
Gene Regulation
and Cancer
C H A P T E R
12
O U T L I N E
12.1 Control of Gene Expression
• Reproductive cloning shows that any cell has the potential to become a complete organism, and therapeutic cloning suggests that
embryonic cells and adult stem cells may become any type of specialized cell.•180
–81
• In prokaryotes, gene expression is regulated at the level of transcription by a regulatory gene.•183
• In eukaryotes, gene expression is regulated at many more levels, including DNA unpacking, DNA transcription, mRNA
processing, mRNA translation, and protein activity.•183
• In eukaryotes, transcription factors turn genes on, and transcription activators enhance transcription.•186
• In eukaryotes, signaling between cells stimulates transcription activators.•187
12.2 Cancer: A Failure of Genetic Control
• Cells that actively divide are particularly subject to the development of cancer.•188
• Mutations in proto-oncogenes and tumor suppressor genes bring about cancer.•189
• When proto-oncogenes mutate and become oncogenes, and when tumor suppressor genes mutate, cells divide uncontrollably
and apoptosis does not occur.•189
• Rarely, mutated proto-oncogenes and tumor suppressor genes are inherited, which increases a persons predisposition to
cancer.•191
• Tests for genes that contribute to the development of cancer are sometimes available.•191
The term cancer describes a family of diseases having the common characteristic of uncontrolled cell division. You are probably aware
that lifestyle choices such as smoking or tanning might increase your odds of developing certain types of cancer. However, have you ever
considered that some viral infections can also increase your chances of cancer? A group of viruses termed oncogenic viruses can
contribute to the possibility of certain cancers. These viruses are characterized by their ability to insert viral DNA into human chromosomes.
Humans have several genes, called proto-oncogenes, that help regulate cell division. When oncogenic viral DNA inserts a mutated
proto-oncogene, called an oncogene, into a human chromosome, the cell cycle may occur repeatedly, and a tumor may result.
Human papillomavirus (HPV) is a well-known oncogenic virus. This virus comes in many forms and often causes benign warts. However,
several strains of this virus are known to be oncogenic, and are the primary cause of cervical cancer. Women infected with oncogenic
strains of this HPV typically have no symptoms, but an annual Pap smear can detect changes in cervical cells that are consistent with HPV
infection. When detected early, infected cells can be removed.
Although the possibility of cancer strikes fear in many individuals, numerous genetic changes are usually required in order for cancer to
develop. In this chapter, you will learn about some of the genetic changes that contribute to the development of cancer.

12.1
Control of Gene Expression
The events of the cell cycle and DNA replication ensure that every cell of the body receives a copy of all the genes. This means that every nucleus in your
body has the potential to become a complete organism. To demonstrate this potential, let’s take a look at the process of reproductive and therapeutic
cloning.
Reproductive and Therapeutic Cloning
In reproductive cloning, the desired end is an individual that is exactly like the original individual. The cloning of plants has been routine for some time
(Fig. 12.1). However, at one time it was thought that the cloning of adult animals would be impossible because investigators found it difficult to have the
nucleus of an adult cell “start over,” even when it was placed in an enucleated egg cell. (An enucleated egg cell has had its own nucleus removed.)
In March 1997, Scottish investigators announced they had cloned a Dorset sheep, which they named Dolly. How was their procedure different
from all the others that had been attempted? Again, an adult nucleus was placed in an enucleated egg cell; however, the donor cells had been starved.
Starving the donor cells caused them to stop dividing and go into a resting stage (the G
0
stage of the cell cycle). The G
0
nuclei are amenable to
cytoplasmic signals for initiation of development (Fig. 12.2). Now it is common practice to clone farm animals that have desirable traits (Fig. 12.3), and
even to clone rare animals that might otherwise become extinct.
In the United States, no federal funds can be used for experiments to clone human beings. And at this point, even cloning farm animals is
wasteful—in the case of Dolly, out of 29 clones only one was successful. Also, cloned animals may not be healthy. Dolly was put down by lethal
injection in 2003 because she was suffering from lung cancer and crippling arthritis. She had lived only half the normal life span for a Dorset sheep.
In therapeutic cloning, the desired end is not an individual organism, but mature cells of various cell types. The purposes of therapeutic cloning
are (1) to learn more about how specialization of cells occurs, and (2) to provide cells and tissues that could be used to treat human illnesses such as
diabetes, spinal cord injuries, and Parkinson disease.
Therapeutic cloning can be carried out in several ways. One way follows the same procedure as reproductive cloning, except that embryonic cells,
called embryonic stem cells, are isolated, and each is subjected to a treatment that causes it to develop into a particular type of cell, such as red blood
cells, muscle cells, or nerve cells (see Fig. 12.2b). Ethical concerns exist about this type of therapeutic cloning because if the embryo had been allowed
to continue development, it would have become an individual. People are especially concerned when researchers short-cut this procedure and use
embryos that began life through in vitro fertilization in fertility clinics. Another way to carry out therapeutic cloning is to use adult stem cells. Stem cells
are found in many organs of an adult’s body. For example, the skin has stem cells that constantly divide and produce new skin cells, while the bone
marrow has stem cells that produce new blood cells. The goal is to develop techniques that would allow any adult stem cell to become any type of
specialized cell. In order to bring this about, researchers need to know how to control gene expression.
Levels of Gene Expression Control
The human body contains many types of cells that differ in structure and function. Each cell type must contain its own mix of proteins that make it
different from all other cell types. Therefore, only certain genes are active in cells that perform specialized functions, such as nerve, muscle, gland, and
blood cells.
Some of these active genes are called housekeeping genes because they govern functions that are common to many types of cells, such as glucose
metabolism. But otherwise, the activity of selected genes accounts for the specialization of cells. In other words, gene expression is controlled in a cell,
and this control accounts for its specialization (Fig. 12.4).
Let’s begin by taking a look at gene regulation in prokaryotes because prokaryotes utilize only one level of control. They control the transcription
of their genes.
Gene Expression in Prokaryotes
The bacterium Escherichia coli lives in the intestine and can quickly adjust its production of enzymes according to what we eat. If someone drinks a
glass of milk, E. coli immediately begins to make three enzymes needed to metabolize lactose. All three enzymes are under the control of one
promoter, a short DNA sequence where RNA polymerase first attaches when transcription occurs. The French microbiologists François Jacob and
Jacques Monad called such a cluster of bacterial genes, along with the DNA sequences that control their transcription, an operon. They received a
Nobel prize in 1961 for their investigations because they were the first to show how gene expression was controlled—specifically, how the lac operon
was
controlled
(Fig.
12.5).
In their model, a regulatory gene located outside the operon codes for a repressor, a protein with the potential to bind to the operator (a short
sequence of DNA near the promoter). The activity of the lac operon is not needed much of the time, and the regulatory gene codes for a repressor
that ordinarily binds to the operator. Now, RNA polymerase cannot attach to the promoter, and transcription cannot occur.
What turns the operon on when lactose is present? Lactose binds to the repressor, inactivating it; specifically, a change in shape prevents the
repressor from attaching to the operator. Now RNA polymerase is able to bind to the promoter, and transcription of the genes needed for lactose

metabolism can occur. Binding of RNA polymerase is further ensured by another regulatory protein called CAP, which is not shown in Figure 12.5.
In this way, these enzymes are produced only when they are needed to metabolize lactose.
Other bacterial operons, such as those that control amino acid synthesis, are usually turned on. For example, in the trp operon, the regulatory
gene codes for a repressor that ordinarily is unable to attach to the operator. Therefore, the genes needed to make the amino acid tryptophan are
ordinarily expressed. When tryptophan is present, it binds to the repressor. A change in shape activates the repressor and allows it to bind to the operator.
Now the operon is turned off.
Gene Expression in Eukaryotes
In bacteria, a single promoter serves several genes that make up a transcription unit, while in eukaryotes, each gene has its own promoter where RNA
polymerase binds. Bacteria rely purely on transcriptional control, but in eukaryotes, a variety of mechanisms regulate gene expression. These eukaryotic
mechanisms affect whether the gene is expressed, the speed with which it is expressed, and how long it is expressed.
Some mechanisms of gene expression occur in the nucleus, and others occur in the cytoplasm (Fig. 12.6). In the nucleus, DNA unpacking,
transcription, and mRNA processing all play a role in determining which proteins are made in the cytoplasm.
In the cytoplasm, translation of mRNA into a polypeptide at the ribosomes can occur right away or be delayed. The mRNA can last a long time or
be destroyed immediately, and the same holds true for a protein. These mechanisms control the quantity of gene product and/or how long it is active.
DNA Unpacking•Eukaryotes utilize chromatin packing as a way to keep genes turned off. Some inactive genes are located within darkly staining
portions of chromatin, called heterochromatin. A dramatic example of inactive heterochromatin is the Barr body in mammalian females. Females have
a small, darkly staining mass of condensed chromatin adhering to the inner edge of the nuclear envelope. This structure, called a Barr body after its
discoverer, is an inactive X chromosome.
How do we know that Barr bodies are inactive X chromosomes that are not producing gene product? Suppose 50% of the cells in a female have one
X chromosome active, and 50% have the other X chromosome active. Wouldn’t the body of a heterozygous female be a mosaic, with “patches” of
genetically different cells? This is exactly what happens. For example, human females who are heterozygous for an X-linked recessive form of ocular
albinism have patches of pigmented and nonpigmented cells at the back of the eye. And women who are heterozygous for hereditary absence of sweat
glands have patches of skin lacking sweat glands. The female calico cat also provides dramatic support for a difference in X-inactivation in its cells (Fig.
12.7).
Active genes in eukaryotic cells are associated with more loosely packed chromatin, called euchromatin. You learned in Chapter 8 that in
eukaryotes, a nucleosome is a portion of DNA wrapped around a group of histone molecules. When DNA is transcribed, a transcription activator pushes
aside the histone portions of nucleosomes so that transcription can begin (Fig. 12.8). In other words, even euchromatin needs further modification
before transcription can begin. The presence of nucleosomes limits access to DNA, and euchromatin becomes genetically active when histones no
longer bar access to DNA. Only then is it possible for a gene to be turned on and expressed in a eukaryotic cell.
DNA Transcription•Transcription in eukaryotes follows the same principles as in bacteria, except that many more regulatory proteins per gene are
involved. The occurrence of so many regulatory proteins allows for greater control, but it also allows for a greater chance of malfunction.
In eukaryotes, transcription factors are DNA-binding proteins that help RNA polymerase bind to a promoter. Several translation factors are
needed in each case, and if one is missing, transcription cannot take place. All the transcription factors form a complex that also helps pull double-stranded
DNA apart and even acts to position RNA polymerase so that transcription can begin. The same transcription factors, but not the same mix, are used over
again at other promoters, so it is easy to imagine that if one malfunctions, the result could be disastrous to the cell. The genetic disorder Huntington
disease is a devastating psychomotor ailment caused by a defect in a transcription factor.
In eukaryotes, transcription activators are DNA-binding proteins that speed transcription dramatically. They bind to a DNA region called an
enhancer that can be quite a distance from the promoter. A hairpin loop in the DNA can bring the transcription activators attached to enhancers into
contact with the transcription factor complex (Fig. 12.9).
It’s possible for a single regulatory protein to have a decisive effect if it is the last protein needed to turn on a gene. A s an example,
investigators have found one DNA-binding protein, MyoD, that alone causes fibroblasts to become muscle cells in various vertebrates. Another
DNA-binding protein, called Ey, can bring about the formation of not just a single cell type but a complete eye in flies (Fig. 12.10).
mRNA Processing•After transcription, messenger RNA (mRNA) is processed before it leaves the nucleus and passes into the cytoplasm. Removal of
introns and splicing of exons can vary the type of mRNA that leaves the nucleus. For example, both the hypothalamus and the thyroid gland produce a
hormone called calcitonin, but the mRNA that leaves the -nucleus is not the same in both types of cells. Radioactive -labeling studies show that they
vary because of a difference in the exons they contain (Fig. 12.11). Evidence of different patterns of mRNA splicing is found in other cells, such as those
that produce neurotransmitters and antibodies.
The speed of transport of mRNA from the nucleus into the cytoplasm can ultimately affect the amount of gene product realized following
transcription. Evidence indicates there is a difference in the length of time it takes various mRNA molecules to pass through a nuclear pore.
mRNA Translation•The cytoplasm contains proteins that can control whether translation of mRNA takes place. For example, an initiation factor,
known as IF-2, inhibits the start of protein synthesis when it is phosphorylated by a specific protein kinase. Environmental conditions can delay
translation. Red blood cells do not produce hemoglobin unless heme, an iron-containing group, is available.
The longer an mRNA remains in the cytoplasm before it is broken down, the more gene product is produced. During maturation, mammalian red
blood cells eject their nuclei, and yet they continue to synthesize hemoglobin for several months thereafter. The necessary mRNAs must be able to persist
all this time. Differences in the guanine cap or poly-A tail determine how long a particular transcript remains active before it is destroyed by a ribonuclease
associated with ribosomes. Hormones cause the stabilization of certain mRNA transcripts. An mRNA called vitellin persists for 3 weeks instead of 15 hours
if it is exposed to estrogen.
Protein Activity•Some proteins are not active immediately after synthesis. For example, insulin is a single long polypeptide that folds into a
three-dimensional structure. Only then is a sequence of about 30 amino acids enzymatically removed from the middle of the molecule. This leaves two
polypeptide chains bonded together by disulfide (S±S) bonds, and an active insulin molecule results (Fig. 12.12).
Feedback control usually regulates the activity of proteins. Many proteins are short-lived in cells because they are degraded or destroyed. The cell
has giant enzyme complexes, called proteosomes, that carry out this task.
Signaling Between Cells in Eukaryotes
In multicellular organisms, cells are constantly sending out chemical signals that influence the behavior of other cells. Dur ing animal development,
these signals determine the specialized role a cell will play in the organism. Later, the signals help coordinate growth and day-to-day functions.
Plant cells also signal each other so that their responses to environmental stimuli, such as direct sunlight, are coordinated .
Typically, a cell-signaling pathway begins when a chemical signal binds to a receptor protein in a target cell’s plasma membrane. The signal causes
the receptor protein to initiate a series of reactions within a signal transduction pathway. The end product of the pathway (not the signal) directly affects
the metabolism of the cell. For example, growth is only possible if certain genes have been turned on by regulatory proteins.
In Figure 12.13, a signaling cell secretes a chemical signal that binds to a specific receptor located in the receiving cell’s plasma membrane. The
binding activates a series of reactions within a signal transduction pathway. The last reaction activates a transcription activator that enhances the
transcription of a specific gene. Transcription leads to the translation of mRNA and a protein product that, in this case, stimulates the cell cycle so that
growth occurs.
A protein called Ras functions in signaling pathways that lead to the transcription of many genes, several of which promote the cell cycle. Ras is
normally inactive, but the reception of a growth factor leads to its activation. Ras becomes active when it is phosphorylated.
12.2
Cancer: A Failure
of Genetic Control
As described in Chapter 8, cancer is a genetic disease requiring several mutations, each propelling cells toward the development of a tumor, an
accumulation of cancer cells that no longer have the characteristics of normal cells (Table 12.1). Several mutations are usually needed to disrupt the
many redundant regulatory pathways that normally ensure a cell remains normal in structure and function. Because of this sequence, it takes several
years for cancer to develop, and the likelihood of cancer increases as we age.
Cells that are already highly specialized, such as nerve cells and cardiac muscle cells, do not become cancer cells; rather, carcinogenesis (the
development of cancer) is more likely to begin in cells that have the capacity to enter the cell cycle. Fibroblasts and cells lining the cavities of the lung,
liver, uterus, and kidney are able to divide when stimulated to do so. Adult stem cells continue to divide throughout life. These include blood-forming
cells in the bone marrow and basal cells of the skin and digestive tract. Continuous division of these cells is required because blood cells live only a few
days, and cells that line the intestine and cells that form the outer layer of the skin are continually sloughed off. The development of colon cancer, in
particular, has been studied, and Figure 12.14 is based on those studies. This figure shows that a single abnormal cell begins the process toward the
development of a tumor. And along the way, the most aggressive cell becomes the dominant cell of the tumor.
Cancer cells release a growth factor that causes neighboring blood vessels to branch into the cancerous tissue, a process called angiogenesis.
Cancer cells also produce enzymes that degrade the basement membrane and then they invade underlying tissues. Unlike most tissue cells, cancer cells
are motile and able to travel through the blood or lymph vessels to other parts of the body, where they start distant tumors.
Proto-Oncogenes and Tumor Suppressor Genes
Recall that the cell cycle, as discussed on page 114, consists of interphase, followed by mitosis. Cyclin is a molecule that has to be present for a cell to
proceed from interphase to mitosis. When cancer develops, the cell cycle occurs repeatedly, in large part, due to mutations in two types of genes.
Figure 12.15 shows that:
1. Proto-oncogenes: code for proteins that promote the cell cycle and inhibit apoptosis. They are often likened to the gas pedal of a car because they
cause acceleration of the cell cycle.
2. Tumor suppressor genes: code for proteins that inhibit the cell cycle and promote apoptosis. They are often likened to the brakes of a car

because they inhibit acceleration of the cell cycle.
Proto-Oncogenes Become Oncogenes
When proto-oncogenes mutate, they become cancer-causing genes called oncogenes. These mutations can be called “gain-of-function” mutations
because overexpression is the result (Fig. 12.16). Whatever a proto-oncogene does, an oncogene does it better.
A growth factor is a signal that activates a cell-signaling pathway, resulting in cell division. Some proto-oncogenes code for a growth factor or
for a receptor protein that receives a growth factor. When these proto-oncogenes become oncogenes, receptor proteins are easy to activate and even may
be stimulated by a growth factor produced by the receiving cell. Several proto-oncogenes code for Ras proteins that promote mitosis by activating
cyclin. Ras oncogenes are typically found in many different types of cancers. Cyclin D is a proto-oncogene that codes for a cyclin directly. When this
gene becomes an oncogene, cyclin is readily available all the time.
p53 is a transcription factor instrumental in stopping the cell cycle and activating repair enzymes. If repair is impossible, the p53 protein
promotes apoptosis, programmed cell death. Apoptosis is an important way carcinogenesis is prevented. A certain proto-oncogene codes for a protein
that functions to make p53 unavailable. When this proto-oncogene becomes an oncogene, no matter how much p53 is made, none will be available.
Many tumors are lacking in p53 activity.
Tumor Suppressor Genes Become Inactive
When tumor suppressor genes mutate, their products no longer inhibit the cell cycle nor promote apoptosis. Therefore, these mutations can be called
“loss of function” mutations (Fig. 12.16).
The retinoblastoma protein (RB) controls the activity of a transcription activator for cyclin D and other genes whose products promote entry into
the S phase of the cell cycle. When the tumor suppressor gene p16 mutates, the RB protein is always functional and the result is, again, too much active
cyclin D in the cell. The cell experiences repeated rounds of DNA synthesis without the occurrence of mitosis.
The protein Bax promotes apoptosis. When a tumor suppressor gene Bax mutates, the protein Bax is not present, and apoptosis is less likely to occur.
The gene contains a run of eight consecutive G bases, making it subject to mutations.
Other Genetic Changes
Cancer cells also undergo other genetic changes that result in certain other specific characteristics.
Absence of Telomere Shortening
A telomere is a repeating DNA sequence (TTAGGG) at the end of the chromosomes that can be as long as 15,000 base pairs. Telomeres have been likened
to the protective caps on the ends of shoelaces. Rather than keeping chromosomes from unraveling, however, telomeres stop chromosomes from fusing to
each other. Each time a cell divides, some portion of a telomere is lost; when telomeres become too short, the chromosomes fuse and can no longer
duplicate. Then the cell is “old” and dies by apoptosis.
Embryonic cells and also certain adult cells, such as stem cells and germ cells, have an enzyme called telomerase that can rebuild the telomeres.
The gene that codes for telomerase is turned on in cancer cells, which allows them to keep dividing over and over again. Telomerase is believed to
become active after a cell has already started proliferating wildly.
Angiogenesis
Angiogenesis, as noted earlier, is the formation of new blood vessels. To grow larger than, say, a pea, a tumor must have a well--developed capillary
network to bring it nutrients and -oxygen. Some of the growth factors released by cancer cells, such as vascular endothelial growth factor (VEGF), bind
to receptors on the epithelial cells of blood vessels. Thereafter, the epithelial cells grow and divide, producing new blood vessels that branch and send
capillaries into the center of a tumor.
The new blood vessels supply tumor cells with the nutrients and oxygen they require for rapid growth, but they also rob normal tissues of nutrients
and oxygen. Two highly effective drugs, called angiostatin and endostatin, have been shown to inhibit angiogenesis and are now being studied as
possible anticancer drugs.
Metastasis
A benign tumor is a disorganized, usually encapsulated mass that does not invade adjacent tissue. Cancer in situ is a -tumor located in its place of origin,
before any invasion of normal tissue has occurred (Fig. 12.17).
Malignancy is present when metastasis establishes new tumors distant from the primary tumor. To metastasize, cancer cells must make their way
across a basement membrane and into a blood vessel or lymphatic vessel. Cancer cells produce proteinase enzymes that degrade the basement
membrane and allow them to invade under-lying tissues.
Due to mutations of particular genes, cancer cells tend to be motile because they have a disorganized internal cytoskeleton and lack intact actin
filament bundles. After traveling through the blood or lymph, cancer cells start tumors elsewhere in the body. Not many cancer cells achieve this feat,
but when they do, the prognosis for recovery becomes doubtful.
Hereditary Forms of Cancer
The environmental influences discussed in Chapter 8 (radiation, organic chemicals, and viruses) are major risk factors for the development of cancer.
These agents can cause mutations in tumor suppressor genes and proto-oncogenes so that cancer is more likely. Although cancer is usually a somatic
disease, meaning that it develops only in body cells, some instances of an inherited predisposition to cancer are known.
Inheritance Patterns for Cancer
BRCA1 and BRCA2•In 1990, DNA linkage studies on large families in which females tended to develop breast cancer identified the first gene
associated with breast cancer. Scientists named this gene breast cancer 1 or BRCA1 (pronounced brak-uh). Later, they found that breast cancer in other
families was due to another breast cancer gene they called BRCA2. These genes are tumor suppressor genes that are known to behave as if they are
autosomal recessive alleles. If one mutated allele is inherited from either parent, a mutation in the other allele is required before the predisposition to
cancer is increased. Because the first mutated gene is inherited, it is present in all cells of the body, and then cancer is more likely wherever the second
mutation occurs. If the second mutation occurs in the breast, breast cancer may develop (Fig. 12.18). If the second mutation is in the ovary, ovarian
cancer may develop if additional cancer-causing mutations occur.
RB Gene•The RB gene is also a tumor suppressor gene. It takes its name from its association with an eye tumor called a retinoblastoma, which first
appears as a white mass in the retina. A tumor in one eye is most common because it takes mutations in both alleles before cancer can develop (Fig.
12.19). Children who inherit a mutated allele are more likely to have tumors in both eyes.
RET Gene•An abnormal RET gene, which predisposes a person to thyroid cancer, can be passed from parent to child. RET is a proto-oncogene known
to be inherited in an autosomal dominant manner—only one mutated allele is needed to increase a predisposition to cancer. The remainder of the
mutations necessary for thyroid cancer to develop are acquired (not inherited).
Testing for These and Other Genes
Genetic tests are available for the presence of the BRCA genes, the RET gene, and the RB gene. Persons who have inherited these genes may decide
to have surgery by choice rather than necessity or be examined frequently for signs of cancer.
Genetic tests are also available for other types of mutated genes that help a physician diagnose cancer. For example, a ras oncogene can be
detected in stool and urine samples. If the test for ras oncogene in the stool is positive, the physician suspects colon cancer, and if the test for ras
oncogene in the urine is positive, the physician suspects bladder -cancer.
Telomerase, you may recall, is the enzyme that keeps telomeres a constant length in cells. The gene that codes for telomerase is turned off in
normal cells but is active in cancer cells. Therefore, if the test for the presence of telomerase is positive, the cell is likely cancerous.
T H E C H A P T E R I N R E V I E W
Summary
12.1 Control of Gene Expression
Every cell in the body has a complete set of genes as is proven by the process of cloning, when a 2n nucleus is able to bring about the development of an
organism exactly like the 2n nucleus donor or any specialized tissue that matches those of the donor.
Reproductive and Therapeutic Cloning
In reproductive cloning, the desired end is an individual genetically identical to the original individual. To achieve this type of cloning, the early embryo is
placed in a surrogate mother until the individual comes to term.
In therapeutic cloning, the cells of the embryo are separated and treated so that they develop into specialized tissues that can be used to treat
human disorders. Alternately, adult stem cells can be treated to become specialized tissues.
Levels of Gene Expression Control
The specialization of cells is not due to the presence/absence of genes; it is due to the activity/inactivity of genes.
Gene Expression in Prokaryotes•In prokaryotes, gene expression is controlled at the level of transcription.
• In the lac operon, the repressor usually binds to the operator. Then RNA polymerase cannot bind, and the operon is turned off. When lactose is
present, it binds to the repressor, and now RNA polymerase binds to the promoter, and the genes for lactose metabolism are transcribed.
• In the trp operon, the genes are usually transcribed because the repressor is inactive. When tryptophan is present, the repressor becomes active,
and RNA polymerase cannot bind to the promoter.
Gene Expression in Eukaryotes•Gene expression in eukaryotes occurs at various levels:
• DNA unpacking—heterochromatin becomes active.
• DNA transcription—transcription factors present at the promoter, and transcription activators (bound to enhancers) are brought in contact and form
a transcription initiation complex.

• mRNA processing—alternate patterns of processing; how quickly mRNA leaves the nucleus.
• Translation of mRNA—how long the mRNA stays active.
• Protein activity—activation of a protein may require further steps and degradation of the protein can occur at once or after a while.
Signaling Between Cells in Eukaryotes•Eukaryotic cells constantly communicate with their neighbors by secreting signals.
• Signals are received at receptor proteins located in the plasma membrane.
• A signal transduction pathway (enzymatic reactions) leads to a change in the behavior of the cell. For example, when stimulated via a transduction
pathway, a transcription activator stimulates transcription of a particular gene.
12.2 Cancer: A Failure of Genetic Control
Cancer Is a Genetic Disease
Evidence is strong that cancer is a genetic disease. A series of mutations, particularly in cells that can divide, lead to cancer cells with these
characteristics:
• No contact inhibition
• Uncontrolled growth
• Are nonspecialized
• Have abnormal chromosomes
• Do not undergo apoptosis
Proto-Oncogenes and Tumor Suppressor Genes
• Proto-oncogenes ordinarily promote the cell cycle and inhibit apoptosis. Mutation results in a gain of function for these activities.
• Tumor suppressor genes ordinarily suppress the cell cycle and promote apoptosis. Mutation results in loss of function for these activities.
Mutations in both types of cells result in
• Excess cyclin to stimulate the cell cycle
• No p53 to bring about apoptosis
Other Genetic Changes
• Telomeres at ends of chromosomes don’t shorten because telomerase is present.
• Angiogenesis, formation of new blood vessels, occurs because cancer cells release VEGF.
• Metastasis, spread of cancer to remote areas, occurs because cells are mobile.
Hereditary Forms of Cancer
Mutagens such as radiation, organic chemicals, and viruses, are also major risk factors for the development of cancer.
Inheritance Patterns for Cancer•Certain genes are known to cause cancer, including:
• BRCA1 and BRCA2, which are tumor suppressor genes. If one mutated gene is inherited and a second mutates, cancer is more likely.
• The RB gene is also a tumor suppressor gene. When a mutated allele is inherited, tumors in both eyes are more likely.
• An abnormal RET gene, which predisposes a person to thyroid cancer, can also be passed from parent to child.
Testing for Cancer Genes•Genetic tests are available to help a physician diagnose cancer. These genes are turned off in normal cells, but are active in
cancer cells. For example, if the test for the presence of telomerase is positive, the cell is cancerous.
Thinking Scientifically
1. A person with Turner syndrome is XO and phenotypically female. Typical symptoms of Turner syndrome include rudimentary sexual development,
extra folds of skin on the neck, and short stature. If normal (XX) women have only one active X chromosome, why would someone with a single X
chromosome exhibit this syndrome?
2. Most cat breeders know that male tortoiseshell cats are very rare. In addition, these cats are inevitably sterile. Explain these two observations.
Testing Yourself
Choose the best answer for each question.
1. During reproductive cloning, a(n) _________ is placed into a(n) _________.
a. enucleated egg cell, adult cell nucleus
b. adult cell nucleus, enucleated egg cell
c. egg cell nucleus, enucleated adult cell
d. enucleated adult cell, egg cell nucleus
2. The major challenge to therapeutic cloning using adult stem cells is
a. finding appropriate cell types.

b. obtaining enough tissue.
c. controlling gene expression.
d. keeping cells alive in culture.
3. Cancer cells are immortal because
a. they produce telomerase.
b. their tumor suppressor genes stimulate apoptosis.
c. they have an unlimited food supply.
d. they can complete mitosis without cytokinesis.
4. Label this operon with these terms: active repressor, lactose metabolizing genes, operator, RNA polymerase promoter, regulatory gene, mRNA
5. Which of the following is not a gene expression control mechanism in eukaryotes?
a. mRNA processing
b. rate of ribosome synthesis
c. longevity of mRNA
d. speed of passage of mRNA through nuclear pores
6. What types of offspring would you expect from a female tortoiseshell cat and a black male? Remember that this coat color gene is due to
heterozygous alleles on the X chromosomes.
a. tortoiseshell females and black males
b. all black cats
c. tortoiseshell females, black females, orange males, black males
d. orange females and black males
For questions 7
–11, match the examples to the gene expression control mechanisms in the key. Each answer can be used more than once.
Key:
a. DNA unpacking
b. mRNA processing
c. translation of mRNA
d. protein activity
7. Insulin does not become active until 30 amino acids are cleaved from the middle of the molecule.
8. The mRNA for vitellin is longer-lived if it is exposed to estrogen.
9. Genes in Barr bodies are inactivated.
10. Calcitonin is produced in both the hypothalamus and the thyroid gland, but in different forms due to exon splicing.
11. A translation initiation factor inhibits translation when it is phosphorylated by kinase.
12. Suppose a woman is heterozygous for the RB
gene that causes retinoblastoma. Her husband’s mother was homozygous for the mutant RB allele,
but his father’s side of the family has no history of retinoblastoma. What is the probability that this couple will have a child who does not carry the
mutant RB allele?
a.
1
⁄
4
c.
3
⁄
4
b.
1
⁄
2
d. 0
13. A cell is likely to be cancerous if it tests positive for a (an)
a. elongation factor.
c. enhancer.
b. transcription factor.
d. telomerase.
14. Which of the following correctly describes reproductive cloning?
a.
The desired end is an individual that is a genetic improvement over the original individual.
b. The desired end is mature cells of various cell types.
c.
An embryonic nucleus is placed in an enucleated and starved embryonic donor cell.
d.
Products of this process show signs of being unhealthy and may age more rapidly.
e.
Ethical concerns surround the embryo, which could become an individual if allowed.
15. The product of therapeutic cloning differs from the product of reproductive cloning in that it is
a. various mature cells, not an individual.
b. genetically identical to the somatic cells from the donor.
c. various mature cells with the ability to become an individual.
d.
an individual that is genetically identical to the donor of the nucleus.
e.

a result of starving donor cells, causing them to stop dividing and go into a resting state.
16. What is a Barr body?
a. an example of gene regulation using chromatin packing
b. an inactive X chromosome
c. an X chromosome not producing gene products
d. explains tortoiseshell cats with patches of black and orange fur
e. All of these are correct.
17. How is transcription controlled in eukaryotic cells?
a. through the use of signal transduction pathways
b. by means of cell signaling
c. using transcription factors and activators
d. when chromatin is packed to keep genes turned on
e. None of these are correct.
18. An operon is a short sequence of DNA
a. that prevents RNA polymerase from binding to the promoter.
b. that prevents transcription from occurring.
c. and the sequences that control its transcription.
d. that codes for the repressor protein.
e.
that functions to prevent the repressor from binding to the operator.
19. Which of these correctly describes the function of a regulatory gene for the lac operon?
a. prevents transcription from occurring
b. a sequence of DNA that codes for the repressor
c. prevents the repressor from binding to the operator
d. keeps the operon off until lactose is present
e. Both b and d are correct.
20. What is the purpose of cell signaling in eukaryotes?
a. to change the behavior of the receiving cell
b. to initiate a set of reactions through the receptor protein
c. to turn on a gene
d. to transmit a nerve impulse, for example
e. All of these are correct.
21. Eukaryotic cells signal one another through the use of
a. repressor proteins acting upon operons.
b.
molecules that bind to a receptor protein in the membrane of a target cell.
c.
transcription factors and activators acting upon RNA polymerase.
d.
RNA polymerase acting upon transcription promoters and enhancers.
e.
nerve impulses that cause a receiving cell to change its behavior.
22. Which of these is not a possible cause of cancer?
a. radiation
d. genes
b. metastasis
e. oncogenes
c. viruses
23. Which of the following is not an example of why cancer cells are a failure of genetic control?
a.
They exhibit contact inhibition and do not grow in multiple layers.
b.
They lack specialization and do not contribute to the function of a body part.
c.
They have enlarged nuclei and may have an abnormal number of chromosomes.
d.
They fail to undergo apoptosis even though they are abnormal cells.
e.
They release a growth factor that causes neighboring blood vessels to branch into the tumor and therefore feed the cancerous tissue.
Go to www.mhhe.com/maderessentials for more quiz questions.

Bioethical Issue
Over the past decade, genetic tests have become available for certain cancer genes. If women test positive for defective BRCA1 and BRCA2 genes, they
have an increased risk for early-onset breast and ovarian cancer. If individuals test positive for the APC gene, they are at greater risk for the development
of colon cancer. Other genetic tests exist for rare cancers, including retinoblastoma and Wilms tumor.
Advocates of genetic testing say that it can alert those who test positive for these mutated genes to undergo frequent mammograms or
colonoscopies. Early detection of cancer clearly offers the best chance for successful treatment. Others feel that genetic testing is unnecessary because
nothing can presently be done to prevent the disease. Perhaps it is enough for those who have a family history of cancer to schedule frequent checkups,
beginning at a younger age.
People opposed to genetic testing worry that being predisposed to cancer might threaten their job or health insurance. They suggest that genetic
testing be confined to a research setting, especially since it is not known which particular mutations in the genes predispose a person to cancer. They are
afraid, for example, that a woman with a defective BRCA1 or BRCA2 gene might make the unnecessary decision to have a radical mastectomy. The lack
of proper counseling also concerns many. In a study of 177 patients who underwent APC gene testing for susceptibility to colon cancer, less than 20%
received counseling before the test. Moreover, physicians misinterpreted the test results in nearly one-third of the cases.
Should everyone be aware that genetic testing for cancer is a possibility, or should genetic testing be confined to a research setting? If genetic testing
was offered to you, would you take advantage of it?
Understanding the Terms
angiogenesis•190
cell-signaling pathway•187
cyclin•189
enhancer•186
euchromatin•184
growth factor•189
heterochromatin•184
metastasis•190
oncogene•189
operon•182
proto-oncogene•189
regulatory gene•183
repressor•183
reproductive cloning•180
signal transduction pathway•187
telomere•190
therapeutic cloning•181
transcription activator•186
transcription factor•186
tumor suppressor gene•189
Match the terms to these definitions:
a. _______________
Set of bacterial genes under the control of a single promoter.
b. _______________
Protein that binds to the operator of an operon.
c. _______________
Darkly stained and tightly packed chromatin.
d. _______________
Protein in eukaryotes that helps RNA polymerase bind to a promoter.
e. _______________
DNA region to which transcription activators bind to speed transcription in eukaryotes.
f. _______________
Signal that activates a cell-signaling pathway.
g. _______________
Repeating DNA sequence at the end of a chromosome.
Chromosomal abnormalities are rampant in cancer cells.
A papillomaviral infection can lead to cervical cancer.
Several mutations are usually necessary for cancer cells to develop.
Check Your Progress
Contrast reproductive cloning with therapeutic cloning.
Answer:•The goal of reproductive cloning is to create an individual that is genetically identical to the 2n nucleus donor. The goal of therapeutic cloning is to study how cells
become specialized and to provide specialized cells for medical procedures.

Check Your Progress
Contrast when the lac operon is turned on with when the trp operon is turned off.
Answer:•The lac operon (which breaks down lactose) is turned on only when lactose is present. The trp operon (which synthesizes tryptophan) is turned off only when
tryptophan is present.
Check Your Progress
List the levels of gene expression that can be controlled in eukaryotes.
Answer:•DNA unpacking, transcription of DNA, mRNA processing, translation of mRNA, protein activity.
Check Your Progress
1.
Contrast heterochromatin with euchromatin.
2. Explain how mRNA processing can influence the types of gene products made by a cell.
3. Explain how a mammalian red blood cell can synthesize hemoglobin after it ejects its nucleus.
Answers:•1. Heterochromatin is tightly packed and darkly stained, and generally contains inactive genes. Euchromatin is loosely packed and lightly stained, and generally
contains active genes.•2. The exons that are spliced out may differ in different cell types, resulting in varying types of products after translation.•3. Its mRNA survives for
many months, allowing translation to make hemoglobin.
Check Your Progress
1. Describe the function of transcription factors in eukaryotes.
2.
Describe the relationship between transcription activators and enhancers.
3. Describe the relationship between a signal transduction pathway and gene expression.
Answers:•1. Transcription factors help RNA polymerase bind to promoter regions, allowing transcription to begin.•2. Transcription activators are proteins that bind to
DNA regions called enhancers to speed
transcription.•3. The signal transduction pathway is a series of reactions that changes the signal to one that affects the activity of a cell such as increased expression of
genes.
Check Your Progress
1. Explain why cancer incidence increases with age.
2.
Explain why specialized cells are less likely to be involved in cancer than other cells.
3.
Contrast proto-oncogenes with tumor suppressor genes.
Answers:•1. Cancer results from an accumulation of mutations over time.•
2. Specialized cells are not programmed to undergo the cell cycle anymore, so cancer (which is a cell cycle control disease) is less likely to develop.•
3. When proto-oncogenes are active, the cell cycle occurs normally and apoptosis is restrained. When tumor suppressor genes are active the cell cycle is restrained and
apoptosis is promoted.
Check Your Progress
1.
Explain how telomeres influence a cell’s life span.
2.
Explain why angiogenesis is important for cancer growth.
3. Compare and contrast the BRCA gene with the RB gene.
Answers:•1. Telomeres become shorter after each cell division. When the telomere becomes too short, chromosomes fuse together and the cell dies.•
2. The formation of new blood cells supplies a tumor with the nutrients it needs for growth.•3. Both are tumor suppressor genes. BRCA is associated with breast cancer,
while RB causes retinoblastoma.
Figure 12.1•Cloning carrots.
Carrot cells from a root will produce cloned copies of carrot plants in cell culture. This demonstrates that all the nuclei in a carrot root contain all the genes of a carrot
plant.
Figure 12.4•Gene expression in specialized cells.
All the cells in your body contain all the genes, but gene expression is controlled, and only certain genes are expressed in each type of cell. Housekeeping genes are
expressed in all cells; in addition, cells express those genes that account for their specialization.
Figure 12.5•The lac operon.
a. The regulatory gene codes for a repressor. The repressor binds to the operator and prevents RNA polymerase from attaching to the promoter. Therefore, transcription
of the structural genes does not occur. b. When lactose is present, it binds to the repressor, changing its shape so that it becomes inactive and can no longer bind to the
operator. Now RNA polymerase binds to the promoter, and the structural genes are expressed.
Figure 12.8•DNA unpacking.
Compact chromatin is made accessible for transcription when a transcription activator works with a chromatin remodeling complex to push aside the histone portions of
nucleosomes.
Figure 12.7•X-inactivation in mammalian females.
In cats, the alleles for black or orange coat color are carried on the X chromosome. Random X-inactivation occurs in females. Therefore, in heterozygous females, 50%
of the cells have an allele for black coat color, and 50% of cells have an allele for orange coat color. The result is calico or tortoiseshell cats that have coats with patches
of both black and orange.
Figure 12.9•Transcription factors and transcription activators.
Transcription factors form a complex at a promoter and a transcription activator binds to an enhancer. RNA polymerase binds to the promoter but transcription does not
begin until DNA loops, bringing all regulatory proteins together.
Figure 12.10•Ey gene.
In fruit flies, the expression of the Ey gene, in the precursor cells of the leg, triggers the development of an eye on the leg.
Figure 12.14•Development of cancer.
Development of cancer requires several mutations, each contributing to the development of a tumor. Normally cells exhibit contact inhibition and controlled growth. They
stop dividing when they are bordered by other cells. Tumor cells lack contact inhibition and pile up forming a tumor. Cancer cells do not fulfill the functions of a
specialized tissue and they have abnormal chromosomes. Even so, they do not undergo apoptosis (cell death) and keep on dividing forever. Eventually they form tumors
in other parts of the body.
Figure 12.17•Benign tumor versus cancer in situ.
a. A benign tumor is encapsulated. b. Cancer in situ is not encapsulated and is more likely to become a metastatic tumor.
Figure 12.2•Two types of cloning.
a. The purpose of reproductive cloning is to produce an individual that is genetically identical to the one that donated a 2n nucleus. The 2n nucleus is placed in an
enucleated egg, and after several divisions, the embryo comes to term in a surrogate mother.
b. The purpose of therapeutic cloning is to produce specialized tissue cells. A 2n nucleus is placed in an enucleated egg,
and after several divisions, the embryonic cells (called embryonic stem cells) are separated and treated to
become specialized cells.
Figure 12.3•Cloned farm animals.
These pigs were cloned using 2n nuclei from an adult swine. All nuclei contain all the genes of an organism or
else cloning wouldn’t be possible.
Figure 12.6•Control of gene -expression in eukaryotic cells.
Gene expression which can be turned on by an external signal (red) is controlled at various levels in eukaryotic cells.
Figure 12.12•Protein activity.
The protein insulin is not active until an enzyme removes a portion of the initial polypeptide. The resulting two polypeptide chains are held together by two disulfide
(S±S) bonds between like amino acids.
Figure 12.11•Processing of mRNA transcripts.
Because the primary mRNAs are processed differently in these two cells (a) and (b), distinct proteins result.
Figure 12.13•Cell-signaling pathway.
A chemical signal is received at a specific receptor protein.
A signal transduction pathway terminates when a transcription activator is stimulated. It enhances transcription of a specif ic gene.
Translation of the mRNA
transcript follows and a protein product results. The protein product is the response to the signal.
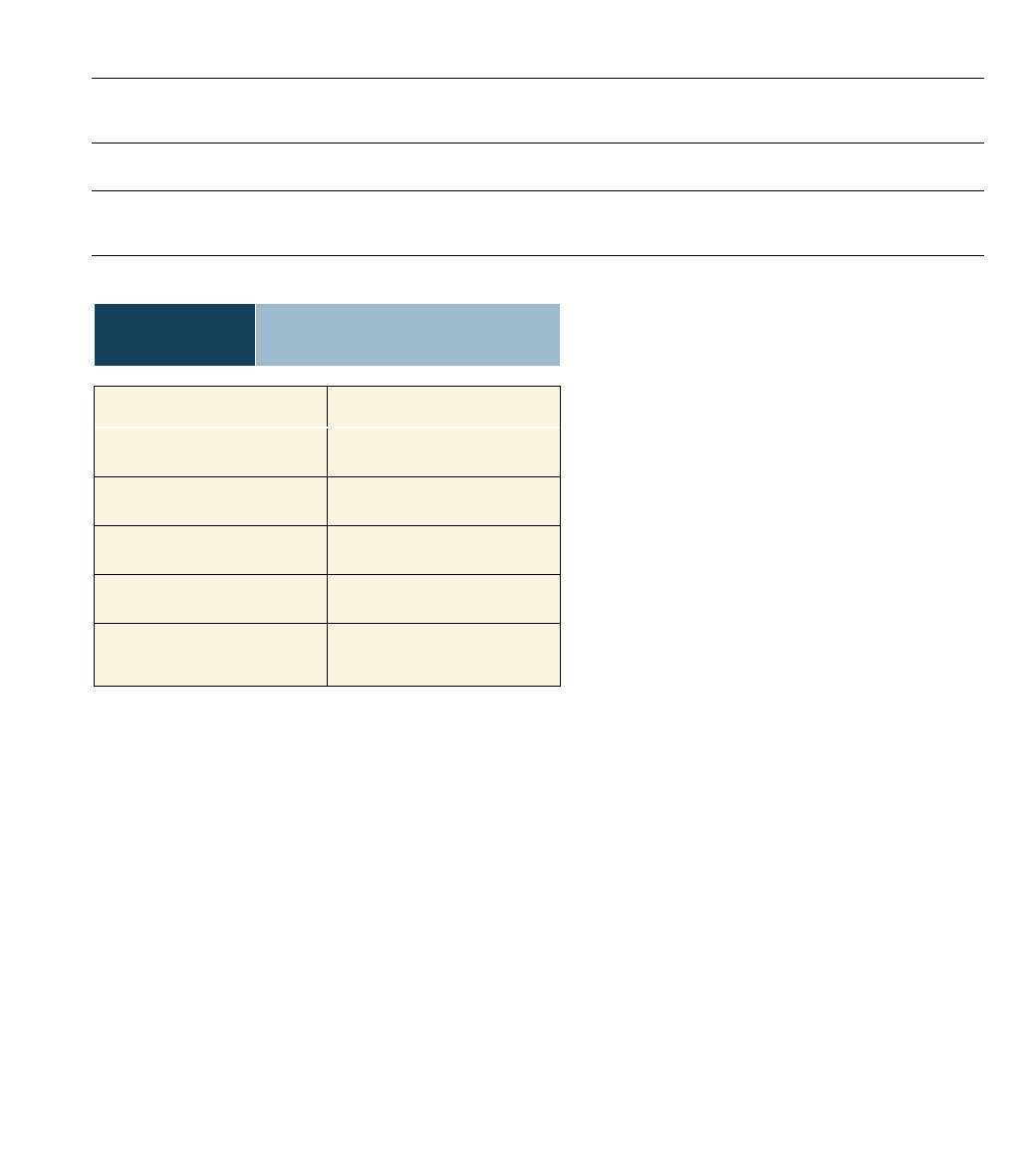
Figure 12.15•Normal cells.
In normal cells, proto-oncogenes code for sufficient cyclin to keep the cell cycle going normally, and they code for proteins that inhibit p53 and apoptosis. Tumor
suppressor genes code for proteins that inhibit cyclin, and promote p53 and apoptosis.
Figure 12.16•Cancer cells.
Notice that oncogenes and mutated tumor suppressor genes have the same end effects: unrestrained cell cycle and apoptosis does n ot occur.
Figure 12.18•Breast cancer can run in families.
These three sisters have all had breast cancer. Genetic tests can identify women at risk for breast cancer so that they can choose to have frequent examinations to allow
for early detection.
Figure 12.19•Inherited retinoblastoma.
A child is at risk for an eye tumor when a mutated RB allele is inherited, even though a second mutation in the normal allele is required before the tumor develops.
Table 12.1
Normal Cells Compared
to Cancer Cells
Normal Cells
Cancer Cells
Contact inhibition
No contact inhibition
Controlled growth
Uncontrolled growth (tumor)
Specialized cells
Nonspecialized cells
Normal chromosomes
Abnormal chromosomes
Undergo apoptosis
No apoptosis
lymphoma
cancer
cell
blood
cancer cells
cervical
smear
SEM of breast cancer cell
Wyszukiwarka
Podobne podstrony:
Essentials of Biology 1e appendix b
Essentials of Biology 1e appendix b
Essentials of Biology 1e index
Essentials of Biology 1e c 17
Essentials of Biology 1e mad86161 ise
Essentials of Biology 1e c 09
Essentials of Biology 1e c 28
Essentials of Biology 1e c 08
Essentials of Biology 1e c 04
Essentials of Biology 1e c 15
Essentials of Biology 1e c 27
Essentials of Biology 1e credits
Essentials of Biology 1e c 29
Essentials of Biology 1e c 16
Essentials of Biology 1e c 31
Essentials of Biology 1e c 22
Essentials of Biology 1e c 18
Essentials of Biology 1e c 19
Essentials of Biology 1e c 32
więcej podobnych podstron