
In Re view
In Vivo Magnetic Resonance Spectroscopy and Its
Application to Neuropsychiatric Disorders
Jef frey A Stan ley, PhD
1
Key Words: in vivo, pro ton, phos pho rus, spec tros copy, schizo phre nia, bi po lar, de pres sion,
autism.
O
ur un der stand ing of bio chemi cal and mo lecu lar un der -
pin nings lead ing to neu rop sy chi at ric dis or ders is con -
tinu ally grow ing, ow ing in part to con tri bu tions from in vivo
mag netic reso nance spec tros copy (MRS) to the psy chi at ric
field. In vivo spec tros copy is the only non in va sive tech nique
that can di rectly as sess the liv ing bio chem is try in lo cal ized
brain re gions (1). In ad di tion, re cent ad vance ments in mag -
netic reso nance (MR) hard ware and soft ware tech nolo gies
have greatly im proved the qual ity of spec tros copy data, es pe -
cially the spa tial and bio chemi cal reso lu tion, and the ac cu racy
and pre ci sion of quan ti fy ing the bio chem is try. The suc cess of
in vivo spec tros copy over the past dec ade has led to use ful
find ings con cern ing the pa tho physi ol ogy of dif fer ent neu rop -
sy chi at ric dis or ders. The pur pose of this re view is first to pro -
vide a brief de scrip tion of spec tros copy and what it can do,
fol lowed by a lit era ture re view of key find ings of in vivo spec -
tros copy stud ies in schizo phre nia, af fec tive dis or ders, and
autism. It will con clude with a dis cus sion of pos si ble fu ture di -
rec tions for in vivo spec tros copy in neu rop sy chi at ric dis or -
ders. (For a list of ab bre via tions and ac ro nyms used in this
pa per, see page 318)
What is Magnetic Resonance Spectroscopy?
Both MRS and mag netic reso nance im ag ing (MRI) tech nolo -
gies are gov erned by the prin ci ples of nu clear mag netic
W Can J Psy chia try, Vol 47, No 4, May 2002
315
In vivo mag netic reso nance spec tros copy (MRS) is the only non in va sive im ag ing tech nique
that can di rectly as sess the liv ing bio chem is try in lo cal ized brain re gions. In the past dec -
ade, spec tros copy stud ies have shown bio chemi cal al tera tions in vari ous neu rop sy chi at ric
dis or ders. These first- generation stud ies have, in most cases, been ex plora tory but have
pro vided in sight ful bio chemi cal in for ma tion that has fur thered our un der stand ing of dif fer -
ent brain dis or ders. This re view pro vides a brief de scrip tion of spec tros copy, fol lowed by a
lit era ture re view of key spec tros copy find ings in schizo phre nia, af fec tive dis or ders, and
autism.
In schizo phre nia, phos pho rus spec tros copy stud ies have shown al tered me tabo lism of
mem brane phos pholip ids (MPL) dur ing the early course of the ill ness, which is con sis tent
with a neu ro de vel op men tal ab nor mal ity around the criti cal pe riod of ado les cence when the
ill ness typi cally be gins. Chil dren and ado les cents who are at in creased ge netic risk for
schizo phre nia show simi lar MPL al tera tions, sug gest ing that schizo phre nia sub jects with a
ge netic pre dis po si tion may have a pre mor bid neu ro de vel op men tal ab nor mal ity.
In de pendent of medi ca tion status, bi po lar sub jects in the de pres sive state tended to have
higher MPL pre cur sor lev els and a defi cit of high- energy phos phate me tabo lites, which
also is con sis tent with ma jor de pres sion, though these re sults var ied. Fur ther bi po lar stud ies
are needed to in ves ti gate al tera tions at the early stage.
Lastly, as so cia tions be tween pre fron tal me tabo lism of high- energy phos phate and MPL and
neu rop sy cho logi cal per form ance and re duced N- acetylaspartate in the tem po ral and cere -
bel lum re gions have been re ported in in di vidu als with autism. These find ings are con sis tent
with de vel op men tal al tera tions in the tem po ral lobe and in the cere bel lum of per sons with
autism. This pa per dis cusses re cent find ings of new func tions of N- acetylaspartate.
(Can J Psy chia try 2002;47:315–326)

reso nance (NMR). While the more popu lar MRI tech nique
pro vides cross- sectional anat omic im ages based on the tis sue
wa ter con tent, MRS is a tech nique that can meas ure the in vivo
bio chemi cal or me tabo lite con cen tra tion lev els in the hu man
body from a spe cific lo cal ized re gion. As im plied in the name,
MRS re quires a mag netic field and a radio- frequency (RF)
trans mit pulse that is at a par ticu lar reso nant fre quency to ob -
serve the sig nal of spe cific nu clei (for ex am ple, pro ton [
1
H] or
phos pho rus [
31
P]) in the sam ple of in ter est. The end prod uct of
MRS is a “spec trum” with a fre quency axis in parts per mil lion
(ppm) and a sig nal am pli tude axis. Spe cific nu clei con tained
in a me tabo lite give rise to ei ther a sin gle peak or mul ti ple
peaks that are uniquely po si tioned along the fre quency axis,
and the peak po si tion is known as the chemi cal shift; the sig nal
am pli tude of a peak, which is quan ti fied, is di rectly re lated to
the con cen tra tion of that as signed me tabo lite. De tails on the
MR ba sic prin ci ples and ap pli ca tions are de scribed else where
(2–4).
How is Magnetic Resonance Spectroscopy
Applied?
To a cer tain de gree, MRS is a com plex tech nique, but this is
due in part to its ver sa til ity in ap pli ca tion. One has the choice
of se lect ing 1) a par ticu lar nu cleus of in ter est, 2) the mag netic
field strength of the MR sys tem to con duct the ex peri ments,
and 3) the trans mit and re ceive coil con figu ra tion and pulse
se quence for lo cal iza tion (1).
Nu clei of in ter est
The cho sen nu clei will de ter mine what bio chemi cal in for ma -
tion can be as sessed and the spa tial reso lu tion ca pa bil ity as
part of the lo cal iza tion. The MR sig nal sen si tiv ity of the more
popu lar
1
H spec tros copy is about 15 times greater than that of
31
P spec tros copy. As a re sult, the spe cial reso lu tion of
1
H spec -
tros copy at 1.5 tesla, for ex am ple, tends to be within the 1- to-8
cm
3
range, while typi cal
31
P spec tros copy vox els are be tween
27 and 60 cm
3
. By choos ing
1
H spec tros copy, one can as sess
the vi abil ity of neu rons (5,6), glutamate- glutamine neu ro -
trans mis sion cy cling (7,8), the
γ
-am inob utyric acid (GABA)
neu ronal sys tem, and the sec ond mes sen ger me tabo lism by
meas ur ing, re spec tively, the me tabo lite lev els of N-
acetylaspartate (NAA), glu ta mate, glu ta mine, GABA, and
myo-inos itol (Fig ure 1a) (9,10). The
1
H spec tral peaks, phos -
pho cre atine (PCr) and cre atine (Cr), are in dis tin guish able, as
in di cated in Fig ure 1, and PCr and Cr are re ac tants in the cre -
atine ki nase high- energy phos phate re ac tion. There fore,
1
H
spec tros copy is not the pre ferred choice for as sess ing the
high- energy phos phate me tabo lism un less the equi lib rium is
al tered.
On the other hand,
31
P spec tros copy can meas ure the me tabo -
lite lev els of adeno sine tri phos phate (ATP), PCr, and
in or ganic or tho phos phate (Pi), which are as so ci ated with
high- en ergy phos phate me tabo lism (Fig ure 1b) (11,12). One
would ex pect de creased PCr with in creased high- energy utili -
za tion or a defi cit in PCr pro duc tion. Fur ther, in
31
P spec tros -
copy, mem brane phos pholipid (MPL) syn the sis and
mem brane deg ra da tion can be as sessed by meas ur ing the
freely mo bile, water- soluble phos pho monoes ters (termed
“free- PME,” and in clud ing pri mar ily phos pho cho line [PC]
and phos pho ry le tha no la mine [PE]) and phos phodi es ters
(termed “free- PDE,” and in clud ing glyc er ol phos pho cho line
[GPC] and glyc ero le tha no la mine [GPE]), re spec tively
(13,14) (Fig ure 1). In a rat model study of neu ronal de gen era -
tion and re gen era tion (us ing neo na tal le sions in the en tor hi nal
cor tex), higher lev els of free- PMEs were ob served at the time
and at the site of neu ritic sprout ing, sug gest ing that the free-
PME lev els di rectly re flect mem brane syn the sis (15).
1
H spec -
tros copy can also as sess lim ited in for ma tion on MPL me tabo -
lism.The tri meth yl amine
1
H peak or the choline- containing
peak is pri mar ily com posed of GPC (the break down prod uct
of MPL) and PC (the MPL pre cur sor). The con tri bu tion of
cho line (Cho) is be low the de tec tion limit (16); there fore, in
this re view, the choline- containing peak is termed
“GPC+PC.” Stan ley, Pet te grew, and Ke sha van pro vide a
more de tailed re view of the
1
H and
31
P me tabo lites (1).
Mag netic Field Strength
The signal- to- noise ra tio (per ac qui si tion time) is criti cal for
ac cu rate and re li able quan ti fi ca tion. In gen eral, in creas ing the
mag netic field strength leads to an ap proxi mate lin ear in -
crease in the MR sig nal am pli tude; con se quently, con duct ing
the spec tros copy at a rela tively higher mag netic field strength,
such as 3, 4, or even 7 tesla, leads to smaller lo cal ized voxel
sizes (that is, greater spa tial reso lu tion), which mini mizes the
de gree of par tial vol ume of dif fer ent tis sue types within the lo -
cal ized voxel. Moreo ver, at higher field strengths, the de gree
to which dif fer ent peaks over lap one an other is much less (or
the chemi cal shift dis per sion is greater), thus im prov ing the
ac cu racy and pre ci sion of quan ti fy ing these over lap ping
peaks, in clud ing glu ta mate and glu ta mine (17–21).
Lo cal iza tion Method
Meth ods avail able for lo cal iza tion dif fer, rang ing from us ing
a single- loop trans mit or re ceive coil with a sin gle RF pulse as
a se quence to us ing a dual- tuned (
1
H and
31
P) vol ume head coil
with slice- selective RF pulses and gra di ent pulses for spa tially
en cod ing the field of view. The stimu lated ac qui si tion mode
(STEAM) and the point- resolved spec tros copy (PRESS)
pulse se quences are the 2 most com monly used for lo cal iza -
tion with in vivo
1
H spec tros copy (10,21,22). Both these se -
quences ac quire the MR sig nal from the in ter sec tion of 3
or thogo nal slices or slabs and can be ap plied to lo cal ize ei ther
a sin gle voxel or, com bined with phase- encoding gra di ents, to
316
W Can J Psy chia try, Vol 47, No 4, May 2002
The Ca na dian Jour nal of Psy chia try—In Re view
lo cal ize mul ti ple vox els si mul ta ne ously in 2 or 3 di men sions.
Also, the time of echo (TE) pa rame ter of these se quences or
the time given for the MR sig nal to ex po nen tially at tenu ate
prior to ac qui si tion (4) may be long (for ex am ple, 135 or 272
ms), to ac quire only the
1
H me tabo lites with singlets (NAA,
PCr+Cr, and GPC+PC), or it may be short (35 ms or less), to
ac quire the singlets plus the mul ti plets, such as glu ta mate, glu -
t a m i n e ,
myo-inos itol, GABA, as par tate, n-
acetylaspartylglutamate (NAAG), taurine, glu cose, and
scyllo-inos itol (9,10). Choices of lo cal iza tion se quences for in
vivo
31
P spec tros copy in clude image- selected in vivo spec -
tros copy (ISIS) ap plied as a sin gle- or multiple- voxel
W Can J Psy chia try, Vol 47, No 4, May 2002
317
In Vivo Magnetic Resonance Spectroscopy and Its Application to Neuropsychiatric Disorders
Fig ure 1 a) A typical single-voxel short TE (= 20ms) in vivo
1
H spectrum of the prefrontal region acquired
on a 1.5 Tesla sys tem. b) A typical in vivo
31
P spectrum of the prefrontal region acquired using a multivoxel
tech nique. From top to bot tom, the acquired and mod elled apec trum and dif fer ence be tween the two are
dis played.

tech nique (23), spin echo (24,25), and chemi cal shift im ag ing
(CSI) se quences (26). The sur face coil pro vides the most sen -
si tiv ity, fol lowed by the Helm holtz coil and the vol ume coil.
Recent Advancements in the Interpretation of
Spectroscopy Data
The Broad Un der ly ing
31
P Peak
In ad di tion to the free- PME and free- PDE me tabo lites, a typi -
cal in vivo
31
P spec trum of the brain also can con tain a rela -
tively broad un der ly ing peak over the PDE and PME spec tral
re gion (Fig ure 1b), ow ing to larger and less mo bile mole cules
with PDE and PME moie ties, which can be quan ti fied us ing a
post- processing method (27). These mole cules, termed
“broad- PDE” (or PME [
i-
τ
c
] + PDE [
i-
τ
c
]) (27), may re flect
sig nals aris ing from small MPL struc tures, in clud ing mi -
celles, syn ap tic vesi cles, trans port and se cre tory vesi cles as -
so ci ated with the Golgi and en do plas mic re ticu lum (28–32),
and small phos pho ry lated pro teins (31,32). Syn ap tic vesi cles
are en riched in grey mat ter nerve ter mi nals, and trans port and
se cre tory vesi cles are en riched in white mat ter ax ons. As dis -
cussed later, the abil ity to quan tify the broad- PDE com po nent
is im por tant to the study of schizo phre nia.
The Role of N- acetylaspartate
Over the years, the clini cal use ful ness of
1
H spec tros copy has
grown by dem on strat ing de creased NAA in many dif fer ent
neu ro pa tholo gies, in clud ing psy chi at ric dis or ders (5,6). NAA
is syn the sized in neu ronal mi to chon dria from acetyl- CoA and
as par tate by the membrane- bound en zyme L- aspartate N-
acetyltransferase (33,34). Sev eral stud ies have con firmed the
neu ronal lo cal iza tion of NAA (35- 38), in clud ing the study by
Uren jak and col leagues (39), which dem on strated the pres -
ence of NAA in both neu rons and oli goden dro cytes in
de vel op ing brains but only in neu rons in ma ture brains. Con -
se quently, NAA is com monly con sid ered to be a pu ta tive neu -
ronal marker. There are, how ever, in con sis ten cies of NAA as
a marker of vi able neu rons. In creased NAA has been re ported
in Ca na van’s dis ease (40). In this dis ease, there is a de fi ciency
in the NAA cata bolic en zyme (as par toa cy lase), which leads to
neu ro de gen era tion of white mat ter, in clud ing de mye li na tion
(41,42). In ad di tion, de creases in NAA have been shown to be
re versi ble in neu ro logi cal dis eases in volv ing white mat ter
(43,44). Re cent re in ves ti ga tions into the lo cal iza tion of NAA
have re vealed that NAA can be ex pressed in ma ture oli goden -
dro cytes (that is, mye lin) (45), and there is evi dence of in ter -
com part men tal cy cling of NAA be tween neu rons and
oli goden dro cytes (45–48). The NAA is syn the sized in one
com part ment and cata bo lized in the other and may func tion as
a mo lecu lar wa ter pump (49). Con sid er ing these re cent find -
ings, the role or in ter pre ta tion of NAA may no longer in di cate
neu ronal vi abil ity but may re flect the for ma tion and main te -
nance of mye lin (46). To a cer tain de gree, this does com pli -
cate the in ter pre ta tion of NAA al tera tions, es pe cially in
stud ies of neu rop sy chi at ric dis or ders in chil dren and
ado les cents.
Recent Methodological Advancements
The in vivo con cen tra tion of GABA in hu man brain is less
than one- half that of glu ta mine, and the
1
H spec trum of GABA
con tains com plex mul ti ple peaks that over lap with PCr+Cr
and other mul ti ple peaks, in clud ing glu ta mate and glu ta mine
(50). Con se quently, the in vivo GABA quan ti fi ca tion has poor
re li abil ity un less a cus tom ized pulse se quence is used to iso -
late a par ticu lar peak of GABA from other over lap ping peaks
(that is, to ap ply a spectral- difference, editing- type se quence)
(51–55). This tech nique has shown en cour ag ing re sults in the
study of psy chi at ric dis or ders. For the first time, de creased in
318
W Can J Psy chia try, Vol 47, No 4, May 2002
The Ca na dian Jour nal of Psy chia try—In Re view
List of ab bre via tions and ac ro nyms
1
H
pro ton
31
P
phos pho rus
ATP
adeno sine tri phos phate
Cho
cho line
Cr
cre atine
CSI
chemi cal shift im ag ing
DLPFC dor so lateral pre fron tal cor tex
FEMN
first- episode, medication- naive
GABA
gamma- aminobutyric acid
GPC
glyc er ol phos pho cho line
GPE
glyc ero le tha no la mine
ISIS
in vivo spec tros copy
MDD
ma jor de pres sive dis or der
MPL
mem brane phos pholip ids
MR
mag netic reso nance
MRS
mag netic reso nance spec tros copy
NAA
N- acetylaspartate
NAAG
N- acetylaspartylglutamate
NMR
nu clear mag netic reso nance
PC
phos pho cho line
PCr
phos pho cre atine
PDE
phos phodi es ters
PE
phos pho ry le tha no la mine
Pi
in or ganic or tho phos phate
PME
phos pho monoes ters
ppm
parts per mil lion
PRESS point- resolved spec tros copy
RF
ra dio fre quency
STEAM stimu lated ac qui si tion mode
TE
time of echo
T
2
spin-spin relaxation tme

vivo GABA lev els in the oc cipi tal cor tex have been ob served
in medication- free sub jects with ma jor de pres sive dis or der
(MDD), com pared with con trol sub jects (56). This pro vides
evi dence that as so ci ates al tered GA BAer gic neu ro trans mis -
sion with de pres sion (57). Al though not de tailed in this re -
view, God dard and oth ers ap plied this tech nique to study
in di vidu als with a panic dis or der, and ob served de creased
GABA lev els in the oc cipi tal cor tex of un medi cated sub jects
with a panic dis or der but with out ma jor de pres sion, com pared
with matched con trol sub jects (58). This find ing is con sis tent
with stud ies show ing lower GABA lev els in ani mals with
anxiety- like be hav iours.
Fur ther, the spectral- editing tech nique can be cus tom ized to
iso late and quan tify other less promi nent me tabo lites in clud -
ing glu tathione (59), an enzyme- catalyzed an ti oxi dant whose
role is to pro tect the brain against oxi da tive stress (60). In sup -
port of an al tered an ti oxi dant de fense and in creased oxi da tive
in jury in schizo phre nia (61), in vivo glu tathione lev els were
sig nifi cantly lower in the me dial pre fron tal cor tex of schizo -
phre nia sub jects com pared with con trol sub jects (62). Do and
oth ers hy pothe size that the glu tathione defi cit in schizo phre -
nia may lead to de gen era tion of neu ronal pro cesses and loss of
con nec tivity in the pre fron tal cor tex (62).
Magnetic Resonance Spectroscopy and
Neuropsychiatric Disorders
Schizo phre nia
De spite dec ades of re search, the bio logi cal ba sis of schizo -
phre nia re mains un clear. Neu roi mag ing stud ies us ing MRI
pri mar ily have shown cor ti cal grey mat ter re duc tions in fron -
tal and tem po ral lobes, in creased ven tricu lar and sul cal cere -
bro spi nal fluid (CSF), and al tera tions in ba sal gan glia,
thala mus, and cere bel lum vol umes, sug gest ing that schizo -
phre nia is a “net work dis or der” that in volves the het ero mo dal
as so cia tion cor tex and the cor ti co tha la mo cere bel lar cir cuits
(63–65). Func tional neu roi mag ing stud ies have shown re -
duced func tion of criti cal brain struc tures such as the fron tal
cor tex, sug gest ing “hy pofron tal ity” (66). Re cent mod els sug -
gest that the ex ci ta tory neu ro trans mit ter, glu ta mate, and the
in hibi tory neu ro trans mit ter, GABA, play an im por tant role in
schizo phre nia (67–70). Pe riph eral meas ure ments of MPL
also have im pli cated neu ronal cell mem branes in schizo phre -
nia (71). Like wise, there is grow ing sup port for a neu ro de vel -
op men tal ab nor mal ity un der ly ing the neu ro pa tho physi ol ogy
lead ing to schizo phre nia (72–78). Be cause of the neu ro de vel -
op men tal fo cus of many stud ies, the dis cus sion of spec tros -
copy stud ies in schizo phre nia will be lim ited to those stud ies
look ing at the first- episode stage prior to medi ca tion and at the
pre mor bid stage.
First- Episode, Medication- Naive Schizo phre nia Stud ies
The semi nal in vivo spec tros copy study by Pet te grew and col -
leagues (78) ob served al tered MPL me tabo lism and high- en -
ergy phos phate me tabo lism in the com bined right and left
pre fron tal re gion of first- episode, medication- naive (FEMN)
schizo phre nia sub jects, com pared with con trol sub jects. This
was the first study to sup port Fein berg’s hy pothe sis of ab nor -
mal neu ro de vel op ment in schizo phre nia caused by an ex ag -
gera tion of nor mal pre ado les cent syn ap tic prun ing in the
pre fron tal re gion (75–78). The evi dence was based on the de -
vel op men tal pro file of PME and PDE lev els (that is, the nor -
mal de creas ing MPL pre cur sor lev els and in creas ing MPL
break down prod uct lev els with age are ex ag ger ated in FEMN
schizo phre nia pa tients). Re cent neu ro pa thologi cal stud ies
show ing re duced neu ropil in the dor so lateral pre fron tal cor tex
(DLPFC) in schizo phre nia is con sis tent with the over ex ag -
gera tion of syn ap tic prun ing (79). Sub se quent stud ies of
FEMN schizo phre nia pa tients dem on strated simi lar MPL al -
tera tions in the left pre fron tal (80,81) and in the right and left
tem po ral lobe (82). The lat ter sug gests that the ex ag ger ated
syn ap tic prun ing also may in volve the tem po ral lobe.
In view of re cent evi dence on the PDE peak, if the broad- PDE
com po nent is not seg re gated or quan ti fied, then the quan ti fied
PDE (and PME to a lesser de gree) may be a re flec tion of both
break down prod ucts of MPL and the larger, less- mobile mole -
cules with PDE (and PME) moie ties. Con se quently, it is un -
clear whether the in creased PDE in per sons with FEMN
schizo phre nia is due solely to in creased free- PDE lev els. In a
pre limi nary study us ing the method de scribed in (27), de -
creased free- PME lev els and in creased broad- PDE lev els
were ob served in the pre fron tal re gion of per sons with FEMN
schizo phre nia, com pared with con trol sub jects (83). In ter est -
ingly, with a
1
H de cou pling method, an in creased broad- PDE
com po nent, but not the free- PDE, also has been re ported in the
pre fron tal re gion of chronic, medi cated schizo phre nia suf fer -
ers (84). This im plies that the origi nally re ported in creased
PDE is not due to in creased break down prod ucts of MPL but
to in creased phos pho ry lated pro teins, in creased con tent of
syn ap tic and trans port vesi cles, in creased flu id ity in the MPL,
lead ing to a buildup of vesi cles, or in creased mo tion in these
larger mole cules with PME and PDE moie ties. Simi larly,
since the ob served amount of the broad- PDE com po nent de -
pends on the ac qui si tion pa rame ters, field strength, and post-
processing (27), these fac tors may ac count for the in con sis -
ten cies ob served in the PDE re sults of sev eral chronic schizo -
phre nia stud ies (85).
With evi dence point ing to an al tera tion in de vel op ment, the
tim ing of the in sult lead ing to the de vel op men tal al tera tion re -
mains con tro ver sial; it may oc cur ei ther early in life (72–74)
or late post na tally (75–77,86). De ter min ing whether simi lar
MPL al tera tions are pres ent ear lier in the dis or der (for
W Can J Psy chia try, Vol 47, No 4, May 2002
319
In Vivo Magnetic Resonance Spectroscopy and Its Application to Neuropsychiatric Disorders

ex am ple, dur ing the pre mor bid phase) is one ap proach to ad -
dress ing this is sue. A
31
P spec tros copy study re ported in -
creased PDE lev els in the com bined right- and left- frontal
re gion of 14 ado les cents at in creased ge netic risk for schizo -
phre nia (off spring of schizo phre nia suf fer ers), com pared with
con trol sub jects (87). Any sub jects with schizo phre nia symp -
toms were ex cluded, and 2 off spring sub jects had an ad just -
ment dis or der. In a simi lar study, de creased free- PME and
in creased broad- PDE were ob served in the pre fron tal re gion
of chil dren and ado les cents whose par ents had schizo phre nia,
com pared with con trol sub jects (88). In ter est ingly, the de -
creased free- PME was ab sent in 5 of the 16 at- risk off spring
sub jects who were free of psy chi at ric ill ness, which is con sis -
tent with the Klemm and oth ers study (87), sug gest ing that the
pre cur sor lev els of MPL in the pre fron tal re gion may be in di -
ca tors of early (non spe cific) psy chi at ric symp toms. In ad di -
tion, those off spring sub jects with psy chi at ric ill ness
(ex clud ing schizo phre nia) show PME defi cits and in creased
broad- PDE lev els simi lar to those found in FEMN schizo -
phre nia pa tients, sug gest ing that the defi cits may be pres ent
even ear lier than those found in the ado les cent years, at least in
sub jects with an in creased ge netic risk for schizo phre nia. This
would be con sis tent with the early de vel op men tal model of
schizo phre nia (72–74). It is pos si ble, how ever, that sub jects
who go on to de velop schizo phre nia and who do not have the
ge netic pre dis po si tion for the dis or der may dif fer in neu ro de -
vel op ment neu ro pa thol ogy.
Al though the spa tial reso lu tion is greater, the ap pli ca tion of in
vivo
1
H spec tros copy to study per sons with FEMN schizo -
phre nia has been some what dis ap point ing. Based on glu ta -
mate and glu ta mine meas ures, in vivo
1
H spec tros copy has the
po ten tial to as sess di rectly the ex ci ta tory glu ta ma ter gic sys -
tem that has been im pli cated in schizo phre nia. In per sons with
FEMN schizo phre nia, com pared with con trol sub jects, no sig -
nifi cant dif fer ences in glu ta mate have been re ported in the left
pre fron tal (89,90) or the left mid tem po ral lobe (91), us ing a
short TE
1
H spec tros copy method and a pri ori knowl edge for
quan ti fi ca tion. Of these 3 stud ies, in creased glu ta mine in the
left me dial pre fron tal was the only sig nifi cant dif fer ence in -
clud ing nega tive find ings in NAA (90). Glu ta mine may be
one of the least re li able me tabo lites to quan tify (92–94); how -
ever, glu ta mine may prove more sen si tive than would glu ta -
mate in as sess ing the glu ta ma ter gic sys tem, be cause the
meta bolic com part ment of glu ta mate is much greater than the
neu ro trans mit ter com part ment, and glu ta mine is in volved di -
rectly in the glutamate- glutamine cy cling dur ing exoci to sis
(7,8). In 1 other study, Ce cil and col leagues re ported de -
creased NAA / PCr + Cr ra tios and in creased GPC+PC / PCr +
Cr ra tios in the DLPFC and de creased NAA / PCr + Cr and
GPC+PC / PCr+ Cr ra tios in the mid- temporal lobe of FEMN
sub jects, us ing a short TE
1
H spec tros copy method (95). There
are, how ever, weak nesses in this study, in clud ing quan ti fy ing
with out the use of a pri ori knowl edge, us ing ra tios (that is, it is
un clear which is chang ing, the nu mera tor or the de nomi na tor),
and mix ing the sam pling of right and left side among the sub -
jects, which may in tro duce a pos si ble group bias.
In all, there does not ap pear to be any as so cia tion be tween
free- PME as ob served in
31
P spec tros copy and NAA. Re cent
evi dence of NAA’s in volve ment in the for ma tion and main te -
nance of mye lin in di cates that the lack of re duced NAA may
be a rea son able ob ser va tion. In
1
H spec tros copy stud ies of
chronic schizo phre nia, re duced NAA lev els are more con sis -
tently re ported (85), not just in the pre fron tal but also in other
re gions, which would be con sis tent with re cent evi dence
show ing pro gres sive struc tural defi cits in schizo phre nia af ter
ini tial on set (96,97).
Mood Disorders
Bi po lar Dis or der
Over the years, sig nifi cant ad vances have been made to fur -
ther the bio chemi cal and ana tomi cal un der stand ing of bi po lar
dis or der, in clud ing bio chemi cal mecha nisms in re sponse to
treat ment; how ever, the neu ro pa tho physi ol ogy of bi po lar dis -
or der re mains largely un known. Al though not con sis tent in all
stud ies, neu roi mag ing stud ies have re ported struc tural and
func tional brain ab nor mali ties, with greater ef fects in the right
hemi sphere in mood dis or ders, in volv ing the limbic-
thalamic- cortical cir cuit and the limbic- striatal- pallidal-
thalamic cir cuit—both cir cuits as so ci ated with mood regu la -
tion (98–103). In ad di tion, MRI im ages fre quently show dif -
fused white mat ter hyper in tense sig nals in cor ti cal and
sub cor ti cal brain re gions of uni po lar and bi po lar sub jects
(104,105). Func tional neu roi mag ing stud ies have dem on -
strated hy pofron tal ity and ab nor mali ties in blood flow and
glu cose me tabo lism in vari ous sub cor ti cal and limbic- related
re gions (106–108).
In the past dec ade, in vivo spec tros copy stud ies of bi po lar dis -
or ders have been fewer, rela tive to stud ies of schizo phre nia. In
gen eral, most bi po lar dis or der stud ies have in ves ti gated me -
tabo lite al tera tions be tween dif fer ent mood states and on
medi cated sub jects. Early
31
P spec tros copy find ings in clude
de creased PME or MPL pre cur sor lev els in the fron tal re gion
of medi cated euthymic bi po lar sub jects, com pared with con -
trol sub jects (109,110), that were rep li cated in medication-
free bi po lar sub jects (111). The lat ter study also re ported in -
creased PDE in the fron tal re gion and de creased PME in the
tem po ral re gion of euthymic bi po lar sub jects, com pared with
con trol sub jects (112). In con trast, there were no re ported dif -
fer ences in the manic state, com pared with con trol sub jects
(110,113). Us ing a 1D mul ti voxel
31
P spec tros copy tech nique,
Kato and col leagues were un able to re pro duce the de creased
320
W Can J Psy chia try, Vol 47, No 4, May 2002
The Ca na dian Jour nal of Psy chia try—In Re view

PME in euthymic sub jects, com pared with con trol sub jects;
how ever, they re ported in creased PME and de creased PCr in
the left pre fron tal of de pressed bi po lar sub jects, com pared
with con trol sub jects. Simi larly, the right PCr lev els were sig -
nifi cantly lower in the manic and euthymic sub jects (114). In -
ter est ingly, a
31
P spec tros copy study com par ing sub jects with
bi po lar I and II dis or ders with con trol sub jects showed in -
creased PME in hy po manic and de pressed bi po lar II sub jects,
de creased PCr in bi po lar II sub jects of all 3 states, and no dif -
fer ences in bi po lar I sub jects (115).
In vivo
1
H spec tros copy re sults are, for the most part, con sis -
tent with the above
31
P find ings. Evi dence of re duced PCr lev -
els is con sis tent with the in crease of all 3 main
1
H ra tios (NAA
/ PCr+Cr, GPC+PC / PCr+Cr, and
myo-inos itol / PCr+Cr) in
the ba sal gan glia of medi cated bi po lar sub jects, com pared
with con trol sub jects (116). Like wise, sup port of in creased
PME (which in cludes PC con tri bu tion) has been dem on -
strated by in creased GPC+PC / PCr+Cr ra tios in the ba sal gan -
glia (117,118) and right an te rior cin gu late (119) of bi po lar
sub jects. In ad di tion, with out the con found ing PCr+Cr de -
nomi na tor, in creased GPC+PC lev els in the ba sal gan glia of
bi po lar sub jects with de pres sion were also ob served (118).
Simi lar to the nega tive
31
P find ings in the fron tal of sub jects
with bi po lar I dis or ders, there were no sig nifi cant dif fer ences
in the
1
H me tabo lite ra tios in the right ba sal gan glia of bi po lar I
sub jects, com pared with con trol sub jects (120). Al though not
all con sis tent, the re sults of a re cent in vivo
1
H spec tros copy
study of the DLPFC in euthymic medication- free sub jects
have shown a bi lat eral de crease in NAA / PCr+Cr and NAA /
GPC+PC ra tios in bi po lar I sub jects and only left- side-
decreased NAA / PCr+Cr ra tios in bi po lar II sub jects, both
com pared with con trol sub jects (121). Fi nally, bi lat eral in -
creases in NAA and PCr+Cr have been ob served in the thala -
mus of euthymic men with fa mil iar bi po lar I dis or der
com pared with con trol sub jects (122). De icken and oth ers
have noted that the in creased NAA is con sis tent with cer tain
MRI find ings of in creased thala mus vol umes and a posi tron
emis sion to mo gra phy (PET) study show ing in creased syn ap -
tic den sity in the thala mus (122). A point to con sider is that the
1
H spec tros copy data were col lected with a long TE and are
thus more sub ject to al tera tions in the spin- spin re laxa tion
times (T2) than to true metabolite- level changes.
In sum mary, the evi dence of the above bi po lar stud ies dem on -
strates that the fron tal, tem po ral, and ba sal gan glia and the
thala mus are key re gions show ing al tera tions. The is sue of lat -
er al ity in these re gions is un clear and re quires fur ther in ves ti -
ga tion. As dem on strated in the al tered PME and GPC+PC
lev els, the me tabo lism of MPL is im pli cated. The al tera tions
ap pear to be in de pend ent of medi ca tion status (123), ill ness
du ra tion, or ven tricu lar en large ments (124,125). On the
whole, PME and GPC+PC lev els tended to be ab nor mally
higher in the de pressed state (119), and PME tended to be ab -
nor mally lower in the euthymic state, which may rep re sent a
trait ab nor mal ity (112). In de pendent of mood state, the
1
H
PCr+Cr and
31
P PCr lev els are re duced, sug gest ing a defi cit in
the pro duc tion of the high- energy PCr in bi po lar dis or der.
Lastly, there does not ap pear to be any evi dence of al tered
myo-inos itol, which is not what one would ex pect con sid er ing
the evi dence sup port ing the inositol- depletion hy pothe sis in
bi po lar dis or der (126– 128). This hy pothe sis states that lith -
ium in ter acts with the inosi tol phos phate second- messenger
sys tem by act ing as an in hibi tor of the en zyme con vert ing
inosi tol mono phos phates to
myo-inos itol. De tect ing
myo-inosi tol, per haps, re quires greater re gional speci fic ity
and greater sen si tiv ity, which can be achieved at higher mag -
netic field strengths. An area that has been ig nored is in ves ti -
gat ing me tabo lite al tera tions at the early stage of this ill ness,
in clud ing the study of chil dren and ado les cents with bi po lar
dis or der (129,130).
Ma jor De pres sion
In vivo spec tros copy stud ies in MDD are small but do show
con sis ten cies. Col lec tively, these spec tros copy stud ies have
shown al tera tions in the MLP pre cur sor lev els and in high-
energy phos phate me tabo lism in MDD, based on in creased
PME in the fron tal (131), in creased GPC+PC / NAA ra tios in
the ba sal gan glia (118), and re duced ATP in the fron tal (131)
and ba sal gan glia (132), all rela tive to con trol sub jects. The in -
creased MPL pre cur sor lev els in MDD are con sis tent with the
simi lar in crease in bi po lar sub jects with de pres sion, sug gest -
ing that there may be shared al tered mecha nisms un der ly ing
the pa tho physi ol ogy of these dis or ders. Pre limi nary evi dence
as so ci ates MPL pre cur sor lev els and the Ham il ton De pres sion
Rat ing Scale, as dem on strated by a nor mali za tion of ab nor -
mally higher MPL pre cur sor lev els and im prove ment on de -
pres sion scores in geriatric- depressed sub jects af ter a
12- week an ti de pres sant treat ment with acetyl- l- carnitine
(133). Fur ther, with a short TE single- voxel
1
H spec tros copy
tech nique with a pri ori knowl edge for quan ti fi ca tion, de -
creased glu ta mate + glu ta mine lev els and no dif fer ences in
NAA, GPC+PC, Pcr +Cr or
myo-inos itol were re ported in the
an te rior cin gu late of sub jects with MDD, com pared with con -
trol sub jects (134). A sub set of sub jects with se vere de pres -
sion showed a sig nifi cant re duc tion in glu ta mate, sug gest ing
a pos si ble role for the glu ta ma ter gic sys tem in MDD. Be cause
it is un clear whether there is an as so cia tion be tween the de -
creased glu ta mate in the an te rior cin gu late and the de creased
GABA in the oc cipi tal re gion of MDD sub jects (56), fur ther
stud ies are re quired.
Autism
Autism is char ac ter ized by spe cific im pair ments in so cial in -
ter ac tions, com mu ni ca tion, and sym bolic play and re stricted
W Can J Psy chia try, Vol 47, No 4, May 2002
321
In Vivo Magnetic Resonance Spectroscopy and Its Application to Neuropsychiatric Disorders

stereo typed pat terns of be hav iour, in ter est, and ac tiv ity (135).
Autism can be di ag nosed as early as age 18 months, and has a
very early age of on set, usu ally prior to age 3 years. Preva -
lence for per va sive de vel op men tal dis or ders (PDD) ranges
from 10 to 15 per 10 000 births. The symp to matol ogy of
autism is het ero ge ne ous and in cludes a wide range in in tel li -
gence and lan guage (136). Autism has con sis tently been hy -
pothe sized as a dis or der caused by a neu ro de vel op men tal
ab nor mal ity; how ever, the tim ing, lo ca tion, and in volved neu -
ronal sys tems caus ing autism are not fully un der stood. Neu ro -
pa thologi cal stud ies have re ported al tered de vel op ment of
neu rons in the an te rior cin gu late, the lim bic sys tem, and the
brain stem, as well as al tered Purk inje cells in the cere bel lum
(137). In creased brain size ap pears to be the most con sis tent
find ing. Al though nor mal at birth in per sons with autism, the
head size in creases above the norm by age 2 to 3 years, and it is
not un til autism in di vidu als reach young adult hood that the
ab nor mally larger head size falls within nor mal range
(137–141). Neu roi mag ing stud ies have shown in creased
whole brain, cere bral white mat ter, cor ti cal grey mat ter, and
cere bel lar white mat ter vol umes. De creased area in the hip po -
cam pus has been re ported in young chil dren with autism, but
not in older chil dren or ado les cents with autism (140,142).
There has been only 1
31
P spec tros copy study in autism. Com -
pared with age-, sex-, and IQ- matched con trol sub jects, high-
functioning autis tic ado les cents and young adult men showed
lower PCr lev els in the com bined right and left pre fron tal
(143), sug gest ing ei ther an in crease in the use of high- energy
phos phate me tabo lites or a defi cit in PCr pro duc tion. The
α
-ATP peak, which also com prises di- nucelotides and di-
phosphosugars, was sig nifi cantly lower in the sub jects with
autism. Be cause the
β
-ATP peak did not sig nifi cantly dif fer,
the de creased
α
-ATP peak im pli cates the lesser me tabo lites
(di- nucleotides and di- phosphosugars) that are as so ci ated
with lipid and pro tein gly co sy la tion and mem brane syn the sis
(143). Simi larly, poorer per form ance in neu rop sy cho logi cal
and lan guage tests was as so ci ated with lower PCr and PME
lev els, and higher PDE lev els; how ever, these as so cia tions
were only ob served in the autism sub jects.
In a 3- group com pari son of autism, men tally re tarded, and
con trol sub jects, NAA PCr+Cr, NAA / GPC+PC, or GPC+PC
/ PCr+Cr ra tios did not dif fer sig nifi cantly be tween autism
sub jects and con trol sub jects in the right pa rie tal, us ing a long
TE
1
H spec tros copy method (144). The NAA / GPC+PC ra -
tios, how ever, were lower in the men tally re tarded group,
com pared with the other 2 groups. Al though the spec tros copy
find ings were nega tive, both the autism and con trol sub jects
showed in creas ing NAA / GPC+PC ra tios with age, ex cept for
the older autism sub jects (those age 8 to 14 years), who
showed no in crease with age, sug gest ing a pos si ble lack of
de vel op ment in those sub jects. Ot suka and col leagues re -
ported lower NAA lev els in both the right me dial tem po ral
lobe and the left cere bel lum of per sons with autism (be tween
age 2 to 13 years), com pared with con trol sub jects (age 6 to 14
years), us ing a short TE
1
H spec tros copy method (145). The
de creased NAA is con sis tent with in creased cell den si ties in
both the lim bic sys tem and cere bel lar cor tex and with de -
creased Purk inje cells, again sup port ing a lack of de vel op -
ment in autism sub jects. In a small pi lot study us ing a short TE
ac qui si tion method, Chugani and col leagues re ported de -
creased NAA lev els rela tive to the wa ter sig nal in the left cere -
bel lum of autism pa tients, com pared with their sib lings, but no
dif fer ences in ei ther the right fron tal or left tem po ral lobe,
(146). Fi nally, the right and left fron tal, tem po ral and pa rie tal
lobes, the brain stem, and the cin gu late gy rus were as sessed on
per sons with autism aged 2 to 21 years and on con trol sub jects
aged 3 months to 15 years, us ing a short TE
1
H spec tros copy
method (147). Based on quan ti fy ing only the singlets, NAA
lev els rela tive to the wa ter sig nal showed re duced lev els in the
right and left tem po ral lobe of sub jects with autism, with
greater re duc tions in the younger autism sub jects on the right
side.
In sum mary, these in vivo
1
H spec tros copy stud ies in chil dren
and ado les cents with autism show rela tively con sis tent re -
sults, with re duced NAA lev els (as sum ing that the de nomi na -
tor is con stant) in the tem po ral lobe and left cere bel lum.
Con versely, the stud ies show nega tive re sults in the pre fron -
tal, pa rie tal, cin gu late gy rus, and brain stem. These re sults are
in part con sis tent with struc tural MRI stud ies; how ever, there
are sev eral is sues to con sider (142). First, the re sults can not
re flect neu ronal loss solely but may also in di cate the lack of
mye lin de vel op ment, con sid er ing that these stud ies in volve
ado les cents. This raises the ques tion of the de gree to which
white mat ter pa thol ogy is in volved in autism, and this ques -
tion re quires fur ther in ves ti ga tion. Along the same line, with
evi dence of in creased head size and in creased struc tural vol -
umes in young autism suf fer ers (aged 2 to 3 years), one might
ex pect in creased NAA to co in cide with the pos si ble early
over growth. Sec ond, the age range of these stud ies ap pears
quite large con sid er ing that nor mal me tabo lite and ana tomi cal
changes oc cur ring dur ing child hood and ado les cence do not
be have in a lin ear fash ion; there fore, hav ing age as a co vari ate
in the analy sis would not prop erly ad just for age (148–153).
Last, there are sev eral stud ies show ing sex dif fer ences with
age as the brain de vel ops, which em pha sizes the im por tance
of sex- matching with the con trol group (154,155). In all, con -
sid er ing the dy namic na ture of this dis or der, which ap pears to
de vi ate from nor mal de vel op ment, fu ture stud ies should take
full ad van tage of the abil ity to con duct lon gi tu di nal meas ure -
ments and fol low the al tera tions over time in chil dren, ado les -
cents, and young adults with autism. Such stud ies should
322
W Can J Psy chia try, Vol 47, No 4, May 2002
The Ca na dian Jour nal of Psy chia try—In Re view

pro vide greater in sights in the bio chemi cal al tera tions un der -
ly ing the neu ro de vel op men tal ab nor mal ity.
Summary and Future Directions
Tech no logi cally, the spec tros copy field has ad vanced sig nifi -
cantly in that one can take full ad van tage of 1) quan ti fi ca tion
meth ods to re lia bly quan tify
1
H me tabo lites non in ter ac tively
with com plex mul ti ple peaks, in clud ing glu ta mate and glu ta -
mine, us ing a pri ori knowl edge; 2) spec tral ed it ing tech niques
that can quan tify the less- intense me tabo lites, in clud ing
GABA, glu tathione, and taurine, which may shed light on the
pa tho physi ol ogy of neu rop sy chi at ric dis or ders; and 3) higher
mag netic field strengths (for ex am ple, 3, 4, or 7 tesla) to in -
crease signal- to- noise ra tios and dra mati cally re duce the
voxel size.
The first- generation in vivo spec tros copy stud ies have been,
for the most part, ex plora tory. As a re sult, a solid foun da tion
of evi dence for bio chemi cal al tera tions in vari ous psy chi at ric
dis or ders has been gen er ated, which can now be used to test
the next level of ques tions. Spe cifi cally, mod els based on neu -
ro de vel op ment or on glu ta ma ter gic or GA BAer gic sys tems
can be ap plied and tested us ing the cur rent un der stand ing of
the bio chemi cal in for ma tion meas ured by this non in va sive
tech nique.
First- generation stud ies also have been pri mar ily cross-
sectional and, con se quently, have not fully ex ploited the po -
ten tials of in vivo spec tros copy. The non in va sive ness of spec -
tros copy makes it ideal for con duct ing lon gi tu di nal stud ies by
re peat ing meas ure ments over time. For ex am ple, lon gi tu di nal
in vivo spec tros copy stud ies have the po ten tial to dis tin guish
be tween state or trait is sues, acute and chronic medi ca tion ef -
fects, and pro gres sive change ver sus static. These is sues di -
rectly ap ply to the study of neu rop sy chi at ric dis or ders.
Ul ti mately, the goal is to use in vivo spec tros copy for the clini -
cal di ag no sis and treat ment moni tor ing of neu rop sy chi at ric
dis or ders, which is still un der de vel op ment (156). How ever,
this mo men tum of first- generation spec tros copy stud ies,
which have pro vided a unique level of in ter pre ta tion, can only
help fa cili tate the clini cal role of spec tros copy and gen er ate
more hypothesis- driven stud ies in the next gen era tion.
Ref er ences
1.Stan ley JA, Pet te grew JW, Ke sha van MS. Mag netic reso nance spec tros copy in
schizo phre nia: Meth odo logi cal is sues and findings- part I. Biol Psy chia try
2000;48:357–68.
2.Far rar TC, Becker ED. Pulse and Fou rier trans form NMR; in tro duc tion to the ory
and meth ods. New York: Aca demic Press; 1971.
3.Fu kushima E, Roe der SBW. Ex peri men tal pulse NMR: a nuts and bolts ap proach.
Read ing (MA): Addison- Wesley Pub. Co. Ad vanced Book Pro gram; 1981.
4.Ga dian DG. NMR and its ap pli ca tions to liv ing sys tems. 2nd ed. New York: Ox -
ford Uni ver sity Press; 1995.
5.Coyle JT. The nag ging ques tion of the func tion of N- acetylaspartylglutamate.
Neu ro biol Dis 1997;4:231–8.
6.Tsai G, Coyle JT. N- acetylaspartate in neu rop sy chi at ric dis or ders. Prog Neu ro biol
1995;46:531– 40.
7.Mag is tretti PJ, Pel lerin L. Cel lu lar mecha nisms of brain en ergy me tabo lism and
their rele vance to func tional brain im ag ing. Phi los Trans R Soc Lond B Biol Sci
1999;354:1155–63.
8.Roth man DL, Sib son NR, Hy der F, Shen J, Be har KL, Shul man RG. In vivo nu -
clear mag netic reso nance spec tros copy stud ies of the re la tion ship be tween the
glu ta mate- glu ta mine neu ro trans mit ter cy cle and func tional neu ro ener get ics. Phi -
los Trans R Soc Lond B Biol Sci 1999;354:1165–77.
9.Mi chaelis T, Mer boldt KD, Bruhn H, Han icke W, Frahm J. Ab so lute con cen tra -
tions of me tabo lites in the adult hu man brain
in vivo: Quan ti fi ca tion of lo cal ized
pro ton MR spec tra. Ra di ol ogy 1993;187:219–27.
10.Frahm J, Bruhn H, Gyn gell ML, Mer boldt KD, Han icke W, Sau ter R. Lo cal ized
pro ton NMR spec tros copy in dif fer ent re gions of the hu man brain
in vivo. Re -
laxa tion times and con cen tra tions of cere bral me tabo lites. Magn Reson Med
1989;11:47–63.
11.Bess man SP, Gei ger PJ. Trans port of en ergy in mus cle: The phos pho ryl cre atine
shut tle. Sci ence 1981;211:448–52.
12.Wal limann T, Wyss M, Brdic zka D, Ni co lay K, Ep pen berger HM. In tra cel lu lar
com part men ta tion, struc ture and func tion of cre atine ki nase isoen zymes in tis -
sues with high and fluc tu at ing en ergy de mands: The ‘pho sph ocr eatine cir cuit’ for
cel lu lar en ergy ho meo sta sis. Bio chem J 1992;281:21–40.
13.Daw son RMC. En zy matic path ways of phos pholipid me tabo lism in the nerv ous
sys tem. In: Eichberg J, edi tor. Phos pholip ids in nerv ous tis sues. New York:
Wiley; 1985:45–78.
14.Pet te grew JW, Kopp SJ, Min shew NJ, Glonek T, Fe lik sik JM, Tow JP, and oth -
ers.
31
P nu clear mag netic reso nance stud ies of phos pho glyc eride me tabo lism in
de vel op ing and de gen er at ing brain: Pre limi nary ob ser va tions. J Neu ro pa thol Exp
Neu rol 1987;46:419–30.
15.Ged des JW, Pan dey GN, Kel ler JN, Pet te grew JW. Ele vated phos pho cho line and
phos pha ti dyl cho line fol low ing rat en tor hi nal cor tex le sions. Neu ro biol Ag ing
1997;18:305–8.
16.Miller BL. A re view of chemi cal is sues in
1
H NMR spec tros copy: N- acetyl- l- as -
par tate, cre atine and cho line. NMR Bio med 1991;4:47–52.
17.Heth er ing ton HP, Pan JW, Ma son GF, Pon der SL, Twieg DB, Deutsch G, and
oth ers. 2d
1
H spec tro scopic im ag ing of the hu man brain at 4.1 T. Magn Reson
Med 1994;32:530–4.
18.Prost RW, Mark L, Mewis sen M, Li SJ. De tec tion of glu ta mate/glu ta mine reso -
nances by
1
H mag netic reso nance spec tros copy at 0.5 tesla. Magn Reson Med
1997;37:615–8.
19.Bar tha R, Drost DJ, Me non RS, Wil liam son PC. Com pari son of the quan ti fi ca tion
pre ci sion of hu man short echo time (1)h spec tros copy at 1.5 and 4.0 tesla. Magn
Reson Med 2000;44:185–92.
20.Barker PB, Hearshen DO, Boska MD. Single- voxel pro ton MRS of the hu man
brain at 1.5T and 3.0T. Magn Reson Med 2001;45:765–9.
21.Tkac I, An der sen P, Adri any G, Mer kle H, Ugur bil K, Gru et ter R. In vivo
1
H
NMR spec tros copy of the hu man brain at 7 T. Magn Reson Med 2001;46:451–6.
22.Bot tom ley PA. Spa tial lo cal iza tion in NMR spec tros copy in vivo. Ann N Y Acad
Sci 1987;508:333–48.
23.Or didge RJ, Con nelly A, Loh man JB. Image- selected in vivo spec tros copy (ISIS).
A new tech nique for spa tially se lec tive NMR spec tros copy. J Magn Reson
1986;66:283–94.
24.Mer boldt KD, Chien D, Han icke W, Gyn gell ML, Bruhn H, Frahm J. Lo cal ized
31
P NMR spec tros copy of the adult hu man brain
in vivo us ing stimulated- echo
(STEAM) se quences. J Magn Reson 1990;89:343–61.
25.Lim KO, Pauly J, Webb P, Hurd R, Ma covski A. Short TE phos pho rus spec tros -
copy us ing a spin- echo pulse. Magn Reson Med 1994;32:98–103.
26.Brown TR. Prac ti cal ap pli ca tions of chemi cal shift im ag ing. NMR Bio med
1992;5:238–43.
27.Stan ley JA, Pet te grew JW. A post- processing method to seg re gate and quan tify
the broad com po nents un der ly ing the phos phodi es ter spec tral re gion of in vivo
31
P brain spec tra. Magn Reson Med 2001;45:390–6.
28.Mur phy EJ, Bates TE, Wil liams SR, Wat son T, Brin dle KM, Ra ja go palan B, and
oth ers. En do plas mic re ticu lum: the ma jor con tribu tor to the pde peak in he patic
31P- NMR spec tra at low mag netic field strengths. Bio chim Bio phys Acta
1992;1111:51–8.
29.Mur phy EJ, Ra ja go palan B, Brin dle KM, Radda GK. Phos pholipid bi layer con tri -
bu tion to
31
P NMR spec tra in vivo. Magn Reson Med 1989;12:282–9.
30.Kilby PM, Al lis JL, Radda GK. Spin- spin re laxa tion of the phos phodi es ter reso -
nance in the
31
P NMR spec trum of hu man brain. The de ter mi na tion of the con -
cen tra tions of phos phodi es ter com po nents. FEBS Lett 1990;272:163–5.
31.Kilby PM, Bo las NM, Radda GK. 31P- NMR study of brain phos pholipid struc -
tures in vivo. Bio chim Bio phys Acta 1991;1085:257–64.
32.Pet te grew JW, Pan chal ingam K, Klunk WE, McClure RJ, Muenz LR. Al tera tions
of cere bral me tabo lism in prob able Alz heimer’s dis ease: A pre limi nary study.
Neu ro biol Ag ing 1994;15:117–32.
33.Pa tel TB, Clark JB. Syn the sis of N- acetyl- l- aspartate by rat brain mi to chon dria
and its in volve ment in mi to chon drial/cy to solic car bon trans port. Bio chem J
1979;184:539–46.
34.Truck en miller ME, Nam boodiri MAA, Brown stein MJ, Neale JH. N- acetylation
of l- aspartate in the nerv ous sys tem: dif fer en tial dis tri bu tion of a spe cific en -
zyme. J Neu ro chem 1985;45:1658–62.
W Can J Psy chia try, Vol 47, No 4, May 2002
323
In Vivo Magnetic Resonance Spectroscopy and Its Application to Neuropsychiatric Disorders

35.Kol ler KJ, Zac zek R, Coyle J. N- acetyl- aspartyl- glutamate: re gional lev els in rat
brain and the ef fects of brain le sions as de ter mined by a new hplc method. J Neu -
ro chem 1984;43:1136–42.
36.Sim mons ML, Fron doza CG, Coyle JT. Im mu no cy to chemi cal lo cal iza tion of N-
acetyl- aspartate with mono clonal an ti bod ies. Neu ro sci ence 1991;45:37–45.
37.Nad ler JV, Coo per JR. N- acetyl- l- aspartic acid con tent of hu man neu ral tu mours
and bo vine pe riph eral nerv ous tis sues. J Neu ro chem 1972;19:313–9.
38.Mof fett JR, Nam boodiri MA, Can gro CB, Neale JH. Im mu no his to chemi cal lo cal -
iza tion of N- acetylaspartate in rat brain. Neu rore port 1991;2:131–4.
39.Uren jak J, Wil liams SR, Ga dian DG, No ble M. Pro ton nu clear mag netic reso -
nance spec tros copy un am bi gu ously iden ti fies dif fer ent neu ral cell types. J Neu ro -
sci 1993;13:981–9.
40.Ma ta lon R, Mi chals K, Se be sta D, Deanching M, Gashkoff P, Casa nova J. As par -
toa cy lase de fi ciency and N- acetylaspartic aciduria in pa tients with Ca na van dis -
ease. Am J Med Genet 1988;29:463–71.
41.Shaag A, Anik ster Y, Chris tensen E, Glus tein JZ, Fois A, Mi che lakakis H, and
oth ers. The mo lecu lar ba sis of ca na van (as par toa cy lase de fi ciency) dis ease in
Euro pean non- Jewish pa tients. Am J Hum Genet 1995;57:572–80.
42.Baslow MH, Resnik TR. Ca na van dis ease. Analy sis of the na ture of the meta bolic
le sions re spon si ble for de vel op ment of the ob served clini cal symp toms. J Mol
Neu ro sci 1997;9:109–25.
43.De Ste fano N, Mat thews PM, Ar nold DL. Re versi ble de creases in N-
acetylaspartate af ter acute brain in jury. Magn Reson Med 1995;34:721–7.
44.Ja gan na than NR, Tan don N, Raghu na than P, Kochu pil lai N. Re ver sal of ab nor -
mali ties of mye li na tion by thy rox ine ther apy in con geni tal hy po thy roid ism: Lo -
cal ized in vivo pro ton mag netic reso nance spec tros copy (MRS) study. Brain Res
Dev Brain Res 1998;109:179–86.
45.Bhakoo KK, Pearce D. In vi tro ex pres sion of N- acetyl as par tate by oli goden dro -
cytes: Im pli ca tions for pro ton mag netic reso nance spec tros copy sig nal in vivo. J
Neu ro chem 2000;74:254–62.
46.Chak ra borty G, Mekala P, Ya hya D, Wu G, Le deen RW. In traneu ronal N-
acetylaspartate sup plies ace tyl groups for mye lin lipid syn the sis: Evi dence for
myelin- associated as par toa cy lase. J Neu ro chem 2001;78:736–45.
47.Bhakoo KK, Craig TJ, Styles P. De vel op men tal and re gional dis tri bu tion of as -
par toa cy lase in rat brain tis sue. J Neu ro chem 2001;79:211–20.
48.Baslow MH. Func tions of N- acetyl- l- aspartate and N- acetyl- l- aspartylglutamate
in the ver te brate brain: role in glial cell- specific sig nal ing. J Neu ro chem
2000;75:453–9.
49.Baslow MH. Evi dence sup port ing a role for N- acetyl- l- aspartate as a mo lecu lar
wa ter pump in mye li nated neu rons in the cen tral nerv ous sys tem. An ana lyti cal
re view. Neu ro chem Int 2002;40:295–300.
50.Govin da raju V, Young K, Maud sley AA. Pro ton NMR chemi cal shifts and cou -
pling con stants for brain me tabo lites. NMR Bio med 2000;13:129–53.
51.Roth man DL, Pet roff OA, Be har KL, Matt son RH. Lo cal ized
1
H NMR meas ure -
ments of gamma- aminobutyric acid in hu man brain
in vivo. Proc Natl Acad Sci U
S A 1993;90:5662–6.
52.Wil man AH, Al len PS. Yield en hance ment of a double- quantum fil ter se quence
de signed for the ed ited de tec tion of gaba. J Magn Reson B 1995;109:169–74.
53.Kelt ner JR, Wald LL, Fre der ick BD, Ren shaw PF. In vivo de tec tion of gaba in
hu man brain us ing a lo cal ized double- quantum fil ter tech nique. Magn Reson
Med 1997;37:366–71.
54.Al len PS, Thomp son RB, Wil man AH. Metabolite- specific NMR spec tros copy in
vivo. NMR Bio med 1997;10:435–44.
55.Hao L, Peel ing J. Si mul ta ne ous spec tral ed it ing for gamma -am inob utyric acid
and taurine us ing dou ble quan tum co her ence trans fer. J Magn Reson
2000;143:95–100.
56.Sana cora G, Ma son GF, Roth man DL, Be har KL, Hy der F, Pet roff OA, and oth -
ers. Re duced cor ti cal gamma- aminobutyric acid lev els in de pressed pa tients de -
ter mined by pro ton mag netic reso nance spec tros copy. Arch Gen Psy chia try
1999;56:1043–7.
57.Sana cora G, Ma son GF, Krys tal JH. Im pair ment of ga baer gic trans mis sion in de -
pres sion: new in sights from neu roi mag ing stud ies. Crit Rev Neu ro biol
2000;14:23–45.
58.God dard AW, Ma son GF, Al mai A, Roth man DL, Be har KL, Pet roff OA, and
oth ers. Re duc tions in oc cipi tal cor tex gaba lev els in panic dis or der de tected with
1H- magnetic reso nance spec tros copy. Arch Gen Psy chia try 2001;58:556–61.
59.Tra besinger AH, We ber OM, Duc CO, Boesi ger P. De tec tion of glu tathione in the
hu man brain
in vivo by means of dou ble quan tum co her ence fil ter ing. Magn
Reson Med 1999;42:283-9.
60.Meis ter A, An der son ME. Glu tathione. Annu Rev Bio chem 1983;52:711–60.
61.Ma hadik SP, Muk her jee S. Free radi cal pa thol ogy and an ti oxi dant de fense in
schizo phre nia: a re view. Schizophr Res 1996;19:1–17.
62.Do KQ, Tra besinger AH, M. K-K, Lauer CJ, Dy dak U, Hell D, and oth ers. A uni -
fied hy pothe sis of schizo phre nia based on glu tathione defi cit. [ab stract]. Biol
Psy chia try 1999;45:42.
63.An dreasen NC. A uni tary model of schizo phre nia bleul er’s “frag mented phrene”
as schizen cephaly. Arch Gen Psy chia try 1999;56:781–7.
64.McCar ley RW, Wi ble CG, Fru min M, Hi ray asu Y, Le vitt JJ, Fischer IA, and oth -
ers. MRI anat omy of schizo phre nia. Biol Psy chia try 1999;45:1099–119.
65.Ross CA, Pearl son GD. Schizo phre nia, the het ero mo dal as so cia tion neo cor tex
and de vel op ment: Po ten tial for a neu ro ge netic ap proach. Trends Neu ro sci
1996;19:171–6.
66.An dreasen NC, Rezai K, Al liger R, Swayze VW, 2nd, Flaum M, Kirch ner P, and
oth ers. Hy pofron tal ity in neuroleptic- naive pa tients and in pa tients with chronic
schizo phre nia. As sess ment with xe non 133 single- photon emis sion com puted to -
mo gra phy and the tower of lon don. Arch Gen Psy chia try 1992;49:943–58.
67.Lie ber man JA, Sheit man BB, Ki non BJ. Neu ro chemi cal sen si ti za tion in the pa -
tho physi ol ogy of schizo phre nia: defi cits and dys func tion in neu ronal regu la tion
and plas tic ity. Neu rop sy cho phar ma col ogy 1997;17:205–29.
68.Far ber NB, New comer JW, Ol ney JW. The glu ta mate syn apse in neu rop sy chi at ric
dis or ders. Fo cus on schizo phre nia and alz heimer’s dis ease. Prog Brain Res
1998;116:421–37.
69.Ke sha van MS. De vel op ment, dis ease and de gen era tion in schizo phre nia: a uni -
tary pa tho physio logi cal model. J Psy chi atr Res 1999;33:513–21.
70.Lewis DA. Schizo phre nia and dis or dered neu ral cir cuitry. Schizophr Bull
1997;23:529–31.
71.Fen ton WS, Hib beln J, Knable M. Es sen tial fatty ac ids, lipid mem brane ab nor -
mali ties, and the di ag no sis and treat ment of schizo phre nia. Biol Psy chia try
2000;47:8–21.
72.Mur ray RM, Lewis SW. Is schizo phre nia a neu ro de vel op men tal dis or der? Br
Med J 1987;295:681–2.
73.We in berger Dr From neu ro pa thol ogy to neu ro de vel op ment. Lan cet
1995;346:552–7.
74.Mar enco S, We in berger DR The neu ro de vel op men tal hy pothe sis of schizo phre -
nia: fol low ing a trail of evi dence from cra dle to grave. Dev Psy cho pa thol
2000;12:501–27.
75.Fein berg I. Schizo phre nia and late matu ra tional brain changes in man. Psy cho -
phar ma col Bull 1982;18:29–31.
76.Hoff man RE, McGlashan TH. Par al lel dis trib uted proc ess ing and the emer gence
of schizo phrenic symp toms. Schizophr Bull 1993;19:119–40.
77.Ke sha van MS, An der son S, Pet te grew JW. Is schizo phre nia due to ex ces sive syn -
ap tic prun ing in the pre fron tal cor tex? J Psy chi atr Res 1994;28:239–65.
78.Pet te grew JW, Ke sha van MS, Pan chal ingam K, Strychor S, Kaplan DB, Tretta
MG, and oth ers. Al tera tions in brain high- energy phos phate and mem brane phos -
pholipid me tabo lism in first- episode, drug- naive schizo phren ics. A pi lot study of
the dor sal pre fron tal cor tex by in vivo phos pho rus 31 nu clear mag netic reso nance
spec tros copy. Arch Gen Psy chia try 1991;48:563–8.
79.Se lemon LD, Goldman- Rakic PS. The re duced neu ropil hy pothe sis: a cir cuit
based model of schizo phre nia. Biol Psy chia try 1999;45:17–25.
80.Wil liam son P, Drost D, Stan ley J, Carr T, Mor ri son S, Mer skey H. Lo cal ized
phos pho rus 31 mag netic reso nance spec tros copy in chronic schizo phrenic pa -
tients and nor mal con trols [let ter]. Arch Gen Psy chia try 1991;48:578.
81.Stan ley JA, Wil liam son PC, Drost DJ, Carr TJ, Ry lett RJ, Malla A, and oth ers.
An in vivo study of the pre fron tal cor tex of schizo phrenic pa tients at dif fer ent
stages of ill ness via phos pho rus mag netic reso nance spec tros copy. Arch Gen
Psy chia try 1995;52:399–406.
82.Fu ku zako H, Fu ku zako T, Hashiguchi T, Ko dama S, Taki gawa M, Fu ji moto T.
Changes in lev els of phos pho rus me tabo lites in tem po ral lobes of drug- naive
schizo phrenic pa tients. Am J Psy chia try 1999;156:1205–8.
83.Stan ley JA, Ke sha van MS, Pan chal ingam K, McClure RJ, Pet te grew JW. Mem -
brane phos pholipid me tabo lite al tera tions in pre fron tal and ba sal gan glia re gions
in schizo phre nia: An in vivo
31
P and
1
H MRSI study. Pro ceed ings of the 9th An -
nual meet ing of the In ter na tional So ci ety of Mag netic Reso nance in Medi cine.
2001; ISMRM: Ber keley (CA). p1016.
84.Pot warka JJ, Drost DJ, Wil liam son PC, Carr T, Ca naran G, Ry lett WJ, and oth ers.
A 1H- decoupled
31
P chemi cal shift im ag ing study of medi cated schizo phrenic
pa tients and healthy con trols. Biol Psy chia try 1999;45:687–93.
85.Ke sha van MS, Stan ley JA, Pet te grew JW. Mag netic reso nance spec tros copy in
schizo phre nia: meth odo logi cal is sues and findings- part II. Biol Psy chia try
2000;48:369–80.
86.Pet te grew JW, Ke sha van MS, Min shew NJ.
31
P nu clear mag netic reso nance spec -
tros copy: Neu ro de vel op ment and schizo phre nia. Schizophr Bull 1993;19:35–53.
87.Klemm S, Rzanny R, Rie he mann S, Volz HP, Schmidt B, Ger hard UJ, and oth ers.
Cere bral phos phate me tabo lism in first- degree rela tives of pa tients with schizo -
phre nia. Am J Psy chia try. 2001;158:958–60.
88.Stan ley JA, Pet te grew JW, Pan chal ingam K, McClure RJ, Har en ski K, Mon trose
M, and oth ers. Al tered mem brane phos pholipid me tabo lism in both off springs of
schizo phre nia par ents and in first- episode never medi cated schizo phre nia sub -
jects: An in vivo
31
P MRS study. So ci ety for Neu ro sci ence Ab stracts
2000;26:1575.
89.Stan ley JA, Wil liam son PC, Drost DJ, Ry lett RJ, Carr TJ, Malla A, and oth ers.
An in vi vo pro ton mag netic reso nance spec tros copy study of schizo phre nia pa -
tients. Schizophr Bull 1996;22:597–609.
90.Bar tha R, Wil liam son PC, Drost DJ, Malla A, Carr TJ, Cortese L, and oth ers.
Meas ure ment of glu ta mate and glu ta mine in the me dial pre fron tal cor tex of
never- treated schizo phrenic pa tients and healthy con trols by pro ton mag netic
reso nance spec tros copy. Arch Gen Psy chia try 1997;54:959–65.
91.Bar tha R, al- Semaan YM, Wil liam son PC, Drost DJ, Malla AK, Carr TJ, and oth -
ers. A short echo pro ton mag netic reso nance spec tros copy study of the left
mesial- temporal lobe in first- onset schizo phrenic pa tients. Biol Psy chia try
1999;45:1403–11.
92.Provencher SW. Es ti ma tion of me tabo lite con cen tra tions from lo cal ized in vivo
pro ton NMR spec tra. Magn Reson Med 1993;30:672–9.
324
W Can J Psy chia try, Vol 47, No 4, May 2002
The Ca na dian Jour nal of Psy chia try—In Re view

93.Stan ley JA, Drost DJ, Wil liam son PC, Thomp son RT. The use of a pri ori knowl -
edge to quan tify short echo in vivo
1
H MR spec tra. Magn Reson Med
1995;34:17–24.
94.Bar tha R, Drost DJ, Wil liam son PC. Fac tors af fect ing the quan ti fi ca tion of short
echo
in- vivo
1
H MR spec tra: Prior knowl edge, peak elimi na tion, and fil ter ing.
NMR Bio med 1999;12:205–16.
95.Ce cil KM, Lenkin ski RE, Gur RE, Gur RC. Pro ton mag netic reso nance spec tros -
copy in the fron tal and tem po ral lobes of neu ro lep tic na ive pa tients with schizo -
phre nia. Neu rop sy cho phar ma col ogy 1999;20:131–40.
96.Matha lon DH, Sul li van EV, Lim KO, Pfef fer baum A. Pro gres sive brain vol ume
changes and the clini cal course of schizo phre nia in men: A lon gi tu di nal mag netic
reso nance im ag ing study. Arch Gen Psy chia try 2001;58:148–57.
97.Thomp son PM, Vidal C, Giedd JN, Gochman P, Blu men thal J, Ni col son R, and
oth ers. Map ping ado les cent brain change re veals dy namic wave of ac cel er ated
gray mat ter loss in very early- onset schizo phre nia. Proc Natl Acad Sci U S A
2001;98:11650–5.
98.Nor ris SD, Krishnan KR, Ahearn E. Struc tural changes in the brain of pa tients
with bi po lar af fec tive dis or der by MRI: a re view of the lit era ture. Prog Neu rop -
sy cho phar ma col Biol Psy chia try 1997;21:1323–37.
99.Alt shuler LL, Bart zokis G, Grie der T, Cur ran J, Ji me nez T, Leight K, and oth ers.
An MRI study of tem po ral lobe struc tures in men with bi po lar dis or der or schizo -
phre nia. Biol Psy chia try 2000;48:147–62.
100.Dre vets WC. Func tional neu roi mag ing stud ies of de pres sion: The anat omy of mel -
an cho lia. Annu Rev Med 1998;49:341–61.
101.Li otti M, May berg HS. The role of func tional neu roi mag ing in the neu rop sy chol ogy
of de pres sion. J Clin Exp Neu rop sy chol 2001;23:121–36.
102.Stra kowski SM, Del Bello MP, Ad ler C, Ce cil DM, Sax KW. Neu roi mag ing in bi po -
lar dis or der. Bi po lar Dis ord 2000;2:148–64.
103.Soares JC, Mann JJ. The anat omy of mood dis or ders—re view of struc tural neu roi -
mag ing stud ies. Biol Psy chia try 1997;41:86–106.
104.Du pont RM, Jer ni gan TL, Gil lin JC, But ters N, Delis DC, Hes se link JR. Sub cor ti cal
sig nal hyper in ten si ties in bi po lar pa tients de tected by MRI. Psy chia try Res
1987;21:357–8.
105.Alt shuler LL, Cur ran JG, Hauser P, Mintz J, Deni coff K, Post R. T2 hyper in ten si ties
in bi po lar dis or der: Mag netic reso nance im ag ing com pari son and lit era ture meta-
analysis. Am J Psy chia try1995;152:1139–44.
106.Ket ter TA, An drea son PJ, George MS, Lee C, Gill DS, Parekh PI, and oth ers. An te -
rior para lim bic me dia tion of procaine- induced emo tional and psy cho sen sory ex pe -
ri ences. Arch Gen Psy chia try 1996;53:59–69.
107.George MS, Ket ter TA, Post RM. SPECT and PET im ag ing in mood dis or ders. J
Clin Psy chia try 1993;54 (Suppl):6–13.
108.Soares JC, Mann JJ. The func tional neu roa nat omy of mood dis or ders. J Psy chi atr
Res 1997;31:393–432.
109.Kato T, Taka hashi S, Shioiri T, Inubushi T. Brain phos pho rous me tabo lism in de -
pres sive dis or ders de tected by phosphorus- 31 mag netic reso nance spec tros copy. J
Af fect Dis ord 1992;26:223–30.
110.Kato T, Taka hashi S, Shioiri T, Inubushi T. Al tera tions in brain phos pho rous me -
tabo lism in bi po lar dis or der de tected by in vivo
31
P and 7Li mag netic reso nance
spec tros copy. J Af fect Dis ord 1993;27:53–9.
111.De icken RF, Fein G, Wei ner MW. Ab nor mal fron tal lobe phos pho rous me tabo lism
in bi po lar dis or der. Am J Psy chia try 1995;152:915–8.
112.De icken RF, Wei ner MW, Fein G. De creased tem po ral lobe phos pho monoes ters in
bi po lar dis or der. J Af fect Dis ord 1995;33:195–9.
113.Kato T, Shioiri T, Taka hashi S, Inubushi T. Meas ure ment of brain phos phoi nosi tide
me tabo lism in bi po lar pa tients us ing in vivo 31P- MRS. J Af fect Dis ord
1991;22:185–90.
114.Kato T, Shioiri T, Mu rashita J, Ha makawa H, Taka hashi Y, Inubushi T, and oth ers.
Lat er al ized ab nor mal ity of high en ergy phos phate me tabo lism in the fron tal lobes of
pa tients with bi po lar dis or der de tected by phase- en coded 31P- MRS. Psy chol Med
1995;25:557–66.
115.Kato T, Taka hashi S, Shioiri T, Mu rashita J, Ha makawa H, Inubushi T. Re duc tion of
brain phos pho cre atine in bi po lar II dis or der de tected by phosphorus- 31 mag netic
reso nance spec tros copy. J Af fect Dis ord 1994;31:125–33.
116.Sharma R, Venka tasubra ma nian PN, Barany M, Davis JM. Pro ton mag netic reso -
nance spec tros copy of the brain in schizo phrenic and af fec tive pa tients. Schizophr
Res 1992;8:43–9.
117.Kato T, Ha makawa H, Shioiri T, Mu rashita J, Taka hashi Y, Taka hashi S, and oth ers.
Choline- containing com pounds de tected by pro ton mag netic reso nance spec tros -
copy in the ba sal gan glia in bi po lar dis or der. J Psy chia try Neu ro sci
1996;21:248–54.
118.Ha makawa H, Kato T, Mu rashita J, Kato N. Quan ti ta tive pro ton mag netic reso nance
spec tros copy of the ba sal gan glia in pa tients with af fec tive dis or ders. Eur Arch Psy -
chia try Clin Neu ro sci 1998;248:53–8.
119.Moore CM, Breeze JL, Gru ber SA, Babb SM, Fre der ick BB, Vil la fuerte RA, and
oth ers. Cho line,
myo-inosi tol and mood in bi po lar dis or der: A pro ton mag netic reso -
nance spec tro scopic im ag ing study of the an te rior cin gu late cor tex. Bi po lar Dis ord
2000;2:207–16.
120.Ohara K, Isoda H, Suzuki Y, Take hara Y, Ochiai M, Takeda H, and oth ers. Pro ton
mag netic reso nance spec tros copy of the len ticu lar nu clei in bi po lar i af fec tive dis or -
der. Psy chia try Res 1998;84:55–60.
121.Wins berg ME, Sachs N, Tate DL, Adal steins son E, Spiel man D, Ket ter TA. De -
creased dor so lateral pre fron tal N- acetyl as par tate in bi po lar dis or der. Biol Psy chia -
try 2000;47:475–81.
122.De icken RF, Eliaz Y, Fei well R, Schuff N. In creased tha lamic N- acetylaspartate in
male pa tients with fa mil ial bi po lar i dis or der. Psy chia try Res 2001;106:35–45.
123.Yildiz A, Sachs GS, Dorer DJ, Ren shaw PF.
31
P nu clear mag netic reso nance spec -
tros copy find ings in bi po lar ill ness: a meta- analysis. Psy chia try Res
2001;106:181–91.
124.Kato T, Shioiri T, Mu rashita J, Ha makawa H, Inubushi T, Taka hashi S.
Phosphorus- 31 mag netic reso nance spec tros copy and ven tricu lar en large ment in bi -
po lar dis or der. Psy chia try Res 1994;55:41–50.
125.Kato T, Inubushi T, Kato N. Mag netic reso nance spec tros copy in af fec tive dis or -
ders. J Neu rop sy chia try Clin Neu ro sci 1998;10:133–47.
126.Ber ridge MJ, Ir vine RF. Inosi tol phos phates and cell sig nal ling. Na ture
1989;341:197–205.
127.Ber ridge MJ, Dow nes CP, Han ley MR. Neu ral and de vel op men tal ac tions of lith -
ium: a uni fy ing hy pothe sis. Cell 1989;59:411–9.
128.Ber ridge MJ, Dow nes CP, Han ley MR. Lith ium am pli fies agonist- dependent phos -
pha ti dy li nosi tol re sponses in brain and sali vary glands. Bio chem J
1982;206:587–95.
129.Cas ti llo M, Kwock L, Cour voisie H, Hooper SR. Pro ton MR spec tros copy in chil -
dren with bi po lar af fec tive dis or der: Pre limi nary ob ser va tions. AJNR Am J Neu ro -
ra diol 2000;21:832–8.
130.Da vanzo P, Tho mas MA, Yue K, Oshiro T, Belin T, Strober M, and oth ers. De -
creased an te rior cin gu late myo- inositol/cre atine spec tros copy reso nance with lith -
ium treat ment in chil dren with bi po lar dis or der. Neu rop sy cho phar ma col ogy
2001;24:359–69.
131.Volz HP, Rzanny R, Rie he mann S, May S, He ge wald H, Pre us sler B, and oth ers.
31
P
mag netic reso nance spec tros copy in the fron tal lobe of ma jor de pressed pa tients.
Eur Arch Psy chia try Clin Neu ro sci 1998;248:289–95.
132.Moore CM, Chris tensen JD, La fer B, Fava M, Ren shaw PF. Lower lev els of nu cleo -
side tri phos phate in the ba sal gan glia of de pressed sub jects: a phosphorous- 31 mag -
netic reso nance spec tros copy study. Am J Psy chia try 1997;154:116–8.
133.Pet te grew JW, Levine J, Ger shon S, Stan ley JA, Servan- Schreiber D, Pan chal ingam
K, and oth ers. 31P- MRS study of acetyl- l- carnitine treat ment in geri at ric de pres -
sion: pre limi nary re sults. Bi po lar Dis or ders 2002; forth com ing.
134.Auer DP, Putz B, Kraft E, Lipin ski B, Schill J, Hols boer F. Re duced glu ta mate in the
an te rior cin gu late cor tex in de pres sion: An in vivo pro ton mag netic reso nance spec -
tros copy study. Biol Psy chia try 2000;47:305–13.
135.Ameri can Psy chi at ric As so cia tion. Di ag nos tic and sta tis ti cal man ual of men tal dis -
or ders.Wash ing ton (DC): Ameri can Psy chi at ric As so cia tion; 1994.
136.Rum sey JM, Ernst M. Func tional neu roi mag ing of autis tic dis or ders. Ment Re tard
Dev Dis abil Res Rev 2000;6:171–9.
137.Kem per TL, Bau man M. Neu ro pa thol ogy of in fan tile autism. J Neu ro pa thol Exp
Neu rol 1998;57:645–52.
138.Piven J, Arndt S, Bai ley J, Hav er camp S, An dreasen NC, Pal mer P. An MRI study of
brain size in autism. Am J Psy chia try 1995;152:1145–9.
139.Piven J, Nehme E, Si mon J, Barta P, Pearl son G, Fol stein SE. Mag netic reso nance
im ag ing in autism: meas ure ment of the cere bel lum, pons, and fourth ven tri cle. Biol
Psy chia try 1992;31:491–504.
140.Courchesne E, Karns CM, Davis HR, Zic cardi R, Carper RA, Tigue ZD, and oth ers.
Un usual brain growth pat terns in early life in pa tients with autis tic dis or der: an MRI
study. Neu rol ogy 2001;57:245–54.
141.Lain hart JE, Piven J, Wzorek M, Landa R, San tan gelo SL, Coon H, and oth ers. Mac -
ro cephaly in chil dren and adults with autism. J Am Acad Child Ado lesc Psy chia try
1997;36:282–90.
142.Sai toh O, Karns CM, Courchesne E. De vel op ment of the hip po cam pal for ma tion
from 2 to 42 years: MRI evi dence of smaller area den tata in autism. Brain
2001;124:1317–24.
143.Min shew NJ, Gold stein G, Dom browski SM, Pan chal ingam K, Pet te grew JW. A
pre limi nary
31
P MRS study of autism: Evi dence for un der syn the sis and in creased
deg ra da tion of brain mem branes. Biol Psy chia try 1993;33:762–73.
144.Hashi moto T, Ta yama M, Mi yazaki M, Yoneda Y, Yoshi moto T, Ha rada M, and
oth ers. Dif fer ences in brain me tabo lites be tween pa tients with autism and men tal re -
tar da tion as de tected by in vivo lo cal ized pro ton mag netic reso nance spec tros copy. J
Child Neu rol 1997;12:91-6.
145.Ot suka H, Ha rada M, Mori K, Hi saoka S, Nishi tani H. Brain me tabo lites in the
hippocampus- amygdala re gion and cere bel lum in autism: an 1H- MR spec tros copy
study. Neu ro ra di ol ogy 1999;41:517–9.
146.Chugani DC, Sun dram BS, Be hen M, Lee ML, Moore GJ. Evi dence of al tered en -
ergy me tabo lism in autis tic chil dren. Prog Neu rop sy cho phar ma col Biol Psy chia try
1999;23:635–41.
147.Hi saoka S, Ha rada M, Nishi tani H, Mori K. Re gional mag netic reso nance spec tros -
copy of the brain in autis tic in di vidu als. Neu ro ra di ol ogy 2001;43:496–8.
148.Jer ni gan TL, Zi sook S, Hea ton RK, Mo ran ville JT, Hes se link JR, Braff DL. Mag -
netic reso nance im ag ing ab nor mali ties in len ticu lar nu clei and cere bral cor tex in
schizo phre nia. Arch Gen Psy chia try 1991;48:881–90.
149.Pfef fer baum A, Matha lon DH, Sul li van EV, Raw les JM, Zipur sky RB, Lim KO. A
quan ti ta tive mag netic reso nance im ag ing study of changes in brain mor phol ogy
from in fancy to late adult hood. Arch Neu rol 1994;51:874–87.
W Can J Psy chia try, Vol 47, No 4, May 2002
325
In Vivo Magnetic Resonance Spectroscopy and Its Application to Neuropsychiatric Disorders
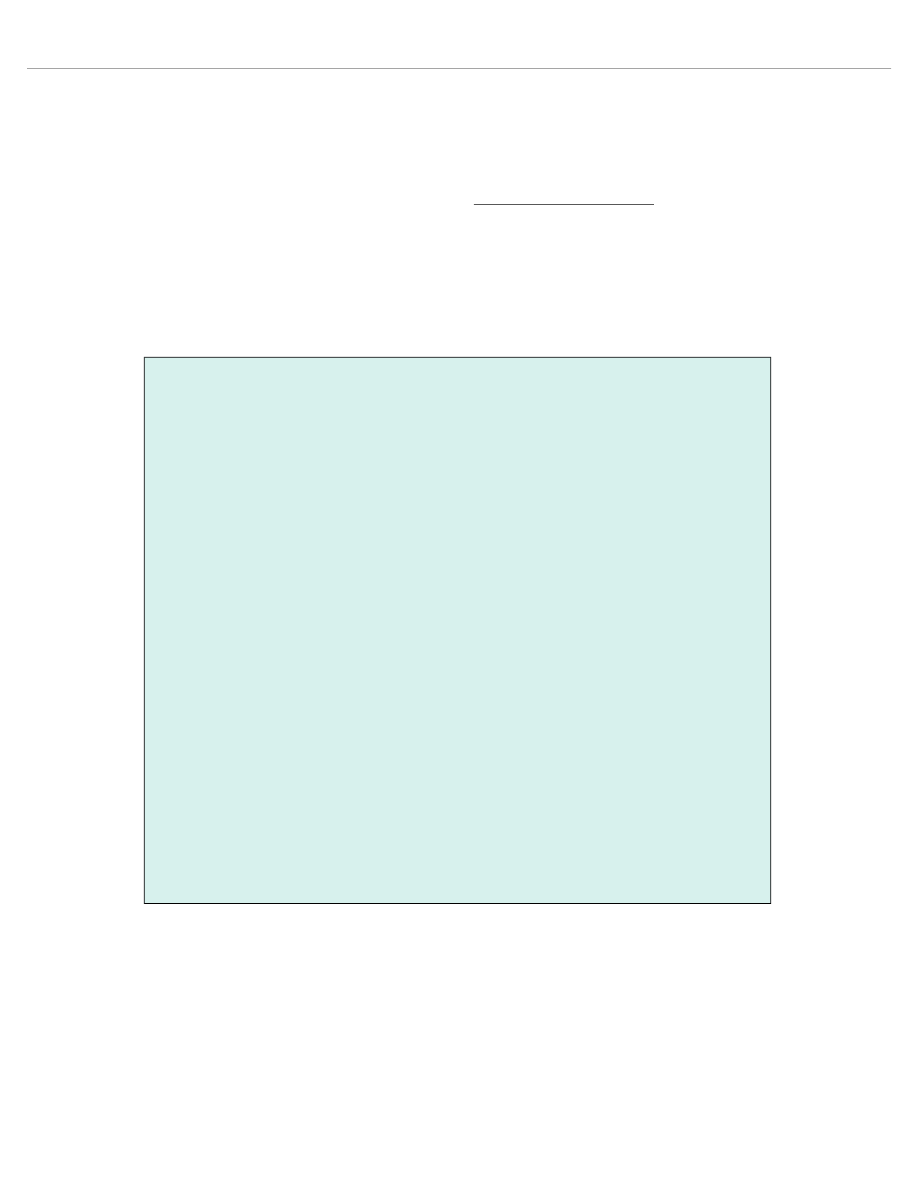
150.Cavi ness VS, Jr., Ken nedy DN, Richelme C, Rade ma cher J, Fili pek PA. The hu man
brain age 7-11 years: a volu met ric analy sis based on mag netic reso nance im ages.
Cereb Cor tex 1996;6:726–36.
151. Re iss AL, Abrams MT, Singer HS, Ross JL, Denckla MB. Brain de vel op ment, gen -
der and IQ in chil dren. A volu met ric im ag ing study. Brain 1996;119:1763–74.
152.Giedd JN, Blu men thal J, Jef fries NO, Cas tel la nos FX, Liu H, Zijden bos A, and oth -
ers. Brain de vel op ment dur ing child hood and ado les cence: a lon gi tu di nal MRI
study. Nat Neu ro sci 1999;2:861–3.
153.Stan ley JA, Min shew NJ, Ke sha van MS, Pan chal ingam K, McClure RJ, Pet te grew
JW. As sess ing the age- dependent pro file in the fron tal and cen trum semio vale re -
gions of healthy nor mal con trols us ing in vivo
31
P MRS. Pro ceed ings of the 8th An -
nual meet ing of the In ter na tional So ci ety of Mag netic Reso nance in Medi cine.
2000; ISMRM: Ber keley (CA). p 1130.
154.De Bel lis MD, Ke sha van MS, Beers SR, Hall J, Frus taci K, Masaleh dan A, and oth -
ers. Sex dif fer ences in brain matu ra tion dur ing child hood and ado les cence. Cereb
Cor tex 2001;11:552–7.
155.Rie he mann S, Volz HP, Wenda B, Hub ner G, Ross ger G, Rzanny R, and oth ers.
Fron tal lobe in vivo 31P- MRS re veals gen der dif fer ences in healthy con trols, not in
schizo phren ics. NMR Bio med 1999;12:483–9.
156.Ross B, Bluml S. Mag netic reso nance spec tros copy of the hu man brain. Anat Rec
2001;265:54–84.
Manuscript received and accepted March 2002.
1
As sis tant Pro fes sor, De part ment of Psy chia try, West ern Psy chi at ric In sti tute
and Clinic, Uni ver sity of Pitts burgh Medi cal Cen ter, Pitts burgh, Penn syl va -
nia.
Ad dress for cor re spon dence: Dr JA Stan ley, Uni ver sity of Pitts burgh, Room
721 Par ran Hall/GSPH, 130 De Soto Street, Pitts burgh, PA 15213
e- mail: stan ley@pitt.edu
326
W Can J Psy chia try, Vol 47, No 4, May 2002
The Ca na dian Jour nal of Psy chia try—In Re view
Rés umé : La spectroscopie par résonance magnétique in vivo et son application
aux troubles neuropsychiatriques
La spec tro sco pie par réso nance magné tique (SRM) in vivo est la seule tech nique non in va sive de visu ali -
sa tion qui peut éval uer di recte ment la bio chimie vi vante des ré gions du cer veau lo cal isées. Au cours des
10 der nières années, les études de spec tro sco pie ont in diqué des modi fi ca tions bio chimiques dans di vers
trou bles neu rop sy chia tri ques. Dans la plupart des cas, ces études de pre mière gé né ra tion ont été ex plora -
toires, mais elles ont fourni des ren seig ne ments bio chimiques éclairants qui ont amé lioré no tre con nais -
sance des diffé rents trou bles du cer veau. Cette étude of fre une brève de scrip tion de la spec tro sco pie,
suivie d’une ana lyse docu men taire des prin ci paux résul tats de la spec tro sco pie pour la schizo phré nie, les
trou bles af fec tifs et l’au tisme.
Dans la schizo phré nie, les études de la spec tro sco pie du phos phore ont révélé un méta bolisme modi fié de
la mem brane phos pholip ide (MPL) du rant la pre mière phase de la mala die, ce qui cor re spond à une
anoma lie neu rodévelop pe men tale aut our de la pé ri ode cri tique de l’a do les cence, où la mala die dé bute
habi tu el le ment. Les en fants et les ado les cents qui sont à risque gé né tique ac cru pour la schizo phré nie pré -
sen tent des modi fi ca tions sem bla bles de la MPL, in diquant ainsi que les su jets schizo phrènes qui ont une
prédis po si tion gé né tique peu vent avoir une anoma lie neu rodévelop pe men tale prémor bide.
Indépen dam ment du mode de médi ca tion, les su jets bi po laires en état dépres sif ten dent à avoir des ni -
veaux pré curseurs de MPL plus élevés et un défi cit des méta bo lites de phos phate à haute én er gie, ce qui
cor re spond égale ment à la dépres sion ma jeure, bien que ces résul tats vari ent. D’autres études sur le trou -
ble bi po laire sont né ces saires pour re cher cher les modi fi ca tions aux pre miers sta des.
En fin, des as so cia tions en tre le méta bolisme préfron tal du phos phate à haute én er gie et la MPL et le
ren de ment neu rop sy cholo gique ainsi qu’une réduc tion de la N- acétylaspartate dans la ré gion tem po -
rale du cer veau et dans le cer velet ont été con statées chez les per son nes at te in tes d’au tisme. Ces résul -
tats cor re spon dent aux modi fi ca tions du dévelop pe ment du lobe tem po ral et du cer velet chez les
per son nes souf frant d’au tisme. Cet ar ti cle dis cute des dé cou vertes ré cen tes des nou velles fonc tions de
la N- acétylaspartate.
Wyszukiwarka
Podobne podstrony:
In vivo MR spectroscopy in diagnosis and research of
In vivo MR spectroscopy in diagnosis and research of
Magnetic Treatment of Water and its application to agriculture
Magnetic Treatment of Water and its application to agriculture
The Danger Theory and Its Application to Artificial Immune Systems
Software Vaccine Technique and Its Application in Early Virus Finding and Tracing
MRS and its application in Alzheimer s disease
Changes in passive ankle stiffness and its effects on gait function in
The main press station is installed in the start shaft and?justed as to direction
Changes in passive ankle stiffness and its effects on gait function in
Non Intrinsic Differential Mode Noise of Switching Power Supplies and Its Implications to Filter Des
Vicci Quaternions and rotations in 3d space Algebra and its Geometric Interpretation (2001) [share
Applied Anthropology and Its Relationship to Anthropology
Petasites kablikianus Tausch ex Berchtold as a pioneer species and its abilities to colonise initial
Winch The Idea of a Social Science And its Relation to Philosophy
Brain to Serum Lithium Ratio and Age An In Vivo Magnetic Resonance Spectroscopy Study
więcej podobnych podstron