luki gdzie zupełnie brak empirycznego świadectwa. W ten
sposób maskuje problem cytowany już Rennie:
„Te skomplikowane składniki owego narządu ruchu mają swoje prekursory w przyrodzie, co
wykazali Kenneth R. Miller z Brown University i inni. Wić bardzo przypomina strukturę, za
pomocą której pałeczka dżumy (Yersinia pestis) wstrzykuje do komórki toksyny. „[31]

Podjednostka Fo pompy rotacyjnej do produkcji ATP.

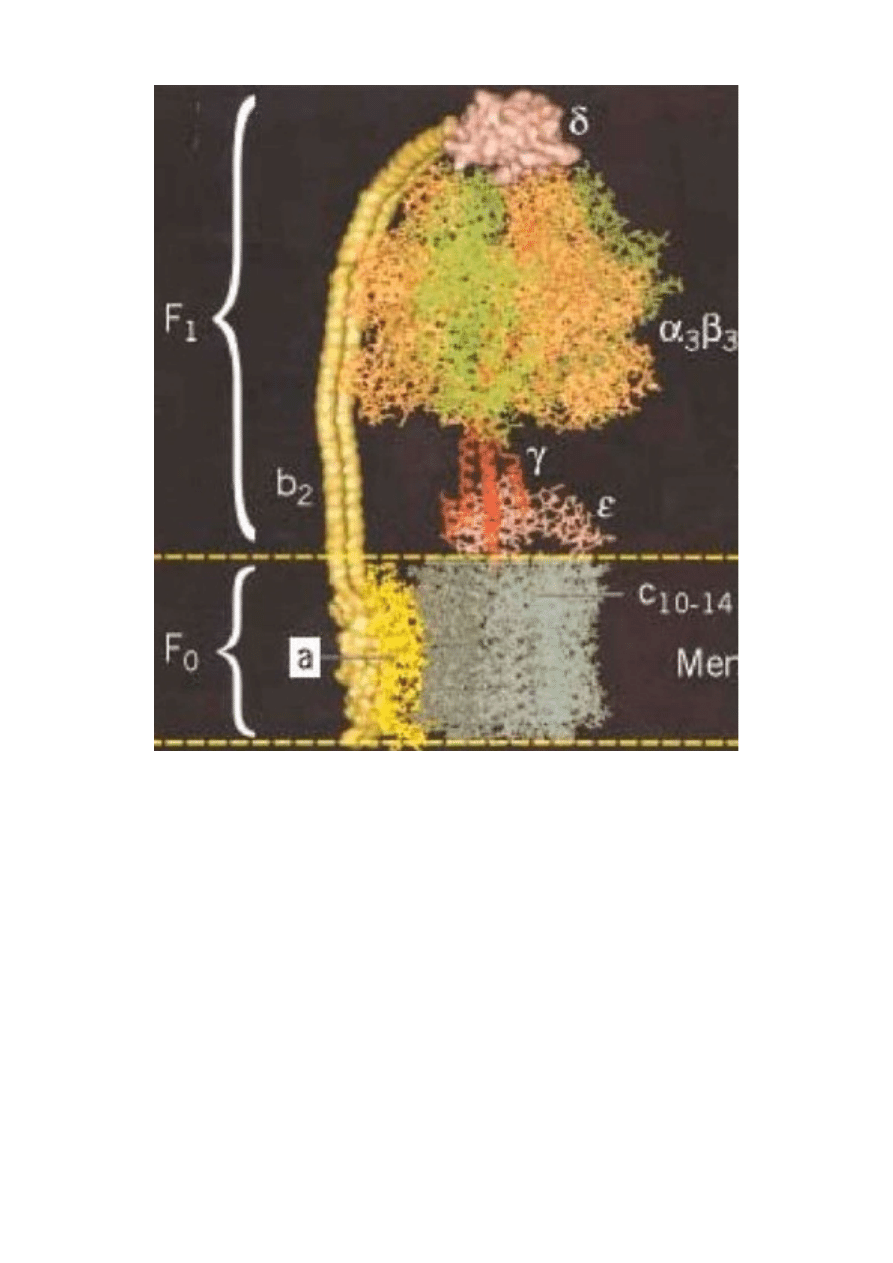
Pompa rotacyjna do produkcji ATP. Podjednostki Fo i F1.
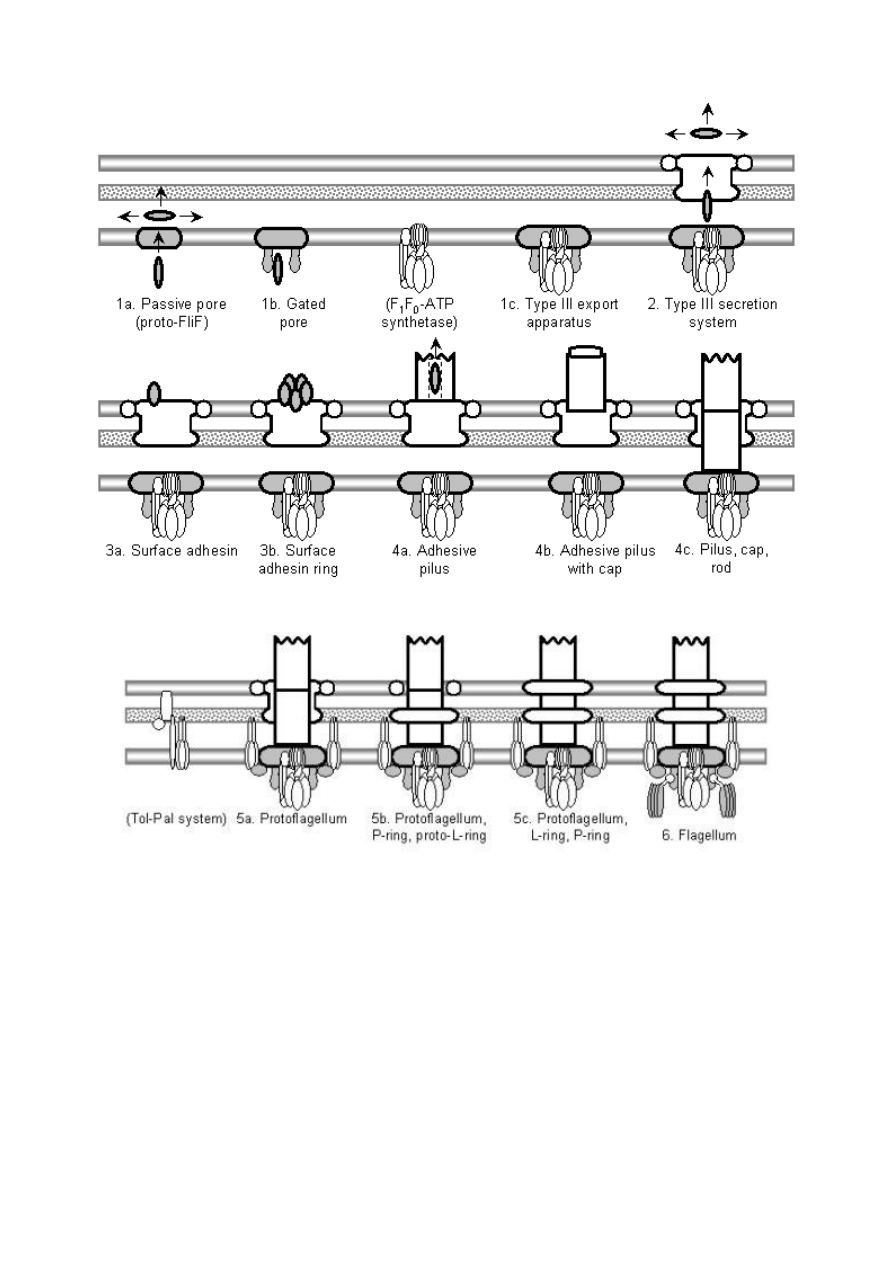
Powyżej schemat koncepcji ewolucji kooptacyjnej (na zasadzie preadaptacji-to jest zmiany
funkcji starego organu) witki bakteryjnej. Pomysł ten zakłada ,że powszechna we wszystkich
żywych organizmach (od najprostszej bakterii po komórki budujące ciało człowieka) pompa
rotacyjna do produkcji ATP, układ jak najbardziej nieredukowalne złożony-choć miał być
„prostszym prekursorem w tej ewolucji”, przekształciła się w układ sekrecyjny typu III (rotacyjny
aparat bakteryjny do infekowania eukariontów), a ten z kolei w wić bakteryjną.
Stwierdzenie, że bakteryjny system lokomocyjny „bardzo przypomina” strukturę
wstrzykującą pałeczki dżumy jest nieco naciągane. Systemy te pełnią zupełnie odmienne
funkcje: silnik protonowy jest narządem lokomocyjnym dostarczającym bakterii napędu w
płynnym środowisku, system wstrzykujący nie ma nic wspólnego z napędem, jest to rodzaj
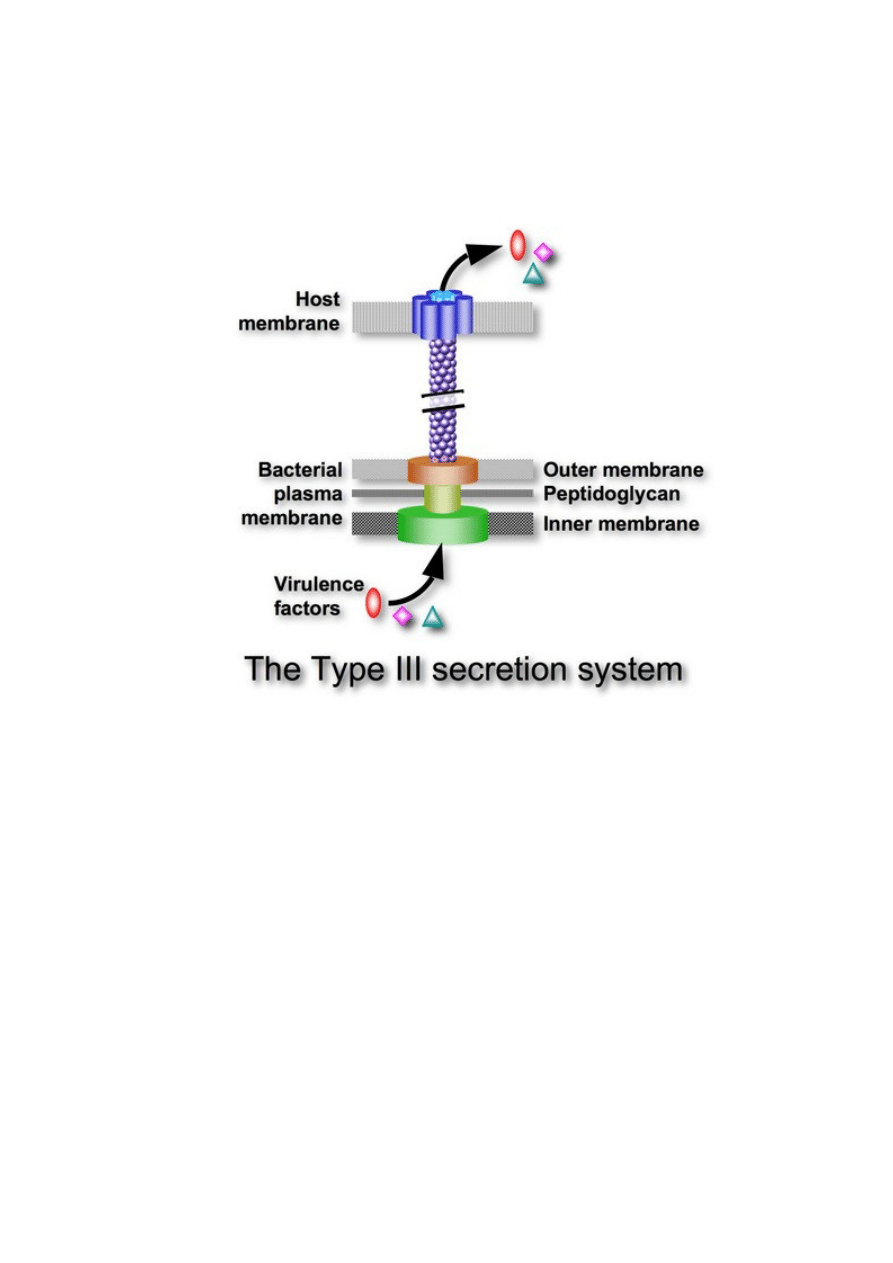
molekularnej pompy służącej do atakowania innych komórek. [32] To „wyraźne
podobieństwo” sprowadza się głównie do tego, że niektóre białka tworzące bakteryjny silnik
protonowy są podobne do niektórych obecnych w strukturze wstrzykującej, gdzie pełnią
zresztą inne funkcje.
Aparat sekrecyjny typu III (aparat wstrzykujący).
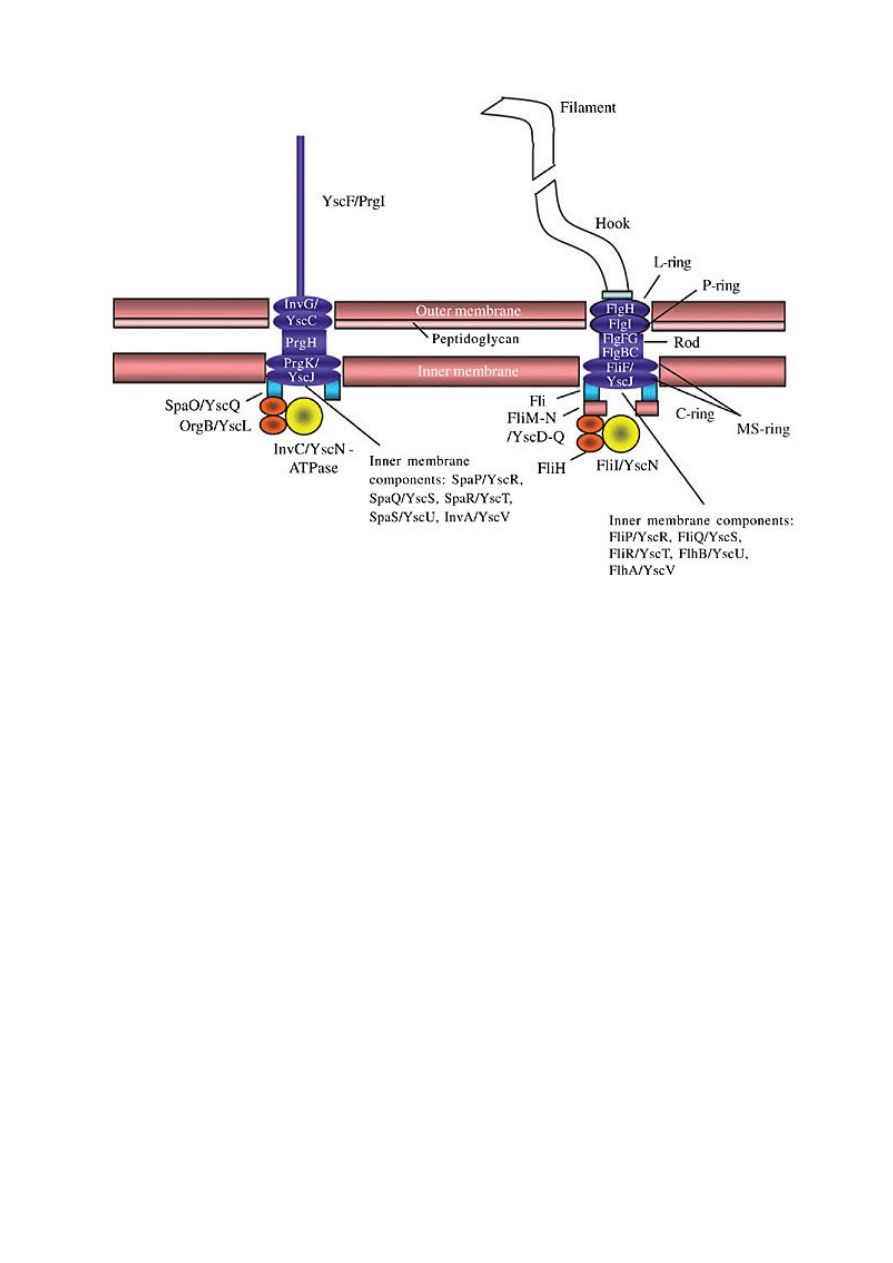
Aparat sekrecyjny typu III i uproszczony schemat wici bakteryjnej.
Najwyraźniej Rennie uznaje, że wystarczy wskazać podobne elementy pełniące różne funkcje
w różnych strukturach, następnie rzucić na nie zaklęcie: „ewolucja” i sprawa rozwiązana.
Dokładnie tak, jakby wskazać łopatki w odkurzaczu i łopatki w silniku turboodrzutowym,
rzucić na nie zaklęcie „ewolucja” i już oto mamy rozwiązanie problemu powstania silnika
turboodrzutowego dzięki ewolucji silnika z odkurzacza. Przypomina to stary argument, że
możemy wierzyć, że ołów przekształca się w złoto, choć nikt nigdy nie widział jak alchemicy
przeprowadzają taką transformację. A dlaczego możemy w to wierzyć? Bo alchemicy mieli
kamień filozoficzny. W przypadku współczesnych ewolucjonistycznych alchemików
transformujących systemy proste w złożone, miejsce kamienia filozoficznego zajmuje po
prostu koewolucja. Nie wiadomo co prawda, jak taka koewolucja zachodziła, ale wierzymy, że
zachodziła. John Bracht, genetyk i biolog komórkowy na University of California ironizuje:
„W tym przypadku teoria ewolucji funkcjonuje jako „teoria-zapchajdziura”, którą zalepia się
luki w wiedzy [evolution-of-the-gaps theory].” [33]
Oprócz życzeniowego myślenia i braku empirycznego poparcia wyjaśnienie to obciążone jest
innymi poważnymi wadami. Bazuje ono jedynie na tym, że niektóre, około dziesięciu, białek
tworzących bakteryjny silnik (jak np.: FlhA, FlhB, FliR, FliQ, FliP) ujawniają znaczącą
homologię (choć nie są identyczne) z białkami obecnymi w strukturze służącej pałeczce dżumy
wstrzykiwać do komórki toksyny. [34] Jednak sam w sobie ten fakt nie dowodzi jeszcze
naturalistycznej ewolucji — przykładowo wiele technicznych urządzeń o różnych funkcjach
także składa się z podobnych komponentów jak śruby, przekładnie, tryby czy tranzystory.
Taka sytuacja nie przeszkadza nikomu wnioskować o rozumnej genezie tego typu urządzeń.
Dlaczego ma więc przeszkadzać w przypadku bakteryjnego silnika protonowego? [35]
Aby dowieść ewolucyjnych spekulacji, nie wystarczy wskazywać podobieństwo niektórych

podzespołów tego systemu z podzespołami innych systemów. Takie podobieństwo może być
równie dobrze interpretowane jako wynikające z wspólnego projektu — podobnie jak
podobieństwo różnych produktów naszej technologii. [36] Konieczny jest jeszcze testowalny
mechanizm, który byłby w stanie przekształcić jedne struktury w inne. A jedyny znany tego
typu mechanizm pochodzi z inżynierii — a więc popiera koncepcję projektu.
Co gorsza, niektórzy ewolucyjni biologowie twierdzą, że to nie bakteryjny silnik wyewoluował
ze struktury wstrzykującej do komórek toksyny, ale że było dokładnie odwrotnie — to z
bakteryjnego silnika miał wyewoluować aparat wstrzykujący, czyli, że bakteryjny silnik
pojawił się wcześniej niż struktura, z której rzekomo miał wyewoluować. [37] Biologowie ci
argumentują następująco:
Bakterie wyposażone w silniki znajdowane są w bardzo szerokim spektrum tych
mikroorganizmów: u bakterii żyjących w średnich temperaturach i u bakterii termofilnych,
bakterii gram-ujemnych i gram-dodatnich oraz u bakterii spiralnych (krętek). [38] Natomiast
aparat wstrzykujący występuje tylko u kilku szczepów bakterii gram-ujemnych.
Aparat wstrzykujący służy do wprowadzania toksyn do komórek eukariotycznych. Zasadne
jest więc przypuszczenie, że najpierw musiały się pojawić eukarionty, a dopiero później
powstał aparat służący do atakowania tych komórek. Występowanie aparatu wstrzykującego
ograniczone jest nie tylko do niektórych szczepów bakterii gram-ujemnych, ale i do tych tylko
patogenów, które atakują rośliny i zwierzęta. Funkcja struktury wstrzykującej zależy ściśle od
kontaktu z tymi wielokomórkowymi organizmami. To indukuje, że struktura ta pojawiła się
po powstaniu roślin i zwierząt. Jest zaś oczywiste, że prokarionty istniały na długo przed
pojawieniem się komórek eukariotycznych. Ponieważ nie ma żadnych dowodów, by
cyjanobakterie sprzed miliardów lat różniły się znacząco od dzisiejszych, można zakładać, że
posiadały one omawiany system napędowy. [39]
U patogenów roślinnych geny rozwijające aparat wstrzykujący są bardziej podobne do ich
własnych genów rozwijających układ napędowy, niż ma to miejsce u patogenów zwierzęcych,
gdzie podobieństwo genów rozwijających aparat wstrzykujący do genów rozwijających silnik
jest znacznie mniejsze. Skłania to niektórych naukowców do wniosku, że aparat wstrzykujący
pojawił się najpierw u patogenów roślinnych, a następnie rozwijające go geny zostały
„pożyczone” patogenom zwierzęcym przez horyzontalny transfer genów. [40]
Niektóre bakterie w temperaturze powyżej 37-38 stopni C rozwijają organelle aparatu
wstrzykującego (w proces ten zaangażowane są częściowo te same geny, które uczestniczą w
rozwoju silnika). Ponieważ zaś bakteryjny silnik jest strukturą bardziej złożoną od aparatu
wstrzykującego (ten drugi składa się z około 10 różnych białek, podczas gdy silnik z około 30-
40) Scott Minnich, mikrobiolog i genetyk z University of Idaho twierdzi w związku z
powyższym, że aparat wstrzykujący jest zdegenerowaną formą protonowego silnika, który
musiał istnieć wcześniej niż aparat wstrzykujący. [41]
W najlepszym przypadku (i przy dokonaniu szeregu nieweryfikowalnych założeń) możemy
mówić o dewolucji silnika protonowego do aparatu wstrzykującego. Ale jeśli teoria ewolucji
ma ambicje wyjaśniać powstawanie systemów złożonych z prostszych, to ten przykład jest
tego przeciwieństwem. Przykłady utraty złożoności nie mogą służyć jako dowody na jej
wzrost.
Oprócz powyższych zastrzeżeń, problem z ewolucjonistycznym wyjaśnieniem jest taki, że
proteiny, które montują bakteryjny silnik protonowy mają jakoby wywodzić się z systemu nie
tylko znacznie mniej złożonego, ale również zupełnie odmiennego w swojej funkcji. Białka te
musiały stopniowo zmieniać swoje oryginalne molekularne interakcje w kierunku kompletnie
odmiennego systemu takich interakcji. Stare funkcje i interakcje musiały być stopniowo
usuwane i zastępowane nowo postającymi funkcjami i interakcjami. Ale biorąc pod uwagę
liczbę białek i ich wzajmnych zależności, które musiały koewoluować wraz z równoległym
powstawaniem i dodawaniem do systemu nowych białek koniecznych do zmontowania
funkcjonalnego bakteryjnego silnika, jednocześnie z zachowywaniem ciągłej funkcjonalności

takiego systemu, ewolucjonistyczne wyjaśnienie w istocie niczym nie różni się od zwykłego
odwołania się do cudu — a właściwie do całego szeregu cudów.
Jeśli ewolucjoniści chcą wyjaśnić powstanie bakteryjnego silnika bez odwoływania się do
cudów, muszą oni wskazać ciąg funkcjonalnych systemów pośrednich wykształcających ten
system. Musiał więc istnieć ciąg systemów wiodących od A = A_1 przez A_2, A_3, …, do A_n =
B, gdzie każdy taki system był funkcjonalnym ewolucyjnym prekursorem dla następnego.
Implikuje to, że musiała istnieć wyjściowa bakteria A (= A_1) nie zawierająca omawianego
systemu napędowego, ani rzecz jasna genów kodujących ten system. Musiała ona w
ewolucyjnym procesie przez ciąg systemów pośrednich A_i i ich modyfikacje A_(i+1)
wytworzyć finalny system, to jest bakterię B (= A_n), czyli naszą E. coli, posiadającą w pełni
funkcjonalny system lokomocyjny i tym samym operony kodujące tę strukturę. Rzecz jasna
musiał być to proces stopniowy, z ciągłym zachowywaniem funkcjonalności rozwijanego
systemu i z równoległą koewolucją funkcji tego systemu.
Jedynie zaś co mają ewolucjoniści to B (to jest E. coli z jej silnikiem protonowym) i C (to jest
Yersinia pestis z jej aparatem wstrzykującym) i żadnego dobrego argumentu dlaczego C
rzeczywiście można ulokować gdzieś w progresywnym ciągu wiodącym od A_1 do A_n, czyli
B.
Najlepszą chyba ocenę neodarwinistycznych hipotez powstania systemów złożonych dał biolog
molekularny Franklin Harold. Jest on ewolucjonistą, dlatego trudno podejrzewać go o
specjalną sympatię dla twierdzeń kreacjonizmu. W swojej książce The Way of the Cell,
wydanej przez renomowane wydawnictwo Uniwersytetu w Oxfordzie, skrytykował koncepcję
rozumnego projektu w biologii. Stwierdził on:
Jako fundamentalną zasadę powinniśmy odrzucić substytut inteligentnego projektu w dialogu
przypadku i konieczności.
Zaraz potem szczerze wyznał:
Jednak musimy przyznać, że nie ma obecnie szczegółowych darwinistycznych wyjaśnień
ewolucji jakiegokolwiek biochemicznego lub komórkowego systemu, są tylko liczne
życzeniowe spekulacje. [42]
Oto teoria ewolucji w działaniu! Gdy nie ma jakichkolwiek dowodów rozwoju jakiegoś
złożonego systemu wedle neodarwinistycznego scenariusza, to po prostu zakłada się
prawdziwość teorii ewolucji, a następnie przedstawia się hipotetyczne scenariusze ewolucji
takiego systemu w oparciu o to właśnie założenie. Jest to błąd rozumowania nazywany petitio
principii — ewolucjoniści najpierw zakładają prawdziwość teorii ewolucji, by na tej
podstawie próbować dowodzić prawdziwości teorii ewolucji na konkretnych przykładach.
Tylko po co, skoro to już zostało założone na wstępie? Po tej operacji wymyślenie jakiejś
mądrze brzmiącej historyjki i zamiana jej — jak za dotknięciem czarodziejskiej (pardon:
ewolucyjnej) różdżki — na „znakomicie potwierdzone, naukowe i akceptowane przez
większość biologów wyjaśnienie” nie stanowi już problemu. [43]
Ile warte są takie neodarwinistyczne historyjki wie również Lynn Margulis, znana biolog,
twórczyni — obecnie powszechnie przyjętej — teorii symbiotycznego pochodzenia komórki
eukariotycznej:
Jak słodka przekąska, która na krótko zaspakaja nasz głód, ale nie dostarcza wartościowego
pożywienia, tak neodarwinizm zaspakaja intelektualną ciekawość abstrakcjami
pozbawionymi faktualnych detali dotyczących czy to metabolizmu, czy biochemii, czy
ekologii, czy historii naturalnej. [44]
Z kolei w Nature biolog ewolucyjny z University of Chicago, Jerry Coyne, stwierdził:
Nie ma żadnych wątpliwości, że złożoność procesów opisanych przez Behe’ego zniechęca do
prowadzenia badań nad nimi i trudno będzie rozwiązać zagadkę ich ewolucji. […] Być może

nigdy nie będziemy w stanie wyobrazić sobie pierwszych protoprocesów. [45]
Jak widać, zasadniczym ewolucyjnym argumentem staje się możliwość wyobrażenia sobie
hipotetycznej ewolucji złożonych biochemicznych systemów. Że bujna wyobraźnia zamiast
solidnej argumentacji to w nauce trochę za mało, zdaje się zdawać sobie sprawę biolog
ewolucyjny z University of British Columbia, Tom Cavalier-Smith, który w Trends in Ecology
and Evolution, zamieścił następujący komentarz:
Dla żadnego z wymienionych przez Behe’ego przypadków nie ma jeszcze wyczerpującego i
szczegółowego wyjaśnienia prawdopodobnych etapów ewolucji obserwowanej złożoności.
Problemy te rzeczywiście poważnie zaniedbywano — choć Behe nieustannie przesadnie
określa to zaniedbanie takimi hiperbolami, jak „głuche i zupełne milczenie”. [46]
Na zaskakującą szczerość dla New Scientist zdobył się także biolog ewolucyjny z University
College w Londynie, Andrew Pomiankowski:
Weź do ręki jakikolwiek podręcznik do biochemii, a znajdziesz może dwa lub trzy odniesienia
do ewolucji. Przeczytaj któryś z nich, a będziesz miał szczęście, jeśli znajdziesz jakieś lepsze
zdanie niż to, że „ewolucja dobiera molekuły najlepiej dostosowane do swoich biologicznych
funkcji”. [47]
Zgadza się z nim biolog molekularny, Robert Dorit zapewniający w American Scientist, że:
W wąskim sensie Behe ma rację argumentując, że jeszcze nie w pełni rozumiemy, jak zaszła
ewolucja silnika wici czy kaskady krzepnięcia krwi. [48]
Że Behe ma rację, nie tylko w wąskim, ale i znacznie szerszym sensie potwierdza również
James Shapiro, biochemik i mikrobiolog z University of Chicago. W 1996 r. w swojej recenzji
książki Michaela Behe’ego Darwin’s Black Box stwierdził on:
Nie istnieją szczegółowe darwinistyczne wyjaśnienia ewolucji jakiegokolwiek
fundamentalnego biochemicznego lub komórkowego systemu, nie licząc zatrzęsienia
życzeniowych spekulacji. To niepojęte, że darwinizm jest akceptowany jako satysfakcjonujące
wyjaśnienie dla tak rozległego tematu — ewolucji — na podstawie tak słabego zbadania, jak
dobrze jego tezy sprawdzają się w specyficznych przypadkach biologicznej adaptacji lub
różnorodności. [49]
„Biolodzy ewolucyjni mają jednak odpowiedź na jego [Behe'ego] zarzuty” [50] — tryumfalnie
obwieszcza John Rennie w Świecie Nauki. Nie wyjaśnił niestety, jakie są to odpowiedzi. Że —
w świetle powyższych cytatów — te odpowiedzi o (ko)ewolucji bakteryjnego silnika
protonowego to typowe ewolucjonistyczne „takie sobie bajeczkki” (just-so story), wymyślane
przez ewolucyjnych magików w celu zamaskowania przed szerszą publiką poważnych
słabości ich teorii. Ewolucyjni biologowie znają na pamięć przepis na „taką sobie bajeczką”,
jednak to pojęcie może być laikom nieznane. Uchylmy nieco rąbka tajemnicy, zaglądając do
ewolucjonistycznej kuchni i podglądając przepis na ewolucyjną „taką sobie bajeczkę” mającą
wyjaśniać np. ewolucję układów nieredukowalnie złożonych. Oto on:
Przepis na dobrą ewolucyjną „taką sobie bajeczkę”
Najpierw przejrzyj biologiczne systemy pod względem ich funkcji i/lub struktury. Znajdź
taki, który wydaje się najlepszy do zrobienia z niego prekursora docelowego systemu i
dosztukuj te systemy razem, nie zawracając sobie specjalnie głowy biochemicznymi i/lub
genetycznymi szczegółami. Umieść taką zszywkę w wymyślonym stworzeniu z odległej
przeszłości, przyjmując wygodnie, że stworzenie to dawno wyginęło. Ogólnikowo, ale
uroczyście przywołaj selekcyjną presję, która działała na zszywkę, a następnie wyobraź ją
sobie jako strukturę plastyczną i podatną na kolejne modyfikacje, które dobór naturalny już
prostą drogą doprowadził do docelowego systemu. Gotowe!
Nie dziwota więc, że William Dembski określił ewolucjonistyczne „wyjaśnienia” powstania

bakteryjnego silnika jako „śmiecie”. [51] W świetle powyższego trudno się również dziwić, że
Behe i inni zwolennicy teorii projektu oczekują czegoś więcej niż naprędce sklecanych
życzeniowych opowiastek. [52]
Wniosek: projekt
Zwolennikom teorii inteligentnego projektu nie wystarczają zaklęcia i opowiadanie „takich
sobie bajeczek”, w które obfituje ewolucjonistyczna literatura próbująca wyjaśnić powstanie
bakteryjnego silnika protonowego. [53] Brak dowodów na bezpośrednią lub niebezpośrednią
ewolucję systemów nieredukowalnie złożonych jest powszechny i systematyczny — podobnie
jak brak dowodów na istnienie krasnoludków. Wspomniany John Bracht podsumowuje:
Prowadzi to do niekończących się, luźnych spekulacji bazujących całkowicie na założeniu, że
dobór naturalny może stworzyć wszystko, czego wymaga projekt. Ta historia, której
prawdziwość po prostu się zakłada, jest nietestowalna i co za tym idzie niefalsyfikowalna.
Jednak dlaczego mamy wierzyć w tego typu opowiastki? [54]
Właśnie, dlaczego mamy wierzyć w ewolucyjną cudotwórczą mitologię przypadku, która sobie
tylko znanym sposobem wytworzyła nanotechnologiczne perełki, skoro dostępne jest
wyjaśnienie bardziej racjonalne i prostsze? Dlaczego nie uznać, że takie systemy zostały
uformowane dzięki koncepcji, która połączyła różne komponenty w jedną, funkcjonalną
całość? Innymi słowy powstały one przez projekt. Dokładnie tak, jak ma to miejsce w
systemach projektowanych przez inżynierów. Twarda emipria bazująca na naszej inżynierii
taki właśnie wniosek podsuwa. Wyjaśnienie to spełnia również wymagania przyczynowej
adekwatności, to znaczy jego moc wyjaśniająca jest adekwatna do skali problemu, czego nie
można powiedzieć o wyjaśnieniach ewolucjonistycznych.
Oprócz bakteryjnego silnika protonowego zwolennicy teorii projektu analizują także inne
molekularne maszyny i systemy pod kątem projektu i nieredukowalnej złożoności.
Przykładowo artykuł biochemika z Uniwersytetu w Cambridge Douglasa Axe, który ukazał
się w Journal of Molecular Biology [55] dokumentuje, że pewne enzymy są niezwykle
wrażliwe na perturbacje. W tym przypadku perturbacje nie tylko po prostu uszkadzają
istniejące funkcje, ale niszczą również wszelką możliwą funkcjonalność takich enzymów.
Implikuje to, że systemów takich nie mógł stopniowo wytwarzać mechanizm neodarwinowski
— są one dla niego niedostępne. Co więcej, analiza (nie)prawdopodobieństwa powstania tego
typu układów w wyniku naturalistycznych procesów ewolucyjnych, wyraźnie wskazuje na
konieczność wniosku o projekcie.
W innym artykule, który ukazał się w bardzo prestiżowym naukowym czasopiśmie Annual
Review of Genetics, w którym publikują tylko zaproszeni przez redakcję autorzy, znani
niemieccy genetycy i szefowie grup badawczych w Max Planck Institute for Plant Breeding
Research, [56] Wolf-Ekkehard Lönnig i Heinz Saedler analizują rolę transpozonów i innych
mobilnych genetycznych elementów w nielosowych reorganizacjach chromosomów i w
związku z tym, w nagłym powstawaniu nowych gatunków. Autorzy dyskutują
antydarwinistyczne koncepcje noblistki Barbary McClintock, zastanawiając się również nad
możliwością częściowo predeterminowanego generowania bioróżnorodności i nowych
gatunków. [57]
Pojawianie się artykułów otwarcie nawiązujących do koncepcji projektu w cenionej
literaturze naukowej głównego nurtu [58] może świadczyć, że (r)ewolucyjna czujność nieco
osłabła, co zresztą spowodowało histeryczne reakcje co bardziej znerwicowanych
ewolucjonistów. [59]
Niektórzy ewolucjoniści, przyparci do muru, bronią się słusznym stwierdzeniem, że brak
dowodu na ewolucję nie jest tym samym co dowód na brak takowej ewolucji. Tym samym
potwierdzają jednak impotencję teorii ewolucji do wiarygodnego i popartego faktami
wyjaśniania genezy złożonych struktur typu bakteryjnego silnika protonowego. [60]

Ostatecznie brak dowodów na istnienie krasnoludków nie jest tym samym co dowód na ich
nieistnienie, co nie przeszkadza zasadnie mniemać, że krasnoludki nie istnieją.
Ostatnią linią ewolucjonistycznej obrony jest „obiecujący materializm” — stwierdzenie, że
być może w przyszłości znajdziemy rozwiązanie tego problemu. Nauka jednak działa na
aktualnych dowodach, a nie na mglistych obietnicach, że takie dowody „może pojawią się w
przyszłości”. Może takie dowody się pojawią, a może przeciwnie — dalsze odkrycia będą
potęgować zagadki związane z ewolucyjnym powstaniem złożonych biochemicznych
systemów.
W zgodzie z obecnym naukowym świadectwem, najbardziej racjonalnym wnioskiem o genezie
bakteryjnego silnika protonowego — podobnie jak innych biologicznych molekularnych
maszyn [61] — jest taki, że jest on strukturą powstałą w wyniku rozumnego projektu.
Michał Ostrowski
Przypisy
[1] Michael J. Behe, Darwin’s Black Box. The Biochemical Challenge to Evolution, Free Press,
New York 1996, s. 69-73. Książka ta została uznana przez National Review i World Magazine
za jedną ze 100 najważniejszych książek XX wieku (zob. Gene Edward Veith and Marvin
Olasky, The century’s top 100 books, World Magazine 4 December 1999, vol. 14, No. 47).
[2] Tak bakteryjny silnik określił jeden z liderów ruchu teorii inteligentnego projektu
matematyk i filozof, William A. Dembski w artykule The Logicall Underpinnings of
Intelligent Design (pdf 175 kb), s. 16, który jest częścią zredagowanej przez Dembskiego i
Michaela Ruse’a książki Debating Design: From Darwin to DNA, wydanej w 2004 roku
nakładem prestiżowego Cambridge University Press. Książka ta zawiera m.in. polemiki
pomiędzy zwolennikami teorii ewolucji i teorii inteligentnego projektu.
[3] Przeliczając to na nasze realia: jeśli bakteria długości 2 mikrometrów porusza się z
prędkością 40 swoich długości na sekundę, to podwodna motorówka długości 2 metrów o
podobnie efektywnym systemie napędowym musiałaby poruszać się z prędkością 180-360
km/godz.
[4] Gdyby powiększyć bakterię do wielkości półtorakilogramowego, podłużnego bochenka
chleba, wici miałyby długość dwóch metrów.
[5] Por. Howard C. Berg, The rotary motor of bacterial flagella, Annual Review of
Biochemistry July 2003, vol. 72, s. 19-54.
[6] Zob. David J. DeRosier, Spinning Tails, Current Opinion in Structural Biology April 1995,
vol. 5, No. 2, s. 187-93.
[7] Por. Jiadong Zhou, Scott A. Lloyd, and David F. Blair, Electrostatic interactions between
rotor and stator in the bacterial flagellar motor, Proceedings of the National Academy of
Sciences USA May 1998, vol. 95, No. 11, s. 6436–6441.
[8] Por. Avraham Oplatka, Do the bacterial flagellar motor and ATP synthase operate as water
turbines?, Biochemical and Biophysical Research Communications 28 August 1998, vol. 249,
No. 3, s. 573-578.
[9] David F. Blair and Scott A. Lloyd, Charged residues of the rotor protein FliG essential for
torque generation in the flagellar motor of Escherichia coli, Journal of Theoretical Biology 7
March 1997, vol. 266, No. 4, s. 733-744.
[10] Robert M. Macnab, The bacterial flagellum: reversible rotary propellor and type III
export apparatus, Journal of Bacteriology December 1999, vol. 181, No. 23, s. 7149-7153.
[11] Por. Robert M. Macnab, How bacteria assemble flagella, Annual Review of Microbiology
October 2003, vol. 57, s. 77-100.

[12] Animowaną rekonstrukcję tego procesu przygotowaną przez Keiichi Namba obejrzeć
można na stronach Bacterial Locomotion and Signal Transduction, Inc. (BLAST).
[13] Por. Piotr Lenartowicz SJ, Rozwój i postęp w świetle empirii biologicznej, [w:]
Humanizm ekologiczny. Kryzys idei postępu wymiar ekologiczny, vol. 2, Politechnika
Lubelska, Lublin 1993, s. 179-180 [173-187].
[14] Liczba Reynoldsa dla E. coli jest mniejsza niż 10-4.
[15] Howard C. Berg, Motile Behavior of Bacteria, Physics Today January 2000, vol. 53, No. 1,
s. 24-29.
[16] Hasło „Maszyna” w: Nowa encyklopedia powszechna PWN, t. 4, Wydawnictwo Naukowe
PWN, Warszawa 1996, s. 118.
[17] Bruce Alberts, The Cell as a Collection of Protein Machines: Preparing the Next
Generation of Molecular Biologists, Cell 8 February 1998, vol. 92, No. 3, s. 291 [291-294].
[18] Adam Wilkins, A Special Issue on Molecular Machines, BioEssays December 2003, vol.
25, No. 12, s. 1146 [1145-1146].
[19] Paul Ricci, Fundamentals of Critical Thinking, Ginn Press, Lexington MA 1986, s. 190.
[20] Francisco Ayala, Darwin’s Revolution, [w:] John H. Campbell and J. William Schopf
(eds.), Creative Evolution?!, Jones and Bartlett, Boston 1994, s. 4.
[21] Ayala, Darwin’s Revolution…, s. 5.
[22] William A. Dembski, Still Spinning Just Fine: A Response to Ken Miller, Progress in
Complexity, Information, and Design January-June 2003, vol. 2.1-2.2.
[23] Richard H. Thornhill, David W. Ussery, A classification of possible routes of Darwinian
evolution, Journal of Theoretical Biology 2000, vol. 203, No. 2, s. 112 [111-116].
[24] D. Dewar, L.M. Davies, and J.B.S. Haldane, Is Evolution a Myth? A Debate between D.
Dewar and L.M. Davies vs. J.B.S. Haldane, Watts & Co. Ltd/Paternoster Press, London 1949,
s. 90.
[25] Behe, Darwin’s Black Box…, s. 39. William Dembski proponuje nieco rozszerzoną
definicję nieredukowalnej złożoności:
System wykonujący podstawową funkcję jest nieredukowalnie złożony, jeśli zawiera
określona liczbę precyzyjnie zintegrowanych, wzajemnie oddziałujących podzespołów, tak że
każdy taki podzespół jest konieczny do zachowania podstawowej funkcji systemu i przez to
oryginalnej funkcji. Liczba tych koniecznych podzespołów jest określana jako
nieredukowalny rdzeń systemu.
(William A. Dembski, No Free Lunch. Why Specified Complexity Cannot Be Purchased
Without Intelligence, Rowman & Littlefield Publishers 2001, s. 285.)
[26] John Rennie, 15 odpowiedzi na nonsensowne tezy kreacjonistów, Świat Nauki wrzesień
2002, nr 9 (133), s. 72 [66-72], podkreślenie dodane. Rennie fałszuje twierdzenie Behe’ego
jakoby przypisywał on elementom składającym się na bakteryjny silnik przydatność tylko
jako podzespołom napędowym. Na podstawie podanych powyżej oryginalnej definicji układu
nieredukowalnie złożonego w rozumieniu Behe’ego, jak i rozszerzonej definicji Dembskiego
nie można wyciągnąć takiego wniosku. Najwyraźniej Rennie nie do końca orientuje się o co
chodzi w koncepcji nieredukowalnej złożoności i co z niej wynika, a co nie.
[27] Richard Dawkins, Wspinaczka na szczyt nieprawdopodobieństwa, Prószyński i S-ka,
Warszawa 1998.
[28] Por. Karol Darwin, O powstawaniu gatunków drogą doboru naturalnego, czyli o

utrzymywaniu się doskonalszych ras w walce o byt, [w:] Dzieła wybrane, t. 2, Państwowe
Wyd. Rolnicze i Leśne, Warszawa 1959, s. 182.
[29] Meandry ewolucjonistycznej logiki szerzej omówiłem w artykule: Królowa nauk.
Wprowadzenie do ewolucjonistycznej logiki, Na Początku… listopad-grudzień 2002, nr 11-
12A, s. 350-355.
[30] Trzymając się analogii Millera i zakładając, że rzeczywiście pułapki na myszy były
funkcjonalnymi prekursorami spinek do krawatów: zarówno pułapka na myszy, jak i
bakteryjny silnik budowane są z odpowiednich części (dla wygody pomińmy kolosalną różnicę
w złożoności tych systemów). Silnik montowany jest przez bakteryjną komórkę, pułapka na
myszy w odpowiednim zakładzie. Jeśli ta analogia ewolucji spinka do krawatu — pułapka na
myszy ma być sensowna musimy uwierzyć, że dawno, dawno temu była sobie
zautomatyzowana fabryka spinek na krawaty, w której dochodziło na taśmie produkcyjnej do
szeregu błędów. W ich wyniku spinki do krawatu zaczęły powoli zmieniać swoją funkcję
przekształcając się stopniowo w pułapki na myszy. I to przy zachowaniu ciągłej
funkcjonalności tych ewoluujących bubli, czyli „spinko-pułapek”.
[31] Rennie, 15 odpowiedzi…, s. 71.
[32] W literaturze anglojęzycznej ten wstrzykujący aparat określa się czasem skrótem TTSS
(type III secretory system).
[33] John Bracht, The Bacterial Flagellum: A Response to Ursula Goodenough (pdf 109 kb),
Progress in Complexity, Information, and Design January-June 2003, vol. 2.1-2.2.
[34] Por. Christoph J. Hueck, Type III Protein Secretion Systems in Bacterial Pathogens of
Animals and Plants, Microbiology and Molecular Biology Reviews June 1998, vol. 62, No. 2, s.
379-433.
[35] Zaznaczyć tu trzeba, że zwolennicy teorii inteligentnego projektu generalnie — choć nie
wszyscy — akceptują pochodzenie od wspólnego przodka. Odrzucają natomiast
neodarwinistyczny mechanizm, który odpowiada jakoby za różnicowanie się życia. Ich
zdaniem źródłem większych funkcjonalnych i strukturalnych transformacji jest rozumny
projekt, a nie makroewolucja. Kazimierz Jodkowski tak wyjaśnia ten problem:
„Pochodzenie od wspólnego przodka nie jest ani konieczne, ani wystarczające, by uznać jakiś
proces za ewolucję. Istnieją nieewolucjoniści, którzy wierzą w pochodzenie od wspólnego
przodka (Michael Behe, David Wilcox), ale istnieją też ewolucjoniści, którzy nie wierzą w
takie pochodzenie (np. G.A. Kerkut, C. Schwabe i G. Warr). Denton wskazuje, że pochodzenie
od wspólnego przodka jest zgodne z niemal każdą filozofią przyrody, także i z
kreacjonizmem:
Prawdą jest, że zarówno podobieństwo homologiczne z prawdziwego zdarzenia, to jest takie,
gdzie zjawisko to ma jasną genetyczną i embriologiczną podstawę (co, jak wyżej widzieliśmy,
jest dużo mniej powszechne, niż to się często zakłada), oraz hierarchiczne wzorce relacji klas
sugerują pewien rodzaj teorii pochodzenia. Ale żadne z nich nie mówi nam niczego na temat,
jak to pochodzenie czy ewolucja mogła zachodzić, czy proces ten był stopniowy, czy nagły,
albo czy mechanizm kauzalny był darwinowski, lamarckowski, witalistyczny czy nawet
kreacjonistyczny. Taka teoria pochodzenia jest dlatego pozbawiona jakiegokolwiek znaczenia
i w równym stopniu zgodna z niemal każdą filozofią przyrody.
Wilcox zwrócił uwagę, że o pochodzeniu od wspólnego przodka wiedziano i przed Darwinem,
a zasługą tego ostatniego było wskazanie przyrodniczego, materialnego źródła dla
różnicowania się świata ożywionego:
Artykuł ten dotyczy struktury biologicznej, a nie związku jej z biologicznym pochodzeniem.
Świadectwo na rzecz strukturalnej różnicy/pochodzenia nie stanowi świadectwa na rzecz
mechanizmu, przy pomocy którego zachodziła transformacja strukturalna. Dlatego te rodzaje

świadectwa empirycznego, które po prostu wskazują na związek i pochodzenie od wspólnego
przodka (np. dane zegara molekularnego, sekwencje kopalne, powiązania chromosomowe i
inne pomiary podobieństwa) nie są ważne dla tej kwestii, o ile nie wskazują na naturę
mechanizmu stwórczego, który produkował nowość w trakcie tego pochodzenia. Świadectwo
na rzecz pochodzenia nie implikuje posiadania wiedzy o morfogenetycznych mechanizmach
zdolnych wyprodukować nowość. Być może lepiej to rozumiano w dziewiętnastym stuleciu,
niż obecnie (Muller i Wagner 1991). I rzeczywiście, do 1850 roku niemal wszyscy badacze
akceptowali pochodzenie od wspólnego przodka (Gillespie 1979; Desmond 1989). Wyjątkową
implikacją teorii Darwina było dlatego nie pochodzenie, ale sugestia, że źródło porządku
biotycznego należy znajdować w porządku przyrodniczym (materialnym).
Sam Darwin przyznawał, że wspólne pochodzenie nie wystarczy, trzeba jeszcze pokazać, jak
ono następowało:
Zastanawiając się nad powstawaniem gatunków pojmiemy łatwo, że przyrodnik, który bada
wzajemne powinowactwo istot organicznych, ich stosunki w okresie zarodkowym, ich
rozmieszczenie geograficzne, ich geologiczne następstwo i inne podobne fakty, dojść może do
wniosku, że gatunki nie zostały stworzone oddzielne, ale, tak jak odmiany, powstały z innych
gatunków. Wniosek ten jednak, choćby nawet dobrze uzasadniony, nie wystarczy jeszcze,
dopóki nie zostanie wykazane, w jaki sposób niezliczone gatunki zamieszkujące powierzchnię
ziemi zostały tak przekształcone, iż uzyskały ową doskonałość budowy i przystosowania, które
słusznie budzą w nas podziw”.
(Kazimierz Jodkowski, Metodologiczne aspekty kontrowersji ewolucjonizm-kreacjonizm,
Realizm. Racjonalność. Relatywizm, t. 35, Wyd. UMCS, Lublin 1998, s. 217-218, podkreślenia
w oryginale, przypisy pominięte.)
Jodkowski cytuje również Delvina Ratzscha:
Przypuśćmy, że współczesna teoria ewolucji ma tak mocno wbudowany mechanizm ślepego
przypadku, że po prostu nie ma sposobu pogodzenia jej z jakimkolwiek rodzajem Boskiego
kierownictwa. A jednak nadal teiści mogliby odrzucać tę teorię ewolucji i przyjmować zamiast
niej teorię, wedle której procesy i prawa przyrodnicze stanowią mechanizm większości
procesu ewolucyjnego, ale od czasu do czasu Bóg ograniczał te prawa i wprowadzał jakąś
kluczową mutację w trakcie wydarzeń. Nawet gdyby Bóg miał interweniować bezpośrednio,
zawieszając prawa przyrody i wprowadzając istotny nowy materiał genetyczny w różnych
momentach, by doprowadzić do wyłonienia się nowych cech i w końcu nowych gatunków, to
ta cudowna i opracowana Boska interwencja nie naruszyłaby sama w sobie takich kluczowych
tez teorii ewolucji jak ta, że wszystkie gatunki pochodzą ostatecznie od jakiegoś wspólnego
przodka. Pochodzenie z genetyczną interwencją jest nadal pochodzeniem — jest to po prostu
pochodzenie z nieprzyrodniczymi elementami.
(Jodkowski, Metodologiczne aspekty…, s. 217.)
Na temat niefalsyfikowalności koncepcji wspólnego pochodzenia patrz również: Paul Nelson
and Jonathan Wells, Is Common Descent an Axiom of Biology?, referat wygłoszony na
konferencji „The Darwinian Paradigm: Problems and Prospects” odbytej w dniach 22-25
czerwca 1993 r. w Pajaro Dunes w Kalifornii.
[36] Zresztą nawet w obrębie neodarwinizmu nie każde podobieństwo interpretuje się jako
świadectwo wspólnego pochodzenia. Czyli inaczej: nie każde podobieństwo oznacza
pokrewieństwo. Znanym fenomenem świata ożywionego jest konwergencja (ewolucja zbieżna,
w terminologii kladystycznej zwana także homoplazją), czyli powstawanie bardzo podobnych
cech i funkcji niezależnie, w organizmach spokrewnionych ze sobą bardzo odlegle. Niektórzy
ewolucjoniści argumentują, że konwergencja jest bardzo powszechną cechą życia, z czego
dopiero teraz zaczynamy w pełni zdawać sobie sprawę. Znany paleobiolog z Uniwersytetu w
Cambridge, Simon Conway Morris w swojej ostatniej książce Life’s Solution: Inevitable

Humans in a Lonely Universe (Cambridge University Press, Cambridge 2003) dowodzi, że
świat ożywiony pełen jest konwergencji obserwowanej na każdym poziomie: molekuł,
systemów komórkowych, cech anatomicznych, a nawet różnych typów inteligencji —
wszystko to ujawnia konwergencję.
[37] Lily Nguyen, Ian T. Paulsen, Jason Tchieu, Christoph J. Hueck, Milton H. Saier Jr.,
Phylogenetic analyses of the constituents of Type III protein secretion systems, Journal of
Molecular Microbiology and Biotechnology April 2000, vol. 2, No. 2, s. 125-44.
[38] Por. Rasika M. Harshey, and Adam Toguchi, Spinning tails: homologies among bacterial
flagellar systems, Trends in Microbiology June 1996, vol. 4, No. 6, s. 226-231.
[39] Pierwsze prokarionty (sinice), bardzo przypominające współczesne cyjanobakterie
(Cyanobacteria), pojawiają się w zapisie kopalnym około 3,5 miliarda lat temu (por. William
J. Schopf, Microfossils of the Early Archean Apex chert: new evidence of the antiquity of life,
Science 30 April 1993, vol. 260, s. 640-646). Niektórzy badacze twierdzą, że prokarionty
pojawiły się jeszcze wcześniej: „W 1997 r. Steve Mojzsis, geolog z Uniwersytetu
Kalifornijskiego w Los Angeles, znalazł jego ślady [życia] w liczących sobie 3,85 mld lat
skałach grenlandzkiej wyspy Akilia. Wiek samej Ziemi wynosi około 4,6 mld lat i początkowo
panujące na jej powierzchni warunki nie mogły sprzyjać istnieniu życia biologicznego”
(Krzysztof Szymborski, Kamienne anioły. Czy życie przybyło z kosmosu — nowe hipotezy,
Polityka wrzesień 2002 [2338]). Zob. również: S.J. Mojzsis, G. Arrhenius, K.D. McKeegan,
T.M. Harrison, A.P. Nutman and C.R.L. Friend, Evidence for life on Earth before 3,800
million years ago, Nature 7 November 1996, vol. 384, s. 55-59 i J.M. Eiler, S.J. Mojzsis, G.
Arrhenius, Carbon isotope evidence for early life, Nature 17 April 1997, vol. 386, s. 665.
[40] C. Stephens, L. Shapiro, Delivering the payload. Bacterial pathogenesis, Current Biology
1 August 1996, vol. 6, No. 8, s. 927-930.
[41] Scott Minnich, Bacterial Flagella: Spinning Tails of Complexity and Co-Option.
[42] Franklin M. Harold, The Way of the Cell: Molecules, Organisms and the Order of Life,
Oxford University Press, Oxford 2001, s. 205.
[43] Petitio principii trenuje również cytowany powyżej John Rennie. Najpierw stwierdza, że
ewolucja bakteryjnego silnika „mogła” polegać na nowej kombinacji skomplikowanych
podzespołów, a skoro mogła to od razu wnioskuje, że pewnie i polegała, no bo przecież
„ewolucja musi być prawdziwa”.
[44] Lynn Margulis, Acquiring Genomes: A Theory of the Origins of Species, Basic Books,
New York 2002, s. 103.
[45] Jerry A. Coyne, God in the Details, Nature 19 September 1996, vol. 383, s. 227-228.
[46] Tom Cavalier-Smith, The Blind Biochemist, Trends in Ecology and Evolution 1997, vol.
12, No. 4, s. 162-163.
[47] Andrew Pomiankowski, The God of the Tiny Gaps, New Scientist, September 1996, vol.
14, s. 44-45.
[48] Robert Dorit, Molecular Evolution and Scientific Inquiry, Misperceived, American
Scientist 1997, vol. 85, s. 474-475.
[49] James A. Shapiro, In the details… What?, National Review 16 September 1996, vol. 48,
Issue 17, s. 62 [62-65].
[50] Rennie, 15 odpowiedzi…, s. 71.
[51] Dembski, The Logicall Underpinnings…, s. 17.
[52] Ujmując materię w symbole, spójrzmy czego potrzebują ewolucjoniści, aby ich

wyjaśnienia stały się czymś więcej niż „takimi sobie bajeczkami” i życzeniowymi
spekulacjami, a tym samym mogły wiarygodnie wyjaśniać powstawanie układów
nieredukowalnie złożonych. W rzeczy samej, problem jest nieco podobny do tego dotyczącego
ewolucji różnych gatunków w zapisie kopalnym niestety niewidocznej. Mając organizmy X i Y
— jeśli wywodzą się one od wspólnego przodka via mechanizm neodarwinowski — musiał
istnieć łańcuch form pośrednich X(0) —> X(1) —> … —> X(m) = X i Y(0) —> Y(1) —> … —
> Y(n) = Y gdzie X(0) = Y(0), czyli wspólny przodek.
Najlepiej dla dowodzenia teorii ewolucji byłoby, gdyby jak najwięcej takich form pośrednich
X(i) i Y(j) było wyraźnie uwidocznionych w kopalnym zapisie. Każda strzałka reprezentuje
przekształcanie się gatunków — oznacza to wytwarzanie niewielkich kumulujących się zmian,
zgodnie z neodarwinowskim mechanizmem. Im więcej takich form pośrednich „uznanych jest
za zaginione”, tym więcej koniecznych jest „takich sobie bajeczek” opisujących takie
stopniowe przemiany i tym bardziej niewiarygodne stają się ewolucjonistyczne wyjaśnienia.
(O sprzeczności kopalnego zapisu z teorią ewolucji, patrz: Don Batten, Teoria przerywanej
równowagi, Na Początku… wrzesień-październik 2000, nr 9-10 (133-134), s. 260-282;
Eksplozja kambryjska kontra ewolucyjne drzewo rodowe; Ewolucja kontra fakty –
zestawienia graficzne).
W przypadku systemu nieredukowalnie złożonego S zawierającego komponenty {S(1), S(2),
…, S(r)} zorganizowane w jakąś nieredukowalnie złożoną konfigurację C, aby wiarygodnie
dowieść ewolucji takiego układu potrzebujemy sekwencyjnego ciągu pośrednich systemów
T[1] —> T[2] —> … —> T[n], gdzie każdy taki system T[i] zawiera z kolei jeden lub więcej
funkcjonalnych biologicznych podsystemów. Zatem T[i] = {T[i](1), … T[i](k)} (k jest zależne
od i i wynosi przynajmniej 1) i każdy podsystem T[i](j) składający się z m komponentów {T[i]
(j)(1), … T[i](j)(m)} (m jest zależne od i i od j) i zorganizowane w jakąś funkcjonalną
konfigurację C[i](j) (niekoniecznie nieredukowalnie złożoną). W takiej sekwencji T[1] —>
T[2] —> … —> T[n], T[n] = {T[n](1)} a T[n](1) = S, czyli finalny system.
Przy założeniu, że system S wyewoluował via mechanizm neodarwinowski, musiał istnieć
system T[1], w którym komponenty S były albo nieobecne, albo działające w zupełnie innym
podsystemie i pełniące zupełnie inne funkcje. Taki system T[1] musiał być punktem
wyjściowym — jeśli system S rzeczywiście wyewoluował dzięki mechanizmom
neodarwinowskim. Każda strzałka w tym schemacie reprezentuje małe kumulujące się
zmiany zgodnie z neodarwinowskim mechanizmem. Najlepiej dla dowodzenia ewolucji
byłoby, gdybyśmy mogli to prześledzić na poziomie pojedynczego komponentu T[i](j)(k)
powoli przekształcającego się wraz z równoczesną stopniową i ciągłą modyfikacją swojej
funkcji (koewolucja) i w ten sposób ewoluującego do komponentu T[i+1](j’)(k’) systemu
T[i+1](j’). Zatem ewolucja T[i] —> T[i+1] musi równolegle zmieniać T[i](j)(k) —> T[i+1](j’)
(k’).
Optymalną sytuacją byłaby taka, gdyby każdy taki system T[i] mógł być bezpośrednio
pokazany na poziomie komponentu T[i](j)(k) razem ze wszystkimi konfiguracjami C[i](j).
Każde przekształcanie systemu — reprezentowane przez strzałki — musi wytwarzać małe
kumulujące się zmiany zgodnie z neodarwinowskim mechanizmem. Te zmiany powinny: być
„niezwykle drobne i stopniowe”, operować na poziomie komponentów poszczególnego
systemu, a funkcje systemu winny być indukowane (rzecz jasna również stopniowo, drobnymi
kroczkami i równolegle z ewolucją komponentów) dzięki zmieniającej się konfiguracji
komponentów. Im więcej form pośrednich w tym schemacie „uznanych jest za zaginione” (czy
to całe systemy, czy komponenty, czy konfiguracje, czy funkcje), tym więcej koniecznych jest
„takich sobie bajeczek” opisujących ewolucję takich systemów i tym bardziej niewiarygodne
stają się ewolucjonistyczne wyjaśnienia.
W tym schemacie wszystko co aktualnie możemy obserwować, to funkcjonalne komponenty
T[i](j)(k)s oraz systemy T[i](j)s obecnych biochemicznych układów. Nie wiadomo jakie były

pośrednie systemy T[i]s lub konfiguracje C[i](j)s. Ale nade wszystko strzałki reprezentujące
stopniowe transformacje zamiatane są po prostu pod dywan dzięki inwokacji do doboru
naturalnego — tak jakby pojęcie doboru naturalnego było magicznym zaklęciem
automatycznie gwarantującym sukces ewolucyjnego procesu. Dla dobrze uformowanych
ewolucjonistów takie zaklęcie jest trickiem pozwalającym wypełnić wszystkie luki pomiędzy
poszczególnymi systemami i w ten sposób „ewolucyjnie” połączyć te systemy. Dla sceptyków
magiczne zaklęcie pozostaje tylko zaklęciem, nawet jeśli nazywiemy je „dobór naturalny”,
jeśli nie jest poparte testowalnym, empirycznym materiałem dowodowym.
[53] Doskonały krytyczny przegląd ewolucjonistycznych wyjaśnień powstania bakteryjnego
silnika daje biolog molekularny piszący pod pseudonimem Mike Gene: Evolutionary Origin
of Bacterial Flagellum Through Cooption: A Critical Survey. Part 1-6.
[54] Bracht, The Bacterial Flagellum…
[55] Por. Douglas D. Axe, Extreme Functional Sensitivity to Conservative Amino Acid
Changes on Enzyme Exteriors (pdf 420 kb), Journal of Molecular Biology 2000, vol. 301, No.
3, s. 585-595.
[56] Max-Planck-Institut für Züchtungsforschung, Carl-von-Linne-Weg 10, D-50829 Köln,
Germany.
[57] Wolf-Ekkehard Lönnig and Heinz Saedler, Chromosome Rearrangements and
Transposable Elements, Annual Review of Genetics December 2002, vol. 36, s. 389-410.
Zobacz również prywatne strony Lönniga zawierające jego liczne artykuły krytyczne wobec
teorii ewolucji i polemiki z ewolucyjnymi biologami.
[58] Jako przykłady niedarwinistycznych interpretacji biologicznej rzeczywistości patrz
również: Stephen C. Meyer, The origin of biological information and the higher taxonomic
categories, Proceedings of the Biological Society of Washington June 2004, vol. 117, No. 2, s.
213-239; Michael J. Denton and Craig J. Marshall, The Laws of Form Revisited, Nature 22
March 2001, vol. 410, s. 417; Michael J. Denton, Craig J. Marshall and Michael Legge, The
Protein Folds as Platonic Forms: New Support for the pre-Darwinian Conception of
Evolution by Natural Law, Journal of Theoretical Biology 7 December 2002, vol. 219, s. 325-
342; David K.Y. Chiu and Thomas H. Lui, Integrated Use of Multiple Interdependent
Patterns for Biomolecular Sequence Analysis, International Journal of Fuzzy Systems
September 2002, vol. 4, No. 3, s. 766-775.
[59] Zob. Alison Abbott, Axeing of website article sparks row at Max Planck, Nature 13 April
2003, vol. 422, s. 460 i Ulrich Kutschera, Designer scientific literature, Nature 8 May 2003, vol.
423, s. 116.
[60] Na temat innych analiz molekularnych systemów z perspektywy kreacjonistycznej patrz:
Michael J. Behe, Biologiczne mechanizmy molekularne. Eksperymentalne poparcie dla
kreacjonizmu, fragment książki Kazimierza Jodkowskiego, Metodologiczne aspekty…, s. 496-
511. (Oryginał: Michael J. Behe, Molecular Machines: Experimental Support for the Design
Inference; Jerry Bergman, ATP: The Perfect Energy Currency for the Cell, Creation
Research Society Quarterly June 1999, vol. 31, No. 1, s. 2-9; Michael J. Behe, Experimental
Support for Regarding Functional Classes of Proteins to be Highly Isolated from Each Other,
The Weekly Standard 7 June 1999, BOOKS & ARTS, s. 35; Joseph W. Francis, Peering into
Darwin’s Black Box: The cell divsion processes required for bacterial life, Origins & Design
2000, vol. 20, No. 1; Jonathan Sarfati, Decoding and editing design: double sieve enzymes,
Creation Ex Nihilo Technical Journal 1999, vol. 13, No. 1, s. 5-7; Jonathan Sarfati, Lobster
eyes — brilliant geometric design, Creation June 2001, vol. 23, No. 3, s. 12-13; Sean Henahan,
ATP Synthase Molecule; Molecular Machines, Index of Illustrations, Graphics, and
Animations; Mike Gene, Irreducible Complexity ReVisited, TeleoLogic, Background
Considerations 5.11.2002, No. 3; Mike Gene, A Teleological Hypothesis Regarding the

Degradosome Machine, TeleoLogic, Biotic Reality, No. 2; Mike Gene, Tubulin and ftsZ: More
than One Way to View Something, Biotic Reality, No 18.
[61] Np. jeszcze mniejszego od bakteryjnego silnika protonowego mechanizmu zwanego F1-
ATPase będącego częścią enzymu ATPazy i najmniejszym znanym tego typu rotacyjnym
silniczkiem w naturze. Jego wymiary wynoszą 10 nanometrów szerokości i 8 nanometrów
wysokości. Zob. Piotr Lenartowicz SJ, Najmniejszy silniczek wszechświata, Na Początku…
czerwiec 1997, nr 6 (87), s. 141-142; Hiroyuki Noji, Ryohei Yasuda, Masasuke Yoshida and
Kazuhiko Kinosita, Direct observation of the rotation of F1-ATPase, Nature 20 March 1997,
vol. 386, s. 299-302 i komentarz do tego tekstu: Steven M. Block, Real engines of creation,
Nature 20 March 1997, vol. 386, s. 217-219; Ryohei Yasuda, Hiroyuki Noji, Masasuke Yoshida,
Kazuhiko Kinosita, Hiroyasu Itoh, Resolution of distinct rotational substeps by
submillisecond kinetic analysis of F1-ATPase, Nature 19 April 2001, vol. 410, s. 898-904;
Jonathan D. Sarfati, Design in Living Organisms: Motors, Creation Ex Nihilo Technical
Journal 1998, vol. 12, No. 1, s. 3-5.
Źródło: Na Początku… wrzesień-październik 2003, nr 9-10 (172-173), s. 328-342. Ostatni
dostęp do linków podanych w artykule: 27 października 2004 r.
Document Outline
- http://bioslawek.wordpress.com/2011/07/17/137/#more-137
- ‘Marzenie nanotechnologów: silnik protonowy bakterii Escherichia coli’
Wyszukiwarka
Podobne podstrony:
więcej podobnych podstron