Transport związków organicznych przez błony
Transport związków organicznych przez błony może odbywać się na wiele sposobów. U bakterii obecne są wspomniane wyżej permeazy - ABC transportery selektywnie pobierające cukry, aminokwasy witaminy i inne potrzebne bakteriom związki.
Ważnym związkiem organicznym, który musi być transportowany przez błonę jest glukoza. Transportery glukozy GLUT1-GLUT4 to błonowe białka będące uniporterami glukozy. Różnią się właściwościami kinetycznymi. GLUT1 ma bardzo małą Km i pobiera glukozę z krwi mniej więcej ze stałą intensywnością. GLUT2 ma Km ok. 13 razy większe, ale dzięki temu jest bardziej czuły na zmiany stężenia glukozy we krwi. Ekspresjonowany jest w hepatocytach i komórkach β trzustki. Pozwala to im na wyczucie zwiększonego stężenia glukozy we krwi po posiłku i odpowiednio rozpoczęcie syntezy glikogenu i wydzielenie insuliny. GLUT4 ekspresjonowany jest w mięśniach i tkance tłuszczowej i lokuje się w błonie komórkowej w odpowiedzi na insulinę, zwiększając dopływ glukozy do komórek tych tkanek.
Odmienny mechanizm pobierania substancji organicznych występuje w komórkach nabłonka jelita cienkiego. Glukoza, inne cukry oraz aminokwasy muszą być pobierane ze światła jelita i przekazywane do krwi nie zależnie od ich stężenia w jelicie i komórkach nabłonka. Dodatkowo, substancje te nie mogą przenikać między komórkami nabłonka, ponieważ komórki te przylegają do siebie bardzo ściśle (tworzą tight junctions). Dlatego też apikalna (czyli ta od strony jelita) błona komórek nabłonkowych jelita posiada wiele specyficznych symporterów, które pobierają cukry, aminokwasy itp. razem z jonami sodowymi. Niskie stężenie sodu w komórce umożliwiające ten transport zapewniane jest przez pompy sodowo-potasowe na bazolateralnej (czyli tej od strony innych tkanek i naczyń krwionośnych) błonie komórki. Natomiast wypływ substancji odżywczych z komórek nabłonka do macierzy zewnątrzkomórkowej i dalej do krwi odbywa się przez uniportery, zgodnie z gradientem stężeń.
36.Dyfuzja prosta, dyfuzja ułatwiona
Dyfuzja prosta odbywa się przez dwuwarstę lipidową i podlegają jej małe niepolarne cząstki oraz niektóre małe polarne np. gazy (N2, CO2, O2), etanol, benzen, do pewnego stopnia woda i mocznik. Zachodzi ona zgodnie z gradientem stężeń, a jej szybkość zależy liniowo od hydrofobowości cząstki i różnicy stężeń.
Dyfuzja ułatwiona zachodzi z udziałem białkowych kanałów, które tworzą hydrofilowe środowisko w środku swojej struktury lub uniporterów. Jej szybkość nie zależy od hydrofobowości cząstki i rośnie hiperbolicznie (tak jak enzymy kinetyki MM) ze stężeniem, bo transportery wysycają się.
Kanały, nośniki, pory.
Kanały to integralne białka błony tworzące w jej poprzek hydrofilowe korytarze dla jonów lub wody. Mogą być specyficzne albo niespecyficzne, bramkowane (napięciem, mechanicznie, chemicznie) albo nie. Działają na zasadzie dyfuzji ułatwionej i przenoszą wiele cząsteczek naraz (10^7-10^8 na sekundę).
Przykłady kanałów to kanały sodowe bramkowane napięciem w neuronach.
Pory również są transmembranowymi białkami pozwalającymi na dyfuzję ułatwioną. Mogą być też zbudowane z beta-baryłek, tak jak bakteryjne poryny. Pory, w odróżnieniu od kanałów są niespecyficzne, przenoszą wydajniej (bo są większe) i są zawsze otwarte.
Przykłady porów: Jądrowy (cząsteczki do 5kDa), połączenie szczelinowe (do 1,5kDa)
Nośniki to białka przenoszące cząsteczki na zasadzie uniportu (transport bierny) symportu albo antyportu (transport aktywny). Od kanałów różnią się tym, że nie muszą przenosić zgodnie z gradientem stężeń. Poza tym są dużo wolniejsze (10^2-10^4 cząstek na sekundę) bo muszą związać ligand, zmienić konformację i uwolnić ligand.
Kanały jonowe - regulacja otwierania
Kanały jonowe moją stan otwarty i zamknięty. Przejście z jednego do drugiego może być spowodowane:
napięciem: np. sodowe, potasowe, wapniowe. Depolaryzacja błony powoduje przesunięcie domen czułych na napięcie i otwarcie kanału, kanały zamykają się spontanicznie, ale potrzebna jest repolaryzacja, żeby uczulić je na następny bodziec
chemicznie: np. nikotynowy receptor acetylocholiny otwiera się po związaniu dwóch cząsteczek acetylocholiny, specyficznie przepuszczając kationy. Zamyka się na skutek zmniejszenia stężenia acetylocholinę w szczelinie synaptycznej.
Mechanicznie: np. bakteryjne kanały jonowe otwierają się w wyniku pęcznienia komórki, żeby przeciwdziałać rozerwaniu komórki przez ciśnienie osmotyczne. Mechanoreceptory układu nerwowego również zawierają kanały bramkowane mechanicznie.
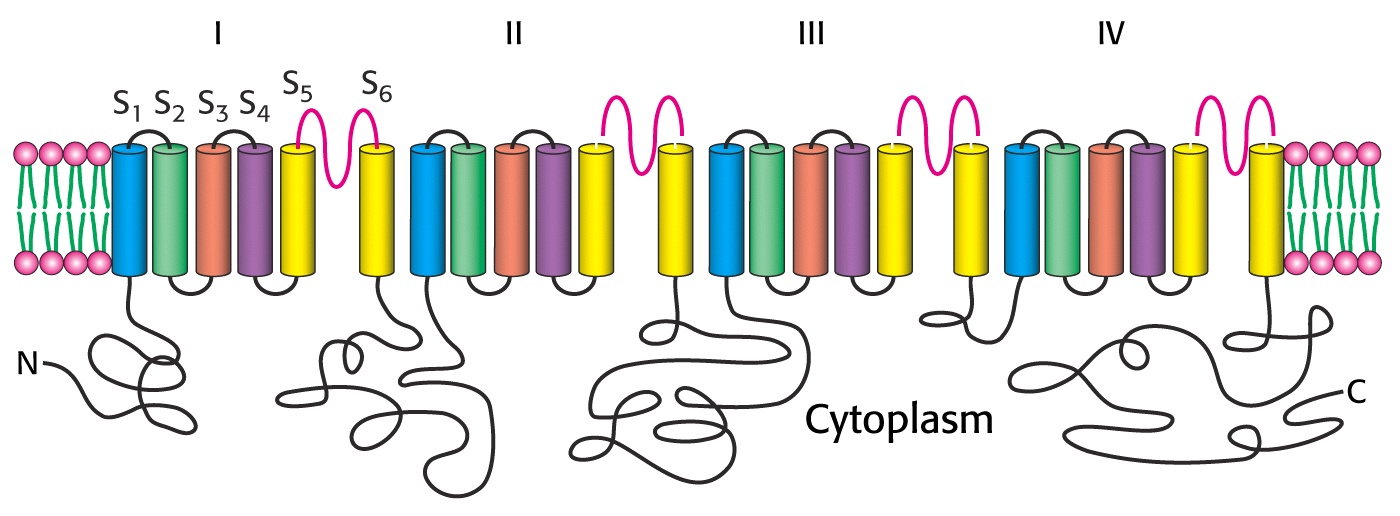
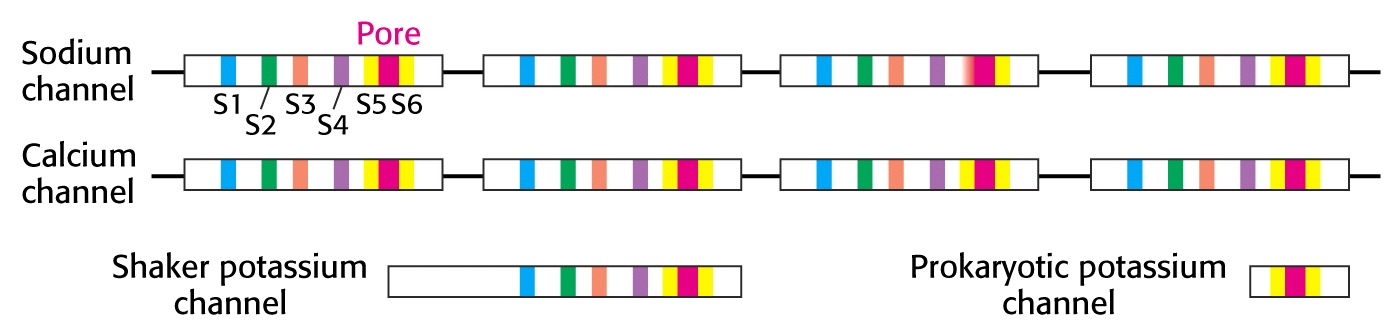
Stany konformacyjne kanału sodowego bramkowanego napięciem
SKOPIOWANE ZE STARYCH ZAGADNIEN BO MI SIE NIE CHCIALO
Kanał zawiera 4 wewnętrzne powtórzenia o podobnych sekwencjach aminokwasów. Każda homologiczna jednostka sekwencji zawiera 5 segmentów hydrofobowych (S1, S2, S3, S5 i S6). Każde powtórzenie zwiera segment S4 o silnym ładunku dodatnim, zawierającym dużo reszt argininy lub lizyny. Te dodatnio naładowane aminokwasy stanowią co trzecią resztę, co sugeruje, że są one umiejscowione po jednej stronie transbłonowej α helisy i są sparowane, przy spoczynkowym potencjale błonowym, z ujemnymi ładunkami na innych helisach transbłonowych. Depolaryzacja prowadzi do spiralnego ruchu segmentu S4, któremu towarzyszy przemieszczenie netto jednego lub dwóch ładunków dodatnich ku zewnątrzkomórkowej stronie błony.
Istnieją przynajmniej trzy stany zamknięte poprzedzające stan otwarty. W czasie każdego z tych przejść (trwających 25μs), wywołanych depolaryzacją poprzez dwuwarstwę lipidową przemieszcza się równoważnik dwóch ładunków.
Stan otwarty trwa tylko około 1ms, nawet wtedy gdy błona utrzymywana jest nadal w stanie depolaryzacji. Stan otwarty jest krótkotrwały, ponieważ przechodzi spontanicznie w stan nieaktywny, który nie może być już otwarty ze względu na potencjał błonowy. Za inaktywację odpowiedzialna jest pętla domeny cytozolowej zawarta między powtórzeniami III i IV polipeptydu tworzącego kanał.
Powrót do stanu zamkniętego, ale zdolnego do aktywacji wymaga ponownej polaryzacji błony. Kanał otwiera się tylko raz w czasie każdego potencjału czynnościowego. Występowanie okresu całkowitej refrakcji zapewnia rozprzestrzenianie się potencjału czynnościowego w jednym tylko kierunku.
Kanał sodowy ma wbudowany wyłącznik czasowy, określający czas przepływu jonów.
Wyszukiwarka
Podobne podstrony:
35 39
35 39
JW 28 34 transport1
35 39
35 39
03 1996 35 39
02 1996 35 39
35 39
35 39
35 39
03 1996 35 39
35 39 (3)
02 1996 35 39
02 1996 35 39
39-ręczny transport surowców, Instrukcje BHP, XXXIV - BUDOWLANKA
FOR ostrzega 35 Reorganizacja sadow odroczona MK JW
więcej podobnych podstron