
Transport asymilatów

1. Transport substancji
organicznych w komórce
i transport
bliskodystansowy
Transport produktów fotosyntezy w roślinie składa
się z trzech współzależnych od siebie etapów:
- załadunku floemu – eksport asymilatów z
chloroplastów do cytozolu, a następnie z komórek
miękiszowych liści do tkanek przewodzących;
- transportu na duże odległości – transport w
samym floemie;
- rozładunku floemu – przemieszczenie
transportowanych związków z floemu do komórek
takich organów, jak korzenie lub owoce –
pozafloemowy transport.

• Substancje organiczne – asymilaty są
rozprowadzane przez żywe komórki łyka.
Głównym elementem przewodzącym łyka
są rurki sitowe; komórki towarzyszące i
miękisz łyka, które współdziałają w
przewodzeniu asymilatów. Transport przez
rurki sitowe odbywa się w dwóch
kierunkach: w górę i w dół rośliny. Jest to
związane z koniecznością doprowadzenia
substancji organicznych do tych części
roślin, które nie przeprowadzają
fotosyntezy, np. merystemów, miękiszu
kory, kwiatów, owoców, korzeni.

• Sok wypełniający rurki sitowe zawiera różne
substancje organiczne i jony np. PO
4
‾ ³, K
+
, Mg
+ ²
.
Ilościowo dominującym przemieszczanym
związkiem jest sacharoza. Wśród niesionych
prądem transportującym substancji znajdują się
między innymi: rafinoza (trisacharyd), aminokwasy i
amidy (zwłaszcza półamid asparagina - będąca
transportową formą związków azotu), kwasy
organiczne, nukleotydy, ATP, ortofosforany,
rozpuszczalne w wodzie witaminy (z grupy B) oraz
substancje regulujące wzrost i rozwój roślin
(fitohormony). Charakterystycznym zjawiskiem
jest brak cukrów prostych (np. heksoz) i
makromolekuł (wyjątek stanowią cząsteczki
wirusów, które mogą być przewodzone rurami
sitowymi względnie szybko) w rurach sitowych.

Transport krótkodystansowy – ruch substancji
między sąsiadującymi z sobą komórkami, tkankami, a
także przedziałami wewnątrzkomórkowymi. Woda i
sole mineralne są transportowane cewkami i
naczyniami
w kierunku od korzenia do wierzchołka rośliny; jest to
prąd wstępujący. Prąd ten jest uwarunkowany
parciem korzeniowym, a przede wszystkim siłą
ssącą liści, dzięki której tworzy się prąd
transpiracyjny umożliwiający rozprowadzenie wody
wraz z rozpuszczonymi w niej solami mineralnymi do
wszystkich części roślin. Transport
krótkodystansowy odbywa się poprzez:
- przemieszczanie się przez plazmodesmy (wypustki
cytoplazmatyczne);
- dyfuzję - bezpośrednio przez same błony biologiczne;
- specjalne pory – akwaporyny, które występują w
błonach biologicznych i są zbudowane ze specjalnych
białek;

2. Specyfika transportu
asymilatów u drzew i
krzewów
Na wiosnę u roślin drzewiastych substancje
zapasowe z korzeni do wierzchołkowych
części rośliny są przewodzone również
tkanką naczyniową. U tych roślin szczególnie
obficie zaopatrywana w związki organiczne
jest miazga, od której zależy przyrost roślin
na grubość. Przez promienie rdzeniowe wiele
substancji przedostaje się do miękiszu
drzewnego, gdzie m.in. odkładane są w
postaci materiałów zapasowych.

3. Mechanizm transportu
dalekiego asymilatów i substancje
transportowane we floemie
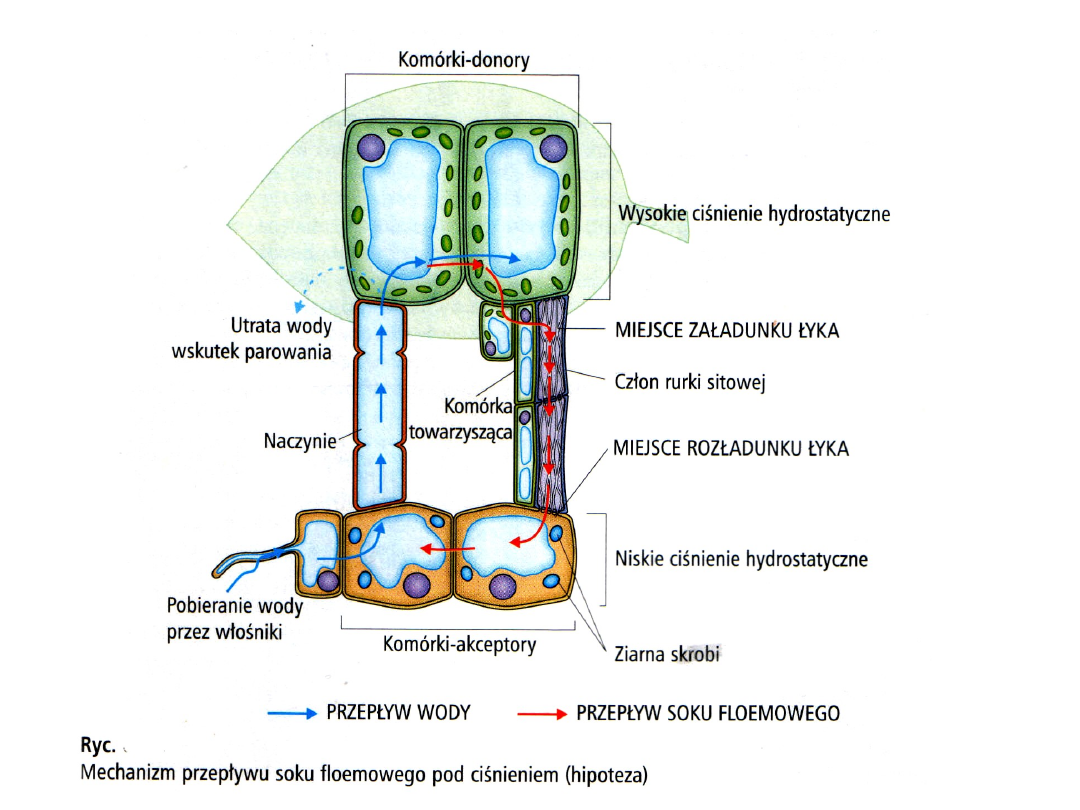
Od końca lat 70. większość badaczy powraca do koncepcji
biernego transportu floemowego, nie wymagającego
bezpośredniego wydatkowania energii do przemieszczania
substancji w rurkach sitowych. Hipotezę (przedstawioną
przez Műncha w 1930 r.) przepływu masowego
(objętościowego) pod ciśnieniem w rurkach sitowych
uznano za najlepiej wyjaśniającą większość faktów
eksperymentalnych. Opiera się ona na stwierdzeniu, że
pory w płytkach sitowych roślin okrytonasiennych są
średnio w 70% puste, czyli drożne. W tej hipotezie siłą
motoryczną ruchu w rurkach sitowych jest gradient
ciśnienia hydrostatycznego, czyli turgorowego, między
punktem załadunku i rozładunku floemu. W teorii przepływu
masowego pod ciśnieniem związki i jony przemieszczają się
w rurkach sitowych wraz z wodą, w każdej rurce tylko w
jednym kierunku. Gradient potencjału wody między rurkami
sitowymi a ksylemem powoduje stałą cyrkulację wody.

4. Od czego zależy
dystrybucja asymilatów w
roślinach?
Dystrybucja substancji pokarmowych w
roślinie zależy od:
- poziomu bezpośredniego zaopatrzenia
w produkty fotosyntezy,
- możliwości remobilizacji okresowo
zakumulowanych związków
organicznych,
- sumarycznego zapotrzebowania
wszystkich akceptorów.

• Reasumując, dystrybucja zależy od aktualnej
pojemności akumulacyjnej każdego akceptora
oraz od zużycia substratów w procesie
oddychania.
• Drugim czynnikiem rzutującym na
dystrybucję substancji pokarmowych jest
waskularny kontakt donora z akceptorem –
tzn. akceptor jest zaopatrywany w substancje
pokarmowe z najbliżej położonych liści.
• Także hormony są czynnikami
uczestniczącymi w regulacji dystrybucji
substancji pokarmowych i proces ten został
nazwany jako „transport nadzorowany przez
hormony”.

5. Rodzaje i charakterystyka
donorów i akceptorów asymilatów
• Organy, które eksportują przez całą dobę związki organiczne
(produkty fotosyntezy lub substancje okresowo akumulowane w
swoich tkankach) – to donory substancji pokarmowych.
Donorami są przede wszystkim asymilujące liście, ale również
organy spichrzowe, gdy są w nich uruchamiane rezerwy
pokarmowe, a także organy starzejące się, przede ich odcięciem
od rośliny (np. żółkniejące liście).
• Organy, których importowane są przez całą dobę substancje
pokarmowe – to akceptory związków organicznych,
produkowanych w procesie fotosyntezy. Są one również nazywane
receptorami lub biorcami. Akceptorami asymilatów są głównie
korzenie, wierzchołki wzrostu, młode, rozwijające się liście,
rozwijające się owoce i nasiona, w których występuje wysokie
zużycie substancji pokarmowych, również organy spichrzowe w
okresie gromadzenia zapasów.
• Tak więc w trakcie rozwoju organów mogą odbywać się
zmiany akceptorów w donory i odwrotnie (np. podczas
rozwoju liści czy organów spichrzowych).

Rodzaje akceptorów:
- akceptory gromadzące duże ilości substancji
pokarmowych, związane z intensywnym
wzrostem organów reprodukcyjnych;
- wegetatywne organy spichrzowe, gromadzące
skrobię (np. bulwy ziemniaka), bądź różne
cukry rozpuszczalne (np. korzeń marchewki);
- akceptory nieodwracalne – organy
odżywiające się heteroficznie (całkowicie lub
częściowo);
- patogeny (np. grzyby, owady, bakterie);
- szkodniki.

• Zapotrzebowanie akceptorów na
substancje pokarmowe zależy od
procesu wzrostu i ich pojemności
akumulacyjnej. Ilość dopływających do
nich substancji wyrażona na jednostkę
masy nazywana jest aktywnością
właściwą. Każdy akceptor jest
integralną częścią organizmu, a jego
aktywność zależy od jego zdolności od
sprawnego rozładunku floemu.

Rodzaje donorów:
- wtórne donory substancji
pokarmowych – organy, które w
okresie rozwoju wegetatywnego były
akceptorami asymilatów, w drugim
roku, w okresie poprzedzającym
przejście w fazę generatywną, stają
się wtórnymi donorami substancji
pokarmowych.

6. Załadunek i rozładunek
asymilatów
• Załadunek floemu, tj. przeniesienie
asymilatów z tkanek donora (liści) do
rury sitowej.

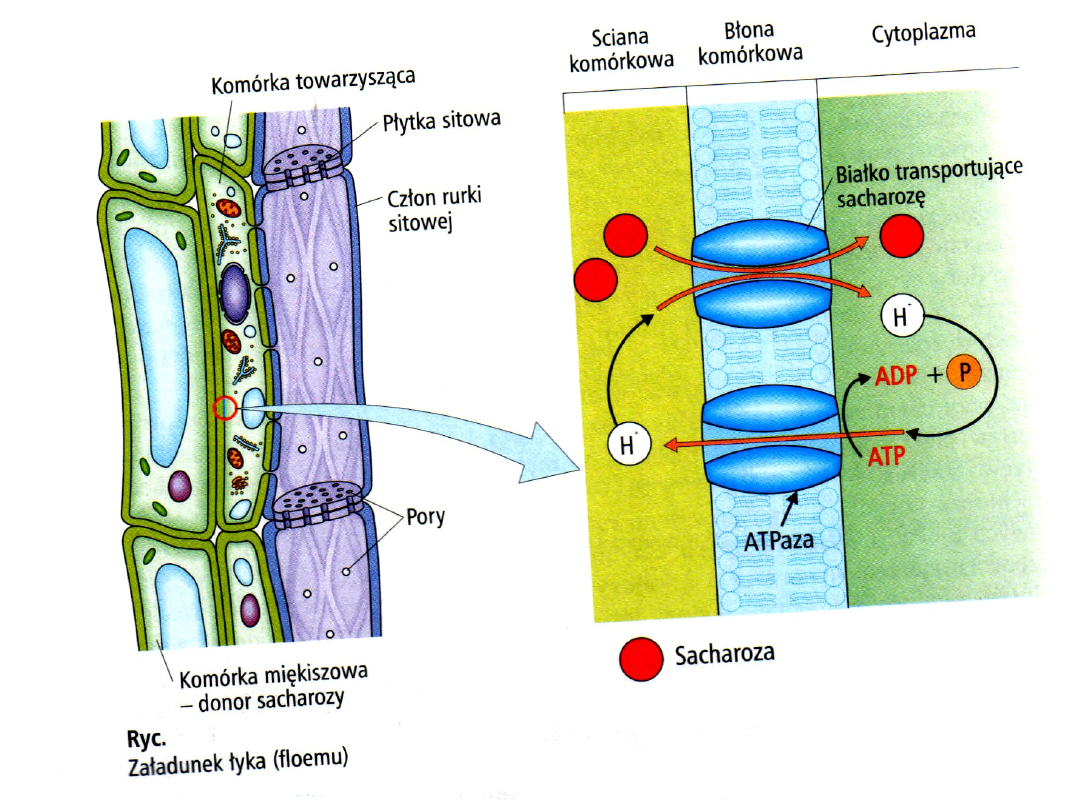
W chloroplastach miękiszu asymilacyjnego liścia
odbywa się synteza cukrów jako produktów
fotosyntezy. Pierwotne cukry to triozofosforany.
Część ich przenika do cytoplazmy, gdzie powstaje z
nich sacharoza. Część sacharozy przechodzi z
cytoplazmy do wodniczki, część do ściany
komórkowej (apoplastu). Głównie apoplastem
odbywa się przepływ roztworu sacharozy w
bezpośrednie sąsiedztwo rurki sitowej. Następuje
ładowanie rurki sitowej, często wbrew stromemu
gradientowi stężenia, co wymaga przeniesienia
cząsteczek sacharozy na drodze transportu
aktywnego. Przejściu sacharozy do rurki sitowej
towarzyszy ruch jonów H+.
W plazmolemie rurki sitowej funkcjonuje pompa
protonowa, a w rurce sitowej znajduje się dużo ATP.
Pompa ta wyrzuca protony z rurki sitowej,
powodując powstanie silnego gradientu tych jonów,
w którym zmagazynowana jest energia. Powrót
protonów do rurki sitowej, zgodnie z ich gradientem
stężenia, uwalnia energię, która umożliwia
równoczesny aktywny przepływ sacharozy.

Załadunek może się odbywać w
dwojaki sposób:
- poprzez symplastyczny kontakt –
przez plazmodesmy między
komórkami miękiszowymi a
komórkami towarzyszącymi i rurkami
sitowymi;
- na drodze apoplastycznej.

Rozładunek floemu – zjawiska podobne, choć o
kierunku odwrotnym, odbywają się również przy
rozładowaniu floemu w tkankach akceptora. Następuje
wypływ sacharozy przez błonę rurki sitowej do
apoplastu komórek akceptora. Rozładowanie floemu
jest prawdopodobnie również procesem aktywnym,
w którym uczestniczą pompy protonowe jako
pierwotne źródło energii.
Akceptory intensywnie rosnące („nieodwracalne”), np.
młode liście są zaopatrywane w substancje
pokarmowe poprzez symplast. Akceptory akumulujące
w swych tkankach substancje pokarmowe głównie
przez apoplast. W organach reprodukujących, w
rozwijających się nasionach, u których brak kontaktu
symplastycznego między komórkami zarodka a
komórkami rośliny macierzystej, z reguły rozładunek
jest typu apoplastycznego.

7. Rola substancji
wzrostowych w dystrybucji
asymilatów
Substancjami wzrostowymi nazywamy hormony, które
odgrywają trojaką rolę w dystrybucji asymilatów:
- jako regulatory indukujące ekspresję odpowiednich
genów warunkujących syntezę białek enzymatycznych
lub modyfikację aktywności enzymów uczestniczących
w transporcie na poziomie załadunku lub rozładunku
floemu;
- jako sygnały przekazujące informacje o przebiegu
procesów fizjologicznych w akceptorach, czyli
pośrednio
o wielkości zapotrzebowania na związki pokarmowe;
- jako regulatory modyfikujące intensywność wzrostu.
Ta grupa efektów dotyczy długotrwałego, lecz
pośredniego oddziaływania hormonów na transport i
dystrybucję substancji pokarmowych.
Document Outline
- Slide 1
- Slide 2
- Slide 3
- Slide 4
- Slide 5
- Slide 6
- Slide 7
- Slide 8
- Slide 9
- Slide 10
- Slide 11
- Slide 12
- Slide 13
- Slide 14
- Slide 15
- Slide 16
- Slide 17
- Slide 18
- Slide 19
- Slide 20
Wyszukiwarka
Podobne podstrony:
8 TRANSP ASYMIL id 47130 Nieznany (2)
skrócony transport asymilatów
Notatki Medycyna word grafy, TRANSPORT ASYMILATOW I SUBSTANCJI ODŻYWCZYCH U ROŚLIN
EŚT 07 Użytkowanie środków transportu
IK Transport a środowisko
Urządzenia transportu pionowego
EKONOMIKA TRANSPORTU IX
Ubezpieczenia związane z transportem drogowym
TRANSPORT GAZU prezentacja
Seminarium 6 Immunologia transplantacyjna farmacja 2
modelowanie procesˇw transportowych
8 Transport ponadnormatywny
transplantologia
Cw 7 IMMUNOLOGIA TRANSPLANTACYJNA
więcej podobnych podstron