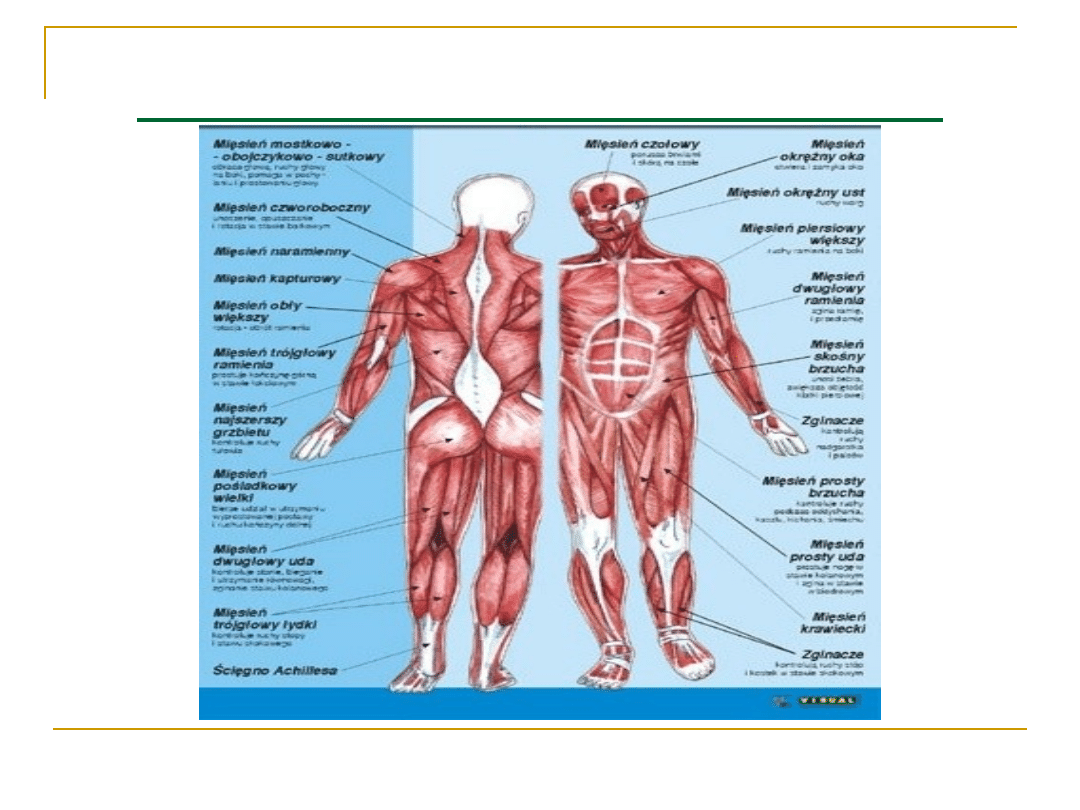
FIZJOLOGIA MIĘŚNI
TKANKA
MIĘŚNIOWA
:
POPRZECZNIE
PRĄŻKOWANA
SZKIELETOWA
GŁADKA
POPRZECZNIE
PRĄŻKOWANA
SERCA
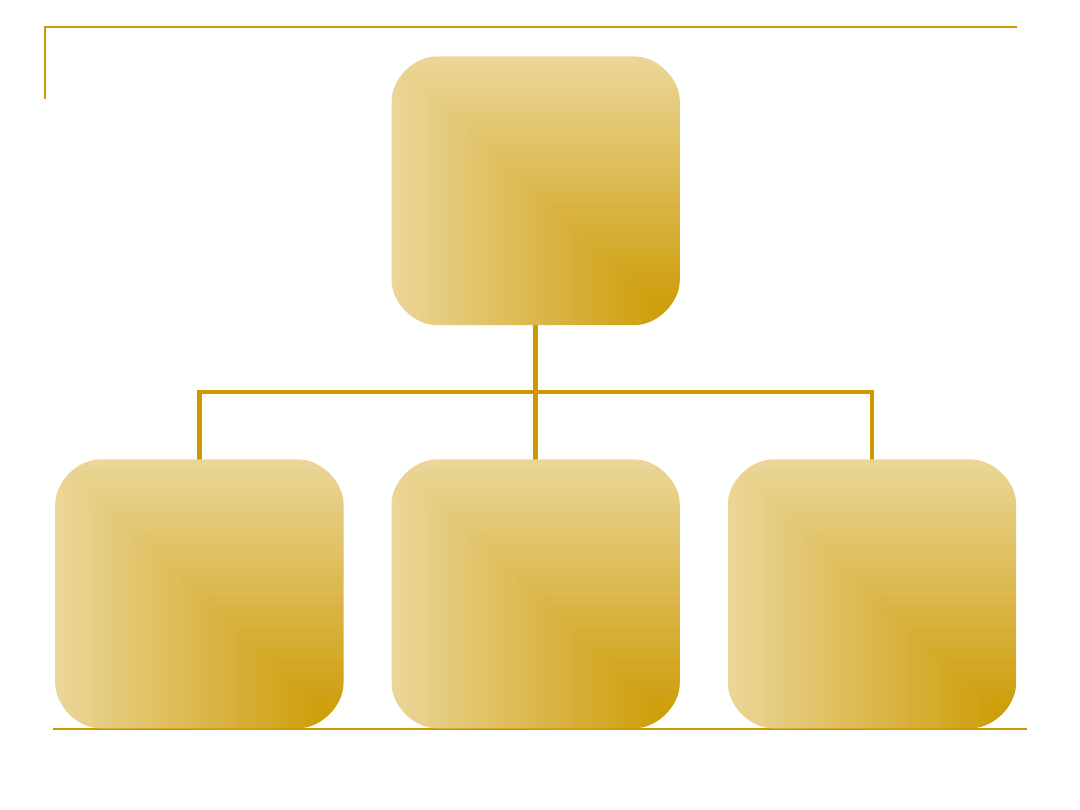
Podział mięśni
Tkanka mięśniowa
poprzecznie prążkowana
gładka
mięśnie szkieletowe
mięsień serca
mięśnie
jednostkowe
mięśnie
wielojedno-
stkowe
mięśnie trzewne

MIĘŚNIE
SZKIELETOWE
Narząd
wykonawczy,
odpowiedzialny
za czynność
motoryczną
człowieka i
utrzymanie
postawy ciała.

Rola w czynności organizmu
mięśni szkieletowych:
Czynność mechaniczna zachodzi w taki sposób aby
czas od pobudzenia do skurczu był jak najkrótszy i
była jednakowa w takich samych warunkach;
Drogi korowo rdzeniowe posiadają nieliczne
synapsy;
Ruchowe włókna nerwowe należą do najszybciej
przewodzących impulsację nerwową
Płytka motoryczna jest synapsą chemiczną
działającą z najmniejszym opóźnieniem;
Mechanizm sprzężenia elektomechanicznego jest
szybki i sprawny.

Czynność bioelektryczna
mięśni szkieletowych:
Potencjał spoczynkowy jest bardziej
elektroujemny i wynosi ok. 90 mV
Depolaryzacja jest nieco dłuższa trwa ok.2
ms;
Całkowity czas trwania potencjału jest
dłuższy- do 10ms.
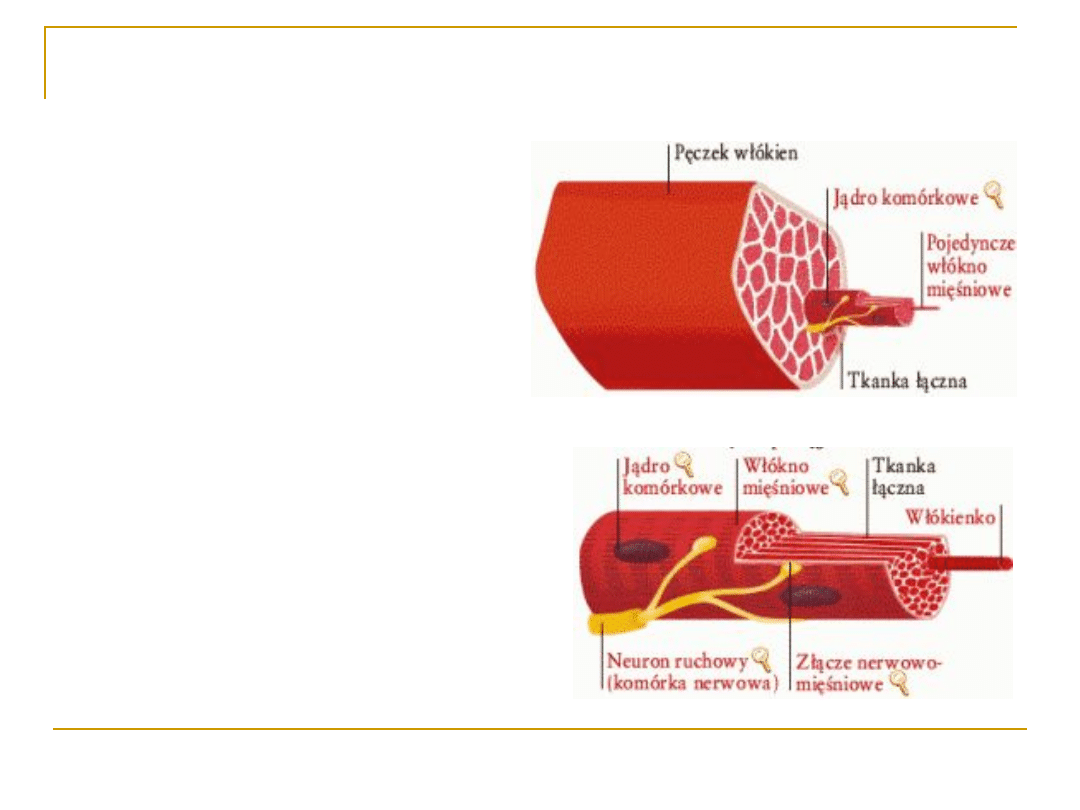
Budowa mięśni
szkieletowych:
Jednostką morfologiczną jest
włókno mięśniowe;
Włókna mięśni mają kształt
cylindryczny, posiadają liczne
jądra komórkowe;
Włókna mięśniowe biegną od
przyczepu do przyczepu mięśnia;
Błonę komórki mięśniowej
nazywamy SARKOLEMMĄ, a
cytoplazmę SARKOPLAZMĄ;
W strukturze włókna
mięśniowego wyróżniamy:
System białek kurczliwych
(przekształcanie energii
chemicznej na mechaniczną)
Błonowy system kontrolny.
Białka kurczliwe
(system białek kurczliwych)
Miozyna
Aktyna
Tropomiozyna
Troponina
Filamenty grube
Filamenty cienkie

Mięśnie szkieletowe – główne cechy
Stanowią główną masę mięśniową w układzie somatycznym
Rozwinięte prążkowanie poprzeczne
Nie mają anatomicznych i czynnościowych powiązań
między poszczególnymi włóknami
Nie kurczą się przy braku pobudzeń nerwowych
Podlegają świadomej kontroli
Budowa mięśni
szkieletowych
Do białek kurczliwych
należą:
MIOZYNA
(tworząca filamenty
grube))
AKTYNA
TROPO
MIOZYNA
TROPONIN
A
-
białko o m.c.
460000 D
-zbud. z 2
identycznych
łańcuchów
meramiozyny
ciężkiej oraz 2 par
łańcuchów
meramiozyny
lekkiej.
-łańcuchy ciężkie
zbudowane są z
globularnej głowy
i ogonów
splecionych wokół
siebie.
-każda główka
posiada miejsca
do łączenia z
aktyną.
-masa
cząsteczkowa
43000 D
-jest cząsteczką
białka
fibrylarnego,
zbudowanego z
monomerów
aktyny G
(globularnej)
-każdy monomer
posiada centrum
aktywne
wykazujące
powinowactwo
do miozyny.
-cząsteczka o m.
cz. 70000 D i
budowie liniowej
-oplata cz. Aktyny
F, zasłaniając
swoją strukturą
centra aktywne
-gdy mięsień jest
w spoczynku
uniemożliwia
powstawanie
aktywnego
kompleksu aktyna-
miozyna.
-zbudowana z 3
podjednostek: I, T,
C.
-troponina przez
podjednostkę T
łączy się z
tropomiozyną, a
przez podjednostkę
I z aktyną
-stabilizuje
połozenie
filamentów
względem siebie
-podjednostka C
wskazuje
powinowactwo do
jonów Ca
2+
.
Tworzące łącznie filamenty cienkie
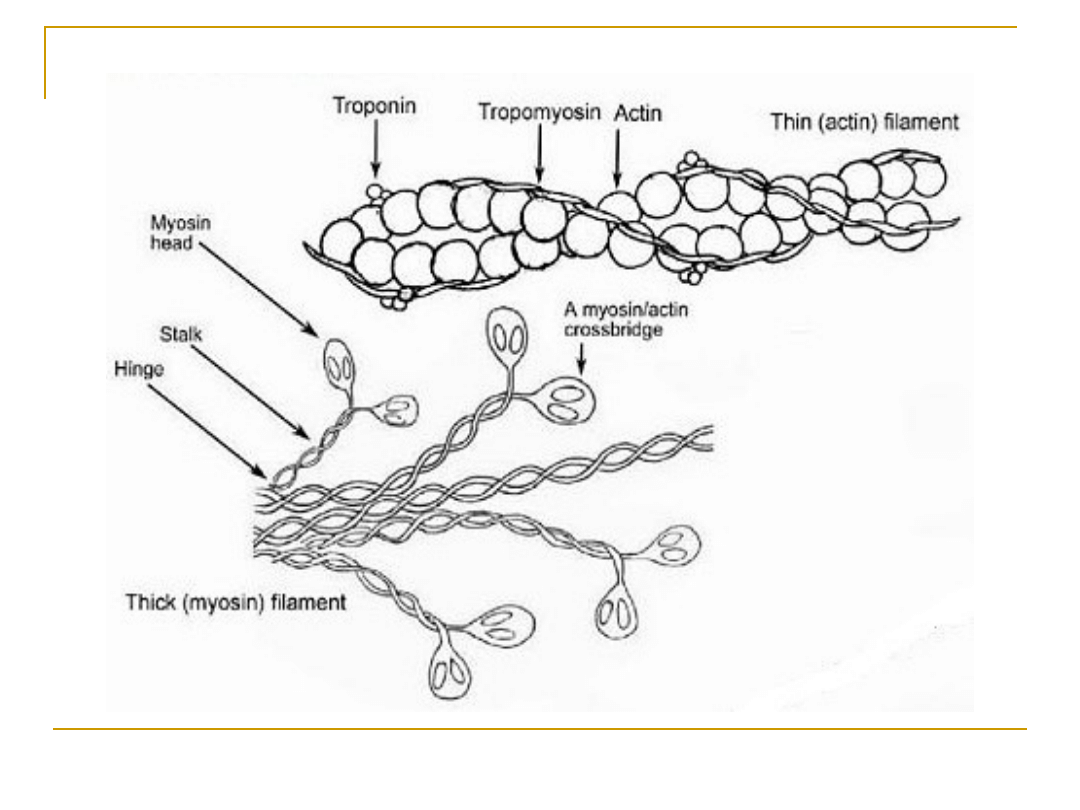

W skład błonowego systemu
kontrolnego wchodzą:
Błona komórkowa
Kanaliki poprzeczne (T)- stanowią
uwypuklenie sarkolemmy. Układ utworzony
przez kanalik poprzeczny T i dwa zbiorniki
końcowe zlokalizowane po obu stronach tego
kanalika nazywany jest TRIADĄ
MIĘŚNIOWĄ.
Siateczka sarkoplazmatyczna- jest bardzo
silnie rozbudowana i tworzy gęstą sieć
oplatającą białka kurczliwe. Składa się ze
zbiorników końcowych oraz ułożonego
pomiędzy nimi systemu kanalików podłużnych.
W jej wnętrzu magazynowane są jony wapnia.
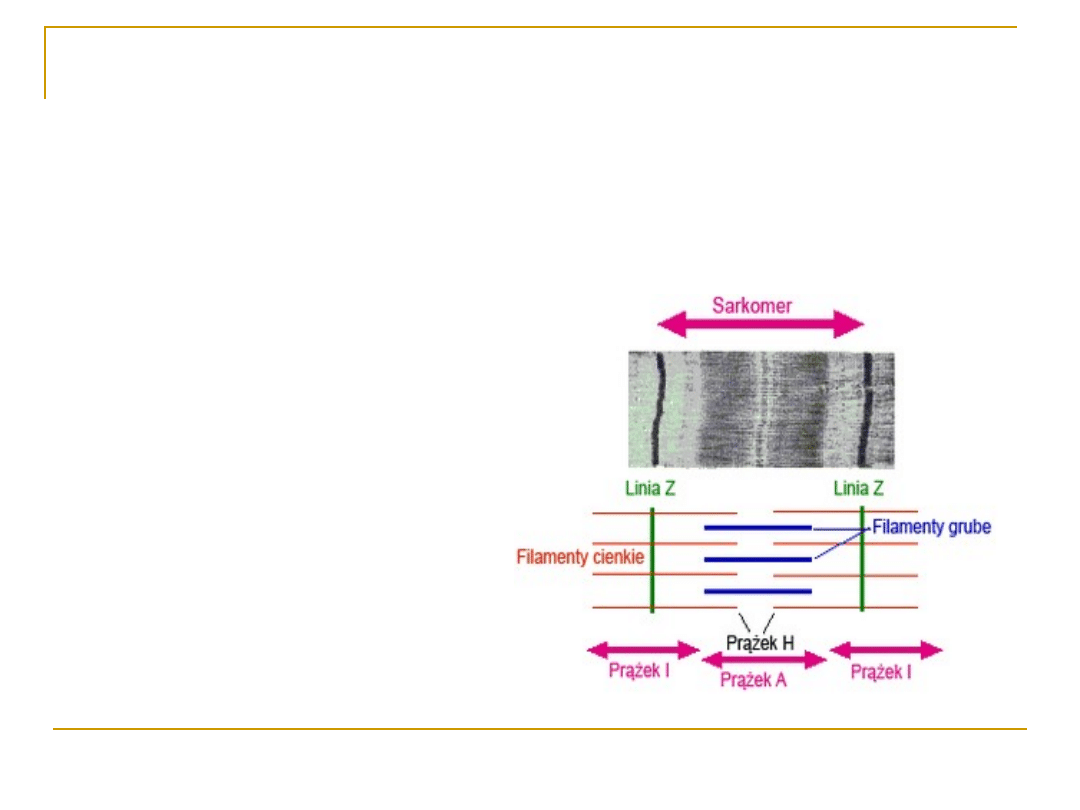
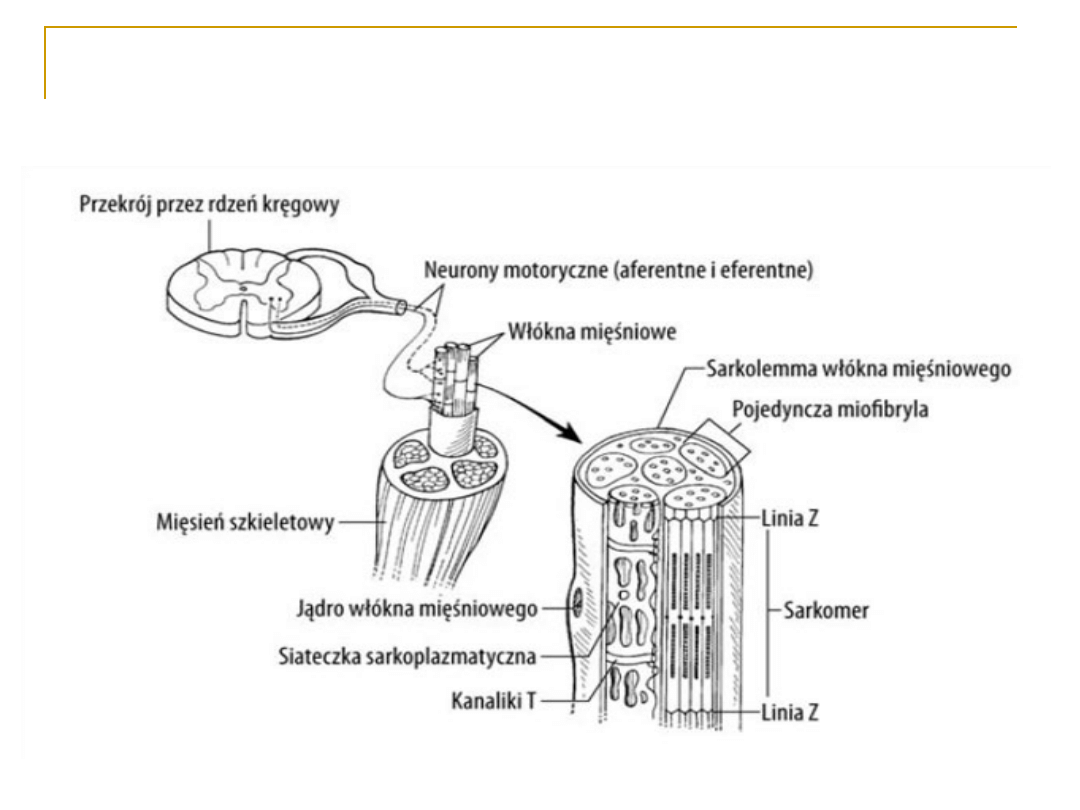
Sarkomer – jednostka czynnościowa
włókna mięśniowego
- prążki jasne
(izotropowe): aktyna
{kind=link}
- prążki ciemne
(anizotropowe):
miozyna
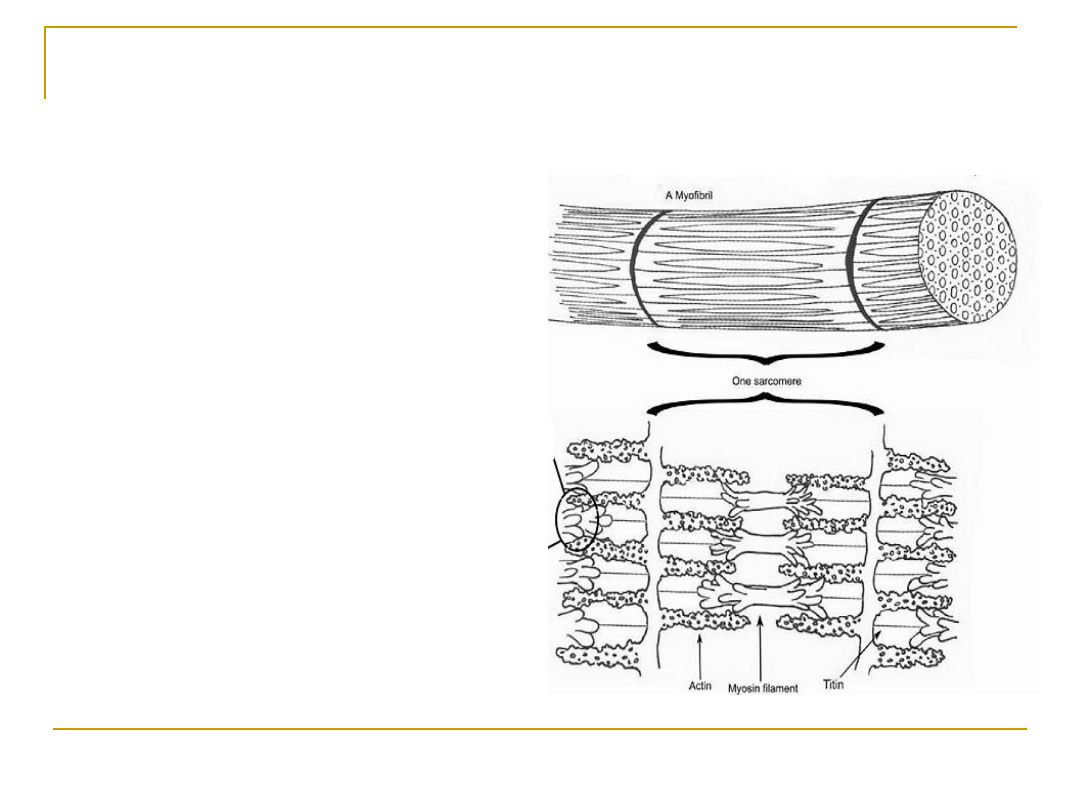
SARKOMER
To jednostki czynnościowe włókien
mięśnia szkieletowego;
Jest ograniczony liniami
granicznymi Z (linie Z to
zagęszczenia sarkoplazmy, przez
które przechodzą filamenty
aktynowe. W obrębie tych linii f. a.
umocowane są za pomocą
włókienek titinowych.)
Obszary sarkomeru, gdzie
występuje tylko aktyna tworzą
prążki jasne I (izotropowe)
Część centralna sarkomeru zajmuje
ciemny prążek A (anizotropowy)
W części centralnej prążka A
widoczna jest strefa przejaśnienia
Hansenowskiego (prążek H), w
którym występuje tylko miozyna.
Teoria ślizgowa HUXLEYA
( molekularny mechanizm skurczu m.
szkieletowego)
Jeżeli pobudzenie nerwowe przestanie docierać do mięśnia, wapń
zostaje wypompowany do retikulum sarkoplazmatycznego; jego
stężenie wraca do wartości spoczynkowych. Możliwy jest bierny
rozkurcz mięśni.

Typy włókien mięśniowych:
EKSTRAFUZALNE
(zewnątrzwrzecionowe)
Stanowią zasadniczą masę
mięśnia, odpowiedzialna
za czynność
mechaniczną; biegną od
przyczepu do przyczepu
na całej długości i
wykazują poprzeczne
prążkowanie.
Przyczepiają się do nich
WRZECIONKA
MIĘŚNIOWE.
INTRAFUZALNE
(wewnątrzwrzecionowe)
Otoczone torebką
łącznotkankową;
poprzecznie prążkowane;
ich czynność mechaniczna
występuje tylko w
częściach biegunowych.
W części centralnej
zakończenia czuciowe
wrażliwe na rozciąganie-
receptory pierścieniowo-
spiralne (pierwotne) oraz
bukietowate (wtórne).

Jednostka motoryczna
Każdą jednostkę motoryczna tworzy MOTONEURON
α, jego wypustka osiowa i wszystkie włókna
ekstrafuzalne unerwiane są przez ten motoneuron.
Mięśnie szkieletowe zbudowane są z małych lub
dużych jednostek.
Każda jednostka motoryczna jest uruchamiana
niezależnie od pozostałych, dlatego możliwa jest
regulacja siły skurczu mięśnia szkieletowego poprzez
włączanie odpowiedniej do wielkości pokonywanego
oporu liczby jednostek. Zjawisko to nazywamy
REKRUTACJĄ JEDNOSTEK MOTORYCZNYCH.
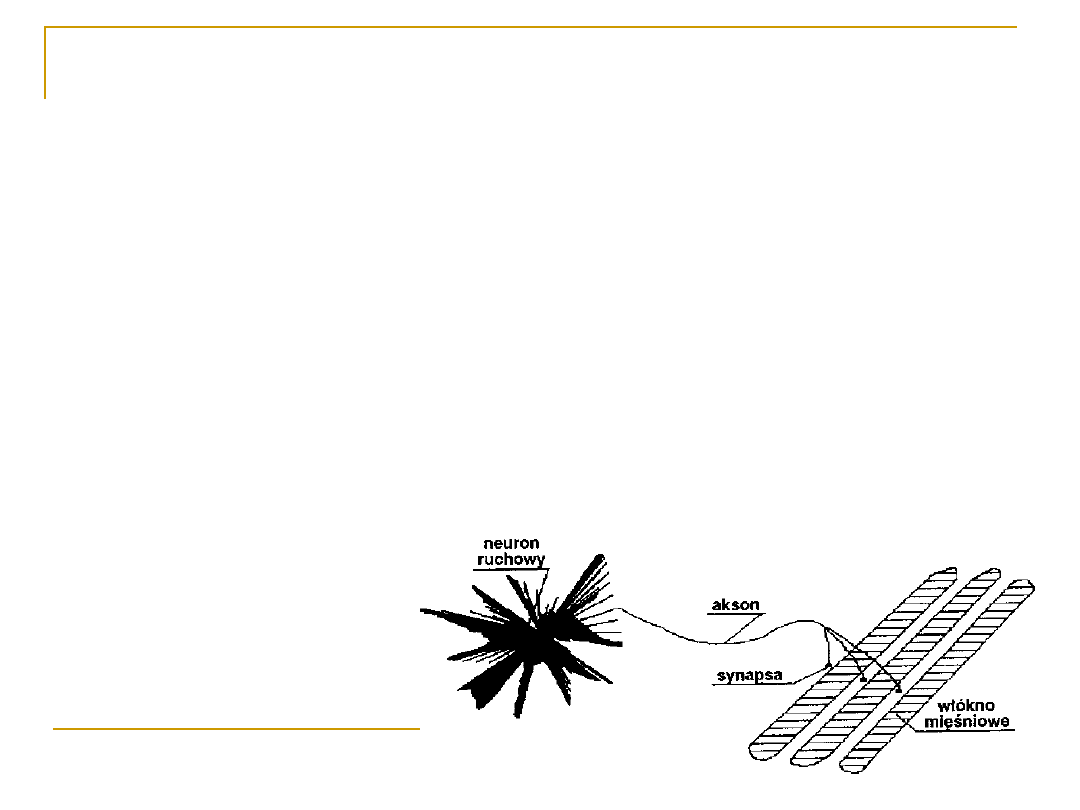
Jednostka motoryczna
Każdy pojedynczy neuron
ruchowy alfa i unerwiane
przez niego
włókna
ekstrafuzalne
tworzą
jednostkę motoryczną
Liczba włókien w jednostce
jest różna, w zależności od
typu mięśni

Typy włókien mięśniowych
(podział II)
Typ I
Czerwone
Nieprecyzyjne, zmiany długości i napięcia zachodzą skokowo,
Duże jednostki motoryczne (kilkaset włókien)
Przystosowane do długotrwałych wolnych skurczów (czas trwania
skurczu pojedynczego średnio ok. 100 ms) utrzymujących postawę
np. mięśnie grzbietu
Duża ilość mioglobiny
Metabolizm oparty na fosforylacji oksydacyjnej, praca bez szybko
narastającego zmęczenia
(zdolność oksydacyjna koreluje z zawartością mitochondriów, naczyń
włosowatych i zawartością mioglobiny
Przykłady: mięśnie statyczne, siłowe , np. grzbietu, obręczy
biodrowej, brzucha i kończyn dolnych

Typy włókien mięśniowych
(podział II)
Typ II
Szybkie
Białe
wykonują precyzyjne, zręczne ruchy
Małe jednostki motoryczne (kilka-kilkadziesiąt włókien)
Mała ilość mioglobiny
Podtyp IIa – metabolizm oparty na glikolizie tlenowej –
odporne na zmęczenie
IIb – metabolizm oparty na glikolizie beztlenowej –
najszybciej ulegają zmęczeniu
Przykłady: mięśnie okoruchowe, mimiczne, odpowiedzialne
za artykulacje mowy, mięśnie kończyn górnych

Mięśnie szkieletowe dzielimy
na:
TYP I (NIEPRECYZYJNE)
Czerwone
Wolne, czas trwania
skurczu pojedynczego
średnio ok.. 100ms
Duża ilość mioglobiny
Metabolizm oparty na
fosforylacji oksydatywnej,
duże zasoby substratów
energetycznych, praca bez
szybko narastającego
zmęczenia
Mięśnie statyczne, siłowe,
zaliczamy do nich: m.
grzbietu, obręczy
biodrowej, brzucha i
kończyn dolnych.
TYP II (PRECYZYJNE)
Białe
Szybkie, czas trwania skurczu
pojedynczego od 7,5 do
kilkudziesięciu ms
Mała ilość mioglobiny
Podtyp II a- dominuje
metabolizm oparty na glikolizie
tlenowej, względnie odporne na
zmęczenie
Podtyp II b- metabolizm oparty
na glikolizie beztlenowej, małe
zasoby substratów
energetycznych, najszybciej
ulęgają zmęczeniu
Zaliczamy do nich: m.
okoruchowe,
mimiczne
.

Mięśnie szkieletowe – główne cechy
Stanowią główną masę mięśniową w układzie somatycznym
Rozwinięte prążkowanie poprzeczne
Nie mają anatomicznych i czynnościowych powiązań
między poszczególnymi włóknami
Nie kurczą się przy braku pobudzeń nerwowych
Podlegają świadomej kontroli

Płytka motoryczna
Rodzaj synapsy chemicznej służącej przekazaniu
pobudzenia z aksonu komórki nerwowej na komórkę
mięśnia szkieletowego; synapsa, w której błoną
postsynaptyczną jest wyspecjalizowany fragment
sarkolemmy , o licznych wpukleniach, zawierający
kanały jonowe bramkowane chemicznie ;
neuroprzekaźnikiem w p. m. jest acetylocholina
działająca na receptory cholinergiczne
nikotynowe typu mięśniowego w błonie
postsynaptycznej.
Sprzężeniem elektromechanicznym nazywamy
mechanizm powstawania aktywnego kompleksu
AKTYNA- MIOZYNA i skurczu mięśnia poprzez
wzrost stężenia jonów wapnia w sarkoplaźmie.

Układ sarkotubularny
(błonowy system kontrolny)
Błona komórkowa (sarkolemma) -
kanaliki T
Siateczka śródplazmatyczna (retikulum
sarkoplazmatyczne)
Triada mięśniowa
: kanalik poprzeczny T + dwa zbiorniki
końcowe siateczki sarkoplazmatycznej zlokalizowane po
obu stronach kanalika
Receptory :
dihydropirydynowy, rianodynowy
Receptor dihydropirydynowy - zależne od napięcia kanały
wapniowe
Receptor rianodynowy - niezależny od napięcia kanał wapniowy
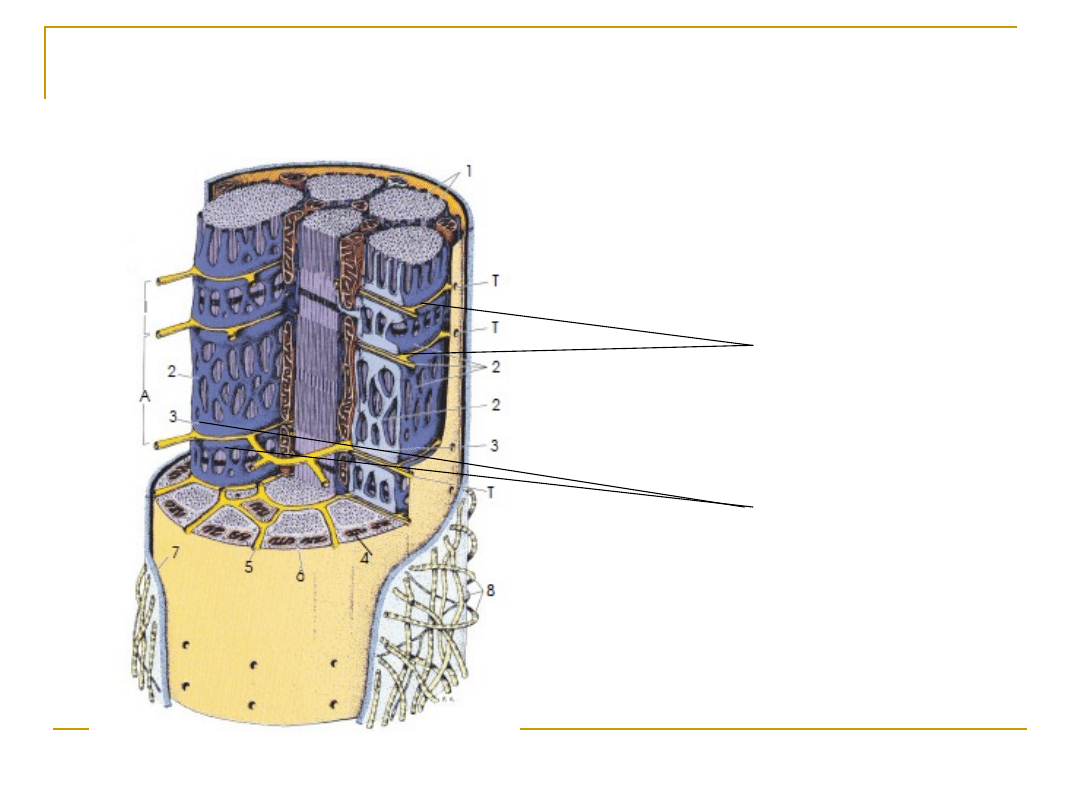
Układ sarkotubularny
Kanaliki T
Zbiorniki końcowe
siateczki
sarkoplazmatycznej
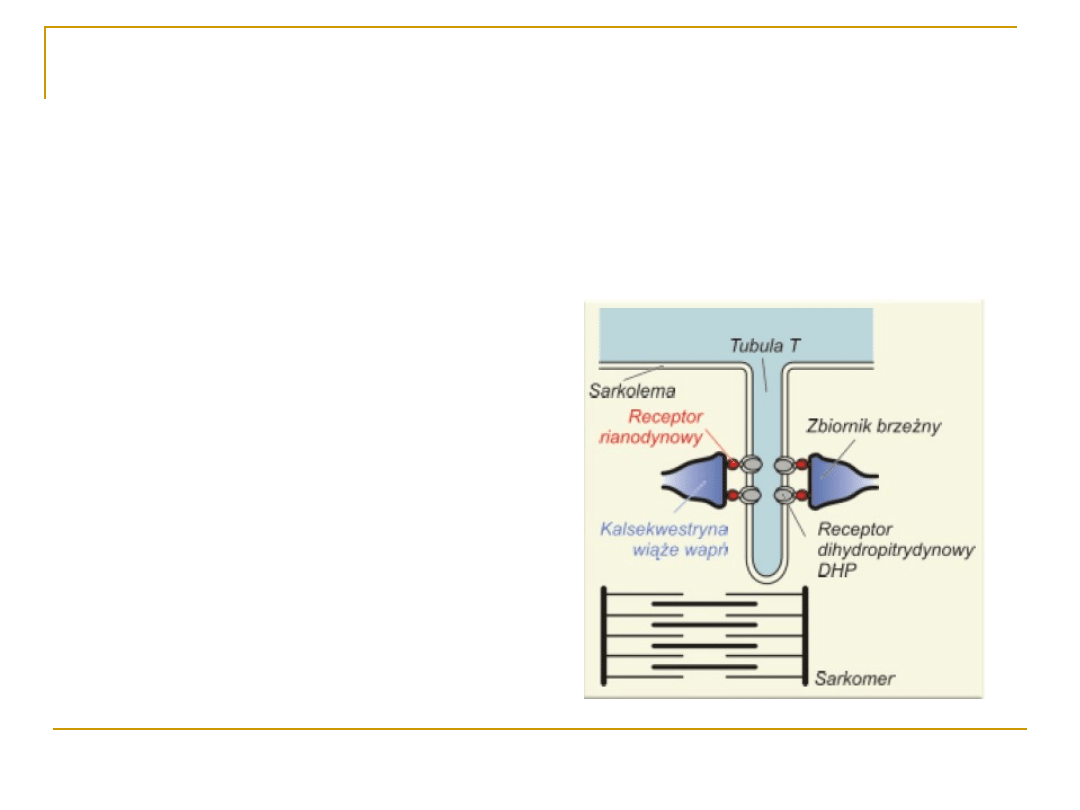
Rola układu sarkotubularnego w
skurczu mięśnia
1. Depolaryzacja kanalików T
2. Zmiana konformacji receptorów
dihydropirydynowych w
sarkolemmie
3. Zmiana konformacji receptorów
rianodynowych,
otwarcie kanałow wapniowych
4.
Wypływ Ca
2+
z siateczki
sarkoplazmatycznej do
sarkoplazmy
5. Skurcz włókna mięśniowego

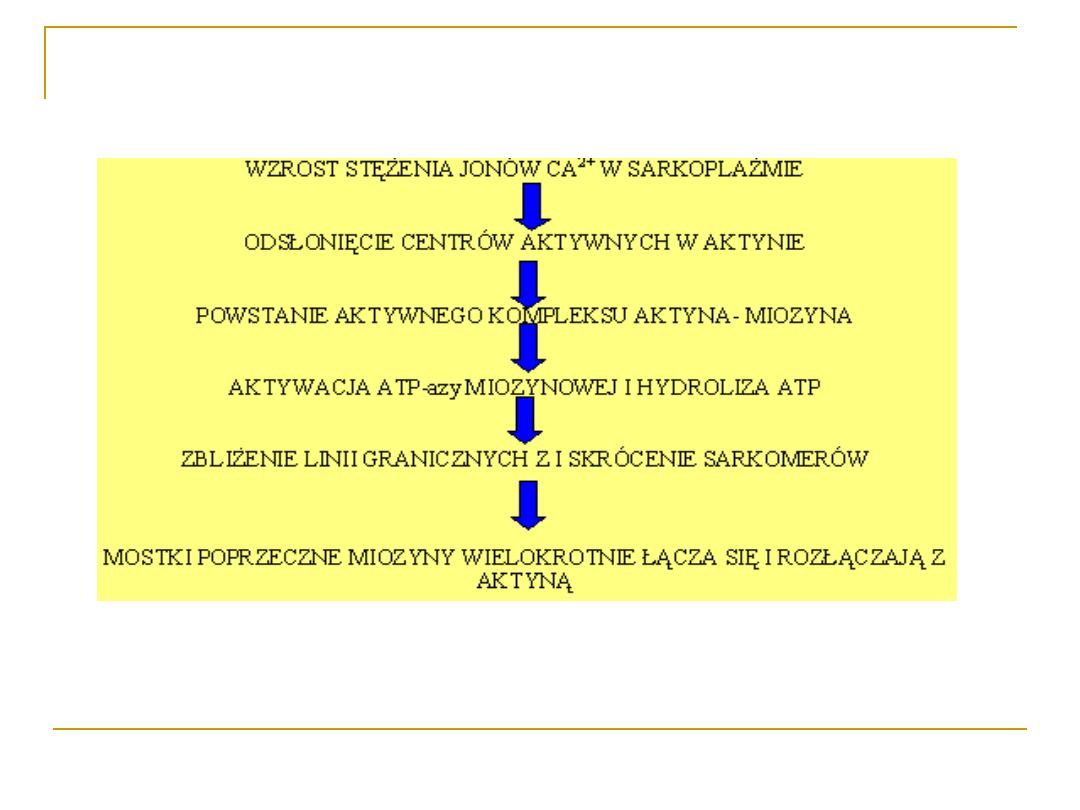
Molekularny mechanizm skurczu
mięśnia-podsumowanie
1. Uwalnianie jonów Ca
2+
za zabiorników końcowych siateczki
sarkoplazmatycznej, ich dyfuzja do grubych i cienkich
filamentów.
2. Wiązanie jonów Ca
2+
z troponiną C, odsłanianie miejsc
połączeń miozyny z aktyną.
3. Osłabienie wiązania troponiny I z aktyną.
4. Przesunięcie boczne tropomiozyny.
5. Odsłonięcie miejsc połączeń między aktyną a główkami
miozyny.
6. Aktywacja ATP-azy miozynowej.
7. Hydroliza ATP, uwolnienie energii.
8. Zgięcie mostka poprzecznego u nasady główki miozyny i
jednoczesne pociągnięcie nici aktynowej wzdłuz miozynowej.
9. Zbliżenie linii granicznych Z i skrócenie sarkomerów
(proces powtarzany jest tak długo dopóki utrzymywane jest
odpowiednie stężenie jonów wapnia w sarkoplazmie).

Typy włókien mięśniowych
Włókna ekstrafuzalne:
Stanowią
zasadnicza masę mięśnia
Wykazują poprzeczne
prążkowanie
Przebieg od przyczepu do
przyczepu
Unerwiane przez
motoneurony alfa
Włókna intrafuzalne
„przyczepiają się” do
włókien ekstrafuzalnych,
tworząc wrzecionka
Poprzeczne prążkowanie
tylko w częściach
biegunowych
W części centralnej:
zakończenia czuciowe
wrażliwe na rozciąganie
Unerwiane przez
motoneurony gamma

Rodzaje skurczów mięśni
szkieletowych:
1
. W zależności od
częstotliwości bodźców
pobudzających:
a)
Skurcze pojedyncze- następują w
momencie pobudzenia włókna
mięśniowego pojedynczym
impulsem. Wyróżniamy w nim fazę
skurczu i rozkurczu. Skurcz taki
można wywołać tylko w warunkach
IN VITRO, natomiast w warunkach
IN VIVO skurcze te nie występują.
b)
Skurcze tężcowe:
•
Niezupełne- powstają poprzez
docieranie kolejnych pobudzeń w
fazie rozkurczu.
•
Zupełne- powstają poprzez
docieranie kolejnych pobudzeń w
fazie skurczu.
2. W zależności od
warunków w jakich
mięsień jest pobudzany
:
a)
Skurcz izotoniczny- zachodzi kiedy
przy pobudzaniu mięśnia jeden lub
oba przyczepy są swobodne.
Następuje zmiana długości mięśnia,
bez zmiany jego napięcia.
b)
Skurcz izometryczny- zachodzi
kiedy przyczepy są unieruchomione i
mięsień nie może się skracać.
Obserwujemy wtedy tylko zmianę
napięcia mięśnia.
c)
Skurcz auksotoniczny- zachodzi
kiedy podczas skurczu występują
obie składowe- najpierw
izometryczna,a następnie
izotoniczna.

Rodzaje skurczów
Skurcz izometryczny („taka sama
długość”)
Skurcz izotoniczny („takie samo
napięcie”)
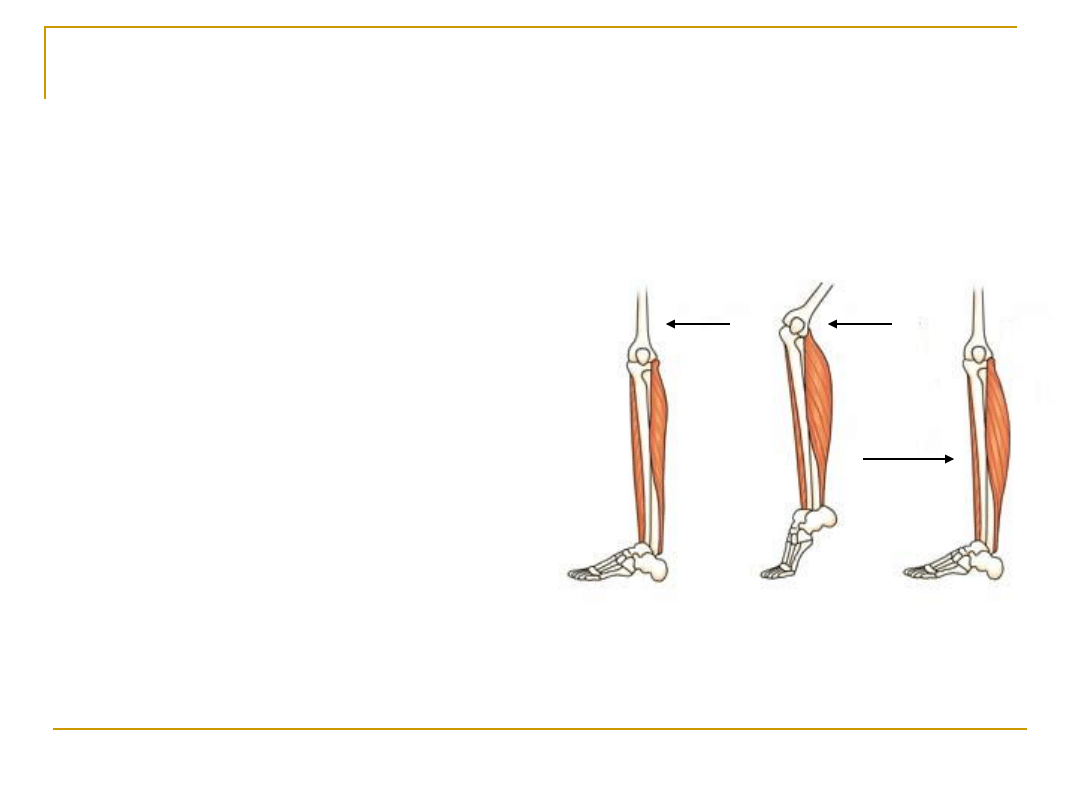
Rodzaje skurczów
Skurcz izometryczny
: nie
zmienia się długość
mięśnia, ale zmienia się
jego napięcie
Skurcz izotoniczny
:
zmienia się długość
mięśnia, ale nie zmienia
się jego napięcie
Mięsień
rozluźnion
y
Mięsień
napięty
Mięsień
nie
rozluźnia
się
powoduja
c skurcz
przebieg skurczu

Sumowanie się skurczów
W warunkach in vivo skurcze pojedyncze
nie
występują
. Przy pewnej częstotliwości bodźca
dochodzi do sumowania się skurczów pojedynczych
Skurcz tężcowy
zupełny
(bodźce pobudzają mięsień
w odstępach czasu krótszych niż trwanie skurczu
Skurcz tężcowy
niezupełny
– pobudzenie mięśnia w
odstępach czasu dłuższych niż czas trwania skurczu
pojedynczego pozwala na częściowy rozkurcz
mięśnia pomiędzy bodźcami

Źródła energii i metabolizm
Bezpośrednim źródłem energii
wykorzystywanym przez mięśnie jest
ATP
Niezbędna jest
resynteza ATP
:
-
rozpad fosfokreatyny
-
Glikoliza - łańcuch oddechowy
-
Glikogenoliza (glikogen glukozo-1-
fosforan)
-
Lipoliza (triglicerydy
wolne kwasy
tłuszczowe)
fosforylaza
lipaza

Źródła energii i metabolizm
Fosfokreatyna + ADP
kreatyna+ATP
Glukoza pirogronian +ATP
Pirogronian CO
2
+H
2
O +ATP
Glikogen glukoza
Tłuszcze kwasy tłuszczowe (B-
oksydacja)
glikoliza
glikogenoliza
lipoliza

Źródła energii pracujących
mięśni:
Jedynym, bezpośrednim źródłem energii wykorzystywanym
przez mięśnie jest ATP. Zasoby ATP w miocycie są
ograniczone i pozwalają na 20-30 sekund intensywnej
pracy, następnie energię czerpiemy ze spożywanych
pokarmów (m. in. węglowodanów).
W warunkach intensywnej pracy, stresu , głodzenia jako
substraty energetyczne wykorzystywane są również białka.
Mięsień wykorzystuje również energię wiązań chemicznych
dla wykonania pracy mechanicznej. Jednocześnie część tej
energii zostaje przekształcona w ciepło i uwolniona do
otoczenia.

Utrzymanie i regulacja napięcia
mięśniowego:
Unerwione mięśnie szkieletowe posiadają stałe bazowe
napięcie. Jest ono utrzymywane na drodze odruchowej
( odruch na rozciąganie) przez wrzecionko mięśniowe.
Za regulację (zmianę) napięcia mięśnia
odpowiedzialny jest układ pozapiramidowy (jądra
kresomózgowia). Z jąder tych wysyłane jest pobudzenie
nerwowe do motoneuronów γ, unerwiających włókna
intrafuzalne. Pobudzenie tych włókien powoduje kurczenie
się ich części biegunowych i jednoczesne wydłużanie
okolicy środkowej, co wiąże się z rozciągnięciem
znajdujących się tam receptorów pierścieniowo-
spiralnych. Konsekwencją tego jest uwrażliwienie
receptorów. W efekcie utrzymywane przez mięsień
napięcie będzie większe.

Dług tlenowy
Niedostateczne zaopatrzenie mięśni w
tlen
„Realizacja” beztlenowych etapów
oddychania (glikoliza)
Przekształcanie pirogronianu w kwas
mlekowy
Kwas mlekowy = zakłócenie
funkcjonowania włókna mięśniowego
(spadek pH)

MIĘŚNIE GŁADKIE
Znajdują się w
obrębie
narządów
wewnętrznych
organizmu. Ich
czynność
przebiega poza
zakresem naszej
świadomości.

BUDOWA mięśni gładkich:
Zbudowane z jednojądrzastych komórek;
Białka kurczliwe nie są przestrzennie
uporządkowane-brak sarkomerów i
poprzecznego prążkowania;
Białkiem akceptorowym dla jonów Ca
2+
jest
kalmodulina;
Występują ciałka gęste, porządkujące
ułożenie filamentów aktynowych;
Siateczka sarkoplazmatyczna słabo
rozwinięta, dlatego zasoby wapnia
wewnątrzkomórkowego są niewielkie.

Cechy mięśni gładkich
Jednojądrzaste komórki
Brak sarkomerów i poprzecznego prążkowania
Białkiem akceptorowym Ca
2+
jest kalmodulina, a nie
troponina
Słabo rozwinięta siateczka sarkoplazmatyczna. Dla
rozpoczęcia skurczu niezbędny jest napływ jonów wapnia ze
środowiska zewnątrzkomórkowego (mięśnie
wapniozależne).
Potencjał spoczynkowy mięsni gładkich jest niestabilny i
waha się pomiędzy -35 do -65 mV
Mogą być pobudzane przez bodźce pozanerwowe np.
hormony
Zakończenia nerwowe nie wytwarzają połączeń
synaptycznych

Mięśnie gładkie
Skurcze mięsni gładkich – niezależne od
naszej woli
Odmienne od mięśni poprzecznie
prążkowanych pod względem budowy,
czynności bioelektrycznej, unerwienia i
mechanizmu skurczu

CZYNNOŚĆ
BIOELEKTRYCZNA mięśni
gładkich:
Potencjał spoczynkowy jest mniej
elektroujemny i niestabilny. Waha się w
granicach -35 do -65 mV.
Pod wpływem zmian stężenia jonów,
hormonów, temperatury potencjał
spoczynkowy może wykazywać
tendencję HIPO lub
HIPERPOLARYZACYJNĄ.

MECHANIZM SKURCZU
mięśni gładkich:
Pod wpływem rozciągania kurczą się nawet
bez udziału innych czynników
pobudzających. Mogą być również
pobudzane przez bodźce pozanerwowe np..
hormony. Zjawisko to nazywamy
AUTOMATYZMEM MIĘŚNI GŁADKICH.
Mechanizm depolaryzacji zależy głównie od
zmian przepuszczalności jonów wapnia,
dlatego narastanie i wygasanie pobudzenia
jest powolne
.

UNERWIENIE mięśni
gładkich:
Zakończenia nerwowe nie wytwarzają
połączeń synaptycznych. Na swoim
przebiegu wytwarzają zgrubienia tzw.
żylakowatości, z których neurotransmiter
pod wpływem przebiegającego impulsu
nerwowego jest uwalniany bezpośrednio
do płynu zewnątrzkomórkowego skąd
dyfunduje do komórek. Taki sposób
unerwienia nazywamy „SYNAPSĄ NA
PRZEBIEGU”.
PODZIAŁ MIĘŚNI GŁADKICH:
MIĘŚNIE GŁADKIE
JEDNOSTKOWE (TRZEWNE, MIOGENNE)
WIELOJEDNOSTKOWE (NEUROGENNE)
POŚREDNIE

Podział mięśni gładkich
Mięśnie trzewne
: ściany przewodu
pokarmowego, moczowody, pęcherz
moczowy, macica
Mięśnie wielojednostowe
: ściany naczyń
krwionośnych, tęczówka
Mięśnie pośrednie
: mięśnie oskrzeli, mięśnie
tętniczek doprowadzających

Mięśnie jednostkowe
:
Znajdują się w przewodzie pokarmowym, układzie moczowym
oraz macicy;
Są skąpo unerwione, odległości między żylakowatościami a
komórkami mięśniowymi są duże, uwolniony neurotransmiter po
wywołaniu reakcji łatwo ulega rozproszeniu;
Mięśnie te łatwo reagują na czynniki humoralne;
Występują w nich komórki rozrusznikowe o właściwościach
OSCYLATOROWYCH, dzięki którym mięśnie te cechują się
automatyzmem;
Pomiędzy komórkami występują liczne połączenia
NISKOOPOROWE, umożliwiające przechodzenie pobudzenia z
komórki na komórkę;
Przewodzenie rozchodzi się z DEKREMENTEM (częściową
utrata energii bodźca), ma ono charakter samo ograniczający się.

Mięśnie wielojednostkowe
:
Są bardzo obficie unerwione- z zakończeń
nerwowych uwalniane są duże ilości
transmitera, który łatwo ulega wychwytowi
zwrotnemu-są całkowicie pod kontrolą układu
nerwowego;
Nie występuje przewodzenie pobudzenia z
komórki na komórkę;
Nie ma automatyzmu;
Zaliczamy do nich: mięśnie wewnętrzne gałki
ocznej, nasieniowodów.

Mięśnie pośrednie
:
Zaliczamy do nich: mięśnie oskrzeli oraz
tętniczek doprowadzających;
Na zewnątrz dominują efekty wywołane przez
neurotransmitery pochodzące z zakończeń
nerwowych, w głębszych warstwach silniejsze
efekty wywołują efekty humoralne lub
automatyzm wywołany obecnością komórek
rozrusznikowych posiadających zdolność do
samodepolaryzacji oraz połączeniami typu
NEXUS występującymi między komórkami
mięśni gładkich.

Mechanizm skurczu
mięśnia gładkiego:
Cechą czynnościową mięśni gładkich jest bardzo małe
zapotrzebowanie na energię. Może być ono ok.. 100 razy, a
nawet kilkaset razy niższe w porównaniu z mięśniami
szkieletowymi
.

PLASTYCZNOŚĆ MIĘŚNI
GŁADKICH
:
Rozciągany biernie mięsień odpowiada początkowo wzrostem,
a następnie obniżeniem napięcia. Właściwość tę nazywamy
PLASTYCZNOŚCIĄ.
W mięśniach gładkich nie ma sarkomerów, więc nie
obowiązuje prawo średnich obciążeń.
Siła skurczu mięśnia gładkiego nie zależy od wstępnego
rozciągnięcia, wynika to z prawa wielkich liczb.
Aktyna i miozyna są rozmieszczone w komórkach mięśni
gładkich w sposób mało uporządkowany, dlatego liczba
przypadkowych połączeń A-M jest mniej więcej taka sama,
niezależnie od stanu skurczu mięśnia.

Botox- zabiegi z wykorzystaniem
toksyny botulinowej (BTX)
Mechanizm działania BTX polega na
blokowaniu uwalniania acetylocholiny z
presynaptycznych zakończeń płytki
nerwowo-mięśniowe
Toksyna powoduje osłabienie skurczy
mięśniowych = likwidacja(spłycenie
zmarszczek mimicznych, kurzych łapek…

Efekty działania botoxu

Choroby mięśni - dystrofie mięśniowe
Dystrofia mięśniowa Duchenne’a
Dystrofia mięśniowa Duchenne’a (DMD) należy do grupy
chorób
dziedzicznych związanych z chromosomem X
Uważa się ją za najcięższą a równocześnie najczęściej
występującą postać kliniczną spośród postępujących
postaci dystrofii mięśniowych
Przyczyną DMD są mutacje w genie dystrofiny. W ich
konsekwencji dochodzi do zmian w ekspresji lub strukturze
tego białka w komórkach mięśniowych a następnie
upośledzenia czynności mięśni szkieletowych
Gen dystrofiny jest największym genem w komórkach
ludzkich, zajmującym prawie 0,1% genomu ludzkiego i 1%
chromosomu X
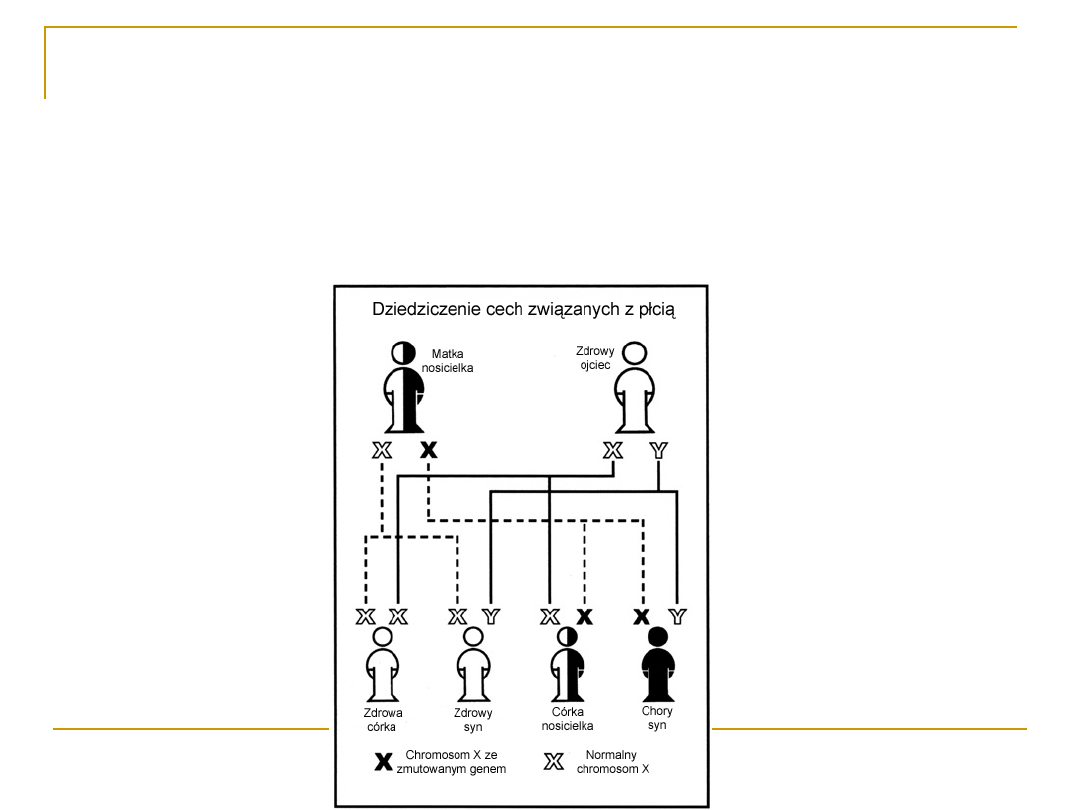
DMD-dziedziczenie

Obraz kliniczny DMD
Częstość występowania 1:
3.500 urodzeń
Pierwsze objawy choroby 3-4
rok życia
(trudności ze wstawaniem,
chodzeniem po schodach,
przerost łydek)
Całkowity zanik mięśni kończyn
dolnych: ok. 10- 12 rok życia
Dysfunkcja mięśni
oddechowych ,mięśnia
sercowego
Brak skutecznych metod terapii
całkowicie eliminujących rozwój
choroby
Średnia długość życia: ok. 20 lat

Dieta a trening siłowy
Przeciętne zapotrzebowanie kaloryczne- 2500
kcal
Aktywność- jako czynnik mający największe
znaczenie w wykorzystywaniu substancji
zapasowych i budulcowych
Pożywienie sportowców uprawiających sporty
siłowe powinno być rożne od posiłków osoby
prowadzącej siedzący tryb życia

Sport wyczynowy/właściwa
dieta
1.Uzupełnienie braku płynów- 0,5l wody / 10kg
masy ciała
2. Zwiększone zapotrzebowanie energetyczne
- łatwo wchłanialne węglowodany.
- Istotny czynnik czasu.
- W celu odnowienia glikogenu
rekompensowanie strat powstałych w wyniku
treningu powinno nastąpić bezpośrednio po nim,
nie poźniej niż w ciągu 2 godzin.

-
3
.
Białka
- główny surowiec budulcowy, (w szczególności mięśni). Dla
budowy mięśni szczególne znaczenie mają:
-
Aminokwasy rozgałęzione
− BCAA – Branched Chain Amino Acids:
leucyna, izoleucyna i walina stanowią ok. 25 % mięśni. ( właściwości
anaboliczne )
-
Glutmina
- jeden z podstawowychskładnikow budulcowych mięsni
szkieletowych.
- Karnityna
- zwiększanie transportu kwasów tłuszczowych do
mitochondriow-cykl reakcji energetycznych.
- Norma dietetyczna wynosi
1 gram białka
na kilogram masy
W treningu siłowym zapotrzebowanie na białko do
4 g
na kilogram
masy ciała, na dobę

Zawartość białka w
produktach spożywczych

Suplementacja diety w treningu
siłowym
Co daje sportowcom suplementacja? Czy
jestniezbędna? Jakie niesie zagrożenia?
Węglowodany
– łatwoprzyswajalne,cukry proste i dwucukry.
- glukoza, maltoza, laktoza i fruktoza.
Białko
-optymalny skład aminokwasów,
- aminokwasy rozgałęzione. Białko pochodzące z
mleka, serów i
jaj
. Ze składnikow pochodzenia roślinnego- białko soi. Większość
odżywek białkowych oparta jest na białku serwatkowym, ponieważ
ma ono największą przyswajalność.

W przypadku substancji energetycznych
najważniejsze jest ich przyswojenie w pierwszych
dwu godzinach po treningu. dostarczenie ze
zwykłymi posiłkami właściwej ilości białek,
zwłaszcza tych najbardziej potrzebnych
wymagałoby zjadania olbrzymiej ilości
pożywienia.
Prawidłowo stosowana suplementacja nie jest
szkodliwa/zagrożenia

„Anaboliki”
Sterydy anaboliczne- steroidy a
działaniu przyspieszajacym rozwój
tkanek- przyspieszeie podziałów
komórkowych
Pochodne testosteronu lub 19-
nortestosteronu
Zaburzenie równowagi hormonalnej
organizmu

Działanie sterydów
anaboliczch
Efekty anaboliczne
- wzrost syntezy białek
- wzrost przyrostun kośćca
-
Zwiększona produkcja erytrocytów
Efekty różnicowania płciowego
-
ginekomastia
-
Zaburzenia popędu seksualnego
-
Przedwczesne starzenie się tkanek
-
I inne…

DZIĘKUJE ZA
UWAGĘ.
Document Outline
- Slide 1
- Slide 2
- Slide 3
- Slide 4
- Slide 5
- Slide 6
- Slide 7
- Slide 8
- Slide 9
- Slide 10
- Slide 11
- Slide 12
- Slide 13
- Slide 14
- Slide 15
- Slide 16
- Slide 17
- Slide 18
- Slide 19
- Slide 20
- Slide 21
- Slide 22
- Slide 23
- Slide 24
- Slide 25
- Slide 26
- Slide 27
- Slide 28
- Slide 29
- Slide 30
- Slide 31
- Slide 32
- Slide 33
- Slide 34
- Slide 35
- Slide 36
- Slide 37
- Slide 38
- Slide 39
- Slide 40
- Slide 41
- Slide 42
- Slide 43
- Slide 44
- Slide 45
- Slide 46
- Slide 47
- Slide 48
- Slide 49
- Slide 50
- Slide 51
- Slide 52
- Slide 53
- Slide 54
- Slide 55
- Slide 56
- Slide 57
- Slide 58
- Slide 59
- Slide 60
- Slide 61
- Slide 62
- Slide 63
- Slide 64
- Slide 65
- Slide 66
- Slide 67
Wyszukiwarka
Podobne podstrony:
Fizjologia, fizjol cw, Budowa mięśnia poprzecznie prążkowanego
Prezętacja budowa mięsni
Budowa mięśnia poprzecznie prążkowanego, weterynaria, Anatomia
budowa miesnia pp
budowa mięśni i ich funkcjonowanie, fizjologia czasu i wypoczynku
Segmentowa budowa miesni
Prezętacja budowa mięsni
budowa mięśni agnieszka antoniewicz gr 20
budowa kości, mięśni, więzadeł
anatomia2, BUDOWA MIKROSKOPOWA MIĘŚNIA
biologiczne podstawy, uklad miesniowy, Budowa i rozw?j aparatu ruchu
więcej podobnych podstron