
Układ bodźcowo-
przewodzący serca

• Odpowiedni rytm i synchronizację skurczów poszczególnych
części serca zapewnia jego układ bodźcowo-przewodzący.
• Układ ten podobnie jak układ nerwowy, przewodzi impulsy
elektryczne, jednakże w odróżnieniu od układu nerwowego
nie składa się z neuronów lecz z komórek mięśniowych
wyspecjalizowanych w przewodzeniu bodźców i odpowiednio
komunikujących się między sobą
• Cechami charakterystycznymi układu bodźcowo-
przewodzącego są jego autonomiczność i automatyzm.
• Impulsy pobudzające do skurczów są periodycznie
generowane przez wyspecjalizowane komórki mięśnia serca,
które nie są pobudzane bodźcami zewnętrznymi.
• Komórki układu przewodzącego serca komunikują się między
sobą elektrycznie za pomocą połączeń tworzonych przez
koneksony który nie powoduje on praktycznie żadnego
opóźnienia przenoszonego impulsu.
• Połączenia pomiędzy komórkami tworzone są na ich
końcach, zwykle jeden koniec komórki kontaktuje się z
dwoma innymi końcami.

• Utworzona w ten sposób sieć połączeń gwarantuje
rozchodzenie się pobudzenia we wszystkich komórkach i
zapobiega „zgubieniu” impulsu w przypadku
nieprawidłowego działania któregoś z elementów sieci.

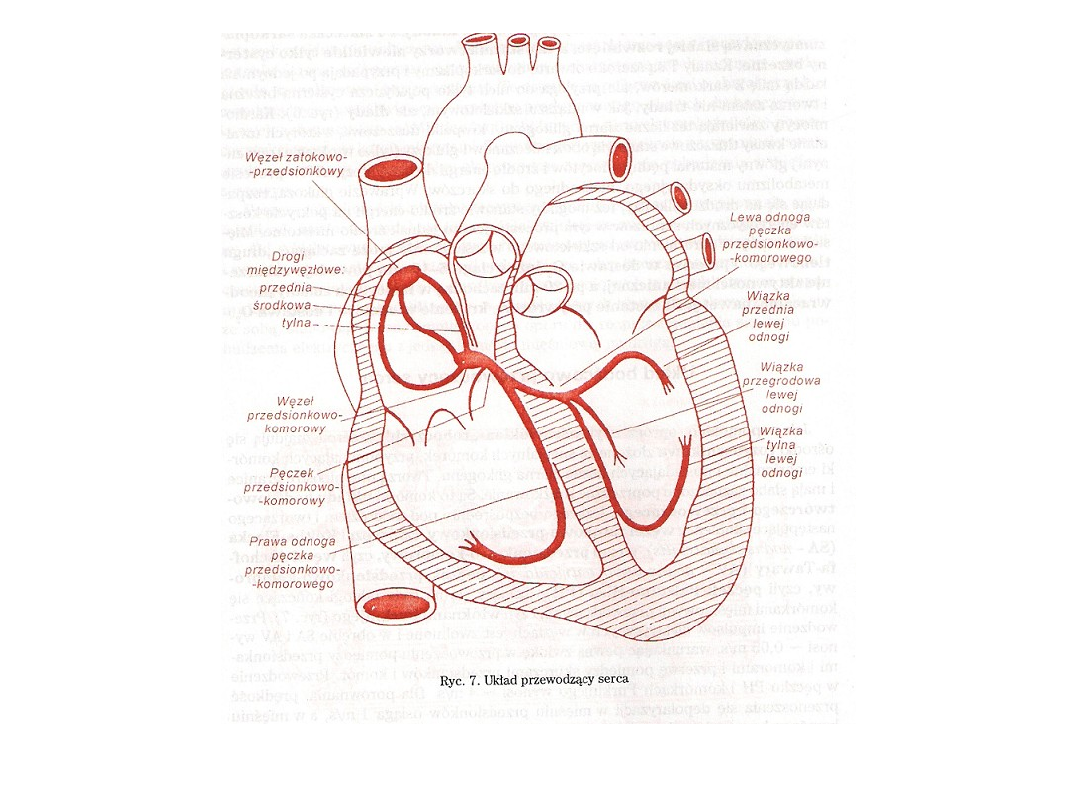
• w skład układu bodźcowo- przewodzącego serca wchodzą
grupy komórek mięśniowych nazywane węzłami i pęczkami.
• Impulsy elektryczne decydujące o częstotliwości skurczów
serca generowane są w węźle zatokowo-
przedsionkowym nazywanym również rozrusznikiem serca.
• Węzeł ten położony jest w pobliżu ujścia żyły głównej górnej
do prawego przedsionka.
• Powstające tu, z częstotliwością wynoszącą około 78
impulsów/min., potencjały czynnościowe inicjują skurcz
przedsionków i jednocześnie za pośrednictwem pęczków
międzywęzłowych docierają do węzła przedsionkowo-
komorowego położonego w dolnej części przegrody
międzyprzedsionkowej.
• Charakterystyczną cechą części komórek wchodzących w
skład węzła przedsionkowo-komorowego jest mała prędkość
rozchodzenia się w nich potencjału czynnościowego, co
wynika bezpośrednio z małej prędkości narastania potencjału
czynnościowego tych komórek.

• w świetle tego mechanizmu jest jasne, że im szybciej błona jest
lokalnie depolaryzowana, tym szybciej powstaje różnica
potencjałów elektrycznych między obszarem pobudzonym i
niepobudzonym, a więc tym szybciej potencjał czynnościowy
przemieszcza wzdłuż błony.
• Dzięki powolnemu charakterowi depolaryzacji dochodzi więc w
strefie przedsionkowo-węzłowej do opóźnienia w przekazywaniu
pobudzenia do mięśni komór o około 0,1 s.
• Komórki węzła przedsionkowo-komorowego też posiadają
zdolność do samoistnego generowania potencjałów
czynnościowych.
• Jednak dzięki wolniejszemu narastaniu generowania potencjału w
fazie powolnej spoczynkowej depolaryzacji, częstotliwość
generowanych impulsów jest mniejsza niż w węźle zatokowo-
przedsionkowym (50/min)
• Zanim więc potencjał czynnościowy zostanie wygenerowany w
węźle przedsionkowo-komorowym jest on pobudzany impulsem
dochodzącym do niego z wyższego piętra układu przewodzącego,
to znaczy z węzła zatokowo-przedsionkowym.

• Rozprzestrzenienie się pobudzenia w mięśniach komór
zapewnia pęczek przedsionkowo-komorowy (Hisa), jego
odnogi i włókna Purkiniego.
• Pęczek Hisa wychodzi z węzła przedsionkowo-komorowego
w postaci jednego pnia i następnie rozdziela się na dwie
odnogi : lewą i prawą.
• Odnogi biegną w kierunku koniuszka serca i w jego pobliżu
zakręcają się w stronie podstawy serca.
• W miarę oddalania się od pnia odnogi rozdzielają się coraz
bardziej, by w końcu stać sie pojedynczymi włóknami
rozchodzącymi się w mięśniu komór.
• Pęczek przedsionkowo-komorowy zbudowany jest z
komórek przewodzących nazywanych włóknami Purkiniego.
• Pęczek Hisa posiada zdolność do samoistnego generowania
potencjałów czynnościowych, jednakże ich częstotliwość
jest jeszcze mniejsza od częstotliwości węzła
przedsionkowo-komorowego (30/min)

• wynika stąd, że pęczek Hisa jest w
stanie przyjąc funkcję rozrusznika
serca, lecz jedynie w przypadku
uszkodzenia wyższych pięter układu
przewodzącego, gdyż w przeciwnym
razie zostanie on pobudzony przez
węzeł przedsionkowo-komorowy.

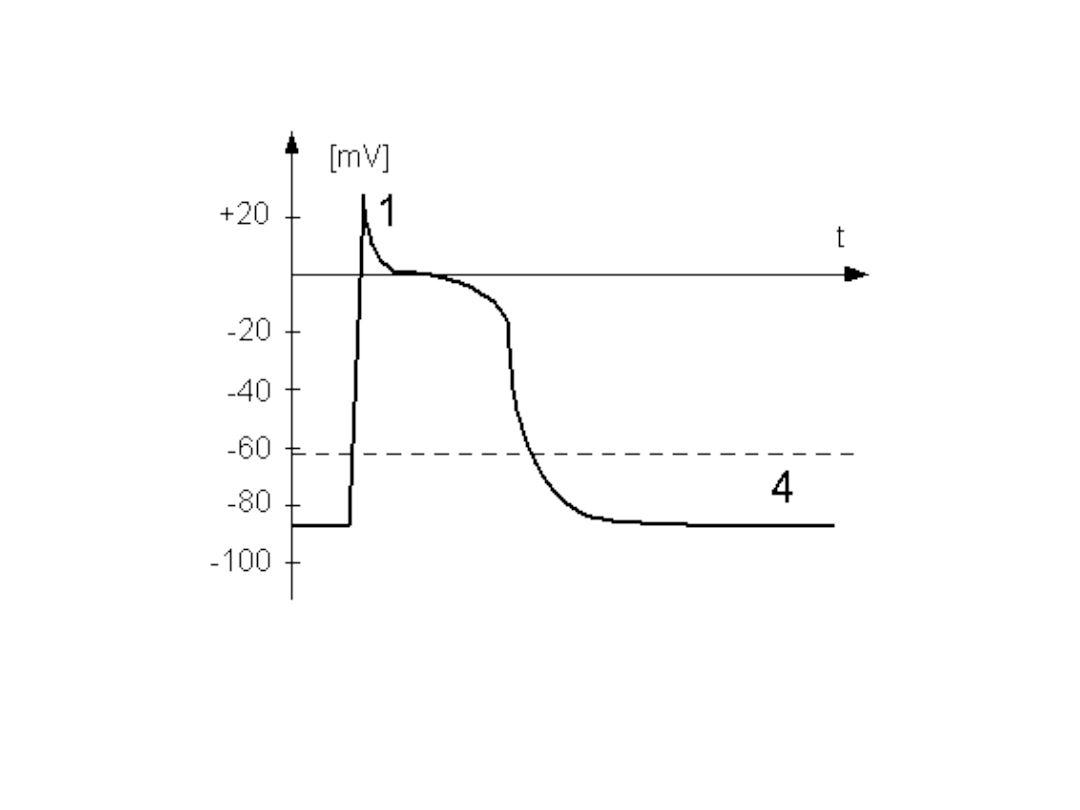
•Potencjał
czynnościowy
mięśniowy komórki
roboczej serca

• Potencjał czynnościowy mięśniowy
komórki roboczej serca różni się
istotnie od potencjału
czynnościowego neuronu lub komórki
mięśnia szkieletowego. W jego
przebiegu wyróżnia sie kilka faz które
związane są z działaniem wielu
typów kanałów jonowych.

Faza 0
• Po pobudzeniu i przekroczeniu przez
potencjał błony wartości progowej;
następuje aktywacja napięciowo-
zależnych kanałów sodowych.
Napływ jonów sodu do wnętrza
komórki powoduje gwałtowną
depolaryzację błony do wartości
+25mV. W związku ze zmianą
potencjału błonowego do wnętrza
komórki zaczyna płynąć prąd
Chlorkowy.

Faza 1
• Prąd chlorkowy oraz zmniejszenie prądu
sodowego są przyczyną spadku potencjału
błonowego.
• Zbytniemu spadkowi potencjału
przeciwdziała dokomórkowy prąd
wapniowy, który zaczyna płynąć przez
aktywowane napięciem kanały wapniowe
typu L. Kanały te otwierają się gdy
potencjał błonowy jest większy od około
-35mV.

Faza 2
• Depolaryzujący prąd wapniowy
równoważy praktycznie
repolaryzujące prądy: chlorkowy oraz
potasowy.
• Dzięki równoważeniu się tych prądów
w tej fazie przez czas około 150ms
potencjał błonowy pozostaje na
stałym poziomie (faza plateau).
• Pod koniec tej fazy inaktywacji
zaczynają ulegać kanały wapniowe.

Faza 3
• Rosnąca przewaga prądów
repolaryzujących powoduje
repolaryzację błony.
• W miarę zbliżania się potencjału
błony do wartości spoczynkowej
maleją prądy: potasowy przewodzony
przez opóżnione kanaly prostownicze
oraz chlorkowy.

Faza 4
• Osiągnięcie przez błonę potencjału
spoczynkowego.
0
2
3
Document Outline
- Slide 1
- Slide 2
- Slide 3
- Slide 4
- Slide 5
- Slide 6
- Slide 7
- Slide 8
- Slide 9
- Slide 10
- Faza 0
- Faza 1
- Faza 2
- Faza 3
- Faza 4
- Slide 16
Wyszukiwarka
Podobne podstrony:
seminarium 36 podpunkt 4wer1 ppt
seminarium 36 podpunkt 5wer1 ppt
seminarium 36 podpunkt 3wer2 ppt
seminarium 36 podpunkt 2wer1 ppt
seminarium 36 podpunkt 1glikozydy ppt
seminarium 36 podpunkt 1wer2bezglikozydow ppt
seminarium 36 podpunkt 1wer1
1 Fizjologia gospodarki wapniowo fosforanowej seminarium dla studentówid 9243 ppt
V rok seminariumt ppt
seminarium7 ppt
Środowiskowo a nowotwory seminarium1 ppt
plik (36) ppt
36 NAUKA I NAUCZYCIELE W SEMINARIUM
seminarium badanie psychiatryczne ppt
01; 14 02 2012; ROK IV; Seminarium nr 1; Dziecko świszczące 2004rid 2757 ppt
2012 10 03 Wprow Seminarium MPid 28102 ppt
V rok seminariumt ppt
seminarium7 ppt
więcej podobnych podstron