
Protein phosphatase 2A C
a
is involved in osteoclastogenesis by regulating
RANKL and OPG expression in osteoblasts
Hirohiko Okamura
,
, Di yang
, Kaya Yoshida
, Tatsuji Haneji
a
Department of Histology and Oral Histology, Institute of Health Biosciences, The University of Tokushima Graduate School, 3-18-15, Kuramoto, Tokushima 770-8504, Japan
b
Department of Oral Health Care Education, Institute of Health Biosciences, The University of Tokushima Graduate School, 3-18-15, Kuramoto, Tokushima 770-8504, Japan
a r t i c l e
i n f o
Article history:
Received 28 June 2012
Revised 4 October 2012
Accepted 23 October 2012
Available online xxxx
Edited by Zhijie Chang
Keywords:
Protein phosphatase 2A
RANKL
OPG
NFAT
Osteoclastogenesis
a b s t r a c t
We examined whether alteration of PP2A C
a
expression in osteoblasts is involved in osteoclast dif-
ferentiation. Reduction of PP2A C
a
in MC3T3-E1 cells (shPP2A) decreased receptor activator of
nuclear factor
j
B ligand (RANKL) expression and increased osteoprotegerin (OPG) expression. The
conditioned medium from shPP2A cells failed to induce NFATc1 as well as the expression of osteo-
clast marker genes cathepsin K and osteoclast-associated receptor (OSCAR) in bone marrow macro-
phage cells. Treatment of bone marrow macrophage cells with the conditioned medium from
shPP2A cells impaired osteoclastogenesis. These results suggest that alteration of PP2A C
a
expres-
sion in osteoblasts modulates the expressions of RANKL and OPG, which are involved in osteoclasto-
genesis via the NFATc1 transcription factor.
Ó 2012 Federation of European Biochemical Societies. Published by Elsevier B.V. All rights reserved.
1. Introduction
The bone is a dynamic tissue that provides mechanical support,
physical protection, and a storage site for systemic mineral homeo-
stasis. Bone remodeling is carried out by osteoblasts and osteo-
clasts derived from independent lineages (mesenchymal and
hematopoietic). Osteoblasts produce both stimulatory and inhibi-
tory factors that tightly regulate osteoclast formation and activity
.
A member of the tumor necrosis factor (TNF) family—receptor
activator of nuclear factor
j
B ligand (RANKL)—has been shown to
regulate osteoclast differentiation and function directly
.
RANKL is found both as a transmembrane molecule on osteoblasts
and as a secreted molecule—soluble RANKL (sRANKL)
. Binding
of RANKL to its receptor, RANK, on the surface of osteoclast precur-
sor cells initiates signals, which leads to osteoclastogenesis. On the
other hand, osteoprotegerin (OPG) is a competitive receptor for
RANKL and has been shown to prevent bone destruction by block-
ing the binding of RANKL with the receptor RANK, thereby inhibit-
ing osteoclast differentiation and activation
. Therefore,
alterations in the ratio of RANKL/sRANKL to OPG produced from
osteoblasts affect bone turnover and homeostasis by controlling
osteoclastogenesis
.
The binding of RANKL/sRANKL to its receptor RANK on the sur-
face of osteoclast precursor cells activates various transcription
factors, including nuclear factor of activated T cells c1 (NFATc1)
. In response to an increase in intracellular Ca
2+
levels,
NFATc1 is dephosphorylated, which allows NFATc1 to translocate
from the cytoplasm to the nucleus
. Translocated active
NFATc1 cooperates with other transcription factors to regulate
the expression of many osteoclast-specific genes such as cathepsin
K, osteoclast-associated receptor (OSCAR), and tartrate-resistant
acid phosphatase (TRAP)
.
We recently reported that the reduction of PP2A C
a
accelerates
bone formation and osteoblast differentiation through the expres-
sion of bone-related genes
. However, it remains unknown
whether reduction of PP2A C
a
expression in osteoblasts has an ef-
fect on osteoclast differentiation. The purpose of this study was to
examine whether PP2A C
a
expression in osteoblasts is involved in
osteoclast differentiation through RANKL/OPG expression and the
NFATc1 transcription factor.
2. Materials and methods
2.1. Materials
Alpha-modified Eagle’s minimal essential medium (
a
-MEM)
was purchased from Invitrogen (Carlsbad, CA). Plastic dishes were
obtained from IWAKI (Chiba, Japan) and fetal bovine serum (FBS)
0014-5793/$36.00 Ó 2012 Federation of European Biochemical Societies. Published by Elsevier B.V. All rights reserved.
http://dx.doi.org/10.1016/j.febslet.2012.10.041
⇑
Corresponding author. Fax: +81 88 633 7342.
E-mail address:
okamura@dent.tokushima-u.ac.jp
(H. Okamura).
FEBS Letters xxx (2012) xxx–xxx
j o u r n a l h o m e p a g e : w w w . F E B S L e t t e r s . o r g
Please cite this article in press as: Okamura, H., et al. Protein phosphatase 2A C
a
is involved in osteoclastogenesis by regulating RANKL and OPG expression
in osteoblasts. FEBS Lett. (2012),

was purchased from JRH Biosciences (Lenexa, KS). Antibodies
against RANKL (sc-9037) and OPG (sc-8468) were purchased from
Santa Cruz Biotechnology (Santa Cruz, CA), while anti-Gapdh anti-
body, ascorbic acid, b-glycerophosphate, Fast Red TR, and naphthol
AS-MX phosphate were all purchased from Sigma-Aldrich (St.
Louis, MO). The other materials used were of the highest grade
commercially available.
2.2. Cell culture and differentiation
PP2A knockdown (shPP2A) and control (shCont) osteoblastic
cells used in this study have been described previously
. Mouse
macrophage RAW264.7 cells were obtained from Riken Cell Bank
(Tsukuba, Japan). shPP2A and shCont cells were cultured in
a
-
MEM supplemented with 10% fetal bovine serum (FBS) at 37 °C un-
der a humidified atmosphere of 5% CO
2
. For each assay, the cells
were plated onto 100-mm culture dishes and cultured with differ-
entiation medium (supplemented with 50
l
M ascorbic acid and
2 mM b-glycerophosphate) for 7 days. The conditioned medium
was harvested, centrifuged to remove cell debris, filtered through
a 0.45-
l
m pore membrane filter (ADVANTEC, Tokyo, Japan), and
stored at 80 °C.
2.3. Real-time PCR analysis
Total RNA was extracted using Trizol, as recommended by the
manufacturer (Invitrogen). Real-time PCR of each gene was per-
formed in triplicate for at least three independent experiments.
Real-time PCR was performed with a 7300 Real-time PCR system
(Applied Bio-systems, Carlsbad, CA) using SYBR Premix Ex Taq™
(Takara Bio, Kyoto, Japan). The sequences of the primers were as
follows:
Mouse
Gapdh
(NM_008084):
(forward,
761)
5
0
-
TGTGTCCGTCGTCGTGGATCTGA-3
0
, (reverse, 910) 5
0
-TTGCTGTT-
GAAGTCGCAGGAG-3
0
; Mouse cathepsin K (NM_007802): (forward,
620)
5
0
-CAGCAGAACGGAGGCATTGA-3
0
,
(reverse,
703)
5
0
-
CTTTGCCGTGGCGTTATACATACA-3
0
; Mouse OSCAR (NM_175632):
(forward, 116) 5
0
-TGCATGCCGTGCTGACTTC-3
0
, (reverse, 223) 5
0
-
AAGGTCACGTTGATCCCAGGAG-3
0
.
2.4. ELISA
ELISA was performed according to the method described by
Hirota et al.
, with some modifications. Briefly, 50
l
l of the con-
ditioned media each from shCont and shPP2A cells were separately
dispensed into a polystyrene ELISA plate (Nunc Immuno Module,
Maxisorptype, Nalge Nunc, Rochester, NY). Subsequently, the
plates were dried completely at 37 °C. After blocking with 1.5% bo-
vine serum albumin in PBS for 1 h at room temperature (RT), these
plates were incubated with 50-
l
l total volume of 1:100 diluted
antibody for RANKL or OPG at RT for 1 h. Normal rabbit or goat
IgG was used as a negative control. After washing three times with
PBS, the plate was incubated with 1:500 diluted horseradish perox-
idase-labeled anti-rabbit (for RANKL) or -goat (for OPG) (Biosource
Co., Camarillo, CA) secondary antibodies for 1 h at RT. The plate
was washed 3 times with PBS and then incubated with peroxidase
substrate, 2,2
0
-azino-bis-(3-ethyl-benzthiazolin sulfonate) at RT
for 15 min. The reaction was measured spectrophotometrically at
405 nm.
2.5. Bone marrow cells
All mice studied were reared in our specific pathogen-free
mouse colony and given food and water ad libitum. Experiments
were humanely conducted under the regulation and permission
of the Animal Care and Use Committee of the University of Toku-
shima, Tokushima, Japan (toku-dobutsu 10051). Bone marrow-de-
rived cells from Balb/c mice (4–6 weeks of age) were incubated
with the conditioned medium from shCont and shPP2A cells. The
cultured cells were fixed and stained with TRAP staining solution.
TRAP-positive multinuclear cells containing more than three nuclei
were counted as osteoclasts.
2.6. Immunocytochemistry
Bone marrow macrophage cells were grown on sterile 18-mm
round glass cover slips placed in 60-mm plastic dishes. The cover
slips were incubated for 30 min with an anti-NFATc1 antibody
(BD Biosciences, San Jose, CA) diluted to 1:200 in the blocking solu-
tion. Then, they were incubated for 30 min with Alexa Fluor 488
goat anti-mouse IgG (Invitrogen) secondary antibody, diluted to
1:500 in 4% BSA. After incubation with 10
l
g/ml Hoechst 33342
for 30 min, the cover slips were mounted with PermaFluor aqueous
mounting medium (Lipshow, Pittsburgh, PA).
2.7. Statistical analysis
All data was expressed as mean ± SD, and a minimum of three
independent experiments were performed for each assay. Analysis
of variance (ANOVA) was used for statistical analysis. Statistical
significance was indicated with ‘‘
⁄
’’, and P values less than 0.01
were considered significant.
3. Results
3.1. Reduction of PP2A C
a
attenuates RANKL expression in osteoblasts
To examine whether PP2A C
a
expression plays a role in the
expressions of RANKL and OPG in osteoblasts, mRNA isolated from
shCont and shPP2A cells were subjected to real-time PCR using
specific primers for these transcripts (
A). The level of RANKL
was found to be significantly lower in shPP2A cells compared to
shCont cells, whereas OPG expression was significantly higher in
shPP2A cells compared to shCont cells. Consistent with the real-
time PCR data, the ratio of sRANKL/OPG in the cultured medium
of shPP2A cells was lower than that of shCont cells, as determined
using ELISA (
3.2. Conditioned medium of shPP2A cells does not induce NFATc1
translocation
During osteoclastogenesis, an activated form of NFATc1 is
known to translocate from the cytoplasm into the nucleus in osteo-
clast precursor cells. Therefore, we investigated whether the condi-
tioned medium from shCont and shPP2A cells could stimulate
NFATc1 translocation. Following a 1 h treatment of bone marrow
macrophage cells with either conditioned medium, the cells were
fixed, permeabilized, and stained using anti-NFATc1 antibody and
Hoechst 33342. We observed green fluorescent signal mostly in
the cytoplasm indicating that NFATc1 expression was localized to
the cytoplasm in untreated cells (
, upper panel). When we
treated bone marrow macrophage cells with the conditioned med-
ium from shCont cells, intense green fluorescence for NFATc1 was
observed in the nucleus (
, middle panel). In contrast, strong
fluorescence was not observed in the nucleus of the cells treated
with the conditioned medium from shPP2A cells (
, bottom pa-
nel). Nuclear staining with Hoechst 33342 was visualized as a red
pseudo-color. Thus, from a merged image, we observed a yellow
fluorescent signal restricted to the nucleus of cells treated with
the conditioned medium from shCont, but not shPP2A cells, thereby
confirming NFATc1 localization in the nucleus of these cells. Same
results were obtained from another experiment, in which we trea-
2
H. Okamura et al. / FEBS Letters xxx (2012) xxx–xxx
Please cite this article in press as: Okamura, H., et al. Protein phosphatase 2A C
a
is involved in osteoclastogenesis by regulating RANKL and OPG expression
in osteoblasts. FEBS Lett. (2012),

ted mouse macrophage cell line RAW264.7 with the conditioned
medium from shCont or shPP2A cells. NFATc1 translocation into
nucleus (yellow fluorescent signal) was observed in the cells trea-
ted with the conditioned media from shCont cells, but not shPP2A
cells (
B). Dephosphorylation of NFATc1 allows NFATc1 to
translocate and activate in the nucleus
. To further examine
phosphorylation status of NFATc1, the cell lysate was collected and
subjected to western blot analysis using NFATc1 specific antibody.
C shows that the conditioned media from shCont cells induced
dephosphorylation of NFATc1 proteins in RAW264.7 cells, which
can be followed by nuclear translocation. In contrast, phosphory-
lated status of NFATc1 (p-NFAT1c1) was still detected as around
120 kDa band in the RAW264.7 cells treated with the conditioned
medium from shPP2A cells.
3.3. Conditioned medium from shPP2A cells does not induce the
expression of NFATc1-related osteoclastogenesis marker genes
We next investigated whether translocalization of NFATc1 re-
sults in differential expression of NFATc1-related genes. Total
RNA was extracted from bone marrow cells treated with the con-
ditioned medium from shCont and shPP2A cells. Real-time PCR
was performed for NFATc1-related genes that are involved in
osteoclast differentiation.
shows that expressions of cathep-
sin K and OSCAR were significantly lower in bone marrow cells
treated with the conditioned medium from shPP2A cells com-
pared to those from shCont cells. The addition of recombinant
sRANKL (100 ng/ml) to the conditioned medium from shPP2A
cells
dramatically
restored
NFATc1
translocation
and
the
Relative gene expression / Gapdh
0
0.5
1
1.5
RANKL
OPG
0
0.5
1
1.5
*
shCont shPP2A
shCont shPP2A
*
shCont
shPP2A
0
0.5
1
sRANKL/OPG (ELISA)
*
shCont shPP2A
sRANKL OPG
A
B
Fig. 1. Expression levels of RANKL and OPG. (A) Total RNA was isolated from shCont or shPP2A cells and examined for RANKL, OPG, and Gapdh transcripts by real-time PCR.
Gene expression was normalized to Gapdh and plotted as the relative value compared to shCont cells (white bar). Each column represents the mean ± S.E.M.
⁄
P < 0.01. (B)
Cultured medium was collected from shCont or shPP2A cells and subjected to ELISA. The ratio of Soluble RANKL (sRANKL) to OPG is represented as the relative value
compared to shCont cells. Each column represents mean ± S.E.M. values.
⁄
P < 0.01.
NFATc1 Hoechst Merge
−
shCont
shPP2A
NFATc1
Hoechst Merge
shCont
shPP2A
p-NFATc1
NFATc1
β-actin
shCont
shPP2A
A
B
C
Fig. 2. Localization of NFATc1 in bone marrow macrophage cells. Bone marrow macrophage (A) and RAW264.7 (B) cells on coverslips were treated with the conditioned
medium collected from shCont and shPP2A cells. The cells were incubated with anti-NFATc1 antibody. NFATc1 was found to localize in the nuclei of cells treated with the
conditioned medium from shCont but not from shPP2A cells. Bar indicates 20
l
m. (C) Bone marrow macrophage cells were treated with the conditioned medium from either
shCont or shPP2A cells. Cell lysates from the cells were collected and subjected to western blots analysis. Immunoblots were developed by using specific antibodies for
NFATc1 and b-actin.
H. Okamura et al. / FEBS Letters xxx (2012) xxx–xxx
3
Please cite this article in press as: Okamura, H., et al. Protein phosphatase 2A C
a
is involved in osteoclastogenesis by regulating RANKL and OPG expression
in osteoblasts. FEBS Lett. (2012),
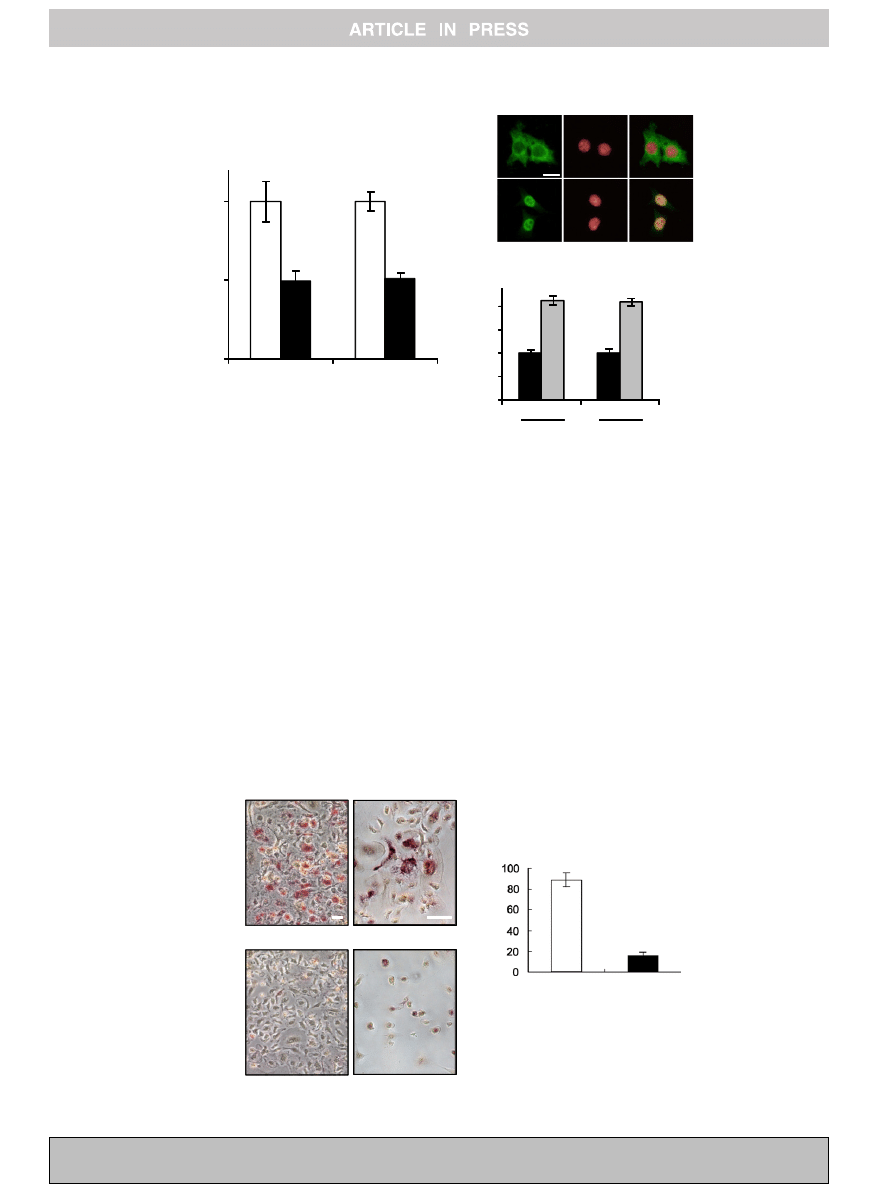
expressions of cathepsin K and OSCAR in bone marrow cells
(
B and C).
3.4. Conditioned medium from shPP2A cells does not induce TRAP-
positive multinuclear cells
Finally, to examine the effect of PP2A in osteoblasts to induce
TRAP-positive multinuclear cells, we treated bone marrow cells
with the conditioned medium from shCont cells or shPP2A cells.
Treatment with the conditioned medium from shCont cells in-
duced the formation of TRAP-positive multinuclear osteoclasts.
However, no effect was observed in the cells treated with the
conditioned medium from shPP2A cells (
B shows
the number of TRAP-positive multinuclear cells in each well of a
96-well plate. The number of TRAP-positive multinuclear cells in
the bone marrow cells treated with the conditioned medium from
shPP2A cells was significantly lower than that in the bone marrow
cells treated with the conditioned medium from shCont cells.
4. Discussion
Osteoblasts control bone metabolism not only by synthesizing
bone matrix proteins and regulating mineralization but also
by orchestrating the process of bone resorption through the
0
0.5
1
*
*
R
el
a
ti
ve
g
ene
e
x
p
re
ss
ion
/ G
a
p
d
h
cathepsin K
OSCAR
shCont shPP2A shCont shPP2A
A
0
0.5
1
1.5
2
*
*
R
e
la
ti
ve
g
e
ne
e
x
pr
e
ss
ion
/
G
a
pdh
ca thepsin K OSCAR
shPP2A shPP2A
sRANKL
−
+
−
+
B
c
NFATc1 Hoechst Merge
shPP2A
shPP2A
+sRANKL
Fig. 3. Expression levels of cathepsin K and OSCAR in osteoclast precursor cells. (A) Bone marrow macrophage cells were treated with the conditioned medium collected from
shCont or shPP2A cells for 24 h. Total RNA was isolated and subjected to real-time PCR by using specific primers for cathepsin K and OSCAR. Gene expression was normalized
to Gapdh and plotted as the relative value compared to shCont cells (white bar). Each column represents the mean ± S.E.M.
⁄
P < 0.01. (B) Bone marrow macrophage cells on
coverslips were treated with the conditioned medium collected from shPP2A cells with or without sRANKL (100 ng/ml) for 1 h. The cells were stained with anti-NFATc1
antibody and Hoechst 33342. NFATc1 was found to localize in the nuclei of cells treated with the conditioned medium from shPP2A cells with sRANKL. Bar indicates 20
l
m.
(C) Bone marrow macrophage cells were treated with the conditioned medium collected shPP2A cells with or without sRANKL (100 ng/ml) for 24 h. Total RNA was isolated
and subjected to real-time PCR by using specific primers for cathepsin K and OSCAR. Gene expression was normalized to Gapdh and plotted as the relative value compared to
shCont cells (white bar). Each column represents the mean ± S.E.M.
⁄
P < 0.01.
T
R
A
P
-p
o
sitiv
e
(c
ells
/w
ell)
shCont shPP2A
*
shCont
shPP2A
a
b
c
d
A
B
Fig. 4. TRAP-positive multinuclear cells induced by treatment with the conditioned medium. (A) Bone marrow macrophage cells were treated with the conditioned medium
derived from shCont (a, b) or shPP2A cells (c, d). The cells were fixed and stained for TRAP expression. High-power magnification images are shown in (b) and (d). (B) The
numbers of TRAP-positive multinuclear cells were counted. Bars indicate 50
l
m. Each column represents mean ± S.E.M. values.
⁄
P < 0.01.
4
H. Okamura et al. / FEBS Letters xxx (2012) xxx–xxx
Please cite this article in press as: Okamura, H., et al. Protein phosphatase 2A C
a
is involved in osteoclastogenesis by regulating RANKL and OPG expression
in osteoblasts. FEBS Lett. (2012),

regulation of osteoclastogenesis
. However, there has been
no information about the relationship between PP2A C
a
expression in osteoblasts and osteoclastogenesis. Therefore, we
investigated whether PP2A C
a
expression in osteoblasts can
play an important role in osteoclastogenesis by using PP2A
C
a
-knocked-down osteoblasts (shPP2A).
In this study, we demonstrated that silencing of PP2A C
a
in
osteoblasts decreased RANKL and increased OPG expression. In
agreement with our real-time PCR results, we also showed that
the sRANKL/OPG ratio in the culture medium was significantly
decreased in shPP2A cells compared to control cells (shCont). The
balance of the RANKL/OPG ratio is known to be crucial for bone
homeostasis
, since the ratio of RANKL/sRANKL to OPG pro-
duced by osteoblasts modulates osteoclast differentiation and
activity
. Increases in the ratio of RANKL/sRANKL to OPG
are expected to favor osteoclastogenesis, while decreases should
favor inhibition of osteoclastogenesis. Our present results indicate
that PP2A C
a
can regulate the expression and production of
RANKL/sRANKL and OPG, leading us to hypothesize that PP2A C
a
in the osteoblasts is involved in osteoclastogenesis.
RANKL/sRANKL-RANK signaling induces various transcription
factors, including NFATc1, which can act as a positive modulator
in osteoclast differentiation
. In this study, we have
shown that NFATc1 translocation and dephosphorylation were in-
duced by the treatment of the conditioned medium from shCont,
but not shPP2A cells. This result indicates that reduction of PP2A
C
a
in osteoblasts impairs its ability to induce NFATc1 dephospho-
rylation and translocation in osteoclast precursors.
NFATc1 induces the expression of target genes by binding to
NFAT-binding sites in the promoter region of genes such as TRAP,
cathepsin K, and OSCAR, which are important for osteoclast differ-
entiation or function
. A positive feedback loop exists during
late-stage osteoclastogenesis, in which NFATc1 induces OSCAR
expression leading to the activation of NFATc1
. Consistent
with previous reports, we showed that the conditioned medium
from shCont cells induced cathepsin K and OSCAR, following
NFATc1 translocation. In contrast, the conditioned medium of
shPP2A cells failed to induce the expression of these genes in bone
marrow macrophage cells. However, the addition of recombinant
sRANKL dramatically recovered NFATc1 translocation and the
expression of NFATc1-related genes in the cells treated with the
conditioned medium from shPP2A cells. These results indicates
that the reduction of the ratio of RANKL/sRANKL to OPG in shPP2A
cells results in impairment of NFATc1 translocation and the
expression of NFATc1-related genes. Moreover, the number of
TRAP-positive multinuclear cells induced by the conditioned med-
ium from shPP2A cells was significantly lower than that from
shCont cells. The same result was obtained when we co-cultured
bone marrow macrophage cells with shCont or shPP2A cells (data
not shown). These results indicate that reduction of PP2A in oste-
oblasts has a negative effect on osteoclastogenesis through the
down-regulation of NFATc1-related osteoclast marker genes.
In our previous study, administration of okadaic acid (OA), a
PP2A inhibitor, to the calvarial region in mice increased bone min-
eral density, mineral apposition, and bone thickness
. We con-
cluded in the previous report that reduction of PP2A activity in
osteoblasts by OA accelerates osteoblast differentiation, which in
turn leads to stimulate bone formation. In the present study, we
showed that inhibition of PP2A activity was involved in impair-
ment of osteoclastogenesis by regulating RANKL and OPG expres-
sion in osteoblasts. These observations may provide a new
explanation that the positive effect of OA on bone formation results
from an inhibitory effect on osteoclastogenesis. On the other hand,
it was reported that, in osteoclasts, the induction of PP2A by Toll-
like receptor 9 ligand blocks RANKL-induced osteoclast differenti-
ation through dephosphorylation of ERK and c-fos
. These
reports and our findings imply that PP2A exert complicated and
pleiotropic effects on osteoblastogenesis and osteoclastogenesis,
respectively. Therefore, further studies are needed to investigate
a detailed molecular mechanism in osteoblast- and osteoclast-dif-
ferentiation controlled by PP2A.
In conclusion, we demonstrated that reduction of PP2A de-
creased the ratio of RANKL/sRANKL to OPG both at the mRNA
and protein level in osteoblasts. Moreover, reduction of PP2A in
osteoblasts failed to induce NFATc1 activation and osteoclasto-
genesis. Our present study provides the first line of evidence that
PP2A C
a
expression in osteoblasts plays a pivotal role in osteo-
clastogenesis via the regulation of NFATc1-related gene expression.
Acknowledgment
Grant-in-Aid for Scientific Research from the Ministry of Educa-
tion (HO, 23592703).
References
[1] Suda, T., Takahashi, N., Udagawa, N., Jimi, E., Gillespie, M.T. and Martin, T.J.
(1999) Modulation of osteoclast differentiation and function by the new
members of the tumor necrosis factor receptor and ligand families. Endocr.
Rev. 20, 345–357.
[2] Komori, T. (2006) Regulation of osteoblast differentiation by transcription
factors. J. Cell. Biochem. 99, 1233–1239.
[3] Asagiri, M. and Takayanagi, H. (2007) The molecular understanding of
osteoclast differentiation. Bone 40, 251–264.
[4] Silva, I. and Braco, J.C. (2011) Rank/Rankl/opg: literature review. Acta
Reumatol. Port. 36, 209–218.
[5] Sims, N.A. and Walsh, N.C. (2012) Intercellular cross-talk among bone cells:
new factors and pathways. Curr. Osteoporos. Rep. 10, 109–117.
[6] Lacey, D.L., Boyle, W.J., Simonet, W.S., Kostenuik, P.J., Dougall, W.C., Sullivan,
J.K., Martin, J.S. and Dansey, R. (2012) Bench to bedside: elucidation of the
OPG–RANK–RANKL pathway and the development of denosumab. Nat. Rev.
Drug Discov. 11, 401–419.
[7] Boyce, B.F. and Xing, L. (2007) The RANKL/RANK/OPG pathway. Curr.
Osteoporos. Rep. 5, 98–104.
[8] Hofbauer, L.C., Khosla, S., Dunstan, C.R., Lacey, D.L., Boyle, W.J. and Riggs, B.L.
(2000) The role of osteoprotegerin and osteoprotegerin ligand in the paracrine
regulation of bone resorption. J. Bone Miner. Res. 15, 2–12.
[9] Fazzalari, N.L., Kuliwaba, J.S., Atkins, G.J., Forwood, M.R. and Findlay, D.M.
(2001) The ratio of messenger RNA levels of receptor activator of nuclear
kappaB ligand to osteoprotegerin correlates with bone remodeling indices in
normal human cancellous bone but not in osteoarthritis. J. Bone Miner. Res. 16,
1015–1027.
[10] Yao, Z., Matsuo, K., Nishimura, R., Xing, L. and Boyce, B.F. (2005) C-Fos/NFAT1
or 2-mediated osteoclastogenesis requires NF-
j
B p50/p52 expression. J. Bone
Miner. Res. (Suppl. 1), S145.
[11] Zhao, Q., Wang, X., Liu, Y., He, A. and Jia, R. (2010) NFATc1: functions and
osteoclasts. Int. J. Biochem. Cell Biol. 42, 576–579.
[12] Takayanagi, H., Kim, S., Koga, T., Nishina, H., Isshiki, M., Yoshida, H., Saiura, A.,
Isobe, M., Yokochi, T., Inoue, J., Wagner, E.F., Mak, T.W., Kodama, T. and
Taniguchi, T. (2002) Induction and activation of the transcription factor
NFATc1 (NFAT2) integrate RANKL signaling in terminal differentiation of
osteoclasts. Dev. Cell 3, 889–901.
[13] Zhang, L., McKenna, M.A., Said-Al-Naief, N., Wu, X., Feng, X. and McDonald,
J.M. (2005) Osteoclastogenesis: the role of calcium and calmodulin. Crit. Rev.
Eukaryot. Expr. 15, 1–13.
[14] Troen, B.R. (2006) The regulation of cathepsin K gene expression. Ann. N.Y.
Acad. Sci. 1068, 165–172.
[15] Kim, J.H., Kim, K., Jin, H.M., Youn, B.U., Song, I., Choi, H.S. and Kim, N. (2008)
Upstream stimulatory factors regulate OSCAR gene expression in RANKL-
mediated osteoclast differentiation. J. Mol. Biol. 383, 502–511.
[16] Kim, J.H., Kim, K., Youn, B.U., Jin, H.M. and Kim, N. (2010) MHC class II
transactivator negatively regulates RANKL-mediated osteoclast differentiation
by downregulating NFATc1 and OSCAR. Cell. Signal. 22, 1341–1349.
[17] Okamura, H., Yoshida, K., Ochiai, K. and Haneji, T. (2011) Reduction of protein
phosphatase 2A C
a
enhances bone formation and osteoblast differentiation
through the expression of bone-specific transcription factor osterix. Bone 49,
368–375.
[18] Hirota, K., Murakami, K., Nemoto, K., Ono, T., Matsuo, T., Kumon, H. and
Miyake, Y. (1998) Fosfomycin reduces CD15s-related antigen expression of
Streptococcus pyogenes. Antimicrob. Agents Chemother. 42, 1083–1087.
[19] Teitelbaum, S.L. (2000) Bone resorption by osteoclasts. Science 289, 1504–
1508.
[20] Aubin, J.E. (1998) Bone stem cells. J. Cell. Biochem. 30–31, 73–82.
[21] Boyce, B.F. and Xing, L. (2007) Biology of RANK, RANKL, and osteoprotegerin.
Arthritis Res. Ther. 9 (S1), 1–7.
H. Okamura et al. / FEBS Letters xxx (2012) xxx–xxx
5
Please cite this article in press as: Okamura, H., et al. Protein phosphatase 2A C
a
is involved in osteoclastogenesis by regulating RANKL and OPG expression
in osteoblasts. FEBS Lett. (2012),

[22] Evans, C.E., Mylchreest, S. and Andrew, J.G. (2006) Age of donor alters the
effect of cyclic hydrostatic pressure on production by human macrophages
and osteoblasts of sRANKL, OPG and RANK. BMC Musculoskelet. Disord. 7, 21.
[23] Karsenty, G. and Wagner, E.F. (2002) Reaching a genetic and molecular
understanding of skeletal development. Dev. Cell 2, 389–406.
[24] Matsumoto, M., Kogawa, M., Wada, S., Takayanagi, H., Tsujimoto, M.,
Katayama, S., Hisatake, K. and Nogi, Y. (2004) Essential role of p38 mitogen-
activated
protein
kinase
in
cathepsin
K
gene
expression
during
osteoclastogenesis through association of NFATc1 and PU.1. J. Biol. Chem.
279, 45969–45979.
[25] Kim, K., Kim, J.H., Lee, J., Jin, H.M., Lee, S.H., Fisher, D.E., Kook, H., Kim, K.K.,
Choi, Y. and Kim, N. (2005) Nuclear factor of activated T cells c1 induces
osteoclast-associated receptor gene expression during tumor necrosis
factor-related activation-induced cytokine-mediated osteoclastogenesis. J.
Biol. Chem. 280, 35209–35216.
[26] Kim, Y., Sato, K., Asagiri, M., Morita, I., Soma, K. and Takayanagi, H. (2005)
Contribution of nuclear factor of activated T cells c1 to the transcriptional
control of immunoreceptor osteoclast-associated receptor but not triggering
receptor expressed by myeloid cells-2 during osteoclastogenesis. J. Biol. Chem.
280, 32905–32913.
[27] Amcheslavsky, A. and Bar-Shavit, Z. (2006) Interleukin (IL)-12 mediates the
anti-osteoclastogenic activity of CpG-oligodeoxynucleotides. J. Cell. Physiol.
207, 244–250.
[28] Amcheslavsky, A. and Bar-Shavit, Z. (2007) Toll-like receptor 9 ligand blocks
osteoclast differentiation through induction of phosphatase. J. Bone Miner.
Res. 22, 1301–1310.
6
H. Okamura et al. / FEBS Letters xxx (2012) xxx–xxx
Please cite this article in press as: Okamura, H., et al. Protein phosphatase 2A C
a
is involved in osteoclastogenesis by regulating RANKL and OPG expression
in osteoblasts. FEBS Lett. (2012),
Document Outline
- Protein phosphatase 2A Cα is involved in osteocl
- 1 Introduction
- 2 Materials and methods
- 3 Results
- 3.1 Reduction of PP2A Cα attenuates RANKL expres
- 3.2 Conditioned medium of shPP2A cells does not induce NFATc1 translocation
- 3.3 Conditioned medium from shPP2A cells does not induce the expression of NFATc1-related osteoclastogenesis marker genes
- 3.4 Conditioned medium from shPP2A cells does not induce TRAP-positive multinuclear cells
- 4 Discussion
- Acknowledgment
- References
Wyszukiwarka
Podobne podstrony:
Molecular chaperones involved in heterologous protein expression in E coli
Merry Christmas is spoken in many languages around the world
The main press station is installed in the start shaft and?justed as to direction
How Chi Is Used In Dim Mak Pressure Point?fence
2 microorganisms involved in bioleaching
Stephani Hecht Hey There Is Fur in My Wedding Cake
Why the US is losing in Afghanistan A H Cordesman 1 Oct 08 doc
There is a hole in my bucket
Latour What Is Given in Experience
Sexual Attitudes and Activities in Women with Borderline Personality Disorder Involved in Romantic R
Why Affirmative Action in Higher Education is Safe in the Courts
Incest and lycanthropy Ferdinands impotency protecting his social rank and the violent ways control
H Juszka i in Sterowanie logiczne z regulacja PID
In Transit by Lillybellis
7lbs in 7 Days by Jason Vale
(ebook) Tillich, Paul Ultimate Concern Tillich in Dialogue by D Mackenzie Brown (existential phil
Famous Five 05 Five Go Off In A Caravan By Enid Blyton
The King in Yellow by Robert W Chambers
więcej podobnych podstron