
Biotechnologia szans¹ dla
zastosowania allelopatii jako
alternatywnej metody zwalczania
chwastów
Agnieszka Gniazdowska
Katedra Fizjologii Roœlin, Wydzia³ Rolnictwa i Biologii,
Szko³a G³ówna Gospodarstwa Wiejskiego, Warszawa
Biotechnology – a chance for using allelopathy as alternative weed
management strategy
S u m m a r y
Plants (crops and weeds) affect each other through allelochemicals, which
may be released from living and dead organisms. Due to an increase in the num-
ber of herbicide –resistant weeds and herbicides’ negative effect on the envi-
ronment, there is an effort being made to design alternative weed management
strategies. Allelopathic studies offer a challenge for a discovery of new com-
pounds with new target side that may be able to control weeds. Crops produc-
ing allelochemicals can interfere with competing weeds sufficiently enough to
allow significant reductions in the use of other weed management options, in-
cluding synthetic herbicides. Some attempts enhancing the allelopathic poten-
tial of crops (rice, sorghum, barley and wheat) to control weeds are presented.
Key words:
allelopathy, allelochemicals, herbicide, weed management.
1. Wstêp – Czym jest allelopatia?
Wzajemne oddzia³ywania pomiêdzy roœlinami s¹ znane i ob-
serwowane od wieków, jednak dopiero Hans Molish w 1937 r.
zdefiniowa³ to zjawisko, wprowadzaj¹c pojêcie allelopatii. Okreœ-
la ono interakcje wystêpuj¹ce w uk³adach: roœliny-roœliny, roœli-
ny-mikroorganizmy oraz mikroorganizmy-mikroorganizmy. S¹ to
P R A C E P R Z E G L ¥ D O W E
Adres do korespondencji
Agnieszka Gniazdowska,
Katedra Fizjologii Roœlin,
Wydzia³ Rolnictwa
i Biologii,
Szko³a G³ówna
Gospodarstwa Wiejskiego,
ul. Nowoursynowska 159,
02-776 Warszawa;
e-mail:
agnieszka_gniazdowska@
sggw.pl
2 (77) 42–53 2007

oddzia³ywania biochemiczne o charakterze zarówno szkodliwym jak i korzystnym
(1). Na pierwszym œwiatowym kongresie allelopatii w 1996 r. rozszerzono tê defini-
cjê i uznano, ¿e allelopatia to zjawisko, w którym zaanga¿owane s¹ wtórne metabo-
lity, wytwarzane przez roœliny, mikroorganizmy i grzyby, wp³ywaj¹ce na wzrost
i rozwój systemów biologicznych i rolniczych. Zwi¹zki allelopatyczne (allelopatiny)
s¹ to produkty metabolizmu wtórnego roœliny-donora, które po dostaniu siê do œro-
dowiska oddzia³uj¹ na znajduj¹ce siê w s¹siedztwie roœliny-akceptory. Dzia³anie al-
lelopatin ujawnia siê na wszystkich poziomach organizacji ¿ywego organizmu: od fi-
zjologicznego przez komórkowy, po molekularny, a pod wzglêdem chemicznym
obejmuj¹ one ró¿norodne zwi¹zki organiczne. S¹ wœród nich wystêpuj¹ce w formie
gazowej terpeny:
a-pinen, b-pinen, kamfen, cyneol obecne np. w tkankach bylic
(roœliny z rodzaju Artemisia) (2), jak te¿ skomplikowane, wielopierœcieniowe seskwi-
terpenoidy-heliannany np. heliannuole, laktony seskwiterpenów np. annuolid F ze
s³onecznika (Helianthus annuus) (3). G³ówne klasy zwi¹zków allelopatycznych zosta³y
szczegó³owo omówione w pracach przegl¹dowych Vyvyan
(4
) oraz Singh i wsp. (5).
Wydzielanie zwi¹zków allelopatycznych do œrodowiska mo¿e odbywaæ siê na ró¿nej
drodze: uwalniania lotnych substancji (ewaporacja); wymywania (³ugowanie) przez
wodê z opadów atmosferycznych, wodê irygacyjn¹ lub rosê; wydzielania przez sys-
tem korzeniowy (eksudacja); rozk³adu obumar³ych czêœci roœlin. Poszczególne drogi
dostawania siê zwi¹zków allelopatycznych do œrodowiska zosta³y omówione w pra-
cach w jêzyku polskim, m.in. przez Wójcik-Wojtkowiak i wsp. (6) oraz Gniazdowsk¹
i wsp. (7). Nagromadzone w glebie zwi¹zki allelopatyczne pochodz¹ zarówno od
roœlin uprawnych, roœlin dziko rosn¹cych, chwastów, jak te¿ mog¹ byæ metabolitami
mikroorganizmów glebowych.
2. Allelopatia chwastów
Od drugiej po³owy XX w. wzros³o zainteresowanie oddzia³ywaniami allelopa-
tycznymi miêdzy chwastami i roœlinami uprawnymi. Dostarczano coraz wiêcej do-
wodów, ¿e roœliny uprawne i chwasty wprowadzaj¹ do œrodowiska zwi¹zki chemicz-
ne, które mog¹ byæ toksyczne zarówno dla nich samych, jak równie¿ dla innych ga-
tunków (6). Pojawi³o siê szereg doniesieñ wskazuj¹cych, ¿e niekorzystny wp³yw za-
chwaszczenia na uprawy jest zwi¹zany nie tylko z konkurencj¹ o œwiat³o, wodê
i zwi¹zki mineralne, ale tak¿e mo¿e byæ wynikiem produkowania przez chwasty al-
lelopatin dzia³aj¹cych jako inhibitory wzrostu i rozwoju roœlin uprawnych. Roœliny
nale¿¹ce do rodzaju Artemisia to chwasty, wystêpuj¹ce powszechnie w Azji, Europie
i Ameryce Pó³nocnej. Bylica pospolita (A. vulgaris) stanowi jeden z najwiêkszych pro-
blemów w szkó³kach we wschodnich Stanach Zjednoczonych Ameryki Pó³nocnej (8).
Posiada zdolnoϾ do wytwarzania silnego systemu korzeniowego i jest odporna na
herbicydy, a tak¿e toleruje powtarzaj¹ce siê koszenie. Z kolei A. campestris, A. annua,
A. herba alba i A. princeps var. orientalis tworz¹ gêste monokultury (9,10). Przyczyn¹
Biotechnologia szans¹ dla zastosowania allelopatii jako alternatywnej metody zwalczania chwastów
BIOTECHNOLOGIA 2 (77) 42-53 2007
43

tego zjawiska mo¿e byæ zdolnoœæ tych roœlin do syntezy du¿ych iloœci olejków ete-
rycznych w specjalnych gruczo³ach zlokalizowanych na powierzchni liœci. Wykazano
w nich obecnoœæ kamfory, kamfenu, cyneolu, a tak¿e artemizininy. Zarówno liœcie
jak i wodne ekstrakty Artemisia silnie hamuj¹ wzrost roœlin uprawnych. Tak¿e gleba
pobrana z miejsc zajmowanych przez bylice hamuje wzrost i rozwój wielu gatun-
ków roœlin oraz grzybów mikorytycznych. Nie poznano, jak dot¹d, molekularnego
pod³o¿a oddzia³ywañ tych substancji, jednak¿e wydaje siê, ¿e charakteryzuj¹ siê
mechanizmami dzia³ania, ró¿nymi od mechanizmów dzia³ania typowych dla komer-
cyjnych herbicydów (11).
Wulpia mysi ogon (Vulpia myuros) pochodz¹ca z krajów œródziemnomorskich
sta³a siê inwazyjnym chwastem Australii. Powoduje dotkliwe straty w plonach
i jakoœci zbó¿, a walka z ni¹ jest utrudniona z uwagi na niewra¿liwoœæ na wiêkszoœæ
syntetycznych herbicydów, ponadto pozosta³oœci tej roœliny wykazuj¹ znaczn¹ tok-
sycznoϾ (12). W wodnych ekstraktach z resztek V. myuros, zidentyfikowano 20 ak-
tywnych zwi¹zków, m.in. katechol, pirogallol, alkohol koniferylowy, kwasy benzo-
esowy, bursztynowy, hydrocynamonowy, salicylowy, protokatechusowy, wanilino-
wy, syringowy, p-hydroksybenzoesowy, dihydroferulowy, p-kumarowy, dihydroka-
wowy, ferulowy. Allelopatiny obecne w du¿ej iloœci, np kwasy: syringowy, wanilino-
wy, bursztynowy wykazuj¹ nisk¹ aktywnoœæ allelopatyczn¹, podczas gdy te obecne
w ma³ych iloœciach: kwas hydrocynamonowy, katechol wykazuj¹ wysok¹ aktywnoœæ
inhibitorow¹. Na podstawie wyników z przeprowadzonych eksperymentów wskazu-
je siê, ¿e w oddzia³ywaniu allelopatycznym u wulpi o wiele wiêksze znaczenie ma
mieszanina zwi¹zków allelopatycznych i powstaj¹ce efekty addytywne lub/i synergi-
styczne ni¿ pojedyncze, bardzo aktywne allelopatiny (13).
Okazuje siê, ¿e agresywnoœæ perzu w³aœciwego (Agropyron regens) wynika nie tyl-
ko z konkurencji, ale ma równie¿ pod³o¿e chemiczne. W wydzielinach korzenio-
wych tej roœliny zidentyfikowano szereg allelopatin, m.in. kwasy: wanilinowy, 4-hy-
drocynamonowy, ferulowy, protokatechusowy, syringowy, a tak¿e kwasy hydroksa-
mowe np. DIBOA (2,4-dihydroksy-1,4(2H)benzoksazyn-3-on), dla których obserwo-
wano najwy¿sz¹ aktywnoœæ allelopatyczn¹ (14).
Euroazjatyckie gatunki z rodzaju Centaurea zachwaszczaj¹ zarówno pastwiska jak
i pola uprawne Ameryki Pó³nocnej i Po³udniowej. Chaber plamisty (C. maculosa) inwa-
zyjny chwast w zachodniej Ameryce Pó³nocnej wytwarza mieszaninê stereoizomerów
(+)-katechinê i (-)-epikatechinê, gromadz¹ce siê w glebie wokó³ korzeni. Katechina ha-
muje rozwój i wzrost rodzimych gatunków traw Ameryki Pó³nocnej, a roœliny na ni¹
wra¿liwe reaguj¹ œmierci¹ komórek, postêpuj¹c¹ od wierzcho³ka korzenia (15).
Badania nad rol¹ allelopatii w inwazyjnym sukcesie niektórych chwastów mog¹
pozwoliæ na skonstruowanie w przysz³oœci genetycznie zmodyfikowanych roœlin
uprawnych, które bêd¹ odporne na allelopatiny wydzielane przez chwasty. Teore-
tycznie, mo¿liwe jest wprowadzenie do genomu roœlin uprawnych takich genów,
których produkty bêd¹ bra³y udzia³ w procesie enzymatycznej detoksykacji allelopa-
tin, na które gatunki uprawne s¹ obecnie wra¿liwe.
Agnieszka Gniazdowska
44
PRACE PRZEGL¥DOWE

3. Allelopatiny w walce z chwastami
Walka z zachwaszczeniem upraw w 2 po³owie XX w. w du¿ej mierze opiera³a siê
na stosowaniu syntetycznych herbicydów. Wzmo¿one u¿ycie syntetycznych herbicy-
dów w rolnictwie spowodowa³o jednak wzrost liczby roœlin odpornych na stosowa-
ne œrodki ochrony i zagro¿enie œrodowiska wynikaj¹ce z nagromadzania siê chemi-
kaliów w glebie i wodach. Heap (16) podaje, ¿e 177 gatunków chwastów (106 dwu-
liœciennych i 71 jednoliœciennych) podleg³o ewolucji prowadz¹cej do uzyskania od-
pornoœci na herbicydy. W Australii ¿ycica (Lolium ridigum), pospolity i niezwykle
agresywny chwast wykazuje odpornoœæ na herbicydy 9 g³ównych grup (16). Podobna
odpornoœæ na herbicydy charakteryzuje tak¿e: owies g³uchy (Avena fatua), szar³at
szorstki (Amaranthus retroflexus), komosê bia³¹ (Chenopodium album), w³oœnicê zie-
lon¹ (Setaria viridis), chwastnicê jednostronn¹ (Echinochloa crus-galli), manneczkê pia-
skow¹ (Eleusine India), mietelnik ¿akulê (Kochia skoparia) oraz przymiot³o kanadyjskie
(Conyza canadensis) (17). Nabycie przez chwasty odpornoœci na herbicydy, definiowa-
nej jako naturalna zdolnoœæ do prze¿ycia i reprodukcji po zastosowaniu letalnej dla
innych roœlin tego samego gatunku dawki herbicydu, stwarza koniecznoœæ szukania
innych – ekologicznych sposobów ograniczania i zwalczania zachwaszczenia.
W krajach cz³onkowskich Unii Europejskiej dokonuje siê przegl¹du œrodków ochro-
ny roœlin, którego celem jest upewnienie siê, ¿e bezpieczeñstwo œrodków ochrony
roœlin dla ludzi, zwierz¹t i œrodowiska oceniane jest na podstawie nowoczesnych
standardów wynikaj¹cych z aktualnego stanu wiedzy. Szacuje siê, ¿e efektem tego
dzia³ania bêdzie zmniejszenie o oko³o 60% liczby obecnie stosowanych substancji
aktywnych. Wœród œrodków, które wycofuje siê ze sprzeda¿y znajduj¹ siê herbicydy
zawieraj¹ce nastêpuj¹ce substancje aktywne: atrazynê, benazolinê, bromacyl, cya-
nazynê, cykloat, dichloroprop, dimefuron, fluoroglikofen, imazapyr, imazetapyr,
metobromuron, napalam, prometrynesetoksydym, symazynê, terbacyl, terbutrynê
(17).
Mechanizm dzia³ania syntetycznych herbicydów dotyczy hamowania podstawo-
wych procesów ¿yciowych roœliny. Herbicydy hamuj¹ proces fotosyntezy (wœród
nich mo¿na rozró¿niæ: inhibitory transportu elektronów, fosforylacji fotosyntetycz-
nej, akceptacji elektronów), proces oddychania, ró¿norodne procesy biosyntezy (do-
tyczy to biosyntezy kwasów t³uszczowych i lipidów, aminokwasów i bia³ek), po-
dzia³y komórkowe i wzrost merystemów. Istnieje te¿ grupa herbicydów o dzia³aniu
zbli¿onym do naturalnej auksyny. Kompleksowy opis mechanizmów dzia³ania wiêk-
szoœci stosowanych herbicydów syntetycznych znajdzie zainteresowany czytelnik
w obszernym opracowaniu w jêzyku polskim (17). Istnieje jednak pilna potrzeba
stworzenia nowych herbicydów spe³niaj¹cych warunki stawiane idealnym œrodkom
ochrony roœlin, takie jak: efektywnoœæ (w dawkach oko³o 500 mg/ha) i szerokie spek-
trum dzia³ania, odpornoœæ na deszcz, elastycznoœæ terminu aplikacji i wysoki po-
ziom bezpieczeñstwa (brak dzia³ania rakotwórczego i toksycznego dla ludzi i zwie-
rz¹t), niskie koszty produkcji (18). Ogromna ró¿norodnoœæ allelopatin wystêpuj¹-
Biotechnologia szans¹ dla zastosowania allelopatii jako alternatywnej metody zwalczania chwastów
BIOTECHNOLOGIA 2 (77) 42-53 2007
45

cych w tkankach roœlinnych, jak siê wydaje, mo¿e byæ podstaw¹ do konstruowania
na ich bazie herbicydów. Spoœród znanych allelopatin, które mog³yby byæ u¿yte jako
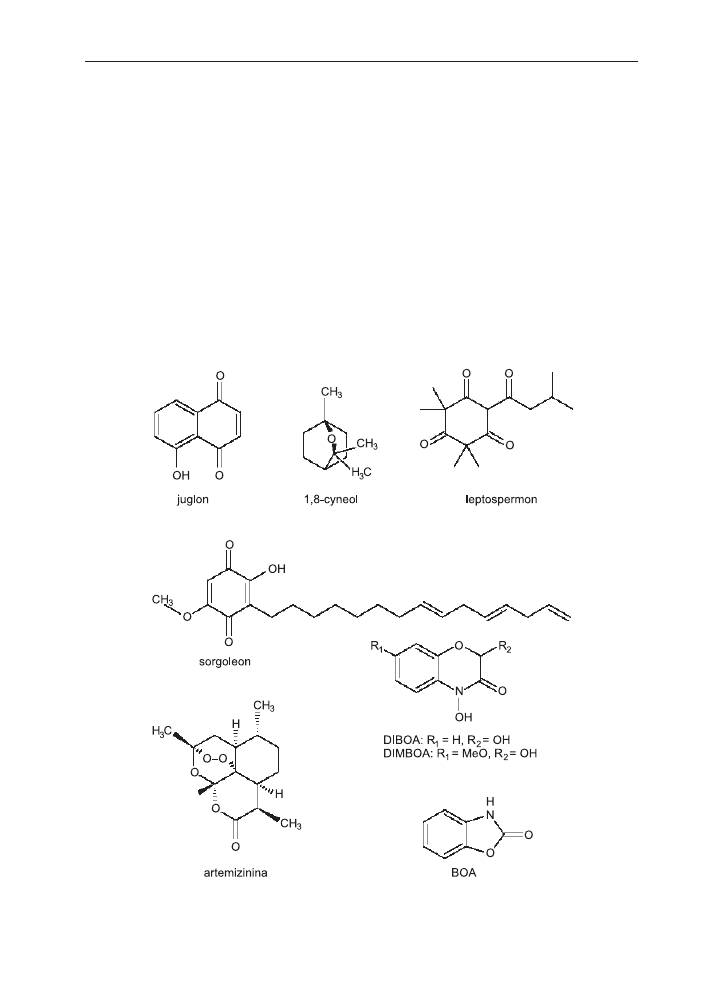
naturalne herbicydy nale¿y wymieniæ: chinony – juglon i sorgoleon; monoterpeny
– 1,8 cyneol i jego analog cinmetylinê, uwa¿an¹ za pierwszy dostêpny na rynku alle-
loherbicyd; laktony seskwiterpenów: artemizininê i dehydrozaluzaninê C (4) (rys.).
Tym bardziej, ¿e biochemiczny mechanizm dzia³ania tych allelopatin jest w du¿ej
mierze poznany (11,19-21). St¹d, wykorzystanie ich do walki biologicznej z chwa-
stami wzbudza coraz wiêksze zainteresowanie (8,22-24). Od lat w rolnictwie zacho-
wawczym stosowane s¹ „konwencjonalne” metody zwalczania chwastów oparte na
allelopatii. Polegaj¹ one na: 1) zastosowaniu roœlin allelopatycznych jako Ÿród³a na-
turalnych œrodków ochrony roœlin: w postaci wyci¹gów, ekstraktów, wyp³uczyn, np.
ze s³omy, posprzêtnych pozosta³oœci roœliny allelopatycznej w systemie bezorko-
wym, rotacji upraw (zmianowanie), upraw z wsiewk¹; 2) tradycyjnej metodzie selek-
cji i hodowli roœlin uprawnych, które bêd¹ wygrywa³y konkurencjê z chwastami
dziêki wysokiemu potencja³owi allelopatycznemu. Genotypy ogórka, ry¿u, sorgo
i jêczmienia s¹ selekcjonowane w celu wzmocnienia zdolnoœci do syntezy metaboli-
tów wtórnych, odpowiadaj¹cych za hamowanie wzrostu i rozwoju chwastów. Jed-
nak, jak na razie, próby te nie da³y komercyjnie zadowalaj¹cych efektów (25,26).
Wraz z rozwojem biotechnologii (biologii molekularnej i chemii organicznej)
otwiera siê jednoczeœnie nowa szansa dla podniesienia efektywnoœci i skutecznoœci
allelopatii stosowanej jako alternatywna metoda ochrony roœlin. Koncepcje wyko-
rzystania obserwowanych w naturze oddzia³ywañ allelopatycznych dotycz¹: 1) izola-
cji i produkcji allelopatin o charakterze herbicydów z roœlin, w których metodami
in¿ynierii molekularnej doprowadzono do zwiêkszonej syntezy tych allelopatin;
2) konstruowania transgenicznych roœlin uprawnych zdolnych do produkcji allelopa-
tin dzia³aj¹cych jako herbicydy; 3) syntezy herbicydów o strukturze chemicznej na-
turalnych allelopatin.
4. Próby okreœlenia molekularnych podstaw potencja³u allelopatycznego
niektórych roœlin uprawnych
Do najintensywniej badanych roœlin uprawnych charakteryzuj¹cych siê wysokim
potencja³em allelopatycznym nale¿¹ ry¿ (Oryza sativa) i sorgo (Sorghum bicolor). Prze-
badano wiele odmian i linii ry¿u pod k¹tem oddzia³ywañ tej roœliny uprawnej na
wzrost pospolitych chwastów (27). Podczas wieloletnich badañ nad allelopati¹ ry¿u
doprowadzono do identyfikacji du¿ej liczby zwi¹zków chemicznych produkowa-
nych przez tê roœlinê i obecnych w jej wydzielinach korzeniowych: momilacton
B (28), rezorcynole, flawony, benzoksazolinony, cykloheksanony i pochodne gliko-
zydowe (29). Ze wzglêdu na mnogoœæ allelopatin wydzielanych przez ró¿ne odmia-
ny ry¿u, jak siê wydaje, trudno mówiæ o konkretnym zwi¹zku odpowiedzialnym za
allelopatiê tej roœliny. Dodatkowo, z uwagi na obecnoœæ tak wielu zwi¹zków allelo-
Agnieszka Gniazdowska
46
PRACE PRZEGL¥DOWE

patyczych w tkankach ry¿u ich dzia³anie mo¿e byæ synergistyczne lub addytywne.
Jednak sklonowanie kompletnego genomu ry¿u (30,31) da³o mo¿liwoœæ identyfikacji
genów zwi¹zanych z produkcj¹ najwa¿niejszych allelopatin ry¿u. Xu i wsp. (32) zi-
dentyfikowali gen koduj¹cy syntazê difosforanu syn-kopalylu odgrywaj¹c¹ g³ówn¹
rolê w syntezie momilactonu. Dayan i wsp. (33) zidentyfikowali enzym uczestni-
cz¹cy w tworzeniu pochodnych rezocynolowych obecnych w wydzielinach korzenio-
wych ry¿u. Uda³o siê im te¿ zidentyfikowaæ 8 sekwencji odpowiadaj¹cych potencjal-
nym genom koduj¹cym ten enzym. Jednym ze zwi¹zków magazynowanych w du-
¿ych iloœciach w tkankach ry¿u jest równie¿ kwas p-kumarowy. Hydrolaza kwasu cy-
namonowego (CA4H) enzym katalizuj¹cy przekszta³cenie kwasu cynamonowego do
p-kumarowego odgrywa g³ówn¹ rolê w biosyntezie zwi¹zków fenolowych u roœlin.
Zaobserwowano, ¿e aktywnoœæ CA4H jest indukowana przez œwiat³o UV (34). U¿yto
zatem specyficznego promotora CASC aktywowanego przez œwiat³o UV w celu skon-
struowania transgenicznego ry¿u, u którego mo¿na bêdzie regulowaæ syntezê feny-
lopropanoidów za pomoc¹ œwiat³a UV (35).
Podjêto równie¿ próby zmapowania loci cech iloœciowych odpowiedzialnych za po-
tencja³ allelopatyczny ry¿u (26,36). Bach-Jansen i wsp. (26) badali loci cech iloœciowych
(QTLs) allelopatii ry¿u w populacji 142 lini rekombinantów powsta³ych z krzy¿owania
dwóch odmian ry¿u ró¿ni¹cych siê znacz¹co potencja³em alleopatycznym w stosunku
do E. crus-galli, wybrano odmianê IAC 165 o wysokim potencjale allelopatycznym i od-
mianê CO 39 o niskim potencjale allelopatycznym. Zbadano 140 markerów genetycz-
nych i zidentyfikowano cztery QTLs rozmieszczone na trzech chromosomach. Dwa
spoœród zidentyfikowanych QTLs s¹ zlokalizowane na chromosomie 3 i, jak siê wydaje,
s¹ stosunkowo ³atwe do manipulacji. Z kolei analiza RFLP (polimorfizm d³ugoœci frag-
mentów restrykcyjnych) przeprowadzona przez Ebana i wsp. (37) pozwoli³a na identy-
fikacjê siedmiu QTLs cechy allelopatii u ry¿u, rozmieszczonych na szeœciu chromoso-
mach. W tym przypadku wariacja fenotypowa wynosi³a od 9,4 do 16,1%. Podobn¹ pró-
bê identyfikacji loci cech iloœciowych allelopatii u pszenicy (Triticum aestivum) wykonali
Wu i wsp. (38). Zbadali oddzia³ywania allelopatyczne pszenicy w stosunku do L. ridigum.
U¿yli 271 linii podwójnych haploidów otrzymanych z roœlin pszenicy o wysokim i ni-
skim potencjale allelopatycznym. Zaobserwowano ró¿nice w aktywnoœci allelopatycz-
nej otrzymanych podwójnych haploidach okreœlane jako procent hamowania wzrostu
korzenia chwastu, kszta³tuj¹ce siê w granicach 23,7-88,3%. Na podstawie analizy RFLP,
AFLP (polimorfizm d³ugoœci amplifikowanego fragmentu) i SSRs (mikrosatelitarny poli-
morfizm krótkich tandemowych powtórzeñ) wykonano identyfikacjê dwóch QTLs alle-
lopatii pszenicy rozmieszczonych na chromosomie 2. Potencja³ allelopatyczny pszeni-
cy podobnie jak ry¿u i ¿yta zwi¹zany jest g³ównie z biosyntez¹ kwasów hydroksamo-
wych, takich jak DIBOA (2,4-dihydroksy-1,4(2H)benzoksazyn-3-on), DIMBOA (2,4-dihy-
droksy-7-metoksy-1,4-benzoksazyn-3-on) i benzoksazolinonów BOA (2(3H)-benzoksa-
zolinon), MBOA (6-metoksy-2(3H)-benzoksazolinon) (rys.) (39).
Najwiêksz¹ przysz³oœæ, jak siê wydaje, ma sorgo jako Ÿród³o allelopatin i poten-
cjalny przedmiot manipulacji biotechnologicznych. Zaobserwowano, ¿e sorgoleon
Biotechnologia szans¹ dla zastosowania allelopatii jako alternatywnej metody zwalczania chwastów
BIOTECHNOLOGIA 2 (77) 42-53 2007
47

(rys.) jest wydzielany przez komórki w³oœnikowe korzeni sorgo do ryzosfery. Wy-
dzieliny korzeniowe sorgo zawieraj¹ oko³o 85-90% czystego sorgoleonu, a stê¿enie
tego zwi¹zku w glebie na której roœnie sorgo dochodzi do 10
-5
-10
-4
M (19). Sorgole-
on stosowany dolistnie w stê¿eniach podobnych jak atrazyna (0,6 kg ha
-1
) hamowa³
wzrost 14-dniowych siewek ró¿nych jedno- i dwuliœciennych chwastów (40). Podob-
nie, przedsiewne stosowanie sorgoleonu by³o toksyczne dla wiêkszoœci chwastów
(41). Yang i wsp. (42) wykorzystuj¹c dwa ró¿ne systemy uprawy roœlin sorgo, w któ-
rych uda³o siê uzyskaæ siewki sorgo nie wykszta³caj¹ce w³oœników (nie syntety-
zuj¹ce sorgoleonu) i wykszta³caj¹ce prawid³owe w³oœniki (zdolne do biosyntezy sor-
goleonu) zidentyfikowali gen SOR1 warunkuj¹cy syntezê sorgoleonu. Dayan i wsp.
(43) zidentyfikowali nie znan¹ dot¹d desaturazê kwasów t³uszczowych, która mo¿e
byæ odpowiedzialna za unikatowy wzorzec rozmieszczenia nienasyconych wi¹zañ
w ³añcuchu alifatycznym cz¹steczki sorgoleonu. Bia³ko SOR1 bêd¹ce produktem
genu SOR1 pod wzglêdem sekwencji aminokwasowej wykazuje podobieñstwo do
desaturaz omega-3 kwasów t³uszczowych wystêpuj¹cych u ry¿u i pszenicy (42).
Obecnie trwaj¹ prace nad testowaniem funkcji genu SOR1 w Arabidopsis thaliana. Byæ
mo¿e, podstaw¹ w wyjaœnieniu roli genu SOR1 w kreowaniu potencja³u allelopatycz-
nego sorgo bêd¹ doœwiadczenia prowadz¹ce do uzyskania roœlin tego gatunku z nad-
ekspresj¹ lub wyciszeniem genu SOR1. Identyfikacja genu SOR1 mo¿e staæ siê kamie-
niem milowym w zastosowaniu allelopatii jako alternatywnej metody zwalczania
chwastów.
Próby genetycznych manipulacji prowadz¹cych do zwiêkszenia syntezy allelopa-
tin w roœlinach, jak na razie pozostaj¹ niestety na poziomie badañ wstêpnych. Jed-
nak nale¿y pamiêtaæ, ¿e metody in¿ynierii molekularnej mog¹ byæ niezwykle przy-
datnym narzêdziem w uzyskiwaniu ekologicznych herbicydów opartych na zwi¹z-
kach allelopatycznych. Konieczne jest ponadto prowadzenie interdyscyplinarnych
badañ, które obok identyfikacji zwi¹zków allelopatycznych i genów odpowiedzial-
nych za cechy allelopatyczne bêd¹ dotyczy³y okreœlenia mechanizmów dzia³ania al-
lelopatin i reakcji roœlin na ich obecnoœæ w œrodowisku.
5. Allelopatiny jako podstawa dla opracowania struktury nowych
herbicydów
S³onecznik (H. annuus) wykazuje silne dzia³anie allelopatyczne w stosunku do
wielu roœlin uprawnych i pospolitych chwastów (44). Z tkanek tej roœliny wyizolowa-
no ponad 100 ró¿nych zwi¹zków (niektóre charakterystyczne tylko dla s³onecznika,
np. heliannany i heliespirany) wykazuj¹cych dzia³anie podobne do syntetycznych
herbicydów. Zespó³ biochemików z pracowni Maciasa stosuj¹c s³onecznik jako roœ-
linê modelow¹ podj¹³ próby opracowania nowych herbicydów w oparciu na bada-
niach zale¿noœci pomiêdzy struktur¹ zwi¹zku allelopatycznego a jego funkcj¹ fizjo-
logiczn¹ w roœlinie akceptorze. Tak¹ strategiê mo¿na by nazwaæ „podpatrywaniem
Agnieszka Gniazdowska
48
PRACE PRZEGL¥DOWE
przyrody” (44). Zosta³a ona szczegó³owo przedstawiona w polskim opracowaniu
Maciasa i wsp. (45). Podobnie, chemiczne modyfikacje allelopatin, takich jak kostu-
nolidy: kostunolacton i cynaropikryna, produkowanych w tkankach kostusa (Saussurea
lappa) oraz karczocha (Cynara scolymus) pozwoli³y na uzyskanie ponad 20 ró¿nych
pochodnych, wykazuj¹cych aktywnoœæ herbicydow¹ w stosunku do L. rigidum
i E. crus-galli wy¿sz¹ ni¿ syntetyczny Logran (46). Szczególnie aktywne okaza³y siê
pochodne oksyetanolaktonowe otrzymane w wyniku modyfikacji kostunolaktonu
(46).
Przyk³adem skutecznego wdro¿enia do praktyki rolniczej wyników badañ nad
dzia³aniem allelopatin jest mezotrion (2-[4-(methylosulfonylo)2-nitrobenzoilo]cyklo-
heksano-1,3-dion) (nazwa produktu Callisto), herbicyd opracowany przez firmê Syn-
genta AG, u¿ywany do zwalczania chwastów w uprawach kukurydzy. Jest to synte-
Biotechnologia szans¹ dla zastosowania allelopatii jako alternatywnej metody zwalczania chwastów
BIOTECHNOLOGIA 2 (77) 42-53 2007
49
Rys. Wzory strukturalne niektórych allelopatin, mog¹cych znaleŸæ zastosowanie jako naturalne her-
bicydy.

tyczna pochodna leptospermonu (rys.) syntetyzowanego przez korzenie kuflika cy-
trynowego (Callistemon citrinus) (47). Zwi¹zek ten zosta³ odkryty przypadkowo, gdy
jeden z biologów firmy Syngenta zauwa¿y³, ¿e w jego przydomowym ogródku pod
C. citrinus nie rosn¹ inne roœliny. Zafascynowany t¹ obserwacj¹ pobra³ próbki gleby
spod C. citrinus i rozpocz¹³ proces identyfikacji substancji odpowiadaj¹cej za to zja-
wisko, a nastêpnie zsyntetyzowa³ jej pochodne. Próby wykonane z u¿yciem czyste-
go leptospermonu nie by³y obiecuj¹ce, widoczny efekt hamowania wzrostu pospoli-
tych chwastów obserwowano przy bardzo wysokiej dawce allelopatiny, wynosz¹cej
9000 gramów na hektar. Dopiero w przypadku mezotrionu (2-[4-(methylosulfony-
lo)2-nitrobenzoilo]cykloheksano-1,3-dion) bêd¹cego pochodn¹ leptospermonu wy-
kazano wysok¹ skutecznoœæ jako herbicydu stosowanego w dawkach 75-225 gra-
mów na hektar (47). Od 2000 r. Callisto jest dopuszczony do u¿ycia w Stanach Zjed-
noczonych Ameryki Pó³nocnej oraz niektórych krajach Europy w tym równie¿ w Pol-
sce. Jest to przypadek œwietnie ilustruj¹cy mo¿liwoœæ wykorzystania allelopatii do
uzyskiwania œrodków ochrony roœlin, bezpiecznych dla ludzi i œrodowiska natural-
nego. Jednoczeœnie cyjanamid oraz jego sole (g³ównie cyjanamid wapnia) s¹ synte-
tycznymi zwi¹zkami stosowanymi od ponad stu lat jako nawozy i herbicydy, jednak
dopiero w ostatnim czasie obecnoœæ tego zwi¹zku zosta³a wykazana w tkankach
wyki kosmatej (Vicia villosa) (48). Roœlina ta jest obecnie testowana w sadach i na po-
lach ry¿owych Japonii jako naturalna ochrona przed chwastami (49).
6. Ryzyko zwi¹zane z zastosowaniem allelopatii w praktyce rolniczej
6.1. Ewolucyjne wykszta³canie odpornoœci chwastów na allelopatiny
Podobnie jak w przypadku nabywania przez roœliny odpornoœci na syntetyczne
herbicydy nie mo¿na wykluczyæ mo¿liwoœci nabycia przez roœliny (chwasty) odpor-
noœci na allelopatiny. Przyk³ad chabra C. diffusa wskazuje, ¿e jego pozycja jako roœli-
ny inwazyjnej w Stanach Zjednoczonych Ameryki Pó³nocnej jest zwi¹zana prawdo-
podobnie z wra¿liwoœci¹ roœlin amerykañskich na wydzielane allelopatiny. W Euro-
pie natomiast, sk¹d pochodzi, nie wykazuje takiej ekspansywnoœci, co wiêcej roœli-
ny europejskie cechuje znacznie ni¿sza wra¿liwoœæ na wydzielan¹ przez niego kate-
chinê (50). Na podstawie dotychczasowych danych zebranych na przyk³adzie C. diffusa
sugeruje siê, ¿e w toku ewolucji mo¿e dochodziæ do nabywania przez roœliny odpor-
noœci na allelopatiny (50,51). Jednak proces ten, nie jest raptowny. Jego szybkoœæ za-
le¿y od wielu czynników m.in. presji selekcyjnej. Presja selekcyjna wywo³ana przez
allelopatiny z jednej strony, mo¿e mieæ charakter ci¹g³y i szeroki, co sprzyja³oby na-
bywaniu odpornoœci, jednak z drugiej, relatywnie niski poziom efektywnoœci zna-
nych allelopatin w warunkach polowych, obni¿a rzeczywist¹ szansê na ewolucjê tej
odpornoœci (52). Dodatkowo, fakt ¿e wiêkszoœæ allelopatin dzia³a na ró¿ne procesy
Agnieszka Gniazdowska
50
PRACE PRZEGL¥DOWE

fizjologiczne i na ró¿nych poziomach organizacji komórkowej równie¿ obni¿a praw-
dopodobieñstwo nabycia takiej odpornoœci.
6.2. Problemy zwi¹zane z wprowadzaniem genów allelopatii
Próby konstrukcji transgenicznych roœlin zdolnych do syntezy allelopatin napo-
tykaj¹ na wiele trudnoœci. Jedn¹ z nich jest obecnoœæ w roœlinie, substratu dla enzy-
mu bêd¹cego produktem genu warunkuj¹cego potencja³ allelopatyczny. Nawet, je-
¿eli substrat jest w tkance, zwiêkszenie potencja³u allelopatycznego roœliny mo¿e
byæ utrudnione z powodu zjawiska autotoksycznoœci. Unikanie autotoksycznoœci
polega na detoksykacji i/lub magazynowaniu allelopatin, np. w wakuolach. Kolejn¹
barierê stanowi tkankowo/komórkowo specyficzna biosynteza allelopatin, wyma-
gaj¹ca u¿ycia do transformacji promotora warunkuj¹cego ekspresjê wprowadzone-
go genu w œciœle okreœlonych komórkach, np. w³oœnikach lub komórkach wydzielni-
czych, tak aby mo¿liwe by³o wydzielanie allelopatiny do œrodowiska i by nie by³a
ona obecna w plonie rolniczym. Ponadto, manipulacje genetyczne prowadz¹ce do
zwiêkszenia syntezy allelopatin mog¹ doprowadziæ do niekorzystnych zmian we
wzroœcie i rozwoju roœliny (ograniczony wzrost, obni¿one plonowanie), a nadmier-
na produkcja metabolitów wtórnych mo¿e obni¿yæ wartoœæ smakow¹ plonu. Nie
mo¿na pomin¹æ faktu, ¿e zastosowanie transgenicznych roœlin allelopatycznych mo-
¿e wywo³aæ bezpoœrednie zagro¿enie dla ludzi zwi¹zane z dzia³aniem produktów
wprowadzonych genów jako alergenów lub potencjalnymi zmianami jakie mog¹ byæ
przez nie wywo³ane w œrodowisku. Wydaje siê jednak, ¿e ewentualne wzmo¿enie
cechy allelopatycznej w roœlinach uprawnych takich jak np. ry¿ czy sorgo nie powin-
no nieœæ za sob¹ niebezpieczeñstwa dla zdrowia ludzkiego, poniewa¿ zró¿nicowa-
nie stosowanych odmian pod wzglêdem potencja³u allelopatycznego waha siê od
skrajnie niskiego do bardzo wysokiego. Co najwa¿niejsze, roœliny, które pozostaj¹
w centrum uwagi biotechnologów, np. sorgo zdolne s¹ do syntezy allelopatin w wy-
specjalizowanych tkankach lub komórkach, dziêki czemu nie odnajduje siê allelopa-
tin w plonie rolniczym wykorzystywanym do produkcji po¿ywienia.
7. Podsumowanie
Poznanie biochemicznych i molekularnych mechanizmów oddzia³ywañ allelopa-
tycznych pomiêdzy roœlinami oraz genetycznego pod³o¿a warunkuj¹cego potencja³
allelopatyczny stanowi podstawê zastosowania allelopatii w rozwijaj¹cym siê rolnic-
twie zrównowa¿onym. Izolacja, okreœlenie struktury chemicznej allelopatin oraz
wyjaœnienie szlaku biosyntezy tych zwi¹zków w organizmach roœlinnych mo¿e wyty-
czyæ kierunki poszukiwania i produkcji herbicydów nowej generacji. Po³¹czenie wie-
dzy o genetycznych podstawach allelopatii roœlin uprawnych z postêpem w dziedzi-
Biotechnologia szans¹ dla zastosowania allelopatii jako alternatywnej metody zwalczania chwastów
BIOTECHNOLOGIA 2 (77) 42-53 2007
51

nie konstruowania roœlin genetycznie zmodyfikowanych mo¿e doprowadziæ w przy-
sz³oœci do otrzymania roœlin, których uprawa nie bêdzie wymaga³a stosowania do-
datkowych œrodków ochrony przed chwastami. Badaniom zmierzaj¹cym do osi¹g-
niêcia tych celów musz¹ jednak nadal towarzyszyæ próby selekcji takich genotypów
roœlin uprawnych, w których wysokiemu potencja³owi allelopatycznemu towarzy-
szyæ bêdzie wysoka jakoœæ plonów.
Literatura
1. Inderjit, Duke S. O., (2003), Planta, 217, 529-539.
2. Tellez M. R., Canel C. Rimando A. M., Duke S. O., (1999), Phytochemistry, 52, 1035-1040.
3. Macias F. A., Varela R. M., Torres A., Galindo J. L. G., Molinillo J. M. G., (2002), Chemical Ecology of
Plants: Allelopathy in Aquatic and Terrestrial Ecosystems, Eds. Inderjit, Mallik A. U., 73-87, Birkhauser
Verlag, Basel, Boston, Berlin.
4. Vyvyan J. R., (2002), Tetrahedron, 58, 1631-1646.
5. Singh H. P., Batish D. R., Kohli R. K., (2003), Crit. Rev. Plant Sci., 22, 239-311.
6. Wójcik-Wójtkowiak D., Politycka B., Weyman-Kaczmarkowa W., (1998), Allelopatia, Wyd. AR,
im. A. Cieszkowskiego, Poznañ.
7. Gniazdowska A., Oracz K., Bogatek R., (2004), Kosmos, 53, 207-218.
8. Weston L. A., Duke S. O., (2003), Crit. Rev. Plant Sci., 22, 367-389.
9. Yun K. W., Kil B., (1992), J. Chem. Ecol., 18, 1933-1940.
10. Yun K. W., Maun M. A., (1997), Can. J. Bot., 75, 1903-1912.
11. Dayan F. E., Hernandez A., Allen S. N., Moraes R. M., Vroman J. A., Avery M. A., Duke S. O., (1999),
Phytochemistry, 50, 607-614.
12. An M., Pratley J. E., Haig T., (2001), J. Chem. Ecol., 27, 395-409.
13. An M., Pratley J. E., Haig T., (2000), J. Chem. Ecol., 26, 1465-1476.
14. An M., Pratley J. E., Haig T., Lin D. L., (2005), Nonlinear. Biol. Toxicol. Med., 3, 245-260.
15. Bais H. P., Vepachedu R., Gilroy S., Callaway R., Vivanco J. M., (2003), Science, 301, 1377-1380.
16. Heap I., (2005), www.weedscience.com.
17. Praczyk T., Skrzypczak G., (2004), Herbicydy, PWRiL, Poznañ.
18. Evans D. A., (1999), Pesticide Chemistry and Bioscience: the Food Environment Challenge, Eds. Brooks G. T.,
Roberts T. R., 124-128, The Royal Society of Chemistry, Cambridge, UK.
19. Czarnota M. A., Paul R. N., Dayan F. E., Nimbal C. I., Weston L. A., (2001), Weed Tech., 15, 813-825.
20. Jose S., (2002), Chemical Ecology of Plants: Allelopathy in Aquatic and Terrestial Ecosystems, Eds. Inderjit,
Mallik A. U., 149-172, Birkhäuser Verlag, Basel, Boston, Berlin.
21. Duke S. O., (1999), Phytochemistry, 52, 805-513.
22. Bhowmik P. C., Inderjit, (2003), Crop Prot., 22, 661-671.
23. Bond W., Grundy A. C., (2001), Weed Res., 41, 383-405.
24. Duke S. O., Dayan F. E., Romagni J. G., Rimando A. M., (2000), Weed Res., 40, 99-111.
25. Duke S. O., Rimando A. M., Baerson S. R., Scheffler B. E., Ota E., Belz R., (2002), J. Pest. Sci., 27,
298-306.
26. Olofsdotter M., Bach-Jensen L., Courtois B., (2002), Plant Breed., 121, 1-9.
27. Dilday R. H., Mattice J. D., Moldenhauer K. A., Yan W., (2001), J. Crop Prod., 4, 287-301.
28. Kato-Noguchi H., Ino T., (2003), Phytochemistry, 63, 551-554.
29. Kong C., Liang W., Xu X., Hu F., (2004), J. Agric. Food Chem., 52, 2861-2865.
30. Goff S. A., Ricke D., Lan T-H, Pressing G., Wang R., Dunn M., Glazebrook J., Session A., Oeller P.,
Varma H., Hadley D., Hutchison D., Martin C., Katagiri F., Lange B. M., Moughamer T., Xia Y., Bu-
dworth P., Hong J., Migiel T., Paszkowski U., Hang S., Colbert M., et. al., (2002), Science, 296,
92-100.
Agnieszka Gniazdowska
52
PRACE PRZEGL¥DOWE

31. Yu J., Hu S., Wang J., Wong G. K., Li S., Liu B., Deng Y., Dai L., Zhou Y., Hang X., Cao M., Liu J., Sun
J., Tang J., Chen Y., Huang X., Lin W., et. al., (2002), Science, 296, 79-92.
32. Xu M., Hellwig M. L., Prusic S., Coates R. M., Peters R. J., (2004), Plant J., 39, 309-318.
33. Dayan F. E., Cook D., Baerson S. R., Rimando A. M., (2005), Proceedings of the 4
th
Word Congress on Al-
lelopathy, Eds. Harper J. D. I., An M., Wu H., Kent J. H., 175-181, Charles Sturt University, Wagga Wa-
gga, NSW, Australia.
34. Kim H. Y., Shin H. Y., Sohn D. S., Lee I. J., Kim K. U., Lee S. C., Jeong H. J., Cho M. S., (2000), Korean
J. Crop Sci., 46, 22-28.
35. Kim K. U., Shin D. H., (2002), www.fao.org./documents/.
36. Bach-Jensen L., Courtois B., Shen L., Li Z., Olofsdotter M., Mauleon R. P., (2001), Agr. J., 93, 21-26.
37. Ebana K., Yan W. G., Dilday R. H., Namai H., Okuno K., (2001), Breed. Sci., 51, 47-51.
38. Wu H., Pratley J., Ma W., Haig. T., (2003), Theor. Appl. Genet., 107, 1477-1481.
39. Wu H., Haig T., Pratley J., Lemerle D., An M., (2000), J. Chem. Ecol., 26, 2141-2154.
40. Nimbal C. I., Yerkes C. N., Weston L. A., Weller S. C., (1996), Pest. Biochem. Physiol., 54, 73-83.
41. Weston L. A., Czarnota M., (2001), J. Crop Prod., 4, 363-377.
42. Yang X., Scheffler B. E., Weston L. A., (2004), J. Exp. Bot., 55, 2251-2259.
43. Dayan F. E., Kagan I. A., Rimando A M., (2003), J. Biol. Chem., 278, 28607-28611.
44. Macias F. A., Molinillo J. M. G., Galindo J. C. G., Varela R. M., Simonet A. M., Castellano D., (2001),
Allelopathy in Agroecosystems, Eds. Kohli R. K., Singh H. P., Batish D. R., 237-256, Food Products Press
an Imprint of the Howarth Press, Inc. New York, London, Oxford.
45. Macias F. A., Simonet A. M., Oleszek W., (2001), Biochemiczne oddzia³ywania œrodowiskowe, red. Ole-
szek W., G³owniak K., Leszczyñski B., 47-60, AM, Lublin.
46. Macias F. A., Vinolo V. M. I., Molinillo J. M. G., (2005), Proceedings of the 4
th
Word Congress on Allelopa-
thy, Ed. Harper J. D. I., An M., Wu H., Kent J. H., 477-480, Charles Sturt University, Wagga Wagga,
NSW, Australia .
47. Cornes D., (2005), Proceedings of the 4
th
Word Congress on Allelopathy, Eds. Harper J. D. I., An M., Wu
H., Kent J. H., 569-572, Charles Sturt University, Wagga Wagga, NSW, Australia.
48. Kamo T., Hiradate S., Fujii Y., ( 2003), J. Chem. Ecol., 29, 275-283.
49. Fujii Y., Hiradate S., (2005), Proceedings of the 4
th
Word Congress on Allelopathy, Ed. Harper J. D. I., An
M., Wu H., Kent J. H., 73-76, Charles Sturt University, Wagga Wagga, NSW, Australia.
50. Callaway R. M., Ridenour W. M., (2004), Front Ecol. Environ., 2, 436-443.
51. Gniazdowska A., (2005), Kosmos, 54, 221-226.
52. Scheffler B. E., Duke S. O., Dayan F. E., Ota E., (2001), Recent Adv. Phytochem., 35, 257-273.
Biotechnologia szans¹ dla zastosowania allelopatii jako alternatywnej metody zwalczania chwastów
BIOTECHNOLOGIA 2 (77) 42-53 2007
53
Wyszukiwarka
Podobne podstrony:
PTASIE GNIAZDA(1) ppt
Orle Gniazdo
gniazda rozszerzen
Interfejs programowy Gniazda BSD
Podstawowe wymiary gniazd wkretow
Ptasie gniazda, PRZEPISY Pizza i cos jeszcze
GNIAZDO ŚWIATEŁ URSUS C360
gniazdka
Gniazdo nad głową(1), wiosna
Magistrala i gniazda rozszerzające
GNIAZDA I PORTY ETC
sprawozdanie pomiary bledow ksztaltu i polozenia osi otworow gniazd lozyskowych
7 pinowe gniazdo w samochodzie i przyczepie Opis
Programowanie sieciowe przy użyciu gniazdek w?lphi 3 MPLMRFGCOQC4VOMKHU5DAT5YKUDWUHLWUTINXRI
Interfejs gniazd
obwody szkic tech gniazda
Opis pinow gniazda OBD2
lokalizacja gniazd diagnostycznych
więcej podobnych podstron