
RUTH CHARLES
THE EXPLOITATION OF CARNIVORES AND OTHER FUR-
BEARING MAMMALS DURING THE NORTH-WESTERN
EUROPEAN LATE UPPER PALAEOLITHIC AND MESOLITHIC
Summary.
The exploitation of large mammals, particularly large herbi-
vores, has dominated perceptions of Upper Palaeolithic and Mesolithic
subsistence behaviour in north-western Europe. This paper critically reviews
the evidence for the exploitation of a complementary resource which has
received little attention within the archaeological literature — carnivores and
other fur-bearing mammals. Evidence for exploitation of individual species is
described and discussed. A model is then developed to explain the apparent
expansion of the subsistence base to include a wide range of fur-bearing
mammals during the Lateglacial and Mesolithic. This paper concludes by
arguing that although the use of carnivore meat and pelts cannot be viewed as
a dominant trend in European hunter-gatherer subsistence practices, their
contribution to hunter-gatherer economies cannot be ignored.
The reconstruction of European Palaeo-
lithic and Mesolithic economic activities has
been a primary concern of researchers over
the last 150 years. Relatively little attention
has been paid to evidence for the exploitation
of carnivores and other fur-bearing mammals,
especially those which do not normally form a
part of modern western European dietary
practices. However, detailed examination of
faunal assemblages from a series of Late
Upper Palaeolithic and early Mesolithic sites
in north-western Europe indicates that such
mammals were exploited both for their meat
and pelts. The material discussed here origi-
nates from archaeological sites dating to the
Lateglacial and early Postglacial and asso-
ciated with Creswellian, Magdalenian, Ma-
glemosian
and
Ertebølle
archaeological
residues (Fig. 1). In most cases the sites
discussed are those currently under review by
the author (Gough’s Cave, the Robin Hood
Cave, Trou de Chaleux, Trou des Nutons,
Trou du Frontal, Grotte de Sy Verlaine, Grotte
du Cole´opte`re, and Star Carr), although
additional examples have been drawn from
the published literature. In the former in-
stances faunal assemblages have been exam-
ined with the specific objective of identifying
direct evidence for the human exploitation of
animals via the evidence of butchery marks on
bones.
Recent research on the Danish Mesolithic,
particularly the Ertebølle, has led the way in
documenting the exploitation of such mam-
mals (Rowley-Conwy 1980 & in press;
Trolle-Lassen 1986 & 1987). The exploitation
OXFORD JOURNAL OF ARCHAEOLOGY 16(3) 1997
ß Blackwell Publishers Ltd. 1997, 108 Cowley Road, Oxford OX4 1JF, UK
and 350 Main Street, Malden, MA 02148, USA.
253
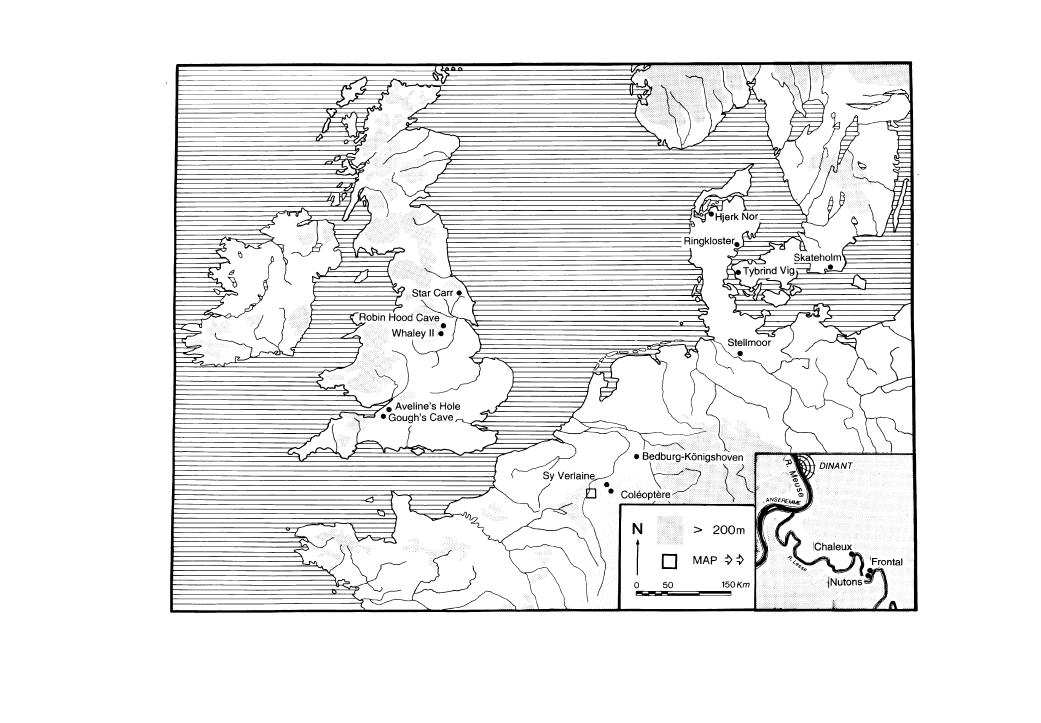
Figure 1
Location map of sites discussed within the text.
THE
EXPLOITATION
OF
CARNIVORES
AND
OTHER
FUR-BEARING
MAMMALS
OXFORD
JOURNAL
OF
ARCHAEOLOGY
254
ß
Blackwell
Publishers
Ltd.
1997

of one particular fur-bearing species (Lepus
timidus) has also been reported during the
British Lateglacial (Charles & Jacobi 1994).
Recent work indicates that the small scale
exploitation of such mammals was more
widespread during the Late Upper Palaeo-
lithic and early Mesolithic. However, in most
of the assemblages discussed here there is
also abundant evidence for the human ex-
ploitation of a number of ungulate species
which formed the primary subsistence base at
these sites, and which have dominated
perceptions of the hunting/trapping activities
which took place.
In contemporary western European society
carnivores are rarely, if ever, viewed as a
potential food source, and only a limited
range of fur-bearing mammals are exploited
for their pelts. However, there is no reason
why these preferences should be reflected in
the European prehistoric archaeological re-
cord. Indeed, the review that follows demon-
strates that whilst a range of carnivore
species may not have been a regular dietary
staple, they were on occasion exploited both
for their meat and pelts. Modern poachers
and countrymen questioned in the Oxford
region suggested that carnivore meat is
avoided today because it ‘tastes bad’, how-
ever, none of these informants had actually
tried carnivore meat, believing the nature of
the prohibition to be self evident. Contem-
porary non-European societies rarely main-
tain dietary taboos similar to those prevalent
in Europe, and the use of dogs, cats and a
variety of rodents as food can be documented
in many parts of the world. Similarly a
variety of rodents and other mammals rarely
considered edible by modern Europeans were
enjoyed by both ancient Romans and Greeks.
Grahame Clark’s seminal volume Prehis-
toric Europe: the economic basis (1952) first
drew to prominence the potential contribu-
tion of small game animals to Palaeolithic
and Mesolithic communities. However, there
are relatively few detailed discussions of the
archaeological evidence for this activity.
What follows is a review of the evidence
for the exploitation of a variety of carnivores
and fur-bearing mammals from the Late
Upper Palaeolithic and early Mesolithic of
north-western Europe (used here to mean
Britain, Benelux, Germany and southern
Scandinavia). Each species will be dealt with
individually and the nature of the evidence
discussed in detail.
URSIDAE
Brown bear (Ursus arctos)
In general, it is uncommon to find cut-
marked bear bones from Late Upper Palaeo-
lithic and early Mesolithic contexts, although
bears are frequently present in faunal assem-
blages. It is usually assumed that these
predators were denning in caves at times
when humans were absent. There are few
convincing accounts of the human exploita-
tion of bears in the archaeological literature.
Suggestions that Middle Palaeolithic sites
such as Ista´llo´sko¨ and Szeleta belonged to a
‘cave bear cult’ in south-eastern Europe are
dubious. Instead it seems probable that the
bear remains found in these caves came from
animals which died naturally during hiberna-
tion and that bears were the main residents of
these sites for much of Middle Palaeolithic
time (Allsworth-Jones, 1986) rather than
there being any direct link with humans.
Re-examination of such ‘cave-bear cult’
faunas for direct butchery evidence may
prove to be a useful path for future research,
offering an approach which could demon-
strate whether humans had contributed in any
way to the accumulation of these bear
remains.
The paucity of clear evidence of bear
RUTH CHARLES
OXFORD JOURNAL OF ARCHAEOLOGY
ß Blackwell Publishers Ltd. 1997
255

exploitation
during
the
Pleistocene
and
Holocene may be explained by bears’
tendency to avoid direct contact with hu-
mans, unless they pose a threat, and their
choice of dens in relatively secluded areas.
The hunting of bears is a highly dangerous
pastime, even with modern guns and rifles.
Modern weapons have to be of a large
calibre, and often a number of shots are
necessary to kill an adult (Nelson 1973, 122–
124; Domico 1988, 170), a luxury rarely
available whilst being charged by a bear. The
use of snares as a means of trapping bears has
been documented by Nelson (1973, 116–
117), as have a range of techniques for
hunting bears in their dens. Almost all
require the use of projectiles, and it is
assumed that Late Pleistocene and early
Holocene bear exploitation
would have
involved the use of an efficient projectile
technology and/or the opportunity to take
bears by surprise either via traps or when
found hibernating.
Brown bears are present in the Late
Magdalenian faunas from the Trou de
Chaleux (Hulsonniaux), the Trou du Nutons
(Furfooz), the Trou du Frontal (Furfooz) and
the Grotte de Sy Verlaine (Hamoir) in
Belgium. They are also present in the
Creswellian-associated faunas from Gough’s
Cave (Cheddar) and the Robin Hood Cave
(Creswell) in Britain. Of these, the only
specimens which can be directly linked to
human activity are those recovered from the
Trou de Chaleux in Belgium.
The number of identifiable specimens
present (NISP) count for bear at Chaleux is
relatively low (65; 1.78% of the overall
assemblage; full details of assemblage com-
position are given in Charles 1994), and the
body part representation data indicates that
only selected parts of three bear carcasses
were recovered. The remains of at least two
adults and one juvenile were present. In the
case of the juvenile only 3 metapodia with
unfused proximal epiphyses were recognised,
suggesting the presence of an isolated paw
rather than an entire skeleton. Both adult
bears are represented by parts of the skull
(mandibles, a maxilla and loose teeth), three
vertebrae, an innominate, sacrum, part of the
forelimb, the lower portion of the hind limb
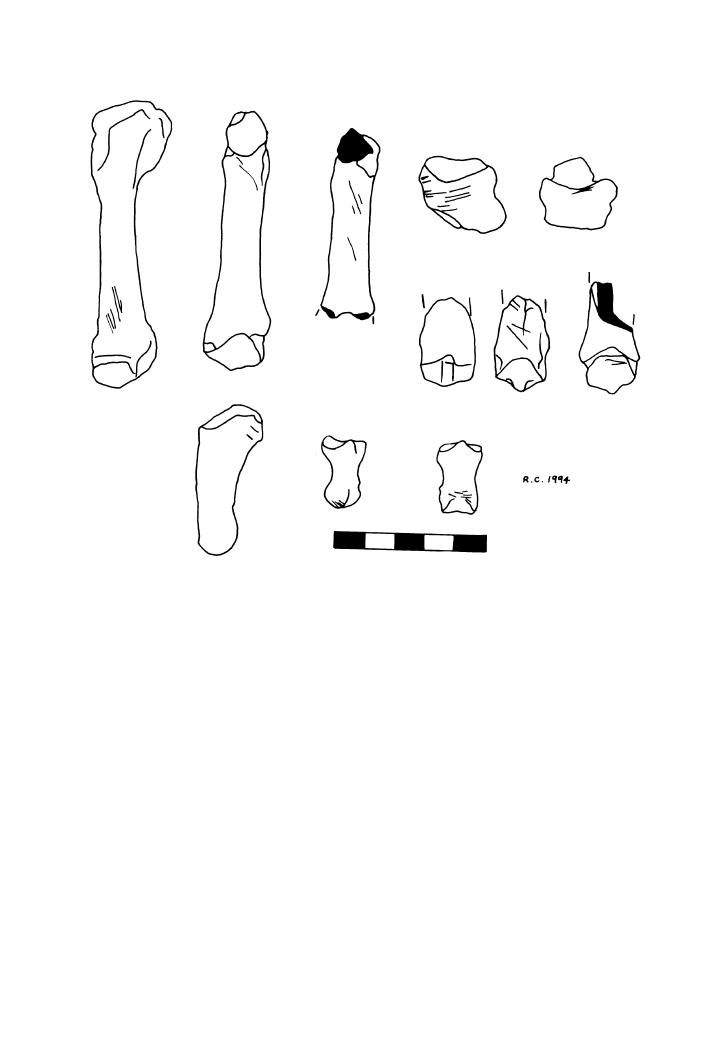
and the phalanges. Butchery marks were
observed on the bones of at least two of the
three individuals (an adult and the juvenile;
Fig. 2). With the exception of a left mandible,
all the butchery marks were confined to the
paws and for the most part correspond with
skinning, although marks across two tarsals
and three phalanges are indicative of dis-
articulation. Amongst these are three meta-
tarsals belonging to the adult (all cut) which
were originally part of the same paw.
Brown bear dens are usually selected for
their remoteness from centres of human
population, and may be in long-term use by
successive generations of bears. Depending
on environmental conditions, bears will begin
hibernation between September and early
November and continue until the April of the
following year. Cubs may den with their
mothers for up to two winters and this may
well be the situation with at least one adult
and the juvenile from Chaleux. Consequently
the taking of the two bears from Chaleux
could indicate autumn or winter seasonality
for this particular ‘event’ at the site. Before
hibernation commences, bears lay down an
large amount of fat to see them through the
winter months. Both Domico (1988) and
Partridge (1992) give the figure of 90 lb
(41 kg) as an upper limit for food consump-
tion within a 24 hour period. Nelson’s
account of the north American Kutchin
Indians (1973, 115–116) noted that there
were only certain periods of the year during
which these animals were considered ‘fit’ for
eating, primarily when they had large fat
THE EXPLOITATION OF CARNIVORES AND OTHER FUR-BEARING MAMMALS
OXFORD JOURNAL OF ARCHAEOLOGY
256
ß Blackwell Publishers Ltd. 1997
reserves (autumn and early winter), although
they were taken during other parts of the year
for their skins. From a human perspective,
bears have many uses. Their fat may well
have been a valuable resource for the human
residents of the cave. Their skins are tough
and hard wearing, and their stored fat is
known to make good fuel for lamps, has a
high calorific value and may also be
extremely useful in the waterproofing of
skins. Coles (1991, 136) notes that bear
grease was a good adhesive agent for the
pigments used in prehistoric art (although
there is no evidence for cave painting at
Chaleux or any of the Belgian caves). Their
hides are at their best between the autumn
and spring, and have been observed being
used for mattresses and for insulation among
contemporary north American Indians (Nel-
son 1973, 127).
Evidence for the Mesolithic exploitation of
brown bears is equally rare, but does exist. At
Medvedia Cave near Ruzı´n in Slovakia, the
skeletal remains of at least one bear were
found associated with composite arrowheads
(bone points with imbedded microliths, Ba´rta
1989). For the time being, the bears from
Chaleux and Medvedia Cave remain the only
unambiguous examples of the Late Pleisto-
cene and early Holocene human exploitation
Figure 2
Cut-marked bear bones from the Trou de Chaleux.
RUTH CHARLES
OXFORD JOURNAL OF ARCHAEOLOGY
ß Blackwell Publishers Ltd. 1997
257

of brown bear.
CANIDAE
Wolf (Canis lupus) and dog (Canis familiaris)
Wolves are the largest wild canids, weigh-
ing between 16 and 60 kg. They have been
known to live for up to 13 years and have a
well defined mating season between January
and April. Originally the wolf had one of the
widest distributions of any known mammal,
being distributed throughout the northern
hemisphere north of 15º lattitude. Their
habitat tolerance is extremely variable, po-
pulation densities being proportional to those
of their prey. Generally they live in family
groups (packs) with complex social organisa-
tion (Mech 1970), pack size is also variable,
upper limits being 30 individual although
groups of eight to twelve are more common
(Ginsberg & MacDonald 1990). Flexibility
appears to be the main aspects of wolf
behaviour, and this is reflected in their diets,
which focus on large ungulates when avail-
able, but will also include carrion, smaller
prey and rubbish discarded by humans.
Wolf and/or dog remains have been
recorded at many of the sites under discus-
sion here. Canids were recovered from the
Lateglacial layers at the Trou de Chaleux,
Trou des Nutons, Trou du Frontal, Grotte de
Sy Verlaine, Gough’s Cave and the Robin
Hood Cave; the domestic dog from Star Carr
is well known in the literature (Degerbøl
1961). Although the presence of semi-feral
dogs during the northern European Upper
Palaeolithic has been suggested by Benecke
(1987), this has yet to be demonstrated at any
of the Lateglacial sites listed here, and all
canids recovered from the Pleistocene layers
at these sites are identified as Canis lupus. At
both the Trou des Nutons in Belgium and
Gough’s Cave in Britain cut-marks are
present on the wolf remains.
The actual number of specimens showing
butchery evidence is low. A single distal
humerus from the Trou des Nutons shows
multiple parallel cuts along the medial aspect
of the distal end and shaft which correspond
with both meat removal and disarticulation.
The only cut specimens from Gough’s Cave
are two phalanges (Natural History Museum
Acc. No. M.13796), with transverse cuts
present across the palmate surfaces, relating
to skinning. A further example is given by
Rowley-Conwy (in press) from Ringkloster,
where four dog crania were recovered
showing cut marks relating to skinning. In
the cases of Gough’s Cave and Ringkloster it
is clear that the animals in question were
processed for their pelts, and certainly for
their meat at the Trou des Nutons. Addition-
ally, canids are present within the assem-
blages from Sy Verlaine (Minimum Number
of Individuals = one), the Trou du Frontal
(MNI = three) and the Trou de Chaleux (MNI
= two). None of these latter specimens was
cut, and the possibility that they represent
‘natural deaths’ within the cave cannot be
excluded.
Red fox (Vulpes vulpes) and arctic fox
(Alopex lagopus)
It is often unclear whether one or two
species of fox are present in European Late
Pleistocene faunal assemblages, although the
red fox (Vulpes vulpes) is usually assumed to
be the only fox present in early Holocene
ones. Clutton-Brock et al. (1976) identified
various point of difference between different
members of the Canidae, but few of these are
applicable to post-cranial elements or in-
complete crania. In general, modern arctic
foxes are smaller than red foxes, one of the
most distinctive differences being that their
muzzles are relatively shorter and broader
THE EXPLOITATION OF CARNIVORES AND OTHER FUR-BEARING MAMMALS
OXFORD JOURNAL OF ARCHAEOLOGY
258
ß Blackwell Publishers Ltd. 1997

than their red counterparts. It is possible to
differentiate these two species osteologically,
but this is only possible on certain anatomical
elements, and requires them to be complete.
One of the main points of differentiation is
within the dental arcade; however, this is
rarely complete in Pleistocene specimens.
Both red and arctic foxes live in dens with
complex burrow systems (Ginsberg & Mac-
Donald 1990). These are used primarily for
shelter and rearing young. In the case of
arctic foxes there is evidence to suggest that
these dens may be in long term use by
succeeding generations. The average life
span of a fox is a few years (11 is given as
a maximum: op. cit., 34), although a den may
be in continual use for up to 300 years.
The arctic fox (Alopex lagopus) occurs in
two colour morphs (‘white’ and ‘blue’),
which change seasonally. Its habitat is today
limited to circumpolar regions, especially
arctic tundra. It is an opportunistic scavenger
and predator, and has been known to
scavenge meat from seal carcasses (op. cit.,
34) and to take seabird eggs.
The colouring of the red fox is quite
variable but not seasonal. Animals have been
observed to have pelts in a range of shades
from grey to bright red, although the most
common is reddish-brown. Red foxes are
distributed throughout the northern hemi-
sphere and much of southern Australia. Their
natural habitat is a dry, mixed landscape,
although they are also found in uplands,
mountains, deserts and sand dunes. In short,
they are a highly flexible and adaptable
species. Their diet is varied, including
beetles, earthworms, small mammals (includ-
ing lagomorphs), birds, fruit and carrion.
They are often blamed for the death of
juvenile livestock (e.g. lambs), although it is
unclear whether they actively hunt these or
taken them as carrion. They are also known
to cache food surpluses (Ginsberg & Mac-
Donald 1990).
Both species of fox are currently trapped
for their pelts in various parts of the world.
The trapping of arctic foxes for their pelts is a
relatively well documented activity during
the Upper Palaeolithic of other regions (cf.
Klein 1973, 56). The evidence from the
Russian sites of Mezin and Avdeevo, consists
of virtually complete fox skeletons (exclud-
ing paws), which Klein suggests were
removed with the skins.
The skinning of animals leaves very few
traces in the archaeological record. Cut
marks may be present at key points of
severance — in the region of the snout, and
perhaps on the lower limbs, where the skin
has been ‘ringed’ to remove it from the body.
The precise location of both points of
severance will vary. This variation may relate
to a wide range of factors, including the
particular butchery ‘tradition’ and techniques
involved, as well as personal ‘style’ on the
part of the butcher/furrier. This being the
case, the recognition of human exploitation
of foxes solely for their pelts via butchery
evidence is likely to be rare — and greater
credence is often placed on body part
representation
information
when
sample
sizes are adequate. However, this does not
preclude the presence of direct butchery
evidence, and where this exists it can only
add credibility to arguments for the exploita-
tion of these fur bearing mammals.
Such evidence is present at many of the
sites under discussion here (the Trou de
Chaleux, Trou des Nutons, the Trou du
Frontal and Star Carr); although only the
Trou de Chaleux yielded sufficiently high
numbers of fox remains to make body part
representation analyses a worthwhile exer-
cise. It is tempting to argue that the fox
remains found at the other sites (the Grotte
du Cole´opte`re, the Grotte de Sy Verlaine,
Gough’s Cave and the Robin Hood Cave)
RUTH CHARLES
OXFORD JOURNAL OF ARCHAEOLOGY
ß Blackwell Publishers Ltd. 1997
259

were also present due to human agency.
However, there is no direct evidence for this,
and in each case the NISP counts for Vulpes
sp. were so low that the remains could easily
be accounted for by one or two individuals
denning in the caves at a later date.
At Chaleux, the foxes were the second
most abundant species (NISP = 473, 12.93%
of the assemblage). Consequently their po-
tential significance within the local Lategla-
cial hunting/trapping economy should not be
underestimated. Of the five fox bones with
butchery marks, one — a left scapula — is
clearly very recent in origin. The remaining
four specimens (two femora, a humerus and a
scapula) show longitudinal marks consistent
with meat removal. It is difficult to establish
whether more (or, indeed all) the foxes at
Chaleux were present due solely to human
agency, or to the actions of several predators
of which humans were just one. As the fox
bones lack any spatial information (as indeed
does all the faunal material from Chaleux)
and the excavator, E
´ douard Dupont, made no
detailed comment on them, it is impossible to
know whether the fox bones were encoun-
tered as disarticulated elements, in articulated
units or as complete skeletons.
An MNI count of 17 was obtained from the
mandibles. Examination of body-part repre-
sentation (Table 1) does not give a clear
indication as to whether complete or almost
complete carcasses were present at the site.
No juvenile bones were recorded and,
although this may indicate that the cave
may not have been a den, this possibility
cannot be fully eliminated. It is interesting
that the first cervical vertebrae (atlas and
axis) are present in identical numbers (eight),
although the % MNI figures for the other
vertebrae are much reduced. Complete long
bones are relatively rare, although they are
present. Proximal and distal ends occur in
significantly different quantities, although
this seems to be linked more with their
relative robustness than with human selection
of different body parts. Fibulae are totally
absent which, given the fragile nature of this
bone, may again be taken as an indication
that bones were fragmented beyond recogni-
tion (or recovery) whilst buried.
Of perhaps greatest note is the relative lack
of metapodia and phalanges. The metapodia
occur only in very low frequencies, as do the
1st phalanges, whilst the 2nd and 3rd are
totally absent. There are three possible
explanations for this: first, that these ele-
ments were not recovered on site due to poor
excavation techniques; second, that these
elements were not identified after recovery;
and finally that they were not originally
present at the site. The first two possibilities
do not seem to be particularly likely and as
recovery from this excavation appear to have
been excellent (Charles 1994), it is assumed
here that the BPR information is a reflection
of what was originally present at the site
rather than a distortion caused by poor
recovery techniques.
Klein’s (1973) account of skinned foxes
from Russian sites seems to hint at the fate of
at least some of the Chaleux foxes. Like their
Russian counterparts, the Chaleux foxes have
lost their paws, a fact highly suggestive of
skinning. Animal remains from Chaleux are
present in high numbers, and the occurrence
of the smaller anatomical elements of most
species in relatively high proportions indi-
cates that this patterning is not due to poor
excavation recovery. It is also worth noting
that only one fox cranium and three maxillae
were present in the collections, in very
noticeable contrast with the number of fox
mandibles, again suggesting that anatomical
elements likely to be attached to the pelts had
been removed from the site.
Despite the circumstantial evidence for
skinning and trapping activities at or near
THE EXPLOITATION OF CARNIVORES AND OTHER FUR-BEARING MAMMALS
OXFORD JOURNAL OF ARCHAEOLOGY
260
ß Blackwell Publishers Ltd. 1997

TABLE 1
Body Part Representation data for fox (Vulpes sp.) from the Trou de Chaleux
RUTH CHARLES
OXFORD JOURNAL OF ARCHAEOLOGY
ß Blackwell Publishers Ltd. 1997
261

Chaleux, very few of the fox bones show any
direct butchery marks. The cut marks on five
fox bones from Chaleux do not indicate
skinning, but rather meat removal (humerus
and femora) and disarticulation (scapula).
Similarly the 3 cut fox bones from the Trou
des Nutons (a left mandible and two distal
right humeri) all indicate meat removal, as do
the marks noted on humeri from the Trou du
Frontal. The red fox bones from Star Carr
come from one individual (Fraser & King
1954, 73) and cut marks present on a phalanx
(Natural
History
Museum
Acc.
No
1958.11.10.43) are consistent with skinning
whereas marks noted on ribs, lumbar verteb-
rae, a humerus, femora and the tibii all
indicate meat removal. Additionally, Row-
ley-Conwy (in press) reports the presence of
a fox skull with transverse cuts across its
front from Ringkloster, relating to skinning.
Overall the evidence for the exploitation of
foxes at these sites is limited, suggesting that
although on occasion foxes were exploited
for both pelts and meat during the Late
Pleistocene and early Holocene this was not a
mainstay of the local economy. Today foxes
are considered vermin, and both vermin and
carnivores are not generally thought of as a
viable food resource in the way that rabbit or
hares are. Whether any such cultural distinc-
tion was made during the Pleistocene is
unclear, although it is worth noting that
within Klein’s sites in Russia the foxes do not
appear to have been used for food, but
instead discarded as complete carcasses.
It seems quite possible that, as well as
being the prey of humans, one or both species
of fox were resident in the Lesse valley caves
during the late Pleistocene and Holocene.
Their dens may be used over relatively long
time periods, and during such periods it is
likely that a selection of vertebrates, inverte-
brates and other food substances will have
been brought by them to the cave, accounting
for components of the faunal assemblage.
Similarly at various times the foxes them-
selves may have been the prey of other
carnivores living in this area. At Star Carr the
picture is somewhat different, with an
isolated red fox carcass brought to the site
and processed for its meat and pelt.
Whilst foxes cannot be viewed a primary
subsistence sources at any of these sites, it is
clear that they did have a role to play in the
subsistence economy of the hunter-gatherers
who used these places, and that they were
exploited for both their furs and their meat.
MUSTELIDAE
Wolverine (Gulo gulo)
Wolverines are today mainly restricted to
the taiga and southern tundra zones of
Eurasia, including parts of the old Soviet
Union, Norway, Sweden, Finland, Mongolia,
northern China, and also extend into the new
world in much of Canada, Alaska and
possibly the Rocky mountains. However, this
is by no means a reflection of their past
distribution. They are highly adaptable ani-
mals, and records from the earlier parts of
this century, as well as the last, suggest that
they were found in more southerly regions in
the recent past (Schreiber et al. 1989, 22–23).
They have extremely large home ranges,
approx. 800–900 km
2
in summer, and per-
haps even larger in winter (ibid., 22). The
wolverine is a predator, capable of taking
large mammals, although in general it is
mainly a scavenger of herbivore carcasses up
to the size of reindeer and red deer. Modern
wolverines are up to 1 metre in length, and
their pelts are dark brown (sometimes almost
black), with a pair of yellowish stripes
running from the shoulders to the rump. In
general, wolverines tend to avoid human
settlements, and to date there is little research
THE EXPLOITATION OF CARNIVORES AND OTHER FUR-BEARING MAMMALS
OXFORD JOURNAL OF ARCHAEOLOGY
262
ß Blackwell Publishers Ltd. 1997

available on their ecology and behaviour
patterns. It seems unlikely that wolverines
came into frequent contact with humans
during the Palaeolithic, and their vicious
reputation no doubt preceded them. Indeed,
wolverine remains are rarely found on
western European Upper Palaeolithic and
Mesolithic sites, and this species has only
been identified at two of the sites considered
here, the Trou de Chaleux and the Trou des
Nutons. Only two specimens (one from
Chaleux and one from the Trou des Nutons)
hint at an association with human activity
(see below).
In many, if not all, of the regions where it
is found today, the wolverine is traditionally
hunted for its fur. Like many animals, they
frequently follow set routes and setting traps
in ‘natural corridors’ such as ravines is
common practice among recent hunters
(Nelson 1973). As wolverines are also one
of the main scavengers from traplines,
wolverine traps are today frequently set near
the ends of trap lines to diminish predation
from other traps. Similarly, kill sites for other
large mammals provide excellent locations to
set traps for wolverines (as well as foxes and
wolves) as the smell attracts scavenging
animals. Because of the wolverine’s strength
and violent nature it is a difficult animal to
keep restrained and extra strong traps and
snares need to be used, although deadfall
traps are also highly effective.
Their fur is thought (R. Jacobi pers. comm.)
to be especially useful in extremely cold
environments, as it does not freeze when
damp, and consequently makes a fine edging
for hoods, amongst other uses. The specimen
from Chaleux is a partial scapula, with
numerous marks which may have been the
result of butchery activities across its posterior
surface. However, the true origin of these
marks could not be determined by micro-
scopic inspection. Examination of the speci-
men by Jill Cook of the British Museum
revealed that the most prominent marks
closely matched marks of natural origin found
on the remains of a cow from Draycott
(Andrews & Cook 1985). Others, less promi-
nent, could only be identified as possible cuts.
The other specimen, a proximal right femur
from the Trou des Nutons had a single,
somewhat ambiguous mark, which, as with
its counterpart from Chaleux, could neither be
confirmed or denied as a cut. if the marks from
both specimens were indeed the result of
butchery activity, then they would correspond
with meat filleting rather than skinning,
although the one would not preclude the
other. However, given the questionable nature
of the marks from these specimens, it remains
unclear whether the wolverine ever formed a
component in any stone age meal in the Lesse
Valley. Similarly it is unclear as to whether
these specimens are parts of the Late Magda-
lenian faunal assemblages, or a more recent
additions. Both the range of species present
alongside recent radiocarbon dating has
indicated that faunal material was accumu-
lated within the same geological units at the
Trou de Chaleux and the Trou des Nutons
during the Lateglacial and the early Post-
glacial (Charles 1994; Rosine Orban pers.
comm.). However, wolverines were certainly
known to Magdalenian hunter-gatherers; one
is engraved on a perforated baˆton from La
Madeleine (Cremades 1992). They have been
identified from other Lateglacial contexts in
north-western Europe such as Chelms Combe
at Cheddar (Andrew Currant pers. comm.) and
Petersfels (Albrecht et al. 1983, 78) and are
thought to have been present in north-western
Europe during the early Postglacial.
Badger (Meles meles)
Present knowledge is extremely patchy
about the temporal and spatial distribution of
RUTH CHARLES
OXFORD JOURNAL OF ARCHAEOLOGY
ß Blackwell Publishers Ltd. 1997
263

badgers during the Holocene and Pleistocene
in western Europe. Their apparent presence
in Late Pleistocene assemblages is usually
taken as evidence for more recent contam-
ination. However, badger fossils have been
recorded from genuine Late Pleistocene and
early Holocene contexts in north-western
Europe, including Petersfels (Albrecht et al.
1983, 80) and Kendrick’s Cave (from which
two perforated badger teeth have been used
as pendants, Roger Jacobi pers. comm.).
Badgers are denning animals and live in
sets; the morphology of these is highly
variable, but they may be extremely large
covering many cubic metres. The tunnels
tend to be linked, and there are many
chambers, often with bedding of vegetation.
Badgers also ‘like’ using caves (Hans Kruuk
pers. comm.). Sets are often shared with
other carnivores, including foxes, otters and
wild cats. These sets are thought to be in long
term use, perhaps being used over centuries.
Indeed, one near Kirkhead Cavern in Cum-
bria is mentioned in the Doomsday Book and
is still in use. It is unclear whether these sets
are used continuously, or episodically by
different badger clans. Although in general
badgers prefer hilly districts, especially
heavily wooded ones (Neal 1948, 122), they
are highly adaptable, and can survive in a
range of habitats and soils. Many of the
Lateglacial caves discussed here, with steep
talus cones, surrounding slopes and proxi-
mity to water, would have proved ideal
locations for badgers.
It is possible that badgers may have played
some part in the accumulation of the faunal
assemblage found in the cave sites. Although
their diet is primarily made up of earthworms
(Kruuk 1989, 39), they are known occasion-
ally to eat a variety of foodstuffs including
salmon, frogs and toads, wood pigeon,
passerines, a variety of small mammals
including rabbits and hares, and carrion up
to the size of sheep and deer. They have even
been observed to kill lambs (Kruuk 1989,
Plate 11) and to take chunks of food,
including carrion, back to cubs in the sets.
In recent times omnivorous tendencies have
been observed amongst badgers in areas
away from extensive agriculture, where they
take carrion, small mammals and vegetable
matter. The cubs are prone to high mortality
rates within the sets, but this varies with local
ecological conditions (Hans Kruuk pers.
comm.). Kruuk has observed corpses of very
young badgers well away from sets, which
must have been taken there by adults, but
thinks that most will be either left inside the
set or eaten by adults. Similarly Kruuk has
noted that ‘small mammal remains can be
passed relatively undamaged in the faeces,
and their latrines inside the set or, in the
cave-based sets in an ‘entrance hall’ (Hans
Kruuk pers. comm.). One particularly inter-
esting feature is that badgers do not chew or
gnaw bones, so it is difficult actually to
demonstrate a direct link between badger
activity and the accumulation of faunal
remains. Given what has been outlined
above, badgers could have been responsible
for at least part of the accumulation of small
mammals and fish bones and even some of
the larger mammal bones, at many of the
Lateglacial caves discussed here, although
this cannot be conclusively demonstrated.
Direct butchery evidence was observed on
badger bones from the Trou de Chaleux,
Trou des Nutons, Gough’s Cave and Star
Carr. In each case only a small number of cut
marked bones were present, and the marks
indicate different processing activities at
different sites (Fig. 3).
An MNI of four was calculated for the
Chaleux badgers, based on the innominates,
scapulae and femora. No juvenile bones were
recorded. In total 66 badger bones were
present (1.8% of the assemblage). Although
THE EXPLOITATION OF CARNIVORES AND OTHER FUR-BEARING MAMMALS
OXFORD JOURNAL OF ARCHAEOLOGY
264
ß Blackwell Publishers Ltd. 1997
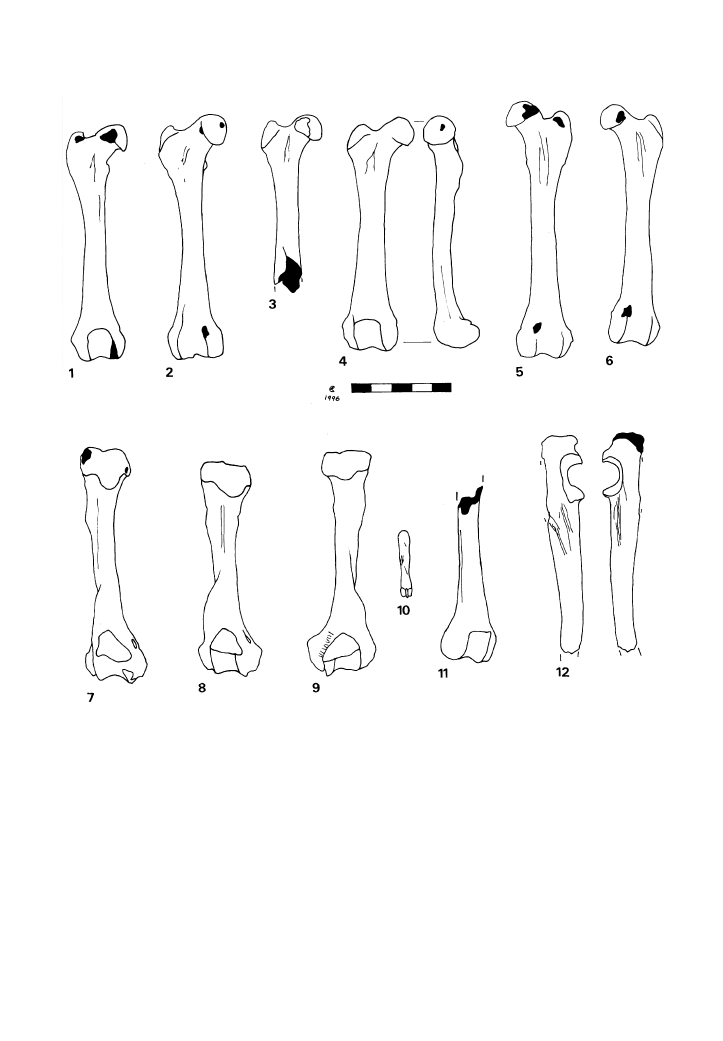
the NISP is too small to formulate any
meaningful discussion of significant trends
in BPR, it is interesting to note in passing that,
as with the foxes from Chaleux, the lower
portions of the badger limbs were totally
absent. Similar arguments to those used in
relation to the fox limbs (above) might be
applied to suggest a case for the skinning of
badgers (and consequently that they too might
have been trapped for their pelts).
The only cut marked badger bones from
Chaleux are five femora. Only longitudinal
Figure 3
Cut-marked badger bones from the Trou des Nutons (1, 7, 8, 11 & 12), Trou de Chaleux (2, 3, 4, 5 & 6) and Star Carr
(9 &10).
RUTH CHARLES
OXFORD JOURNAL OF ARCHAEOLOGY
ß Blackwell Publishers Ltd. 1997
265

groups of cuts were present, almost all of
which were located towards the proximal
ends. The same is true for the five cut bones
from Nutons (a right ulna, two left humeri, a
right femur and a distal left femur) and the
sole cut specimen from Gough’s Cave, a
juvenile humerus with longitudinal cuts
towards the proximal end of the shaft. In
each case these indicate meat removal,
suggesting that whilst badgers may have
been hunted/trapped for their distinctive
pelts, their meat was also of some interest.
Indeed meat removal is the only activity for
which there is any direct evidence at the Late
Upper Palaeolithic sites. In contrast, the two
cut specimens from the early Mesolithic site
of Star Carr show marks which correspond
with skinning (Fig. 3, Nos 9 & 10), although
meat filleting marks are also present on the
distal portion of a humerus. Similarly Row-
ley-Conwy (in press) noted a badger skull
with cut marks relating to skinning from
Ringkloster.
Pine marten (Martes martes) and other small
mustelids
Today there are approximately 53 separate
species of mustelid, which are found primar-
ily in the northern temperate zones of north
America and Eurasia (Schreiber et al. 1989).
These include the badger and the wolverine
(both discussed above) as well as marten,
mink, stoat and weasel. Each member of the
Mustelidae has its own particular set of
environmental preferences and behaviour.
The constraints of space prohibit a detailed
description of each species here.
Mustelids rarely occur in Lateglacial con-
texts in north-western Europe. None were
reported by Currant (1986, 1991) at Cheddar,
neither were any observed by Turner (1991)
in the Neuwied basin Lateglacial faunas.
Although some mustelids have been identi-
fied at the Lesse valley caves, only wolverine
and
badger
remains
(discussed
above)
showed any trace of human modification.
Overall mustelids were only present at these
sites in very low numbers (cf. Charles 1994).
In Mesolithic contexts, however, smaller
mustelids do appear to have figures within
the economy, although accounts are sparse.
At Star Carr the remains of two pine martens
(Martes martes) recovered by Clark show
butchery marks consistent with skinning and
meat removal. The only other evidence for
their exploitation comes from Denmark,
where Rowley-Conwy (1980) has commen-
ted on the specialist exploitation of smaller
fur bearing mammals within the Ertebølle
complex. At Tybrind Vig (Trolle-Larson
1987) the remains of at least 13 complete
pine martens were recovered alongside, a
polecat, and four otters, all apparently
processed for their pelts. Similarly some
articulated skeletons of pine marten were
recovered from Ringkloster (Rowley-Conwy
in press), transverse cuts corresponding with
skinning being present on the front of many
of the crania; overall at least 18 individuals
were recovered from the site.
Prehistoric exploitation of these species
seems to have focused on their pelts. The fact
that they do not appear to occur within
Lateglacial faunas is probably a function of
their distribution and habitat during this time,
rather than the result of any conscious
selection on the part of late ice age trappers.
The apparent increased emphasis on the
exploitation of such species, possibly linked
to the use of specialist trapping camps (cf.
Rowley-Conwy 1980) during the Mesolithic
is probably a reflection of the locally
available resources. The possibility that an
intensification in the trapping of fur bearing
mammals during the Ertebølle made a
valuable a commodity for trade with local
agricultural communities cannot be dis-
THE EXPLOITATION OF CARNIVORES AND OTHER FUR-BEARING MAMMALS
OXFORD JOURNAL OF ARCHAEOLOGY
266
ß Blackwell Publishers Ltd. 1997

counted, and links between the two groups
have been suggested both in terms of lithic
technology and pottery (Zvelebil & Rowley-
Conwy 1986).
FELIDAE
Lynx (Lynx lynx) and Wild Cat (Felis
sylvestris)
The modern range of the wild cat (Felis
sylvestris) includes much of Africa and
Eurasia, the species being highly adaptable
(Kurte´n 1968). In appearance they resemble
the modern domestic tabby, having thick
grey and black banded coats with white
throat patches, and often white patches on the
abdomen. Their range in western Europe has
been severely reduced during recent centuries
due to hunting and trapping, although small
populations have successfully recolonised
parts of Belgium, the Czech Republic,
Slovakia, Germany, France and Scotland.
Their primary diet consists of rodents,
supplemented by rabbits and birds (Anon.
1995, 18).
The lynx (Lynx lynx) is the largest of the
living lynxes, and has today become locally
extinct across much of western Europe.
However, it is primarily a woodland species
and is found in isolated patches of boreal
forest in modern Europe and is widely
distributed across the northern Asiatic taiga
belt, ranging to the south into Asia Minor,
Iran and Tibet (Kurte´n 1968). Adult males
weigh approximately 22 kg, the females
weighing slightly less. Both have greyish
coats, with varying degrees of rust and
yellow; most lynxes are spotted black or
reddish-brown, although occasionally the
pattern is indistinct. Amongst the most
recognisable characteristics are the promi-
nent black ear tufts and shortened tail with a
black tip. They are capable of hunting
ungulates three to four times their own size,
including roe deer and chamois; although
when such species are scarce they will take a
wide range of smaller mammals, especially
hares and pikas. They have a tightly defined
mating season between February and April,
with the young being born during May and
June (Anon. 1995, 13).
Common during much of the Weichselian,
the presence of the lynx within north-western
European Lateglacial faunas has been sus-
pected for some time, and has been con-
firmed as an integral part of the Lateglacial
interstadial fauna from Gough’s Cave, Ched-
dar (Currant et al. 1989; Currant 1991) by
AMS dating (OxA-3411 12650
120 BP
taken from a lynx tibia). Whether lynxes
were also present in north-western Europe
during the early Holocene is still open to
debate, although probable. A cut marked lynx
ulna from Aveline’s Hole, Somerset could
either date to the Late Pleistocene or early
Holocene, as human activity has been
identified and firmly dated at this site during
both periods. However, attempts to date the
lynx ulna via the Oxford AMS unit were
unsuccessful due to low collagen levels in the
specimen (Roger Jacobi pers. comm.). The
transverse cut marks along the shaft of this
specimen correspond with meat filleting, and
it is likely (although undemonstrated) that
this animal was skinned prior to processing.
Wild cat (Felis sylvestris) is also present in
many Late Pleistocene and early Holocene
faunas from western Europe. This species
occurs in all the Late Pleistocene faunas from
the Lesse valley caves, although none of
these specimens show any direct evidence for
human activity. Roger Jacobi (pers. comm.)
has identified two cut and charred distal tibii
of wild cat from Whalley II, associated with
geometric microliths and assumed to be of
mesolithic age. The species is absent from
the Late Pleistocene faunas from The Robin
RUTH CHARLES
OXFORD JOURNAL OF ARCHAEOLOGY
ß Blackwell Publishers Ltd. 1997
267

Hood Cave in Britain and Grotte du Cole´op-
te`re and Sy Verlaine in Belgium and no felids
are recorded in the early Holocene assem-
blage from Star Carr. Outside the British
Isles the most convincing evidence for the
human exploitation of wild cat during the
Mesolithic comes from the Danish Ertebølle
sites of Tybrind Vig (Trolle-Lassen 1987)
and Hjerk Nor (Rowley-Conwy 1980, in
press), which yielded high quantities of bones
from fur bearing species, with a notable bias
towards wild cat at Hjerk Nor. The latter site
was suggested to be a special purpose camp
focussing on the exploitation of fur bearing
mammals, which again links in with the
suggestion (above) that a part of the Ertebølle
economy centred on the exploitation of fur
bearing mammals.
RODENTIA
Beavers (Castor fiber)
Beaver remains have been recorded from
Palaeontological
contexts throughout the
Pleistocene, and there is a continuous record
of this species in western Europe throughout
the Middle and Late Pleistocene (Kurte´n
1968). It is the largest living rodent, and on
average is approximately 110 cm in length.
As with many of the species discussed above,
human exploitation in Europe has signifi-
cantly effected the distribution of contem-
porary beaver populations; although remnant
populations persist in parts of Scandinavia,
the Rhoˆne and Elbe river networks and across
much of eastern Europe into Eurasia. Beavers
are another forest-dwelling animal, and
prefer boreal forest environments closely
allied with river networks. They instinctively
construct
dams,
regulating
water
levels
around their lodges (ibid.).
Beavers have been recovered from archae-
ological contexts in north-western Europe
dating to the early postglacial, although it is
unclear whether they were present in this
region during any part of the preceding
Lateglacial. The earliest radiocarbon deter-
minations available confirm beaver as part of
the northern European fauna by the 9th
millennium BP (OxA-1119 9320
120 BP
from Gough’s Old Cave, Britain and OxA-
2874 9220
90 BP from Stellmoor,
Germany).
Neither
of
these
specimens
showed any signs of human modification,
although another beaver specimen from
Stellmoor is described as a ‘mandibular
artefact of beaver’ (OxA-2873 7830
80
BP; Hedges et al. 1993). Other finds of
beaver within Lateglacial age faunas are
known — the partial
remains of one
individual was recovered from the Grotte de
Sy Verlaine, although whether this specimen
is truly of Lateglacial age is ambiguous (cf.
Charles 1994).
Coles & Orme (1983) propose a subtle
relationship between beaver and human
modification of the landscape, in which the
presence of beaver dams may have led to
locations becoming desirable places for
Mesolithic exploitation. In addition to the
changes in local landscape, the presence of
beavers themselves may have held some
attraction for hunter-gatherers. Eleven of the
beaver bones from Star Carr show cut marks
corresponding with skinning, meat filleting
and disarticulation (Fig. 4). Rowley-Conwy
(in press) reports a beaver skull with skinning
marks from Ringkloster. Beavers make good
eating (Nelson 1973) and their fat, as that of
other species discussed here, would have had
many uses, as a part of prehistoric diet as
well as a resource for other activities. In
addition to this, beavers of both sexes also
produce castoreum, a yellow fluid produced
in the castor glands located near the anal
opening (ibid.). This is used by the beavers to
mark their scent on the shore around their
THE EXPLOITATION OF CARNIVORES AND OTHER FUR-BEARING MAMMALS
OXFORD JOURNAL OF ARCHAEOLOGY
268
ß Blackwell Publishers Ltd. 1997

dams and lodges. Other glands also located in
this region secrete an oil which is used to
maintain their fur quality. If these glands are
extracted, dried and mixed together, their
secretions can be used to make bait for other
terrestrial fur species (ibid.) or medicine
(Kurte´n 1968).
Ethnographic accounts of north American
trappers detail various methods for trapping
beaver and other small mammals (Davis
1986). One common thread is that they are
particularly difficult animals to capture due
to their complex behavioural patterns, and
complicated trapping methods, differing from
those commonly used on land based mam-
mals, are required (Nelson 1973). Having
first identified the location of beaver lodges
in a region and checked that they are in use,
trappers must next locate beaver runways —
habitually used underwater channels from the
beaver lodge and nearby dens to their feed
pile. In winter these are often under ice, and
can be detected by sound. The runways are
used all year round and they provide an
excellent location for setting snares. Nelson
recounts (ibid., 255–257) that multiple snares
are set by the Kutchin Indians in tiers so that
they fill the runway. Such snares are best
checked every two to three days, although
returns diminished once the beavers become
aware of their presence.
Arctic hare (Lepus timidus) and brown hare
(Lepus europaeus)
Present distribution of arctic hare (Lepus
timidus) ranges across to tundra and con-
iferous forest zones of Eurasian Russia and
west Siberia above 50º north. Their distribu-
tion is determined to some degree by
altitude,
Yalden
(1971)
documented
an
altitudinal split between brown and arctic
hares in which the arctic hares occupied a
higher niche between 280 m and 570 m OD.
Their preferred habitat is a mixture of
Calluna- and Eriophorum-dominated heath,
sheltered from predominant winds and away
from grassy areas, although it is widespread
within boreal forests and arctic forest region
(Kurte´n 1968). Population densities tend to
fluctuate within 7 to 12 year cycles, most
probably
linked
to
fluctuations
in
the
numbers of their major predators (especially
the lynx). A striking feature of Lepus timidus
is the change in pelage during the year —
during the summer months the coat is
Figure 4
Cut-marked beaver bones from Star Carr.
RUTH CHARLES
OXFORD JOURNAL OF ARCHAEOLOGY
ß Blackwell Publishers Ltd. 1997
269

brown, but between autumn and spring the
coats turn white. It is one of the larger
lagomorphs, average size being between 52
to 70 cm.
Brown hares are today widespread across
much of Europe (excluding the Iberian
peninsula) and the Near East; this species is
also gradually extending into Scandinavia,
replacing local populations of arctic hares.
This
species
prefers
open
grasslands,
although as with its arctic counterpart, is
found throughout the European boreal forest
zone. This species does not undergo a colour
change, and as its name suggest is brown in
colour and slightly smaller than arctic hares,
the average size being between 48 and 68 cm
in modern specimens.
The exploitation of hares in prehistory has
rarely caused much comment, although
during the Lateglacial a case can be made
for the recognition of specialist hare trapping/
processing stations (cf. Charles & Jacobi
1994). Boyle’s recent review of Upper
Palaeolithic faunas from south-west France
reveals a marked increase in the numbers of
hares and rabbits during the Late Magdale-
nian and Azilian (1990, 203), and again the
use of some sites as specialised processing
camps is a strong possibility. For example,
Boyle cites the striking example of Pont
d’Ambon, in which the conventional view of
the faunal assemblage is one of red deer
domination (90% of the large herbivore
fauna) however, if calculations are modified
to include the hares and rabbits (represented
by over 6000 bones), then red deer fall to
only 8.1% of the species at the site — rabbits
and hares accounting for almost 90% of the
fauna.
Hares possess a number of potentially
attractive features apart from their meat for
Late Pleistocene trappers. Their pelts could
have provided warm linings and trimmings
for clothing, their bones the raw materials for
a range of tools, and sinews and tendons the
materials for cordage and thread.
The Robin Hood Cave at Creswell Crags is
perhaps one of the better known examples of
a Lateglacial hare exploitation site. Butchery
evidence was used as a key to re-assess the
faunal assemblage, revealing that the only
species whose presence at the site was
demonstrably due to human activity were
arctic hares. Individual cut specimens of
these were selected as samples of AMS
dating (Charles & Jacobi 1994). As only low
numbers of cut hare bones were present and
as hares and rabbits are both extremely lean
animals, it was suggested that use of this site
during the Lateglacial was short-lived, as
humans could not have survived for long on
such a lean diet, the phenomenon of ‘protein
poisoning’ (Speth & Spielmann 1983) being
invoked.
Additionally evidence for the exploitation
of hares has been noted at three further sites
under discussion here, all dating to the
Bølling Interstadial phase of the Lateglacial
— The Trou de Chaleux (three cut speci-
mens), the Trou du Frontal (four cut speci-
mens) and Gough’s Cave at Cheddar (three
cut specimens). Whilst none of these sites
can be seen as a specialist hare processing
camps, it is worth noting that this species
provided a dietary change from the mainstays
of Late Palaeolithic diet recorded at these
sites — the horse, aurochs and red deer.
DISCUSSION
Whilst by no means can it be argued that
carnivores and other fur-bearing mammals
formed a dietary staple for western European
hunter-gatherers during the Late Pleistocene
and early Holocene, it is clear that they did
make a contribution to the human economy.
Rowley-Conwy (1980) has argued that the
exploitation of fur bearing mammals is most
THE EXPLOITATION OF CARNIVORES AND OTHER FUR-BEARING MAMMALS
OXFORD JOURNAL OF ARCHAEOLOGY
270
ß Blackwell Publishers Ltd. 1997

likely to occur during the winter, as this is the
season when their fur is of the highest
quality. Accounts of recent north American
hunters and trappers (Nelson 1973) concur
with this suggestion, and add that fur-
trapping primarily for trading purposes is
only likely to occur in times of abundance
(Winterhalder 1980). It is unlikely, however,
that animals were trapped solely for their
furs, and the evidence presented here con-
firms that meat was removed from many of
the carnivores and fur bearing mammals
discussed above. In addition to providing
pelts for clothing and barter, such animals
could have provided an occasional alternative
to dietary staples such as reindeer, horse, red
deer and aurochsen and/or provided meat
when such herbivores were rare or absent.
Whilst the role of larger mammals cannot
be understated during the European Late
Palaeolithic and Mesolithic, a picture of
increased emphasis on the exploitation of
fur bearing mammals during this period is
emerging alongside the recognition of spe-
cialist exploitation camps such as the Robin
Hood Cave and Hjerk Nor. This suggests a
broadening of the subsistence base. Rather
than arguing for a dramatic shift in the nature
of faunal exploitation during the Lateglacial
and early Postglacial, it is suggested here that
the exploitation of fur bearing mammals and
other carnivores reflects an expansion in the
economic base.
In addition to providing a supplement to
dietary staples of locally available plant
foods, ungulate and marine resources, the
meat from carnivores and fur-bearing mam-
mals could have been used as bait to entice
further animals into traps. This is certainly
possible, although regular use of meat
obtained in this way is unlikely due to the
low minimum number of individuals counts
for each species encountered at each site.
It is probable that an expansion in the
subsistence base can be viewed within the
broader context of environmental change
during the Lateglacial and the eventual
development of Boreal forests in north-
western Europe during the early Postglacial,
bringing with it commensurate changes in
regional ecology. The data discussed within
this paper are drawn from archaeological sites
dating either to one of the Interstadial phases
of the Lateglacial or to the early Postglacial.
The evidence for forest development during
the Bølling Interstadial is poor for north-
western Europe, and so the true nature and
extent of forest/tree cover surrounding the
sites discussed here and dating to that period
remains unclear. Better information is avail-
able for the early Postglacial (Simmons et al.
1981), particularly in the vicinity of Star Carr
(Walker & Godwin 1954, Cloutman 1988,
Cloutman & Smith 1988, Day & Mellars
1994).
The European earlier Holocene is gener-
ally characterised by the increase in plant and
tree cover leading to the eventual appearance
of climax forest during the Atlantic phase of
the Flandrian; alongside this eustatic and
isostatic changes radically transformed the
landscape, including the formation of the
English channel and the Irish Sea (Simmons
et al. 1981). The larger mammals such as red
deer (Cervus elaphus) present in north-
western Europe during the Lateglacial and
early Postglacial are relatively flexible in
their behavioural patterns, and are known to
congregate in large herds in open environ-
ments and to lead far more solitary life styles
in forested ones. Shifts in exploitation
strategies have been suggested (Jacobi et al.
1976; Simmons et al. 1981) as adaptations to
these environmental changes.
A model developed to explain the diversity
of resource procurement amongst the north
American Cree seems the most appropriate
analogy here, in which the vast and widely
RUTH CHARLES
OXFORD JOURNAL OF ARCHAEOLOGY
ß Blackwell Publishers Ltd. 1997
271

(but unevenly) distributed resources of boreal
forests are recognised by foragers familiar
with a large area, but in which it is only
feasible to exploit a small portion in any
given season or year (Winterhalder 1983). By
applying the general principles of Optimal
Foraging Theory (cf. Krebs 1977), a series of
different ‘patches’ within the boreal forest
can be recognised in which the available prey
occurs in small, dispersed ‘packets’. Conse-
quently, the expectation is that foragers
spend a considerable amount of time search-
ing for these ‘packets’, pursue a relatively
large proportion of the species encountered
on any given trip and so have a fairly broad
dietary range and utilise a wide range of
‘patches’.
In such a model one would expect the
primary dietary focus to be towards the
exploitation of larger mammals, further
subsidised by the trapping of smaller mam-
mals. This suggestion is certainly borne out
at many of the sites discussed here — the
Trou de Chaleux, Trou des Nutons, Trou du
Frontal, Gough’s Cave, Ringkloster, Tybrind
Vig and Star Carr). These faunal assem-
blages are dominated by large and medium
sized ungulates, all of which show extensive
butchery marks relating to skinning, meat
removal,
disarticulation
and
subsequent
smashing of bones for marrow. Additionally,
various smaller fur bearing mammals are
present which bear evidence for both skin-
ning and meat extraction. All of these sites
can be interpreted as hunting camps, and in
most cases the use of these locations by late
Pleistocene and early Holocene hunter-gath-
erers is believed to have been seasonal. It
seems most likely that when the opportunity
arose for the exploitation of small game this
was unlikely to be disregarded. Efficient
trapping of such animals could be greatly
enhanced by the use of ‘unattended facilities’
(or traps), as the pursuit of larger mammals
can be combined with the routine checking
of trap lines, thus maximising the potential
returns. Such an explanation is proposed here
to account for the widening of the dietary
niche within north-western European Late
Palaeolithic and Mesolithic groups.
Such a model does not, however, exclude
the presence of specialist trapping camps
aimed primarily at the recovery of pelts (in
this case the Robin Hood Cave and Hjerk
Nor) — and again such sites have been
recognised in both the Late Upper Palaeo-
lithic and Mesolithic, although they are
generally seen as exceptions to the general
‘rule’ of large-mammal dominated sites.
Alongside this the possible expansion of
symbolic expression via clothing and perso-
nal adornment cannot be excluded, although
this aspect of stone age life remains difficult
to quantify. Highly stylised engravings are
known from a variety of Lateglacial and early
Postglacial locations, including Go¨nnersdorf
in Germany (Bosinski 1991), Gough’s Cave
(Charles 1989) and Kendrick’s Cave (Sievek-
ing 1971) in Britain. Gamble (1991) has
suggested that the Magdalenian ‘symbolic
explosion’ of the Lateglacial can be inter-
preted within the context of social knowledge
and communication between pioneer groups
re-colonising the north-western European
mainland, allowing for the maintenance of
social contacts over great distances. He notes
that this use of symbolism within material
culture appears to diminish after the pioneer
phase. However, with the dramatic changes in
landscape coincident with the early postgla-
cial, one can again detect the increasing use of
symbolic media within the archaeological
record. Star Carr in Britain (Clark 1954),
Berlin-Biesdorf (Reinbacher 1956), Hohen
Viecheln (Schuldt 1961) and Bedburg-Ko¨nig-
shoven in Germany (Street 1991) have
yielded worked frontlets of red deer: whilst
the use of these objects remains ambiguous,
THE EXPLOITATION OF CARNIVORES AND OTHER FUR-BEARING MAMMALS
OXFORD JOURNAL OF ARCHAEOLOGY
272
ß Blackwell Publishers Ltd. 1997

they are frequently interpreted as head-gear
used as part of hunting equipment and/or
ritual. Inspection of the ones recovered from
Star Carr revealed that the antlers on these
specimens had been shaved or whittled on
their medial surfaces, which would have
served to lighten the weight of these objects
whilst not diminishing their overall effect
when viewed from below, adding some
circumstantial evidence to the suggestions
that they were indeed worn, and that they
were less likely to be used in a hunting
situation in which the wearer was unlikely to
be viewed from below.
In addition to the isolated occurrences
described above, lithic technology shifted
from the pan-European early Mesolithic
‘broad blade’ industries (consisting primarily
of obliquely blunted points) to the regionally
specific late and later Mesolithic ‘narrow
blade’ industries (made up of a range of
microlithic forms) (Jacobi 1976, Gendel
1989, Arts 1989). The redefinition of the
landscape by marine transgressions, river
development and expansion and forest growth
would have had a profound impact on
Mesolithic communities and their communi-
cations. If population densities changed (cf.
Meiklejohn 1978), and social groups re-
structured as an adaptation to different
resource availability, then a need for the
maintenance of social contact and knowledge
between such groups would have become a
priority. This social change may be invoked as
at least one of the reasons behind an
expansion of symbolic behaviour during the
European Mesolithic.
Whilst it is unlikely that archaeologists can
today reconstruct the precise belief systems
associated with the use of such animals, it is
perhaps possible to acknowledge that in
certain instances a symbolic explanation is
the most appropriate. Examples might in-
clude the dog burials from the late Mesolithic
cemeteries at Skateholm and Bredasten,
Sweden (Larsson 1994) and at Vedbaek,
Denmark (Brinch Petersen 1990) as well as
the child burial placed on a swan’s wing at
the Ertebølle cemetery at Vedbaek, Denmark
(Albrethsen & Brinch Petersen 1976).
Whilst it is true that in many recent and
contemporary societies the hunting/trapping
and subsequent exploitation of small mam-
mals is often undertaken by women and
children, this does not preclude males taking
part in such activities (cf. Szeuter 1988).
Suggestions that an emphasis on the exploi-
tation of smaller mammals equate with an
increased role for women and children in the
food quest cannot be supported here. The
evidence presented shows a direct link
between human activity and the exploitation
of small mammals; cut marks (and other
forms of butchery evidence) can say little
about the gender or biological sex of the
individual who wielded the stone tools which
made them — only that such activities took
place. Whilst the search for female perspec-
tives and roles within Palaeolithic society
remains an important research issue (cf.
Conkey 1991), there seems little opportunity
to develop it within the framework of this
paper, unless the reader wishes to reinforce
stereotypes of Palaeolithic and Mesolithic
societies in which men hunted big game and
women small game. Neither such scenario is
necessarily valid, and the gender of the
hunters, trappers and butchers remains un-
known.
CONCLUSIONS
The evidence presented here relating to the
exploitation of carnivores and other fur
bearing mammals during the late Palaeolithic
and early Mesolithic is drawn from the
author’s personal observations combined
with examples known from the published
RUTH CHARLES
OXFORD JOURNAL OF ARCHAEOLOGY
ß Blackwell Publishers Ltd. 1997
273
literature. By no means can it be seen as a
definitive account. The purpose of this paper
has been to document specific instances
where such mammals have been exploited,
and to draw attention to aspects of Palaeo-
lithic and Mesolithic faunal exploitation
beyond the conventional view of these
populations as big game hunters. This is not
to say that large mammals were not hunted
— they clearly were, and dominate many of
the faunal assemblages discussed here. In-
stead this paper has highlighted another
aspect of prehistoric subsistence, suggesting
that carnivores and fur bearing mammals
provided occasional variety to Late Pleisto-
cene and early Holocene diets as well as a
useful resource for pelts used in clothing
manufacture and trade. It is hoped that
evidence reported here will stimulate an
awareness of the potential within Palaeolithic
and Mesolithic faunal assemblages to provide
information on the role of subsidiary re-
sources, such as those provided by fur
bearing mammals.
Acknowledgements
I would like to thank Peter Rowley-Conwy for
providing access to his unpublished work, Juliet
Clutton-Brock and Pat Carter for giving access to Star
Carr material, Hans Kruuk for his many comments
about badger behaviour, Roger Jacobi for adding to my
knowledge of the British Lateglacial database and
Derek Roe for his many comments on draft versions
of this paper. Figure 1 was drawn by R.M.C. Cook.
Much of the material reported here was analysed as part
of doctoral research undertaken by the author between
1990 and 1994 and funded by The British Academy,
The Queen’s College, Oxford and grants from the
Meyerstein Fund of Oxford University and Christ
Church, Oxford.
Department of Archaeology
University of Newcastle
Newcastle-upon-Tyne NE1 7RU
REFERENCES
ALBRECHT, T., BURKE, H.
and
POPLIN, F.
1983: Naturwis-
senschaftliche Untersuchungen an Magdale´nien-Inven-
taren
vom
Petersfels,
Grabungen
1974–1976
(Tu¨bingen, Tu¨binger Monographien zur Urgeschichte,
Herausgegeben von Hansju¨rgen Mu¨ller-Beck Band 8).
ALBRETHSEN,
S.E.
and
BRINCH
PETERSEN,
E.
1976:
Excavation of a Mesolithic cemetary at Vedbaek,
Denmark. Acta Archaeologica 47, 1–28.
ALLSWORTH-JONES, P.
1986: The Szeletian and the
transition from Middle to Upper Palaeolithic in Central
Europe (Oxford, Oxford University Press).
ANDREWS, P.A.
and
COOK, J.
1985: Natural modification to
bones in a temperate setting. Man 20, 675–691.
ANON
. 1995: Wild Cats (London, IUCN).
ARTS, N.
1989: Archaeology, environment and the social
evolution of later band societies in a lowland area. In
Bonsall, C. (ed.) The Mesolithic in Europe (Edinburgh,
John Donald Publishers Ltd), 291312.
BARTA, J.
1989: Hunting of Brown Bears in the
Mesolithic: Evidence from the Medvedia Cave near
Rusı´n in Slovakia. In Bonsall, C. (ed.) The Mesolithic in
Europe (Edinburgh, John Donald Publishers Ltd), 456–
460.
BENECKE, N.
1987: Studies on Early Dog Remains from
Northern Europe. Journal of Archaeological Science 14,
31–49.
BRATLUND, B.
1991: The bone remains of mammals and
birds from the Bjørnsholm shell-mound. Journal of
Danish Archaeology 10, 97–104.
BRINCH PETERSEN, E.
1990: Nye grave fra Jaegerstenal-
deren. Strøby Egede og Vedbaek (Copenhagen, Natio-
nalmuseets Arbejdsmark), 19–33.
BOSINSKI,
G.
1991: The representation of female
figurines in the Rhineland Magdalenian. Proceedings
of the Prehistoric Society 57(1), 51–64.
BOYLE, K.V.
1990: Upper Palaeolithic Faunas from
South-West France: a zoogeographic perspective (Ox-
ford, British Archaeological Reports, International
THE EXPLOITATION OF CARNIVORES AND OTHER FUR-BEARING MAMMALS
OXFORD JOURNAL OF ARCHAEOLOGY
274
ß Blackwell Publishers Ltd. 1997

Series 557).
CHARLES, R.
1989: Incised ivory fragments and other
Late Palaeolithic finds from Gough’s Cave, Cheddar,
Somerset. Proceedings of the University of Bristol
Spelaeological Society 18(3), 400–408.
CHARLES, R.
1994: Food for Thought: Late Magdalenian
Chronology and Faunal Exploitation in the north-
western Ardennes (Unpublished D.Phil. dissertation,
University of Oxford).
CHARLES, R.
and
JACOBI, R.M.
1994: Lateglacial faunal
exploitation at the Robin Hood Cave, Creswell Crags.
Oxford Journal of Archaeology 13(1), 1–32.
CLARK, J.G.D.
1952: Prehistoric Europe: The economic
basis (London, Methuen & Co. Ltd).
CLARK, J.G.D.
1954: Excavations at Star Carr, an early
Mesolithic site at Seamer, near Scarborough, Yorkshire
(Cambridge, Cambridge University Press).
CLOUTMAN, E.W.
1988: Palaeoenvironments in the Vale
of Pickering part 2: Environmental History at Seamer
Carr. Proceedings of the Prehistoric Society 54, 21–36.
CLOUTMAN, E.W.
and
SMITH, A.G.
1988: Palaeoenviron-
ments in the Vale of Pickering, part 3: Environmental
History at Star Carr. Proceedings of the Prehistoric
Society 54, 37–58.
CLUTTON-BROCK, J., CORBET, G.B.
and
HILLS, M.
1976: A
review of the family Canidae, with a classification by
numerical records. Bulletin of the Natural History
Museum (Zoology) 29(3), 119–199.
COLES, J.M.
1991: Elk and Ogopogo. Belief systems in
the hunter-gatherer rock art of northern lands. Proceed-
ings of the Prehistoric Society 57(1), 129–147.
COLES, J.M.
and
ORME, B.J.
1983: Homo sapiens or Castor
fiber? Antiquity 57, 95–102.
CONKEY, M.W.
1991: Contexts of Action, contexts for
power: Material culture and gender in the Magdalenian.
In Gero, J.M. and Conkey, M.W. (eds), Engendering
Archaeology (Oxford & Cambridge, Basil Blackwell),
57–92.
CREMADES, M.
1992: Analyse et reconstitution technolo-
giques en art mobilier Pale´olithique l’example du
glouton grave´ sur baton perfore´ de la Madeleine
(Dordogne). L’Anthropologie 96(2–3), 319–336.
CURRANT, A.P.
1986: The Lateglacial mammal fauna of
Gough’s Cave, Cheddar, Somerset Proceedings of the
University of Bristol Spelaeological Society 17(3), 286–
304.
CURRANT, A.P.
1991: A Late Glacial Interstadial mammal
fauna from Gough’s Cave, Somerset, England. In
Barton, N., Roberts, A.J., and Roe, D.A. (eds.) The
Late Glacial in north-west Europe: human adaptation
and environmental change at the end of the Pleistocene
(London, CBA Research report 77), 48–50.
CURRANT, A.P., JACOBI, R.M.
and
STRINGER, C.B.
1989:
Excavations at Gough’s Cave, Somerset 1986–7.
Antiquity 63(238), 131–136.
DAVIS, S.A.
1986: Man, molluscs and mammals: a study
of land use and resources in the late Holocene of the
maritime provinces of Canada (Unpublished D.Phil.
dissertation, Oxford University).
DAY, S.P.
and
MELLARS, P.A.
1994: Absolute dating of
Mesolithic human activity at Star Carr, Yorkshire: new
palaeocological studies and identification of the 9600
BP radiocarbon ‘plateau’. Proceedings of the Prehis-
toric Society 60, 417–422.
DEGERBØL, M.
1961: On the find of a preboreal dog
(Canis familiaris L.) from Star Carr, Yorkshire, with
remarks on other mesolithic dogs. Proceedings of the
Prehistoric Society 27, 35–65.
DOMICO, T.
1988: Bears of the World (New York &
Oxford, Facts on File).
FRASER, F.C.
and
KING, J.E.
1954: Faunal remains. In
Clark, J.G.D. (ed.) Excavations at Star Carr (Cam-
bridge, Cambridge University Press) 70–95.
GAMBLE, C.S.
1991: The social context for European
Palaeolithic art. Proceedings of the Prehistoric Society
57(1), 3–15.
GENDEL, P.A.
1989: The analysis of lithic styles through
distributional profiles of variation: examples from the
western European Mesolithic. In Bonsall, C. (ed.) The
Mesolithic in Europe (Edinburgh, John Donald Publish-
ers Ltd), 40–47.
GINSBERG, J.R.
and
MACDONALD, D.W.
1990: Foxes,
Wolves, Jackals, and Dogs. An action plan for the
conservation of Canids. (IUCN/SSC Canid Specialist
Group).
HEDGES, R.E.M., HOUSLEY, R.A., BRONK RAMSEY, C.
, and
VAN KLINKEN, G.J.
1993: Radiocarbon dates from the
Oxford AMS system: Archaeometry Datelist 16.
Archaeometry 35(1), 147–67.
JACOBI, R.M.
1976: Britain inside and outside Mesolithic
Europe. Proceedings of the Prehistoric Society 42, 67–
84.
RUTH CHARLES
OXFORD JOURNAL OF ARCHAEOLOGY
ß Blackwell Publishers Ltd. 1997
275

JACOBI, R.M., TALLIS, J.H.
and
MELLARS, P.A.
1976: The
southern Penine Mesolithic and the ecological record.
Journal of Archaeological Science 3, 307–20.
KLEIN, R.G.
1973: Ice-Age Hunters of the Ukraine
(Chicago, University of Chicago Press).
KREBS, J.
1977: Optimal foraging: Theory and experi-
ment. Nature 268, 583–584.
KRUUK, H.
1989: The Social Badger. Ecology and
behaviour of a group living carnivore (Meles meles)
(Oxford, Oxford University Press).
KURTE
´ N, B.
1968: Pleistocene Mammals of Europe
(London, Weidenfield & Nicholson).
LARSSON, L.
1994: Pratiques mortuaires et se´pultures de
chiens dans les socie´te´s Me´solithiques de Scandinavie
me´ridionale. L’Anthropologie 98(4), 562–575.
MECH, L.P.
1970: The Wolf (New York, Natural History
Press).
MEIKLEJOHN, C.
1978: Ecological aspects of population
size and growth in Late-glacial and early Postglacial
north-western Europe. In Mellars, P.A. (ed.) The early
Postglacial settlement of north-western Europe (Lon-
don, Duckworth).
NEAL, E.
1948: The Badger (London, Collins New
Naturalist Monographs).
NELSON, R.K.
1973: Hunters of the northern Forest:
Designs for survival among the Alaskan Kutchin
(Chicago, The University of Chicago Press).
PARTRIDGE, J.
1992: Management Guidelines for Bears
and Racoons (Bristol, The Association of British Wild
Animal Keepers).
REINBACHER, E.
1956: Eine vorgeschichtliche Hirsch-
maske aus Berlin-Biesdorf. Ausgrabungen und Funde 1,
147–51.
ROWLEY-CONWY, P.A.
1980: Continuity and Change in
the Prehistoric economies of Denmark 3700 bc to 2300
bc (Unpublished PhD Dissertation, University of Cam-
bridge).
ROWLEY-CONWY, P.A.
in press. Meat, furs and skins:
Mesolithic animal bones from Ringkloster, a seasonal
hunting camp in Jutland. Journal of Danish Archae-
ology.
SCHREIBER, A., WIRTH, R., RIFFEL, M.
, and
VAN ROMPAEY, H.
1989: Weasels, Civets, Mongooses, and their relatives.
An Action Plan for the Conservation of Mustelids and
Viverrids (IUCN/SSC Mustelid and Viverrid Specialist
Group).
SCHULDT, E.
1961: Hohen Viecheln. Ein mittel-steinzei-
tlicher wohnplatz in Mecklenburg (Berlin, Deutsche
Akademie de Wissenschaften zu Berlin Sektion fu¨r Vor-
und Fru¨geschichte 10).
SIEVEKING, G. de G.
1971: The Kendrick’s Cave mandible.
British Museum Quarterly 35, 230–50.
SIMMONS, I.G., DIMBLEBY, G.W.
and
GRIGSON, C.
1981: The
Mesolithic. In Simmons, I. and Tooley, M. (eds.) The
Environment in British Prehistory (London, Duck-
worth).
SPETH, J.D.
and
SPIELMANN, K.A.
1983: Energy source,
protein metabolism and hunter-gatherer subsistence
strategies. Journal of Anthropological Archaeology 2,
131.
STREET, M.
1991: Bedburg-Ko¨nigshoven: A pre-boreal
Mesolithic site in the lower Rhineland, Germany. In
Barton, N., Roberts, A.J., and Roe, D.A. (eds.) The Late
Glacial in north-west Europe (London, CBA research
report 77), 256–270.
SZEUTER, C.R.
1988: Small animal exploitation among
desert horticulturalists in north America. Archaeozoo-
logica 2(1), 191–200.
TROLLE-LASSEN, T.
1986: Human exploitation of the Pine
Marten (Martes martes (L.)) at the Late Mesolithic
settlement of Tybrind Vig in western Funen. In
Ko¨nigsson, L.-K. (ed.) Nordic Late Quaternary Biology
and Ecology 24, 119–124.
TROLLE-LASSEN, T.
1987: Human exploitation of fur
animals in Mesolithic Denmark — a case study,
Archaeozoologia 1(2), 85–102.
TURNER, E.
1991: Pleistocene Stratigraphy and Verte-
brate Faunas from the Neuwied Basin region of Western
Germany. Cranium 8(1), 21–34.
WALKER, D.
and
GODWIN, H.
1954: Lake stratigraphy,
pollen-analysis and vegetational history. In Clark,
J.G.D. (ed.) Excavations at Star Carr, an early
Mesolithic site at Seamer near Scarborough, Yorkshire
(Cambridge, Cambridge University Press) 25–69.
WINTERHALDER, B.
1980: Canadian fur bearer cycles and
Cree-Ojibwa hunting and trapping practices. The
American Naturalist 115(6), 870–879.
WINTERHALDER, B.
1983: Boreal Foraging Strategies. In
Steegmann, A.T. (ed.) Boreal Forest Adaptations (New
York: Plenum Press) 201–241.
YALDEN, D.W.
1971: The Mountain Hare (Lepus timidus)
THE EXPLOITATION OF CARNIVORES AND OTHER FUR-BEARING MAMMALS
OXFORD JOURNAL OF ARCHAEOLOGY
276
ß Blackwell Publishers Ltd. 1997

in the peak District. Naturalist 91, 81–92.
ZVELEBIL, M.
and
ROWLEY-CONWY, P.A.
1986: Foragers
and Farmers in Atlantic Europe. In Zvelebil, M. (ed.)
Hunters in transition: Mesolithic societies of temperate
Eurasia and their transition to farming (Cambridge,
Cambridge University Press New Directions in Archae-
ology), 67–93.
RUTH CHARLES
OXFORD JOURNAL OF ARCHAEOLOGY
ß Blackwell Publishers Ltd. 1997
277
Wyszukiwarka
Podobne podstrony:
The Hound of?ath and Other Stories
Howard, Robert E The Gates of Empire and Other Tales of the Crusades
The Presentation of Self and Other in Nazi Propaganda
The Mud of a Jew and Other Hassidic Stories Vol 1
Memes and the Exploitation of Imagination
Yasunari Kawabata The Dancing Girl of Izu and Other Stories (rtf)
Blanchard European Unemployment The Evolution of Facts and Ideas
On The Manipulation of Money and Credit
The Repentance of Compassion and Enlightenment Path
Piórkowska K. Cohesion as the dimension of network and its determianants
1b The Literature of Discovery and Exploration
Laszlo, Ervin The Convergence of Science and Spirituality (2005)
Childhood Trauma, the Neurobiology of Adaptation, and Use dependent of the Brain
annex vi ext of the Protocol of 1997 and Annex VI
The Struggles of EMTs and EMS Workers
The book of dreams and ghosts
więcej podobnych podstron