Hormony roślinne:
Mogą być wydzielane:
- endokrynowo – wydzielanie na zewnątrz komórki
-parakrynowo – do wnętrzna komórki
Podobieństwa
- wszystkie hormony działają w niskich stężeniach
- miejscowe stężenie
- muszą mieć receptor (zwykle białko integralne plazmalemmy)
- nie są białkami/peptydami (w przeciwieństwie do zwierząt), są to proste związki, metabolity wtórne
Różnice:
- plejotropiczne działanie – regulują wiele funkcji (plejotropowe są hormony roślinne – regulują one
wiele funkcji, wiele procesów naraz za pomocą jednego hormonu)
- nie wszystkie rośliny mają wszystkie hormony (mogą nie mieć giberelin, za to wszystkie mają
cytokininy i auksyny. Są one niezbędne do zakończenia cyklu życiowego)
Auksyna - hormon wzrostu
Slajd 1
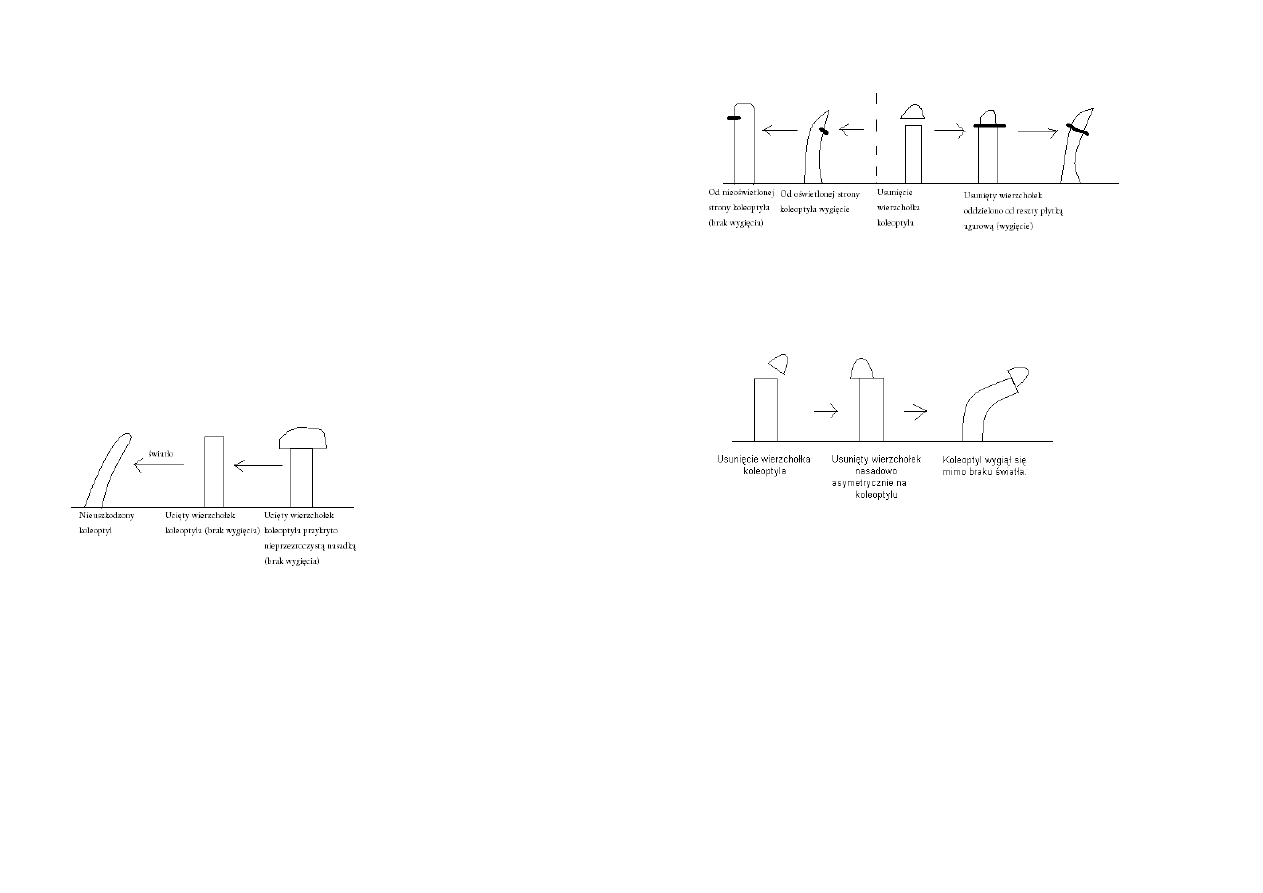
Darwin
Karol i Franciszek Darwinowie badali ruchy fototropiczne koleoptyli mozgi kanaryjskiej.
Wniosek: Czynnik wzrostu produkowany w wierzchołku koleoptyla przemieszcza się do strefy
wzrostu.
Boysen-Jensen (1913)
Wniosek:
Boysen-Jensen wykazał że czynnik stymulujący wzrost przemieszcza się przez agar, ale nie przenika
przez nieprzepuszczalne dla wody materiały.
Paal ustalił, że substancja stymulująca wzrost koleoptyla jest związkiem chemicznym.
Slajd 2
Test wygięciowy koleoptyla owsa - Went (1926)

Went wykazał, że substancja stymulująca wzrost może dyfundować przez bloczki agarowe.
Opracował on też test do oznaczania stężeń substancji wzrostowej (auksyny) w soku roślin.
Identyfikacja auksyny (IAA) dokonali w latach 30-stych XX wieku Kögl i Haagen-Smit (Holandia)
oraz Thimann (USA).
Slajd 3
Fragmenty koleoptyli inkubowano przez 18 h w wodzie (A) i roztworze auksyny (B). Wewnątrz
półprzezroczystego koleoptyla widoczny jest liść.
Slajd 4
Auksyny naturalne
- kwas indolilo-3-octowy (IAA)
- kwas 4-chloroindolilo-3-octowy (4-Cl-IAA)
- kwas indolilo-3-masłowy (IBA)
- kwas chloroindolilo-4-octowy
Slajd 5
Auksyny syntetyczne
- kwas 2,4-dichlorofenoksyoctowy (2,4-D)
- kwas 2-metoksy-3,6-dichlorobenzoesowy (dicamba)
Slajd 6
Aktywność auksynowa
Wszystkie związki wykazujące aktywność auksynową mają charakterystyczny układ ładunków: grupa
karboksylowa ma ładunek ujemny a pierścień aromatyczny ładunek dodatni. Odległość pomiędzy
naładowanymi grupami wynosi 0,5 mm
Slajd 7
Miejscem syntezy IAA są merystemy i szybko rosnące tkanki, szczególnie pędu.
Kumulacja auksyn w zawiązkach liści Arabidopsis:
Gen receptorowy GUS (
β-glukuronidazy
) przyłączono do sekwencji promotorowej DNA (promotor
DR5) odpowiedzialnego za syntezę auksyny i transformowano używając do tego konstruktu liście
Arabidopsis za pomocą plazmidu Ti Agrobacterium tumefaciens (transformacja
wektorowa).
We wczesnym stadium rozwoju hydatody miejsce syntezy auksyn (niebieska plama) zaznaczona
strzałką. Wytworzona auksyna przemieszcza się w kierunku różnicujących się tkanek przewodzących
(grot strzałki).
Slajd 8
B
iosynteza IAA niezależna od tryptofanu: IGP
→
IPA
→
IAA
Trzeba poszukać:D
Indolilo-3-glicerofosforan (IGP)
Slajd 9
Szlaki biosyntezy IAA zależne od tryptofanu
TAM i IPA są głównymi szlakami biosyntezy auksyn u roślin. IAN charakterystyczny jest tylko dla
trzech rodzin: Brassicaceae, Poaceae, Musaceae.
TAM – ważnym materiałem pośrednim jest tryptofan. Szlak prowadzi do IAA
IAN – ważnymi enzymami są nitrylazo-1,3-myrozynazy
IAM – kwas indolilo-3-pirogronowy (IPA). Enzym – meroksydaza tryptofanowa (występuje u
prokaryota, brak u roślin)
Tryptomina – jeden z najważniejszych związków podczas syntezy.
Też trzeba poszukać
IAM uczestniczy w syntezie IAA głównie w kom. bakterii patogenicznych, np. Agrobacterium
tumefaciens czy Pseudomonas sarastanoi.
Slajd 10
Koniugacja i degradacja IAA
- reakcje odwracalne (magazynowanie – istotny jest inozytol)
- reakcje niedowracalne (degradacje)
Służy to do zniwelowania aktywności biologicznej IAA
Metabolizm konjugatów auksyn jest głównym regulatorem poziomu wolnych auksyn np. IAA-mezo-
inozytol w kiełkujących ziarnach kukurydzy przemieszcza się z endospermy nasienia via floem do
wierzchołka koleoptyla gdzie następuje hydroliza i uwolnienia IAA.
Slajd 11
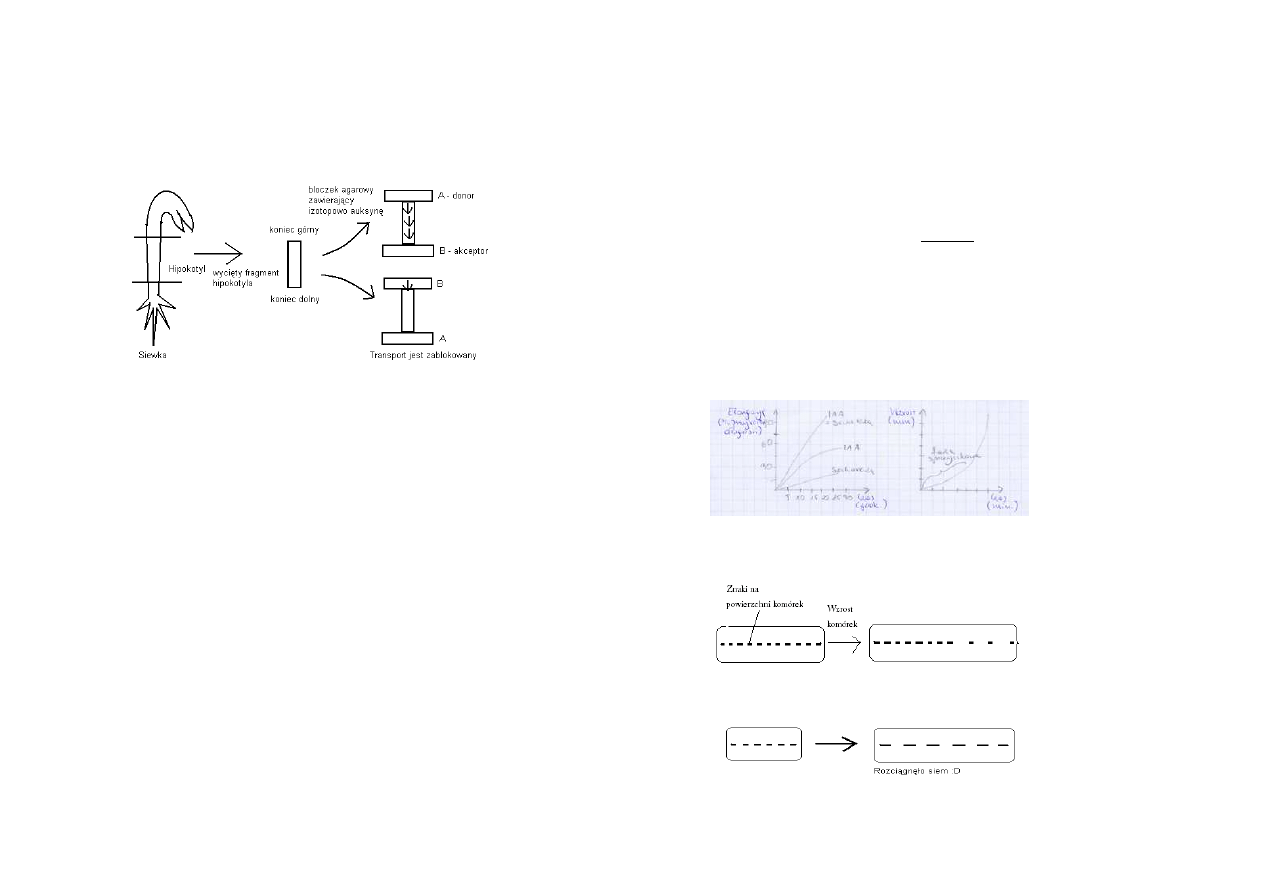
Polarny transport auksyn
- bazypetalnie (od wierzchołka do podstawy, szybki, aktywny) – w komórkach parechymatycznych
wiązek sitowo – naczyniowych pędu
- akropetalny (od podstawy do wierzchołka, wolny, na zasadzie dyfuzji) – we floemie korzenia
Szybkość transportu 2-20 cm/h
Slajd 12
Chemiosmotyczny model polarnego transportu auksyn w nierosnących komórkach
1. IAA przenika przez błonę komórkową na drodze dyfuzji (forma niezjonowana IAAH, zależna
od gradientu stężenia) lub transportowany jest przez specyficzny nośnik (forma zjonowana IAA)
2. W apoplaście odczyn kwaśny (pH=5-5,5) utrzymywany jest dzięki funkcjonowaniu H+-
ATPazy błonowej. Kierunek transportu auksyny ↓
3. W cytoplaźmie (pH=7) dominuje forma anionowa (IAA
-
)
4. IAA
-
transportowany jest z komórki do apoplastu przez inny nośnik zlokalizowany w bazalnej
części komórek. W apoplaście 25% IAA (pKa= 4,75) występuje w formie niezdysocjowanej.
Slajd 13
Dwa typy białek nośnikowych transportujących IAA w komórce roślinnej (Arabidopsis)
1.
P
IN są integralnymi białkami błony komórkowej, współdziałają w transporcie z innymi
białkami. Odpowiadają za napływ IAA do komórki
PIN1 – jest odpowiedzialne za recyrkulację auksyn w merystemach wierzchołkowych pędu,
pośredniczy w transporcie wertykalnym auksyny od wierzchołka wzrostu łodygi w kierunku korzenia
PIN3 – transport boczny IAA spowrotem do tkanek przewodzących
2.
P
GP
PGP1 – wielokierunkowy transport auksyny w wierzchołku korzenia oraz w merystemie
wierzchołkowym pędu. PGP są zależnymi od ATP białkami wielolekowej odporności (ang. multidrug
resistance / p-glycoprotein) (MDR/PGP). Integralne białka błony komórkowej, transportują
hydrofobowe aniony. PGP – wypływ IAA z komórki.
Slajd 14
Model polarnego transportu w małych??? Komórkach merystematycznych
1.
H+-ATPaza w błonie komórkowej pompuje protony do apoplastu. Zakwaszenie apoplastu
wpływa na szybkość transportu poprzez zmianę proporcji IAAH do IAA-
2.
IAAH może wnikać do komórek przez symporter protonowy taki jak AUX1 lub na drodze
dyfuzji. W cytozolu IAA staje się anionem i może przenikać na zewnątrz komórki tylko
poprzez transport aktywny
3.
PGP są zlokalizowane w błonie kom. niepolarnie i mogą transportować aktywnie (zależnie od
ATP) auksyny na zewnątrz komórki.
4.
Synergistycznie wzmocniony transport aktywny występuje wtedy, gdy polarnie
rozmieszczone białko PIN związane jest z białkiem PGP, które przeciwdziałają dyfuzji
zwrotnej IAA.
Slajd 15
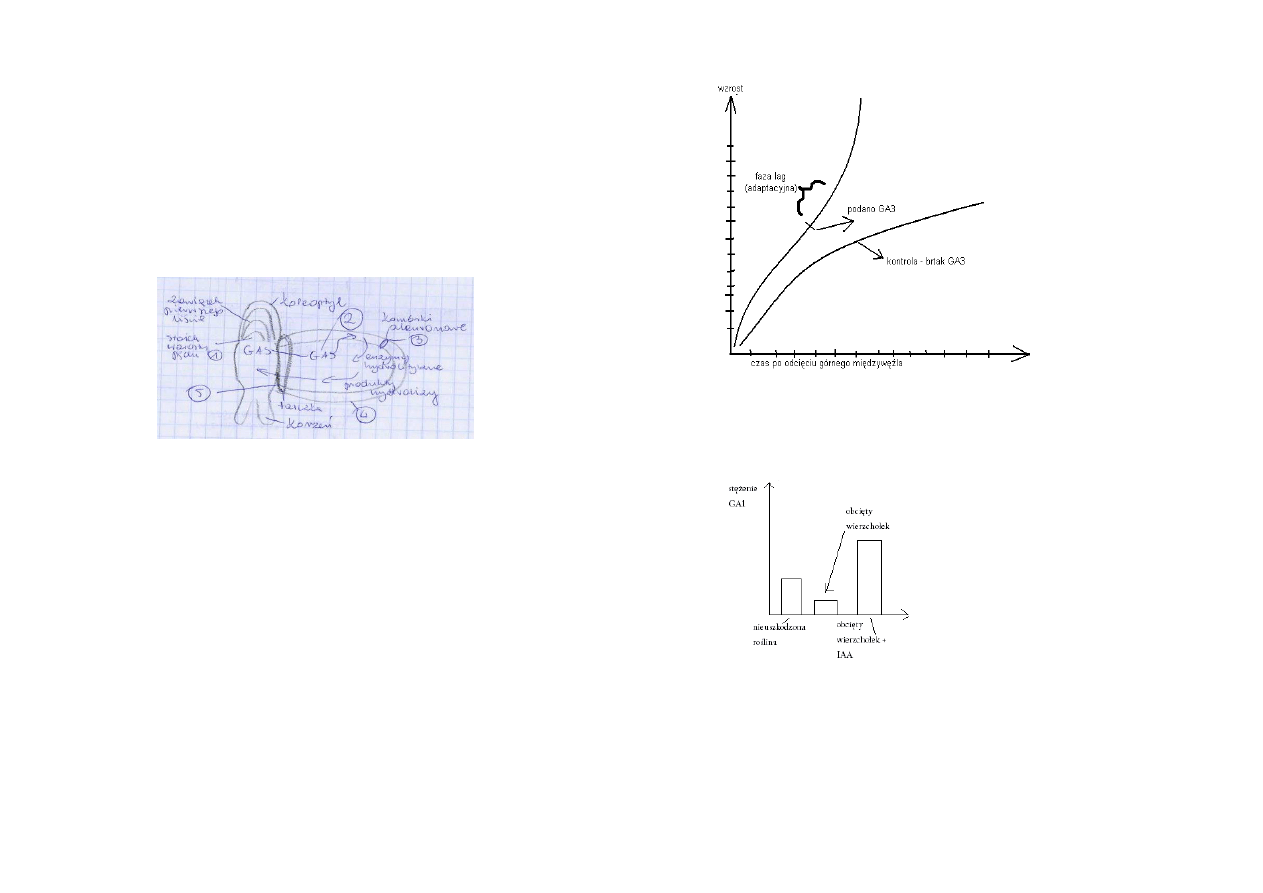
Zależny od auksyny przyrost długości odcinków koleoptyla owsa
Dodatek sacharozy wydłuża czas działąnia auksyn, pełniąc głównie rolę substancji osmotycznie
czynnej zapewniającej odpowiednie ciśnienie turgorowe w trakcie elongacji (może być zastąpiona
KCl)
Zauważ 15 min fazę opóźnienia w odpowiedzi wzrostowej koleoptyli na działanie auksyny (inset).
Slajd 16
Wzrost wierzchołkowy i dyfuzja w korzeniu
Struktura rosnąca wierzchołkowo:
Struktura rosnąca dyfuzyjnie:
S. rosnąca wierzchołkowo – pojedyncze komórki opuszczające roślinę
- łagiewka pyłkowa, protonema (splątek), włośnik
S. rosnąca dyfuzyjnie – wszystko co wewnątrz organizmu, dominujący typ wzrostu
Slajd 17
Ułożenie nowo syntetyzowanych mikrofibryl determinuje kierunek wzrostu korzenia
−
przypadkowe ułożenie mikrofibryli celulozowych
−
poprzeczne ułożenie mikrofibryl celulozowych
Slajd 18
Graficzna prezentacja dwóch równań wiążących proces pobierania wody i powiększanie
komórki z ciśnieniem turgorowym i potencjałem wody.
Komórka rośnie, napiera na ściany, rośnie potencjał, komórka pobiera intensywnie wodę
Slajd 19
Redukcja ciśnienia turgorowego (P) w komórkach wycinków łodygi groszku jest wynikiem
relaksacji (ustąpienia) stresu (napięcia, naprężenia) ścian komórkowych
Auksyna nie obniża wartości progowej, ale wpływa na stan relaksacji ściany komórkowej. Luzuje
mikrofibrylle w ścianie kom, żeby komórka mogła rosnąć, pobudza ją do wzrostu.
Slajd 20
Kwasowa indukcja nieodwracalnego powiększenia izolowanych ścian komórkowych
Środowisko kwaśne luzuje ścianę komórkową, sprzyja wzrostowi; pH = 5,5
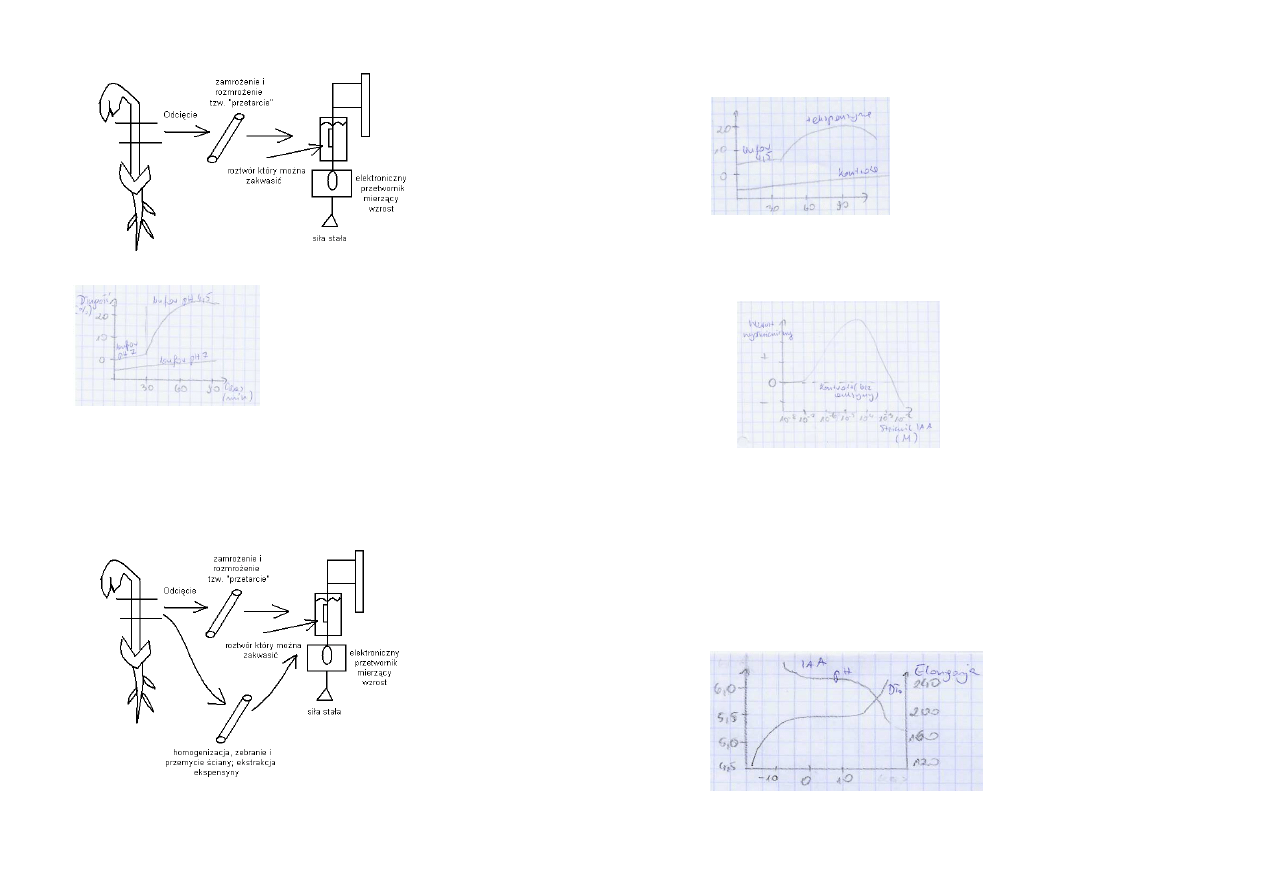
Środowisko kwaśne sprzyja wzrostowi
Slajd 21
Przywracanie właśności rozciągliwości ściany komórkowej pod wpływem ekspansyn
Arabidopsis – 2 rodziny ekspansyn
α-ekspansyny (EXPA)
β-ekspansyny (EXPB)
Slajd 22
Typowa zależność intensywności wzrostu wydłużeniowego wycinków młodych łodyg owsa lub
koleoptyli kukurydzy od stężenia auksyny.
Slajd 23
Pięć warunków potwierdzających słuszność „teorii kwasowego wzrostu”:
1.
Kwaśne bufory (źródło protonów) stymulują przejściowo wzrost wydłużeniowy.
2.
Auksyna poprzez indukcję wydzielania protonów zakwasza ścianę komórkową, ponadto
oistnieje ścisłe podobieństwo kinetyki zakwaszania i wzrostu wydłużeniowego.
3.
Bufory o pH obojętnym hamują wzrost indukowany przez auksynę.
4.
Stymulatory (inne niż auksyna) wydzielania protonów powinny stymulować wzrost.
5.
W ścianie komórkowej obecny jest „czynnik luzujący ścianę” wykazujący optimum działania
w pH kwaśnym.
Slajd 24
Kinetyka Indukowanego przez auksyny wzostu wydłużeniowego i zakwaszenia

Slajd 25
Model indukowanego przsz IAA wydzielania jonów H+
1.
Aktywacja genów: auksyna wiąże się z kompleksem białkowym, który zawiera receptor
auksyny i represor transkrypcji genu H+-ATPazy, wyznakowując jednocześnie represor do
degradacji proteolitycznej w proteosomie. Wzrost poziomu transkrypcji, translacji i
wzmożona sekrecja powoduje zwiększenie ilości H+-ATPazy w błonie komórkowej
2.
Sekrecja białek: białko wiążące auksyny – ABP1 zwiększa sekrecję H+-ATPazy.
3.
Stabilizacja H+-ATPazy i podanie auksyny powoduje zatrzymanie H+-ATPazy w błonie
komórkowej. Proces ten może być redukowany przez białko ABP1.
Slajd 26
Receptor auksyn
Białko TIR1 stanowi podjednostkę kompleksu ligazy ubiknitynowej E3 (SCF
TIR1
). Wiązanie auksyny
z kompleksem SCF
TIR1
powoduje jego aktywację, a następnie ubikwitynację i proteolizę receptora
AUX/IAA.
AUX – receptor, póki nie będzie wycięty będzie powodował represję auksyny (?)
Slajd 27
Zależna od ubikwityny degradacja białka w cytozolu.
Do aktywacji ubikwityny katalizowanej przez E1 potrzebny jest ATP. E1 przenosi ubikwitynę na E2.
Ligaza E3 łączy ubikwitynę z docelowym białkiem przeznaczonym do degradacji. Wyznakowane
białko hydrolizowane jest w proteosomie 26S.
Slajd 28
Auksyny regulują procesy rozwoju roślin:
•
regulują dominację wierzchołkową
- antagonizm z ABA
•
regulują tworzenie zawiązków kwiatów i liśc i oraz ich rozmieszczenia (filotaksja)
- mutant Arabidopsis pin1 tworzy nienormalne kwiaty i liście
•
indukcja procesów różnicowania tkanek przewodzących
- wysokie stężenie IAA → różnicowanie ksylemu i floemu
- niskie stężenie IAA → różnicowanie tylko floemu
•
opóźnienie opadania liści
- antagonizm z etylenem
•
regulacja dojrzewania owoców
•
stosowanie syntetycznych auksyn
- partenokarpia – wynik działania auksyn na niezapylone kwiaty
- heribicydy – mniejsza wrażliwość jednoliściennych na auksyny
Gibereliny – regulatory wysokości roślin i kiełkowania
Slajd 29
Jak to się zaczęło...
- od stuleci hodowcy ryżu znali chorobę ryżu bakanae (jap. Zwariowany ryż) powodowaną przez
patogenicznego grzyba Gibberella fujikuroi
- w latach 30-tych XXw. Japończycy wyizolowali z filtratów pohodowlanych grzyba substancję
stymulującą wzrost roślin, którą nazwano gibereliną A. Obecnie znanych jest ok. 130 giberelin
- w latach 50-tych XX wieku wyizolowano i oczyszczono z filtratów pierwsze trzy gibereliny: kwas
giberelowy (UK, USA) oraz giberelinę A
1
, A
2
, A
3
(Japonia).
- kolejno odkrywane (struktura + pochodzenie) gibereliny otrzymują kolejne numery (GA
1
– Ga
n
)
GA
3
to kwas giberelowy, honorujący odkrywców.
- obecnie znanych jest 130 giberelin.
Slajd 30
1.
Efekt działania GA1 na kukurydzę typu dzikiego i mutanta DWARF (d1)
Giberelina bardzo mocno stymuluje wzrost wydłużeniowy łodygi mutanta dwarf (karzeł). Wielkość
rośliny typu dzikiego po podaniu GA
1
zmienia się w bardzo niewielkim stopniu.
Slajd 31
Kapusta, roślina dnia długiego, tworzy rozety jeśli uprawiana jest w warunkach dnia krótkiego.
Podanie GA3 powoduje wybijanie długiego pędu kwiatowego (nawet 3m)
Slajd 32
Struktura giberelin
Wszystkie gibereliny to pochodne diterpenu – ent-giberelanu – zbudowanego z czterech pierścieni
zawierających łącznie 20-30 atomów C.
Uwaga: przedrostek ent oznacza pochodzenie od ent-kaurenu – tetracyklicznego węglowodoru
będącego enancjomerem występującego w naturze kaurenu.
Slajd 33
Gibereliny regulują procesy wzrostu i rozwoju
•
Stymulacja internodialnego wzrostu pędu
- rośliny genetycznie karłowate (Brassicaceae)
- liczne trawy (Poaceae)
•
Przejście z fazy wzrostu juwenilnego do dojrzałego
−
skracanie fazy juwenilnej szpilkowycj z nawet 20 do 2-3 lat
•
Determinacja (genetycznie uwarunkowanej) płci
- udział w mediacji działania fotoperiodu i składników odżywczych
*jednoliścienne (kukurydza): hamowanie rozwoju męskich i stymulacja żeńskich
kwiatów
*dwuliścienne (ogórek): efek t działania odwrotny
•
Przyspieszanie rozwoju pyłku i wzrostu łagiewki pyłkowej
•
Przyspieszanie rozwoju owoców po zapłodnieniu oraz partenokarpii
- zastosowanie komercyjne np. plantacja winorośli.
•
Przyspieszenie rozwoju nasion i kiełkowania
•
Stosowanie syntetycznych inhibitorów GA
S
ogranicza wzrost.
- hodowle szklarniowe wielu roślin ozdobnych (lilie, chryzantemy, poinsecje)
- cycocel, inhibitor wzrostu pędu zbóż stosowany w UE, unikanie zjawiska lodging.
Lodging – roślina wybujała, z dużym kłosem przed zbiorem, często wylega. Gibereliny zapobiegają
temu, skracają pęd, zapobiegają wyleganiu
Slajd 34
Biosynteza giberelin:
Szlak MVA – cytoplazma, kwas mewalonowy - powstają brasinosteroidy, sterole
Szlak MEP – stroma plastydów, fosforan metyloerytriolu - powstają: GA12 → inne gibereliny fitol
(C20), fitoen (C40) → karotenoidy
Plastydy – w pierwszym etapie pirofosforan izopentylu przekształcany jest w pirofosforan
geranylogeranylu (GGPP), gdzie następnie ulega kolejnym reakcjom cyklizacji, w wyniku których
powstaje ent-kauren
Otoczka plastydów lub siateczka śródplazmatyczna – podczas tego etapu ent-kauren przekształcany
jest do GA
12 .
U większości roślin GA
12
przekształcany jest dalej do GA
53
(hydroksylacja w pozycji
C13).
Cytoplazma – na trzecim etapie biosyntezy, przebiegającymw cytozolu, z GA12 lub GA53, powstają
pozostałe gibereliny. Etap obejmuje reakcje hydroksylacj i utleniania katalizowane przez
dioksygenazy
Slajd 35
GA1 i GA4 są stymulatorami wzrostu dlatego ich prekursory i inne metabolity są nieaktywne.
Mutanty grochu różniące się zawartością GA1 w tkankach wegetatywnych.
GA20 → GA1 → GA8.
Super karzeł: brak GA, nana
Karzeł: zawiera GA20 oraz śladowe ilości GA1, NALE
Wysoki: GA1 (decydujące o wzroście grochu), NALE
Smukły: więcej GA1m NALE sln
LE i le – allele genu wysokości (koduje enzym hydroksylujący GA20 w pozycji 3
β. Mutant nana –
synteza GA zablokowana na etapie KAO (oksydaza ent-kaurenu).
Wszystkie allele są homozygotami
Slajd 36
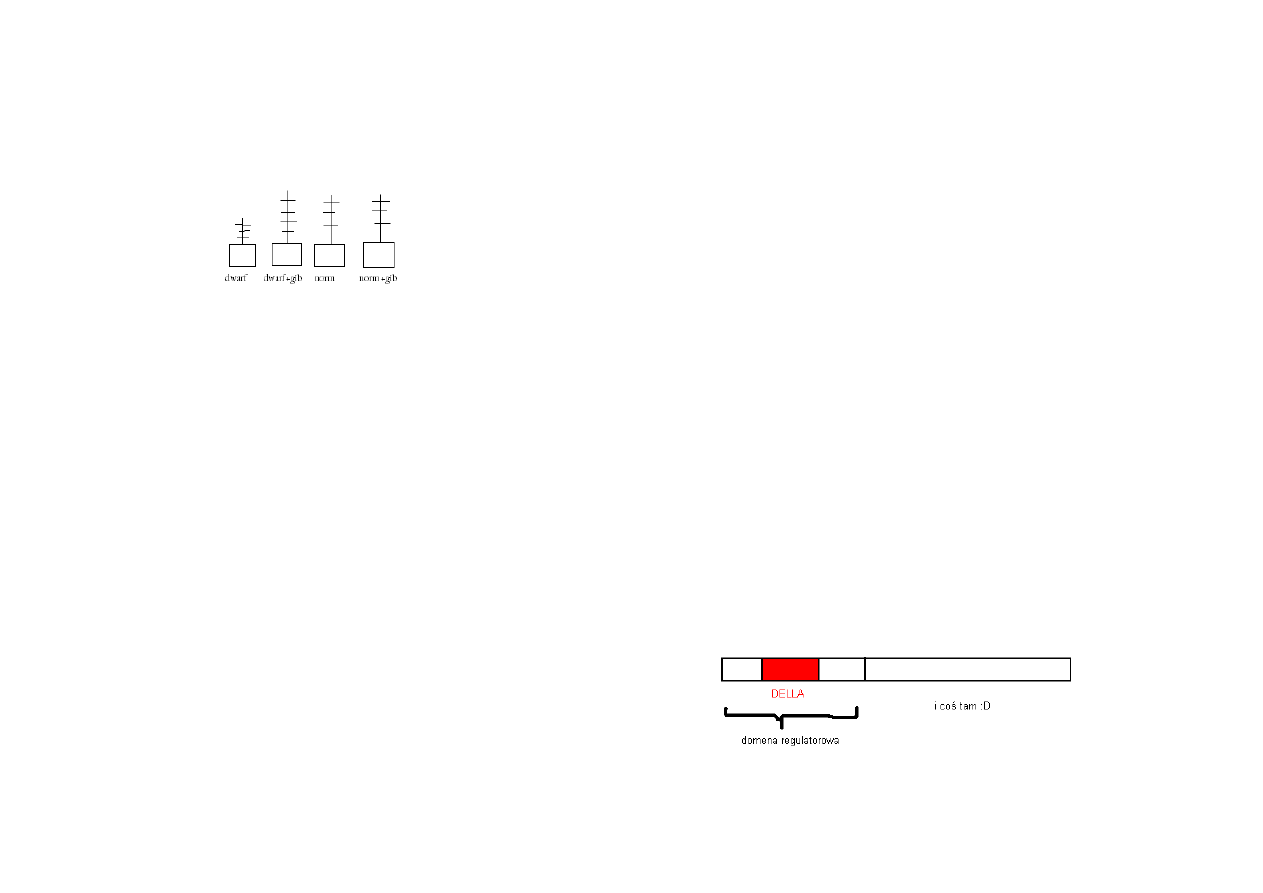
Struktura białka represorowego RGA lub GAI
Slajd 37
Model regulacji ekspresji genów wzrostu ryżu:
Podczas braku giberelin domena DELLA białka represorowego SLR blokuje transkrypcję genów
przez gA
Slajd 38
Struktura ziarniaka jęczmienia, funkcje różnych tkanek podczas kiełkowania:
1.
Gibereliny syntetyzowane sa w zarodku, a nastepnie dyfundują przez tarczkę do skrobiowej
części bielma
2.
Gibereliny dyfundują do warstwy aleuronowej
3.
W komórkach warstwy aleuronowej zachodzi synteza α – amylazy i innych hydrolaz, które
wydzielane są do bielma.
4.
Skrobia i inne makrocząsteczki są hydrolizowane
5.
Produkty hydrolizy makrocząsteczek bielma są wchłaniane przez tarczkę i transportowane do
rozwijającego się zarodka.
Slajd 39
Uniwersalne wtórne przekaźniki w komórkach eukariitycznych:
3`, 5` - cykliczny AMP
3`,5` - cykliczny GMP
Cykliczna ADP-ryboza (cADPR)
Jon wapnia
Tlenek azotu
Inozytolo-1,4,5-trifosforan (IP3)
1,2-diacyloglicerol (DAG)
Slajd 40
Struktura kalmoduliny
Podczas asocjacji kompleks kalmodulina-Ca2+ „owija się” wokół białkoa docelowego zmieniając jego
konformację. Kolmodulina występuje tylko w kom. Eukaryota i posiada na końcach cząsteczki dwie
globularne domeny oddzielone elastyczną helisą-α. Na obydwu globularnych końcach kalmoduliny
znajdują sie po dwa miejsca wiążące jony wapnia.
Slajd 41
GA indukuje syntezę α-amylazy w warstwie aleuronowej ziemniaków
Slajd 42
Gibereliny stymulują wzrost wydłużeniowy górnego międzywęźla łodygi ryżu
Slajd 43
Auksyny stymulują biosyntezę giberelin
Ucięcie wierzchołka łodygi grochu powoduje zmniejszenie stężenia GA1.
Podanie IAA niweluje ten efekt.
Slajd 44
Auksyny stymulują biosyntezę giberelin
IAA (powstający w wierzchołku) stymuluje syntezę GA1 w międzywęźlach grochu.
Kwas indolilo-3-octowy hamuje również degradację GA1.
Slajd 46
Slajd 47
IAA
GA
GR=m(
Ψ
p-Y)
m (rozluźnienie ścian)
Ψp
Y
+
-
+/-
-
-
+
Zakwaszenie ściany kom.
+
-
Aktywność ekspansyn
+
-
Aktywność XET
-
+
Czas ujawnienia efektu
Pierwotny
wtórny
10-12min (koleoptyl Avena)
30-60min(hypokotyl Glycina)
40min (pęd Oryza sativa)
2-3 godziny
Stężenie stymulujące
10
-5
– 10
-6
M (pędy)
10
-9
– 10
-10
M (korzenie,
blokada syntezy etylenu)
10
-10
M (pędy Oryza sativa)
10
-14
M (GA73, anteridia
[plemnia], lygopodium
japonicum [paproć], w
ciemności)
Cykl komórkowy
G1/S (synteza DNA)
G2/M
Slajd 48
Rearanżacja ksyloglukanów
Wyszukiwarka
Podobne podstrony:
więcej podobnych podstron