221
Profile bia³kowe antygenów powierzchniowych Malassezia pachydermatis
Protein profiles of superficial antigens M. pachydermatis strains
Gra¿yna Zió³kowska, Aneta Nowakiewicz
Zak³ad Mikrobiologii Weterynaryjnej, Instytut Biologicznych Podstaw Chorób Zwierz¹t
Wydzia³ Medycyny Weterynaryjnej AR w Lublinie
Wprowadzenie: Malassezia pachydermatis nale¿y do powszechnie wystêpuj¹cych komensali skóry i b³on œluzowych, w pewnych sytuacjach mo¿e
jednak odpowiadaæ za procesy infekcyjne, m.in. u psów zapalenie zewnêtrznego kana³u s³uchowego (otitis) lub zapalenia skóry (dermatitis).
W okreœleniu w³aœciwoœci patogennych dro¿d¿aka mo¿e byæ pomocna analiza porównawcza struktury antygenowej szczepów izolowanych od
zwierz¹t zdrowych klinicznie i z klinicznymi objawami infekcji.
Cel pracy: Uzyskanie i standaryzacja procedury otrzymywania antygenów powierzchniowych M. pachydermatis, okreœlenie ich profili bia³kowych
oraz analiza porównawcza antygenów powierzchniowych szczepów izolowanych od psów z objawami zapalenia zewnêtrznego kana³u s³uchowego
i zdrowych klinicznie.
Materia³ i metody: Badaniami objêto antygeny powierzchniowe szczepów M. pachydermatis, izolowanych od psów zdrowych (n=20) oraz z klinicz-
nymi objawami otitis externa (n=20). Preparaty antygenowe uzyskiwano z 3-dniowej hodowli poszczególnych szczepów, eluowanych wodnym
roztworem merthiolatu przez 1, 3 i 24 godz. Profile bia³kowe preparatów okreœlano na podstawie rozdzia³u elektroforetycznego materia³u w warun-
kach zdenaturowanych (SDS-PAGE) oraz analizy elektroforegramów z zastosowaniem Gel Doc 2000.
Wyniki: Wykazano wysoki stopieñ zró¿nicowania i z³o¿on¹ strukturê poszczególnych preparatów antygenowych. Szczepy izolowane od zwierz¹t
chorych charakteryzowa³y siê wyraŸnie silniejsz¹ ekspresj¹ bia³kowych antygenów powierzchniowych i wiêkszym ich zró¿nicowaniem (œrednia
liczba frakcji wynosi³a 11), w porównaniu ze szczepami komensalicznymi, u których œrednia liczba frakcji wynosi³a 5. Antygeny powierzchniowe
badanych szczepów wykazywa³y obecnoœæ frakcji wspólnych: 68-71 kDa, 57-58 kDa, 45-46 kDa. Stwierdzono ponadto frakcje swoiste, typowe
wy³¹cznie dla szczepów pochodz¹cych od psów z otitis externa; by³y to: 61-64 kDa, 52-53 kDa, 43-44 kDa, 38-39 kDa i 15-17 kDa (tab. II). W pro-
filach bia³kowych antygenów powierzchniowych „patogennych” szczepów M. pachydermatis stwierdzono ponadto wy¿sz¹ czêstotliwoœæ wystêpo-
wania frakcji: 59-60 kDa, 55-56 kDa, 49-50 kDa, 36-37 kDa, 32-33 kDa, 27-28 kDa, 25-26 kDa, 22-23 kDa, 18-20 kDa i 13-14 kDa. Przyjmuj¹c
50% wystêpowania danej frakcji za wartoœæ graniczn¹, obecnoœæ pr¹¿ków 55-56 kDa, 36-37 kDa, 27-28 kDa i 18-19 kDa mo¿e, w po³¹czeniu
z obecnoœci¹ frakcji typowo swoistych, wskazywaæ na patogennoœæ danego szczepu M. pachydermatis.
Wnioski: 1. Szczepy M. pachydermatis izolowane od chorych zwierz¹t charakteryzuj¹ siê wiêksz¹ ekspresj¹ bia³ek. 2. Profile bia³kowe antygenów
powierzchniowych M. pachydermatis cechuje obecnoœæ frakcji typowych wy³¹cznie dla szczepów izolowanych od psów chorych i frakcji wystêpuj¹-
cych u znacznego odsetka szczepów „patogennych”. 3. Korelacja miêdzy pochodzeniem szczepów M. pachydermatis a obecnoœci¹ okreœlonych
frakcji antygenowych w ich profilach bia³kowych sugeruje mo¿liwoœæ wystêpowania bia³kowych markerów patogennoœci u tych grzybów.
S³owa kluczowe: Malassezia pachydermatis, zapalenie zewnêtrznego kana³u s³uchowego, profile bia³kowe
Introduction: Malassezia pachydermatis is one of common commensals of skin and mucous membranes, but in some cases it can cause infec-
tions, such as otitis externa or dermatitis, especially in dogs. Using of comparative analysis of antigenic structure of strains obtained from the
healthy dogs and from those with the clinical symptoms of infection, can be helpful in evaluating the pathogenic features of this yeast.
Aim of study: Obtaining and standardization of the conditions for the surface antigens M. pachydermatis isolation, followed by the protein profiles
analysis of strains obtained from the healthy dogs and from those with the clinical symptoms of otitis externa.
Material and methods: The research material involved the surface antigens of M. pachydermatis strains obtained from the healthy dogs (n=20)
and from those with the clinical symptoms of otitis externa recognized (n=20). The antigenic preparations were obtained from a 3-day culture of
each strain, subjected to the elution with a merthyolate water solution for 1,3 and 24 h. The protein profile preparations were determined on the
grounds of the material electrophoretic separation at the denaturation (SDS-PAGE) and the electrophoregrams analysis with Gel Doc 2000.
Results: The high diversity and complex structure of individual antigen preparations were demonstrated. The strains isolated from the diseased
animals, exhibited a clear stronger expression of surface protein antigens and their higher diversity (amount of fractions was averaged 11) compa-
rison with commensalic strains in which the amount of fraction was averaged 5. Superficial antigens of investigated strains showed the presence
common fractions: 68-71 kDa, 57-58 kDa, 45-46 kDa. In addition a higher frequency of typical fractions for protein profiles of superficial antigens
of M. pachydermatis pathogenic strains was showed 61-64 kDa, 52-53 kDa, 43-44 kDa, 38-39 kDa i 15-17 kDa (tab. II) whereas their presence
wasn’t demonstrated for superficial antigens of commensalic strains. Besides, the protein profiles of superficial antigens of “pathogenic” strains
were characterized, in relation to commensalic strains, by increased frequency of the following fractions: 59-60 kDa, 55-56 kDa, 49-50 kDa, 36-
37 kDa, 32-33 kDa, 27-28 kDa, 25-26 kDa, 22-23 kDa, 18-20 kDa i 13-14 kDa. Assuming that 50% of a specific fraction is a terminal value, the
presence of bands 55-56 kDa, 36-37 kDa, 27-28 kDa i 18-19 kDa can possibly, in combination with the presence of typical fractions, suggest
pathogenicity of specific M. pachydermatis strain.
Conclusions: 1. M. pachydermatis strains obtained from diseased animals are marked by high protein expression. 2. Protein profiles of superficial
antigens are characterized by the presence of fractions found in strains isolated from diseased dogs and fractions found statistically significant in
pathogenic strains. 3. The correlation between the origin of the strains and the presence of a given antigenic fraction in its protein profile can
suggest a possibility of protein markers of pathogenicity in these fungi.
Key words: Malassezia pachydermatis, otitis externa, protein profiles
Adres do korespondencji:
Dr hab. Gra¿yna Zió³kowska, prof. nadzw. AR; Zak³ad Mikrobiologii Weterynaryjnej, Instytut Biologicznych Podstaw Chorób Zwierz¹t
Wydzia³ Medycyny Weterynaryjnej AR; ul. Akademicka 12, 20-033 Lublin; tel.: +48 081 445 66 96, e-mail: grazyna.ziolkowska@ar.lublin.pl
Abstract
Streszczenie
Prace oryginalne
/
Original papers
Mikologia Lekarska 2007, 14 (4): 221-226
Copyright © 2007 Cornetis
www.cornetis.com.pl
ISSN 1232-986X
07ML_04_01_Ziolkowska_Profile_S.indd 221
2007-12-14, 10:38:24
Mikologia Lekarska 2007, 14 (4)
Gra¿yna Zió³kowska, Aneta Nowakiewicz
Protein profiles of superficial antigens M. pachydermatis strains
222
Wprowadzenie
Malassezia pachydermatis (M. pachydermatis), jedyny lipi-
doniezale¿ny gatunek dro¿d¿opodobnych grzybów z rodzaju
Malassezia, nale¿y do powszechnie wystêpuj¹cych komensali
skóry i b³on œluzowych zwierz¹t, g³ównie psów (1, 2). Pod wp³y-
wem zmian w mikroœrodowisku bytowania drobnoustroju, os³a-
bienia mechanizmów obronnych gospodarza, ewentualnie in-
nych, trudnych do okreœlenia przy obecnym stanie wiedzy
czynników, przekszta³ca siê on w formê patogenn¹. Towarzyszy
chorobom na tle zaburzeñ immunologicznych (alergia, atopia)
(3) lub hormonalnych (nadczynnoϾ kory nadnerczy, nadczyn-
noœæ tarczycy, cukrzyca), efektem których jest wzrost wydziela-
nia ³oju lub zmiana jego sk³adu (4). Jako patogen M. pachyder-
matis odpowiedzialny jest w 30-80% przypadków za zapalenia
zewnêtrznego kana³u s³uchowego (otitis externa) oraz w 30%
przypadków ³ojotokowego zapalenia skóry (dermatitis) (5),
zw³aszcza okolic pach, pachwin, brzucha i przestrzeni miêdzy-
palcowych (1). U ludzi poza zmianami skórnymi mo¿e powodo-
waæ fungemie (6), szczególnie u noworodków na oddzia³ach in-
tensywnej terapii (7).
Ze wzglêdu na to, ¿e zarówno otitis externa, jak i dermatitis
mog¹ byæ powodowane u psów tak¿e przez wiele innych czynni-
ków, m.in. Staphylococcus intermedius, S. hyicus, Pseudomo-
nas aeruginosa, E. coli czy Streptococcus spp. (8); bardzo
istotne jest dok³adne ustalenie czynnika etiologicznego choro-
by. Izolacja M. pachydermatis, nawet przy wysokiej zawartoœci
grzyba w pobranym materiale nie zawsze œwiadczy o jego wiru-
lencji. Wykazano bardzo wysoki, oko³o 75% odsetek psów, od
których – bez wzglêdu na obecnoœæ lub brak objawów chorobo-
wych – izolowano M. pachydermatis (2).
Obecnie istotnym kryterium przy ustalaniu czynnika etiolo-
gicznego otitis externa czy dermatitis jest izolacja ze zmian
chorobowych M. pachydermatis w czystej kulturze oraz pozy-
tywna reakcja zwierz¹t na terapiê antygrzybicz¹ (4).
Poszukiwanie fenotypowych cech grzyba odpowiedzialnych
za jego wirulencjê nie przynios³o, jak dot¹d, zadowalaj¹cych
wyników. Badanie struktury antygenowej M. pachydermatis
i porównawcza analiza profili bia³kowych poszczególnych szcze-
pów pozwoli³y wykazaæ pewne prawid³owoœci odrêbne dla grzy-
bów komensalicznych i patogennych (9). Jednak ze wzglêdu na
z³o¿on¹ strukturê antygenow¹ tych grzybów dok³adniejsze ba-
dania by³y utrudnione. Celowe wydawa³o siê wiêc zawê¿enie
badañ do antygenów powierzchniowych M. pachydermatis.
Antygeny te bezpoœrednio uczestnicz¹c w pierwszych etapach
infekcji, jakimi s¹: adherencja do komórek gospodarza, koloni-
zacja tkanek oraz proliferacja grzyba, maj¹ istotny wp³yw na te
procesy, a tym samym odpowiadaj¹ za wirulencjê zarazka (10).
Wykazanie obecnoœci takich frakcji albo przynajmniej silniejsz¹
ich ekspresjê, umo¿liwi byæ mo¿e bardziej precyzyjne typowa-
nie szczepów M. pachydermatis odpowiedzialnych za procesy
chorobowe.
Cel pracy
Celem pracy by³o uzyskanie i standaryzacja procedury otrzy-
mywania antygenów powierzchniowych M. pachydermatis,
okreœlenie ich profili bia³kowych oraz analiza porównawcza an-
tygenów powierzchniowych szczepów izolowanych od psów
z objawami zapalenia zewnêtrznego kana³u s³uchowego i zdro-
wych klinicznie.
Materia³ i metody
Badaniami objêto 40 szczepów M. pachydermatis wyizolo-
wanych bezpoœrednio z materia³u klinicznego. Szczepy wytypo-
wane do badañ izolowano w czystej kulturze (bez obecnoœci
towarzysz¹cej bioty bakteryjnej i grzybiczej) od psów z kliniczny-
mi objawami zapalenia zewnêtrznego kana³u s³uchowego (oti-
tis externa) i reaguj¹cymi pozytywnie na kuracjê antygrzybicz¹
(n=20) oraz od psów zdrowych klinicznie (n=20).
Antygeny powierzchniowe. Badane szczepy M. pachyder-
matis namna¿ano na sta³ym pod³o¿u Sabourauda przez 3 dni
w temp. 37°C. Uzyskany materia³ delikatnie zbierano z pod³o¿a,
oceniano mikroskopowo i zawieszano w ja³owej wodzie destylo-
wanej z dodatkiem 0,02% merthiolatu, w proporcji 10 ml roz-
tworu na 1 p³ytkê hodowli. Tak przygotowane zawiesiny umiesz-
czano na mieszadle magnetycznym i inkubowano przez 1,
3 i 24 godz.y w temp. 37°C, celem powolnej i ³agodnej elucji
antygenów powierzchniowych z komórek grzyba. Nastêpnie
preparaty wirowano w temp. 4°C przez 30 min (4×1000 g),
a supernatanty poddawano 30-krotnemu zagêszczeniu przez
ultrafiltracjê w systemie Amicon-Centriprep 10. Otrzymany ma-
teria³ porcjowano po 200 µl i przechowywano w temp. -20°C.
Profile bia³kowe. Profile antygenów powierzchniowych po-
szczególnych szczepów okreœlano na podstawie rozdzia³u elek-
troforetycznego w warunkach zdenaturowanych (SDS-PAGE) wg
Laemmli (11), w ¿elach poliakrylamidowych. Elektroforezê prze-
prowadzano przy sta³ym natê¿eniu 25 mA przez oko³o 90 min.
Próbki badanego materia³u zawieszano w buforze lizuj¹cym
(0,5 M Tris-HCl pH 6,8, 10% SDS, glicerol, 2-b merkaptoetanol,
0,05% b³êkit bromofenolowy) w stosunku 1:1 i ogrzewano
przez 3 min w temp. 90°C, a nastêpnie w iloœci 20 µl wprowa-
dzano do studzienek startowych. Jako wzorca mas cz¹steczko-
wych u¿yto zestaw Page ruled unstained protein ladder (Fer-
mentas). ¯ele barwiono roztworem b³êkitu kumazyny (CBB
R250), a analizê poszczególnych elektroforegramów przepro-
wadzono z zastosowaniem GEl-Doc 2000.
Wyniki
Profile bia³kowe antygenów powierzchniowych Malassezia
pachydermatis okreœlono na podstawie analizy elektroforegra-
mów materia³ów uzyskanych na drodze elucji 3-dniowych ho-
dowli, 40 uprzednio wytypowanych szczepów grzyba. Wyjœcio-
wa pula badanych preparatów obejmowa³a materia³ uzyskany
w wyniku 1-, 3- i 24-godzinnego procesu elucji masy komórko-
wej poszczególnych szczepów. Wstêpna ocena uzyskanych
elektroforegramów (ryc. 1 i 2) pozwoli³a wytypowaæ do dalszej
analizy jedynie wyniki uzyskane z rozdzia³u materia³ów pocho-
dz¹cych z 3-godzinnego procesu elucji.
Rozdzia³y elektroforetyczne bia³kowych frakcji antygenów
powierzchniowych ilustruj¹ ryciny 1 i 2.
Wykazano wysoki stopieñ zró¿nicowania i z³o¿on¹ strukturê
poszczególnych preparatów antygenowych. Frakcje bia³kowe,
których liczba, w zale¿noœci od badanego szczepu, waha³a siê od
2 do 22, zawarte by³y w przedziale od 104 do 13 kDa. Szczepy
izolowane od zwierz¹t chorych charakteryzowa³y siê wyraŸnie
silniejsz¹ ekspresj¹ bia³kowych antygenów powierzchniowych
i wiêkszym ich zró¿nicowaniem (ryc. 2, tab. II). Liczba frakcji bia³-
kowych wynosi³a w tym przypadku œrednio 11, przy czym warto-
œci dla poszczególnych szczepów waha³y siê od 3 do 22.
Szczepy komensaliczne wykazywa³y obecnoœæ, œrednio
5 frakcji bia³kowych, przy wahaniach od 2 do 13 pr¹¿ków, w za-
le¿noœci od badanego antygenu powierzchniowego (ryc. 1, tab.
I). Bez wzglêdu na pochodzenie szczepów (od zwierz¹t zdro-
wych lub chorych) ich antygeny powierzchniowe wykazywa³y
obecnoœæ frakcji wspólnych: 68-71 kDa, 57-58 kDa, 45-46 kDa
(tab. I i II). Stwierdzono ponadto frakcje swoiste, typowe dla
szczepów pochodz¹cych od psów z otitis externa; s¹ to: 61-64
kDa, 52-53 kDa, 43-44 kDa, 38-39 kDa i 15-17 kDa (tab. II i III),
a których obecnoœci nie wykazano w antygenach powierzchnio-
07ML_04_01_Ziolkowska_Profile_S.indd 222
2007-12-14, 10:38:28

223
Gra¿yna Zió³kowska, Aneta Nowakiewicz
Profile bia³kowe antygenów powierzchniowych Malassezia pachydermatis
wych szczepów komensalicznych (tab. I). Frakcje te wystêpowa-
³y z ró¿n¹ czêstotliwoœci¹ w badanych szczepach (tab. III); naj-
czêœciej, bo u 45% szczepów, stwierdzano bia³ka o masie cz¹-
steczkowej 43-44 kDa i 38-39 kDa, a nastêpnie (wed³ug
malej¹cej czêstotliwoœci) 52-53 kDa (35%), 15-17 kDa (30%)
oraz 61-64 kDa (20%).
Profile bia³kowe antygenów powierzchniowych „patogen-
nych” szczepów M. pachydermatis, charakteryzowa³a ponadto
podwy¿szona, w stosunku do szczepów komensalicznych, czê-
stotliwoœæ wystêpowania frakcji: 59-60 kDa, 55-56 kDa, 49-50
kDa, 36-37 kDa, 32-33 kDa, 27-28 kDa, 25-26 kDa, 22-23
kDa, 18-20 kDa i 13-14 kDa (tab. I i II). Wspó³zale¿noœci te ilu-
struje ryc. 3. Przyjmuj¹c 50% wystêpowania danej frakcji za
wartoœæ graniczn¹, obecnoœæ pr¹¿ków 55-56 kDa, 36-37 kDa,
27-28 kDa i 18-19 kDa mo¿e, w po³¹czeniu z obecnoœci¹ frakcji
typowo swoistych, wskazywaæ na patogennoœæ danego szcze-
pu M. pachydermatis.
Omówienie
Prowadzone od wielu lat badania struktury antygenowej grzy-
bów z rodzaju Malassezia wykaza³y jej z³o¿ony charakter, a tak¿e
wysoki stopieñ zmiennoœci (12, 13). Bior¹c pod uwagê, ¿e wiêk-
szoœæ antygenów zwi¹zana jest z powierzchni¹ lub œcian¹ ko-
mórkow¹ grzyba (14), a ich ekspresja zale¿y miêdzy innymi od
wieku i warunków hodowli (13), istotne wydawa³o siê ustalenie
optymalnych parametrów uzyskiwania reprezentatywnej puli
antygenów (15). Wczeœniej przeprowadzone badania w³asne
(16), a tak¿e prace innych autorów (9, 15) wykaza³y, ¿e w przy-
padku rodzaju Malassezia, logarytmiczna faza wzrostu grzyba,
tj. 2-6 dzieñ inkubacji, zapewnia najbogatszy zestaw antygenów,
przy czym optymalny czas wzrostu dla M. pachydermatis ustalo-
no na trzy dni (16). Z³o¿ona struktura antygenowa grzybów, ze
wzglêdu na wysok¹ liczebnoœæ frakcji bia³kowych (g³ówne anty-
geny grzybów) utrudnia precyzyjn¹ ich analizê i interpretacjê,
Ryc. 1. Profile bia³kowe antygenów powierzchniowych szczepów M. pachydermatis izolowanych od zwierz¹t zdrowych
Fig. 1. Protein profiles of superficial antigens M. pachydermatis strains obtained from healthy dogs
a
b
c
Ryc. 2. Profile bia³kowe antygenów powierzchniowych szczepów M. pachydermatis izolowanych od zwierz¹t chorych
Fig. 2. Protein profiles of superficial antigens M. pachydermatis strains obtained from diseased dogs
a
b
c
Ryc. 3. Frakcje bia³kowe o wysokiej czêstotliwoœci wystêpowania w szczepach
izolowanych od psów z objawami otitis externa
Fig. 3. Protein fraction in high frequency for strains isolated from dogs with
otitis externa
13-14 kDa
18-20 kDa
22-23 kDa
25-26 kDa
27-28 kDa
32-33 kDa
36-37 kDa
49-50 kDa
55-56 kDa
59-60 kDa
40%
10%
50%
15%
45%
10%
20%
10%
75%
40%
30%
15%
90%
30%
40%
20%
65%
30%
35%
10%
0%
20%
40%
60%
80%
100%
szczepy izolowane od
zdrowych psów / strains
isolated from healthy dogs
szczepy izolowane od
chorych psów / strains
isolated from diseased dogs
07ML_04_01_Ziolkowska_Profile_S.indd 223
2007-12-14, 10:38:30
Mikologia Lekarska 2007, 14 (4)
Gra¿yna Zió³kowska, Aneta Nowakiewicz
Protein profiles of superficial antigens M. pachydermatis strains
224
dlatego te¿ w niniejszej pracy, badania zawê¿ono wy³¹cznie do
antygenów powierzchniowych poszczególnych szczepów. Anty-
geny te wchodz¹ce w sk³ad zewnêtrznej warstwy œciany komór-
kowej bior¹ miedzy innymi udzia³ w procesach interakcji dro¿-
d¿aków z komórkami gospodarza (13), czyli s¹ odpowiedzialne
za pierwsze, bardzo istotne stadium patogenezy (10) oraz cha-
rakteryzuj¹ siê wysokim stopniem swoistoœci (17).
Wykorzystuj¹c to, ¿e antygeny powierzchniowe stanowi¹
frakcjê bia³kow¹ stosunkowo luŸno zwi¹zan¹ z komórk¹ grzyba
i mog¹ ³atwo przechodziæ do œrodowiska, zastosowano opraco-
wan¹ wczeœniej procedurê wodnej elucji grzyba (13), pozwala-
j¹c¹ na zachowanie naturalnej struktury i w³aœciwoœci antyge-
nowych ekstrahowanych bia³ek (16). Elucja antygenów przepro-
wadzona dla znacznej puli szczepów (n=40) równolegle w 3
przedzia³ach czasowych pozwoli³a na ustalenie optymalnego
czasu ekstrakcji do badania struktury antygenów powierzch-
niowych M. pachydermatis. Czas ten okreœlono jako 3 godz., co
potwierdzaj¹ wyniki wczeœniejszych badañ w³asnych (16). Krót-
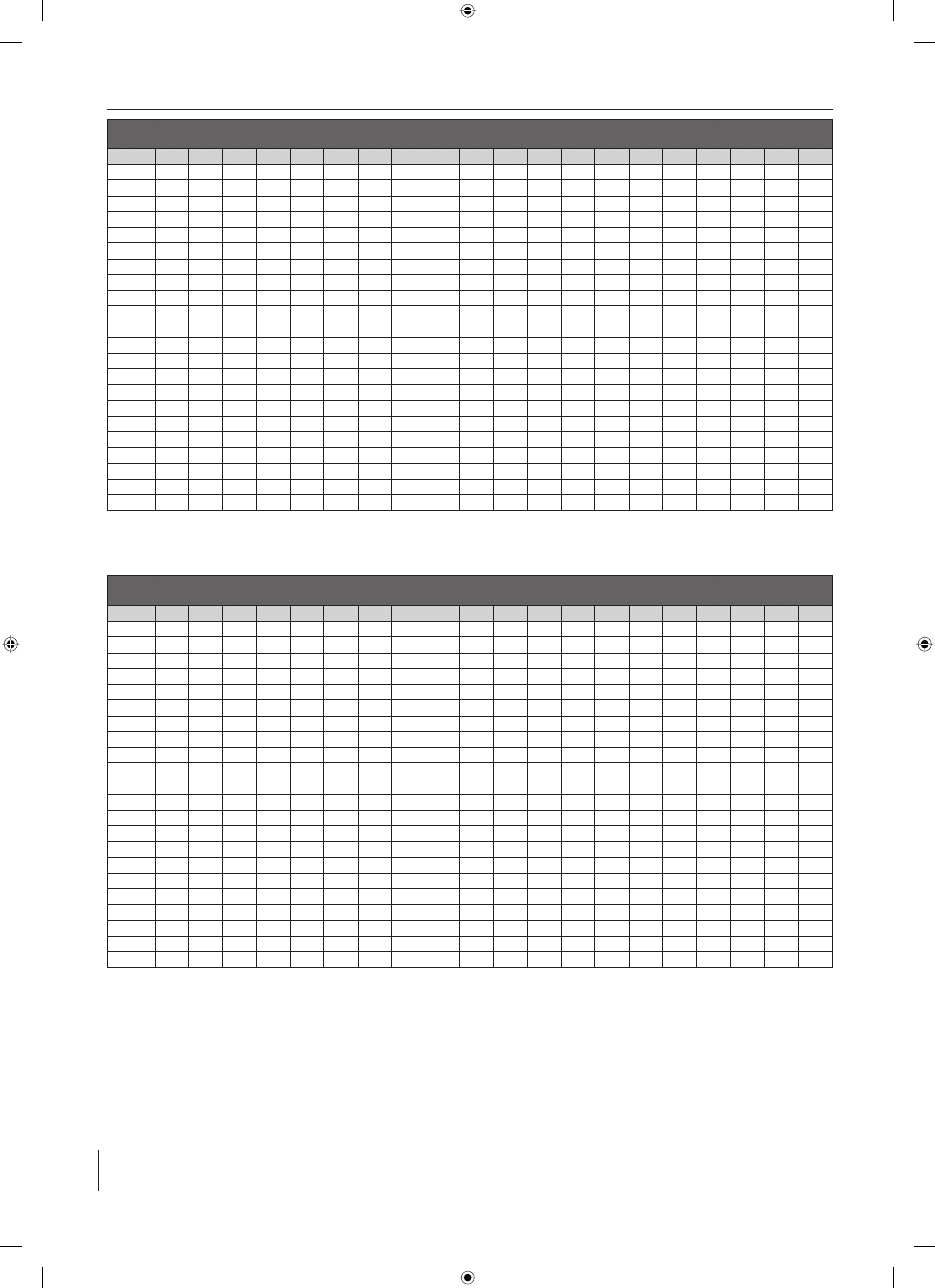
Tabela I: Antygeny powierzchniowe szczepów M. pachydermatis izolowanych od zdrowych psów
Table I: Superficial antigens of M. pachydermatis strains isolated from healthy dogs
kDa
59
66
23a
94
44
90
124
27
65
26a
64
75
141
86
138
73
63
40
16a
76
77-78
+
+
+
+
74-76
+
68-71*
+
+
+
+
+
+
+
+
+
+
+
+
+
+
65-66
+
+
+
+
+
61-64**
59-60
+
+
57-58*
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
55-56
+
+
+
+
+
+
52-53**
49-50
+
+
+
+
45-46*
+
+
+
+
+
+
+
+
+
+
+
+
+
+
43-44**
38-39**
36-37
+
+
+
+
+
+
34-35
+
+
+
+
+
+
+
+
+
32-33
+
+
+
27-28
+
+
+
+
+
+
+
+
25-26
+
+
22-23
+
+
18-20
+
+
+
15-17**
13-14
+
+
Objaœnienie / Explanations
* – frakcje charakterystyczne dla gatunku M. pachydermatis / characteristic fractions for M. pachydermatis species
** – frakcje typowe dla szczepów izolowanych od zwierz¹t chorych / typical fractions for strains isolated from diseased animals
Tabela II: Antygeny powierzchniowe szczepów M. pachydermatis izolowanych od chorych psów
Table II: Superficial antigens of M. pachydermatis strains isolated from diseased dogs
kDa
103
89
133
77
38
147
55
104
47a
14a
37
81
74
46
78
32
36
43
28
26
77-78
+
+
+
+
+
74-76
+
+
68-71*
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
65-66
+
+
+
+
+
61-64**
+
+
+
+
59-60
+
+
+
+
+
+
+
57-58*
+
+
+
+
+
+
+
+
+
+
+
+
55-56
+
+
+
+
+
+
+
+
+
+
+
+
+
52-53**
+
+
+
+
+
+
+
49-50
+
+
+
+
+
+
+
+
45-46*
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
43-44**
+
+
+
+
+
+
+
+
+
38-39**
+
+
+
+
+
+
+
+
+
36-37
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
34-35
+
+
+
+
+
+
+
+
+
+
+
+
32-33
+
+
+
+
+
+
27-28
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
25-26
+
+
+
+
22-23
+
+
+
+
+
+
+
+
+
18-20
+
+
+
+
+
+
+
+
+
+
15-17**
+
+
+
+
+
+
13-14
+
+
+
+
+
+
+
+
Objaœnienie / Explanations
* – frakcje charakterystyczne dla gatunku M. pachydermatis / characteristic fractions for M. pachydermatis species
** – frakcje typowe dla szczepów izolowanych od zwierz¹t chorych / typical fractions for strains isolated from diseased animals
07ML_04_01_Ziolkowska_Profile_S.indd 224
2007-12-14, 10:38:32

225
Gra¿yna Zió³kowska, Aneta Nowakiewicz
Profile bia³kowe antygenów powierzchniowych Malassezia pachydermatis
szy czas elucji (1 godz.) masy grzyba nie zapewnia³ wystarczaj¹-
cej zawartoœci bia³ka w badanych preparatach (bardzo s³aba
wizualizacja rozdzia³u elektroforetycznego). Przed³u¿enie pro-
cesu ekstrakcji (24 godz.) pozwala³o, co prawda, na uzyskanie
materia³u o wysokiej zawartoœci bia³ka, ale jednoczeœnie zró¿-
nicowanego pod wzglêdem pochodzenia. Obok antygenów po-
wierzchniowych w preparatach mog³y znajdowaæ siê bowiem
g³êbiej po³o¿one komponenty komórki grzyba, a tak¿e frakcje
uwolnione podczas ewentualnego procesu autolizy (16).
Wykazano, ¿e antygeny powierzchniowe M. pachydermatis
charakteryzowa³y siê na ogó³ bogat¹ i zró¿nicowan¹ struktur¹.
Analiza poszczególnych elektroforegramów pozwoli³a ustaliæ, ¿e
szczepy izolowane od psów z otitis externa cechowa³a silniejsza
ekspresja antygenów powierzchniowych o wyraŸniej zró¿nicowa-
nej strukturze w porównaniu ze szczepami komensalicznymi.
Œrednia liczba frakcji bia³kowych wynosi³a odpowiednio 11 i 5.
Podobne wyniki uzyskano badaj¹c profile bia³kowe homogena-
tów komórek M. pachydermatis (18), co mo¿e sugerowaæ, ¿e za
wy¿sz¹ ekspresjê bia³ek odpowiedzialne by³y prawdopodobnie
patogenne w³aœciwoœci zarazka (19). Do epitopów bia³kowych
antygenów powierzchniowych, zwi¹zanych wy³¹cznie ze szcze-
pami patogennymi, zaliczono w obecnych badaniach frakcje:
61-64 kDa, 52-53 kDa, 43-44 kDa i 38-39 kDa, których nie wy-
kazywano w profilach szczepów komensalicznych. Mo¿na rów-
nie¿, do pewnego stopnia, uwzglêdniæ frakcje 55-56 kDa, 36-37
kDa, 27-28 kDa i 18-20 kDa, wykazywane u ponad 50% bada-
nych szczepów patogennych, przy nieznacznym procentowym
ich udziale w szczepach komensalicznych. Badania z tego zakre-
su, prowadzone przez innych autorów s¹ stosunkowo nieliczne
i w wiêkszoœci dotycz¹ szczepów Malassezia izolowanych od lu-
dzi (20, 21). Z okreœlonych, jak dot¹d, 12 alergenów, tj. Mala s1,
Mala s5 – Mala s12 i Mala f2 – Mala f4, o wysoce zró¿nicowa-
nych masach cz¹steczkowych, mieszcz¹cych siê w przedziale
16,2 kDa-86,1 kDa (22, 23), a¿ 7 mo¿na porównaæ, z du¿ym
prawdopodobieñstwem, do badanych frakcji antygenów po-
wierzchniowych M. pachydermatis. Zbli¿one masowo pr¹¿ki
w analizowanych obecnie elektroforegramach odnieœæ mo¿na
do alergenów Mala s12 (
»
67 kDa), Mala s11 (
»
22,4 kDa), Mala
s9 (
»
36,7 kDa), Mala s8 (
»
19,2 kDa), Mala s5 (
»
18,2 kDa),
Mala f3 (
»
20 kDa) i Mala s1 (
»
37 kDa). Z puli tej wg Andersson
i wsp. (1) Mala s1, Mala s5 i Mala s8 s¹ charakterystyczne dla M.
pachydermatis, co potwierdzone zosta³o przez wykazanie w ge-
nomie grzyba regionów koduj¹cych te alergeny (20).
Stosuj¹c metody immunodetekcji z wykorzystaniem surowic
pochodz¹cych od psów z infekcj¹ M. pachydermatis, Chen
i wsp. (9) wykazali obecnoœæ frakcji bia³kowych 45, 52, 56 i 65
kDa charakterystycznych jedynie dla szczepów izolowanych od
zwierz¹t chorych. Z kolei Bond i wsp. (24) w podobnych bada-
niach okreœlili inny przedzia³ masowy dla tych antygenów. Frak-
cje reaguj¹ce z przeciwcia³ami IgG psów z objawami zapalenia
skóry na tle Malassezia nale¿a³y do bia³ek o masach 219, 110,
71 i 45 kDa. Antygeny bia³kowe o masach cz¹steczkowych
188, 66, 58, 57, 38, 28 i 17 kDa by³y natomiast typowe dla
patogennych szczepów M. pachydermatis izolowanych przez
Habibah i wsp. (25) od psów z klinicznymi objawami dermatitis.
Podobne zale¿noœci odnotowywano w badaniach antygenów
bia³kowych zwi¹zanych z patogennoœci¹ grzybów z rodzaju Ma-
lassezia wystêpuj¹cych u ludzi (10, 22).
Do czynników prawdopodobnie odpowiedzialnych za stosun-
kowo niewielk¹ powtarzalnoœæ wyników uzyskiwanych przez
ró¿ne oœrodki badawcze zaliczyæ nale¿y wysok¹ heterogennoœæ
wœród szczepów Malassezia spp., ró¿ne procedury ekstrakcji
frakcji antygenowych grzyba, zró¿nicowan¹ odpowiedŸ immu-
nologiczn¹ u poszczególnych pacjentów (przy metodach immu-
nodetekcji) (25), a tak¿e niejednakow¹ dok³adnoœæ oznaczeñ
mas cz¹steczkowych (26). Du¿e znaczenie mo¿e mieæ równie¿
to, ¿e stosowane w przewa¿aj¹cej czêœci badañ metody immu-
noblottingu do oznaczeñ epitopów bia³kowych, zwi¹zanych
z patogennoœci¹ grzyba, nie zawsze mog¹ wykazaæ ich pe³ny
profil. Szczepy Malassezia, bêd¹c bowiem komensalami skóry
i b³on œluzowych, stymuluj¹ uk³ad immunologiczny gospodarza
do produkcji przeciwcia³ (10). Swoiste immunoglobuliny nie
mog¹ wiêc œwiadczyæ o patogennoœci zarazka. Ponadto dro¿-
d¿aki te, podobnie jak inne grzyby, indukuj¹c w organizmie od-
powiedŸ typu komórkowego, mog¹ cechowaæ siê ekspresj¹
frakcji antygenowych, których detekcja nie zawsze jest mo¿liwa
za pomoc¹ przeciwcia³ (13).
Przedstawione wyniki obecnych badañ, prezentuj¹ce z jed-
nej strony frakcje bia³kowe charakterystyczne dla gatunku M.
pachydermatis (68-71 kDa, 57-58 kDa, 45-46 kDa), z drugiej
zaœ frakcje, zwi¹zane – jak siê wydaje – z patogennoœci¹ grzy-
ba (61-64 kDa, 52-53 kDa, 43-44 kDa, 38-39 kDa i 15-17
kDa), ze wzglêdu na odmienn¹ metodykê ekstrakcji materia³u
oraz detekcjê wy³¹cznie za pomoc¹ SDS-PAGE, mog¹ byæ tylko
czêœciowo skonfrontowane z innymi pracami z tego zakresu.
Pewne analogie zaobserwowane miêdzy oznaczonymi frakcja-
mi a alergenami Mala s i Mala f (1) czy te¿ antygenami wyizolo-
wanymi przez innych autorów (15, 25) wymagaj¹ potwierdzenia
w dalszych badaniach, obejmuj¹cych m.in. elucjê poszczegól-
nych frakcji, ich sekwencjonowanie oraz ewentualne oznacze-
nie regionów genomu koduj¹cych ich ekspresjê. Takie badania
pozwol¹ byæ mo¿e na ró¿nicowanie szczepów Malassezia
w aspekcie ich patogennoœci, co znacznie u³atwi prawid³ow¹
diagnostykê infekcji.
Wnioski
1. Szczepy M. pachydermatis izolowane od chorych zwierz¹t
charakteryzuj¹ siê wiêksz¹ ekspresj¹ bia³ek.
2. Profile bia³kowe antygenów powierzchniowych M. pachy-
dermatis cechuje obecnoϾ:
• frakcji typowych wy³¹cznie dla szczepów izolowanych od
psów chorych,
• frakcji wystêpuj¹cych u znacznego odsetka szczepów „pato-
gennych”
3. Korelacja miêdzy pochodzeniem szczepów M. pachyder-
matis a obecnoœci¹ okreœlonych frakcji antygenowych w ich
profilach bia³kowych sugeruje mo¿liwoœæ wystêpowania bia³ko-
wych markerów patogennoœci u tych grzybów.
Praca zosta³a zaprezentowana na XII Miêdzynarodowym Sympozjum Nauko-
wo-Szkoleniowym Sekcji Mikologicznej Polskiego Towarzystwa Dermatologicz-
nego „MIKOLOGIA 2006”, Bia³owie¿a, 20-24.09.2006.
Piœmiennictwo
1. Bond R., Lamport A.J., Lloyd D.H.: Colonization status of Malassezia pachyderma-
tis on the hair and in the hair follicle of healthy beagle dogs. Res. Vet. Sci., 2000,
68, 291-293.
Tabela III: Frakcje bia³kowe typowe dla szczepów izolowanych od psów
z objawami otitis externa
Table III: Typical protein fractions for strains isolated from dogs with the
symptoms of otitis externa
Masa cz¹steczkowa
Molecular mass
[kDa]
Odsetek szczepów wykazuj¹cych
okreœlone frakcje bia³kowe
Percentage strains characterised
by typical protein fractions
61-64 (4)
20%
52-53
(7) 35%
43-44
(9) 45%
38-39
(9) 45%
07ML_04_01_Ziolkowska_Profile_S.indd 225
2007-12-14, 10:38:37
Mikologia Lekarska 2007, 14 (4)
Gra¿yna Zió³kowska, Aneta Nowakiewicz
Protein profiles of superficial antigens M. pachydermatis strains
226
2. Guillot J., Bond R.: Malassezia pachydermatis: a review. Med. Mycol., 1999, 37,
295-306.
3. Mouldin E.A., Morris D.O., Goldsmidt E.: Retrospective study; the presence of
Malassezia in feline skin biopsies. A clinicopathological study. Vet. Dermatol.,
2002, 13, 7-13.
4. Patterson A.D., Frank L.A.: How to diagnose and treat Malassezia dermatitis in
dogs. Vet. Med., 2002, 8, 612-622.
5. Staroniewicz Z., Król J., Cierpisz J.: Flora bakteryjna i grzybicza w otitis externa
u psów. Medycyna Wet., 1995, 51, 667-670.
6. Gueho E., Boeghout T., Ashbee H.R., Guillot J., Van Belkum A., Faergemann J.: The
role of Malassezia species in the ecology of human skin and as pathogens. Med.
Mycol., 1998, 36, 220- 229.
7. Belkum A., Boekhout T., Bosboon R.: Monitoring spreed of Malassezia infections
in a neonatal intensive care unit by PCR-mediated genetic typic. J. Clin. Microbio-
logy, 1994, 32, 2528-2538.
8. Dowling M.P.: Antimicrobial therapy of skin and ear infection. Can. Vet., 1996, 37,
695-699.
9. Chen T., Halliwell R.E.W., Pemberton A.D., Hill P.B.: Identification of major aller-
gens of Malassezia pachydermatis in dogs with atopic dermatitis and Malassezia
overgrowth. Vet. Dermatol., 2002, 13, 141-150.
10. Ashbee H.R., Evans E.G.V.: Immunology of diseases associated with Malassezia
species. Clin. Microbiology Rev., 2002, 15, 21-57.
11. Laemmli U.K.: Cleavage of structural proteins during the assembly of the head of
bacteriophage T4. Nature, 1970, 227, 680-685.
12. Jensen-Jarolim E., Poulsen L.K., With H., Kieffer M., Ottevanger V., Skov P.S.: Ato-
pic dermatitis of the face, scalp and neck: type I reaction to the yeast Pityrospo-
rum ovale?. J. Allergy Clin. Immunol., 1992, 89, 44-51.
13. Zió³kowska G., Nowakiewicz A., Puchalski A.: Profile bia³kowe preparatów antyge-
nowych uzyskiwanych ze szczepów Malassezia pachydermatis. Medycyna Wet.,
2004, 60, 1118-1123.
14. Fukazawa Y., Kagaya K.: Molecular bases of adhesion of Candida albicans. J. Med.
Vet. Mycol., 1997, 35, 87-99.
15. Coutinho S., Souza T., Paula C.D.: Protein profiles of Malassezia pachydermatis
isolated from dogs. Mycopathologia, 1997, 139, 129-135.
16. Zió³kowska G.: Charakterystyka i znaczenie praktyczne egzoantygenów Micro-
sporum canis. Rozprawa habilitacyjna. Rozprawy Naukowe AR w Lublinie nr 236.
2000.
17. Cunnigham A., C., Leeming J.P., Ingham E., Gowland G.: Differentiation of three
serovars of Malassezia furfur. J. Appl. Bacteriol., 1990, 68, 439-446.
18. Nowakiewicz A.: Wspó³czesne metody ró¿nicowania diagnostycznego szczepów
z rodzaju Malassezia izolowanych z przypadków klinicznych. Rozprawa doktor-
ska. Rozprawy Naukowe AR w Lublinie, 2005.
19. Romani L., Howard D.H.: Mechanisms of resistance to fungal infections. Curr.
Opin. Immunol., 1990, 126, 627-632.
20. Andersson A., Scheynius A., Rasool O.: Detection of Mala f and Mala s allergen
sequences within the genus Malassezia. Med. Mycol., 2003, 41, 479-485.
21. Onishi Y., Kuroda M., Yasueda H., Saito A., Sono-Koyama E., Tunasawa S., Hashi-
da-Okado T., Yakigara T., Uchida K.: Two-dimensional electrophoresis of Malasse-
zia and isolation of Mal f4 homologs with mitochondrial malate dehydrogenase.
Eur. J. Biochem., 1999, 261, 148-154.
22. Zargari A., Midgley G., Back O., Johansson S.G.O., Scheynius A.: Ig-E reactivity to
seven Malassezia species. Allergy, 2003, 58, 306-311.
23. Zargari A.: Identification and characterization of allergen components of the op-
portunistic yeast Malassezia furfur. Repro. Print AB Stockholm, 1998.
24. Bond R., Lloyd D.H.: Immunoglobulin G responses to Malassezia pachydermatis in
healthy dogs and dogs with Malassezia dermatitis. Vet. Rec., 2002, 150, 509-512.
25. Habibah A., Catchpole B., Bond R.: Canine serum immunoreactivity to M. pachy-
dermatis in vitro is influenced by the phase of yeast growth. Vet. Dermatol., 2005,
16, 147-152.
26. Ashbee H.R.: Recent developments in the immunology and biology of Malassezia
species. Immunol. Med. Microbiol., 2006, 47, 14-23.
Praca wp³ynê³a do Redakcji: 2007.03.28. Zaakceptowano do druku: 2007.09.02.
07ML_04_01_Ziolkowska_Profile_S.indd 226
2007-12-14, 10:38:39
Wyszukiwarka
Podobne podstrony:
więcej podobnych podstron