Tematyka obowiązująca do egzaminu z kursu MECHANIZMY EWOLUCJI 2008/2009
1. Znaczenie doboru sztucznego w teorii ewolucji Darwina
W połowie XIX wieku panował powszechnie pogląd, że organizmy nie zmieniają się z czasem w sposób istotny i wyglądają tak samo jak w dniu stworzenia. Były wszak pewne wyjątki przemawiające przeciwko temu poglądowi. Jednym z nich okazała się umiejętność hodowców uzyskiwania w ciągu kilku pokoleń ogromnej różnorodności odmian i ras udomowionych zwierząt i roślin. Osiągano to wskutek selekcji określonych cech i krzyżowania jedynie tych osobników, które te cechy posiadały. To postępowanie określa się mianem doboru sztucznego. Zjawisko to wywarło znaczny wpływ na Darwina.
2. Znaczenie pojęcia ewolucji
3. Warunki konieczne dla przebiegu ewolucji
Według Darwina dla istnienia doboru naturalnego potrzebne są cztery cechy:
- nadprodukcja - liczba wydawanego potomstwa jest większa niż liczba osobników osiągających zdolność do reprodukcji
- zmienność - w potomstwie występują różnice międzyosobnicze
- współzawodnictwo - organizmy konkurują ze sobą o ograniczone, dostępne dla nich zasoby
- przeżyć aby się rozmnażać - osobniki odznaczające się najkorzystniejszymi cechami mają największą szansę przeżycia i reprodukcji.
4. Czynniki doboru naturalnego, ich różnorodność
5. Znaczenie pojęcia dostosowania
Dostosowanie to zdolność organizmu wynikająca z jego genotypu, do skutecznego współzawodnictwa i przekazywania genetycznych zasobów następnym pokoleniom. Określamy symbolem W w przedziale 0-1. Odwrotnością jest współczynnik doboru s.
6. Adaptacja i specjacja jako skutki działania doboru naturalnego
7. Cechy nabyte nie mogą być materiałem ewolucji
Możliwość dziedziczenia cech nabytych odrzucił wielki teoretyk biologii, August Weismann (1834-1914). Genetycy od początku zaprzeczyli możliwości dziedziczenia zmian modyfikacyjnych, dowodząc, że tylko zmiany zachodzące w materiale genetycznym są przekazywane z pokolenia na pokolenie, zmiany te zaś, czyli mutacje, mają charakter losowy, a nie przystosowawczy. Fenotyp powstaje w wyniku skomplikowanych współdziałań między czynnikami genetycznymi i środowiskowymi i dlatego uformowanej już postaci i zachowania się osobnika nie można powrotem przełożyć na kod białkowy.
8. Dobór jako proces śródpokoleniowy, ewolucja jako proces międzypokoleniowy
9. Kumulatywny charakter procesu ewolucyjnego
Kumulatywny charakter doboru polega na tym, że dobór w każdym pokoleniu „faworyzuje” fenotypy korzystne dla organizmu. Takie stopniowe gromadzenie cech korzystnych może po pewnym okresie czasu doprowadzić do stworzenia cech bardziej skomplikowanych - np. skomplikowanych organów. Przykładem są małpy piszące dzieło Shakespear'a. Pamiętać jednak należy, że dobór nie wybiera ze wszystkich wyników ale ulepsza je krok po kroku, zachowując w kolejnych pokoleniach te najbardziej sensowne.
10. Proces ewolucyjny ma charakter oportunistyczny
11. Mutacje jako podstawowe źródło zmienności, losowy charakter mutacji
Mutacje to podstawowe źródło zmienności. Mutacje są spontaniczne i przypadkowe. Mutacje w komórkach somatycznych nie są losowe. Mutacje jako zdarzenia losowe nie muszą być zgodne z kierunkiem ewolucji.
12. Zmienność genotypowa i fenotypowa
Zmienność fenotypowa (niedziedziczna ) to fluktuacyjna (modyfikacje i fenokopie) polega na zdolności wytwarzania przez organizmy o tym samym genotypie różnych fenotypów w zależności od warunków środowiska, w których następuje rozwój osobników.. Modyfikacje mają zazwyczaj charakter nietrwały i zależą od czynników środowiska oraz plastyczności genotypu. Jeśli fenotyp jakiegoś genotypu zostaje zmieniony przez warunki środowiska w ten sposób, że przypomina fenotyp innego genotypu, to organizm ten nazywamy fenokopią.
Zmienność genotypowa (dziedziczna) to rekombinacyjna (zygotyczna) i mutacyjna. Czyli występowanie różnic w materiał genetycznym pomiędzy osobnikami. Przez rekombinacje genetyczną rozumiemy każdy proces, w którego wyniku kontakt dwojga organizmów lub komórek prowadzi do powstania osobników rodzicielskich. Rekombinacje nie prowadzą do wytworzenia nowych alleli, ale do ciągłego ich przetasowania i powstawania różnych genotypów. U organizmów eukariotycznych rekombinacja może nastąpić przez mejozę lub w wyniku losowego doboru osobników rodzicielskich i losowego łączenia się gamet w czasie zapłodnienia.
Zmienność mutacyjna prowadzi do powstawania nowych genów, odrębnych układów genów w chromosomach czy wreszcie odrębnych układów chromosomalnych.
13. Komponenty zmienności fenotypowej, normy reakcji
14. Zmienność ciągła i nieciągła (skokowa)
Zmienność można podzielić na nieciągłą, gdy dla danej cechy da się wydzielić 2 lub więcej dobrze odgraniczonych klas lub ciągłą, jeśli pod względem badanej cechy osobniki trudno podzielić na wyraźne klasy.
Zmienność nieciągła charakteryzuje zwykle cechy jakościowe i nosi nazwę polimorfizmu. Przykładem są grupy krwi AB0 u człowieka.
Zmienność ciągłą mają cechy o charakterze ilościowym, jak np. rozmiary ciała. Cechy ciągłe mają zwykle skomplikowane podłoże genetyczne uwarunkowane przez wiele genów, dlatego noszą nazwę cech poligonowych. Stopień zmienności takich cech opisuje się takimi parametrami, jak zakres zmienności (wartość najmniejsza i największa), średnia 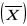
i wariancja (V). Wariancje oblicza się ze wzoru:
gdzie N - liczba badanych osobników, Xi - wartość cechy zmierzona u każdego z nich, 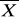
- średnia. Wariancja określa jak bardzo poszczególne wartości odbiegają od średniej . Jest zatem miarą stopnia zmienności; im większa wariancja, tym większa zmienność.
15. Różnica selekcyjna i reakcja na dobór
Selekcja może być stabilizująca - prowadzi do zmniejszenia zmienności, faworyzując osobniki zbliżone do średniej rozkładu normalnego, kosztem osobników o wartościach skrajnych. Selekcja kierunkowa - jeden fenotyp zastępuje stopniowo drugi, powodując selekcję o charakterze jednokierunkowym, np. w kierunku większych rozmiarów. Przykład to Biston betularia. Selekcja rozdzielająca wyróżnia fenotypy skrajne. Działa w kilku kierunkach, prowadząc do dywergencji, czyli wydzielania w obrębie populacji odrębnych grup populacji. Przykład to Zięby z Galapagos.
16. Odziedziczalność, metody oszacowania
Odziedziczalność (h2, heritability) rozumiane w wąskim znaczeniu określa udział wariancji addytywnej w wariancji fenotypowej:
Pojęcie to pozwala określić, jak duża część zmienności ma podłoże genetyczne, a zatem może zostać odziedziczona w następnych pokoleniach.
Odziedziczalność nie określa liczby genów , pozwala natomiast ocenić, jaka część obserwowanej zmienności ma podłoże genetyczne, przez co daje możliwość porównywania komponentów genetycznych różnych cech. Odziedziczalność jest różna w różnych środowiskach, a także dla różnych populacji, gdyż zależy od stopnia ich heterozygotyczności. Na przykład w linii samopylnych roślin wszystkie osobniki są homozygotyczne i identyczne genetycznie, wobec tego zarówno VG = 0, jak i h2 = 0, czyli ich zmienność fenotypowa wynika tylko ze zmienności niegentycznej.
17. Dobór naturalny w populacjach zięb Darwina
Te brunatne, nie wyróżniające się urodą ptaki wywodzą się od wspólnego przodka, którym był gatunek ptaków nasionożernych z Ameryki Południowej. Czternaście znanych gatunków tych ptaków wykształciło całkowicie odmienne style życia, charakterystyczne dla wielu gatunków, które żyją w innych rejonach świata i nigdy nie miały możliwości zasiedlenia Wysp Galapagos. Prawdopodobieństwo pochodzenia tych różnorodnych ptaków od wspólnego przodka nasunęło Darwinowi myśl, że gatunki powstają w wyniku doboru naturalnego.
18. Zmiany kierunku działania doboru w czasie, przykład krępaka brzozowego Biston betularia
Badania nad tą ćmą prowadził w Anglii H.B.D. Kettlewell. W roku 1850 większość drzew na Wyspach pokryte było białym porostem. Ćmy o jasnym ubarwieniu były lepiej dostosowane, a ptaki wyłapywały ćmy ciemne (ten sam gatunek, inna odmiana). Uprzemysłowienie wielu rejonów spowodowało zniknięcie porostów i pociemnienie kory drzew. Wtedy bardziej widoczne dla ptaków stały się ćmy jasne. W rejonach uprzemysłowionych 90% osobników stanowi odmiana ciemna a w rejonach miejskich jest na odwrót.
19. Efekty kojarzeń wsobnych, ich przyczyny; ewolucja mechanizmów zapobiegających kojarzeniom wsobnym
20. Klasyczny i równowagowy model zmienności genetycznej
21. Polimorfizm genetyczny
Polega na utrzymaniu się w populacji więcej niż jednego allelu w tym samym locus. Jeśli co najmniej jeden z alleli utrzymuje się w częstości >1% to mamy już do czynienia z polimorfizmem.
22. Polimorfizm przejściowy i zrównoważony
P. przejściowy występuje gdy zmieniają się warunki ekologiczne i jeden allel eliminowany jest
przez inny w wyniku doboru. Także w trakcie utrwalania się mutacji neutralnych.
23. Naddominacja; inne czynniki utrzymujące i redukujące zmienność genetyczną populacji
Naddominacja występuje wtedy, gdy fenotyp heterozygoty jest bardziej skrajny niż fenotypy obu homozygot. Na przykład w przypadku allelu anemii sierpowatej (HbS) u homozygot HbSHbS występuje anemia prowadząca do śmierci już w młodości, a homozygoty normalne Hb+ Hb+ s bardziej podatne na malarię niż heterozygoty Hb+HbS; te ostatnie mają więc największe szanse przeżyci i pozostawienia potomstwa.
24. Dobór zależny od częstości, dobór apostatyczny
Dobór zależny od częstości może działać na dwa sposoby
Faworyzujący formy rzadsze ( dobór apostatyczny)
Faworyzujący formy częstsze
W stanie równowagi dostosowanie W wszystkich genotypów jest identyczne, a spada po odchyleniu od stanu równowagi. Fenotyp występujący częściej ma niższe dostosowanie, co ostatecznie musi doprowadzić do polimorfizmu zrównoważonego.
25. Podstawowe modele genetyki populacji: pojęcie puli genowej, frekwencja genotypów i alleli
26. Frekwencje genotypów jako funkcje frekwencji alleli
27. Równowaga Hardy-Weinberga, czynniki zaburzające równowagę H-W
Prawo Hardy'ego-Weinberga określa częstość występowania genotypów w całej swobodnie krzyżującej się populacji. Zgodnie z tym prawem sam proces dziedziczenia nie powoduje w dużych populacjach zmian częstości występowania alleli, utrzymującej się na stałym poziomie z pokolenia na pokolenie. Frekwencja genotypów w potomstwie (frekwencja BB + Bb + bb):
Muszą być spełnione następujące warunki aby równowaga nie była zabużona:
- organizmy są diploidalne
- pokolenia nie zachodzą na siebie
- osobniki kojarzą się losowo (populacja jest panmiktyczna)
- populacja jest bardzo duża
- dobór naturalny nie wpływa na badany locus.
28. Populacja panmiktyczna, efekt Wahlunda
29. Tempo zmian ewolucyjnych zależy od wariancji częstości alleli i dostosowania genotypów; współczynniki selekcji
30. Ograniczona wielkość populacji jako główny czynnik losowych zmian genetycznych w populacjach
31. Losowość łączenia się gamet i losowość przeżywania osobników jako przyczyny zmian losowych w populacjach
32. Dryf genetyczny, jego kumulacyjny charakter
Powstanie losowych zmian ewolucyjnych w małej rozmnażającej się populacji określono mianem dryfu genetycznego. Powoduje on zmiany puli genów populacji z pokolenia na pokolenie. Wpływa losowo na zmiany frekwencji genów.
33. Ewolucja bez działania doboru naturalnego
34. Wpływ dryfu genetycznego na stopień zmienności śródpopulacyjnej i stopień zróżnicowania między populacjami
Z jednej strony dryf genetyczny może obniżać zmienność genetyczną w obrębie populacji, z drugiej strony zwiększać zmienność między lokalnymi populacjami.
35. Procesy losowe w czasie kolonizacji nowych obszarów, efekt założyciela
Zasada założyciela sprawia, że izolowane populacje mogą się znacznie różnić pod względem frekwencji alleli od populacji tego gatunku żyjących gdzie indziej, przy czym ta odmienność jest raczej przypadkowa niż przystosowawcza. Przykładem jest brak grupy B w krwi Aborygenów - mała populacja założycielska nie miała allelu IB.
36. Zależność tempa utraty zmienności genetycznej z populacji od wielkości populacji i wyjściowej zmienności
37. Tempo zastępowania alleli neutralnych
Przy częstości mutacji wynoszącej u na allel na pokolenie, w każdym pokoleniu spodziewać się należy 2Nu mutacji na populację (czyli w mutacji 500tys. osobników - 10 mutacji na pokolenie). Ponieważ jest to proces ciągły trwający tysiące pokoleń, to liczba mutacji, które w każdym pokoleniu powstają (2Nu) i prawdopodobieństwa ich utrwalenia 1/(2N), czyli zastąpienia istniejących tam alleli przez allel zmutowany. Wynika z tego, że zastępowanie jednych neutralnych alleli przez nowo powstałe mutacje odbywa się w tempie:
na populację na pokolenie, czyli jest równe częstości mutacji na pokolenie na jeden allel.
Czyli utrwalenie mutacji występuje z częstością raz na 100 tysięcy pokoleń ale proces utrwalania może mieć różny czas.
38. Teoria neutralnych mutacji Kimury
Teoria Kimury tłumaczy losy mutacji i alleli neutralnych. Allele te mogą się wymieniać w populacji w tempie równym częstości mutacji. W dużej populacji mutacje pojawiają się często:
przyjmuje się u = 10-5 a więc w populacji 500 tys. osobników mutacja pojawi się 10 razy w każdym pokoleniu - bo mamy milion alleli. W małych populacjach pojawia się o wiele rzadziej. Natomiast czas utrwalenia - równy 4X liczbie osobników - w dużych populacjach jest o wiele dłuższy. W małej populacji jeden z alleli utrwala się szybko co tłumaczy homogeniczność populacji niewielkich.
39. Charakter zmienności genetycznej na poziomie molekularnym w świetle teorii Kimury
Zmienność genetyczna jest tym niższa im mniejsza jest populacja. Teoria ta wyjaśni utrzymywanie się więcej niż jednego allelu na jednym locus w populacjach dużych, czyli heterogeniczność tych populacji oraz homogeniczność populacji małych.
40. Pozytywny związek między stopniem zmienności genetycznej a tempem ewolucji molekularnej
41. Obserwacje potwierdzające znaczący udział procesów losowych w ewolucji molekularnej
Szczególną cechą zmian ewolucyjnych zachodzących na poziomie molekularnym jest pozytywna korelacja miedzy czasem dywergencji (rozszczepienia linii ewolucyjnej) dwóch grup a stopniem zróżnicowania ich białek i różnych odcinków DNA. Taką korelacje uzyskuje się zestawiając czas rozszczepienia linii filogenetycznych dla pary porównywalnych taksonów, uzyskamy z danych paleontologicznych, ze stopniem zróżnicowania (sekwencja DNA) molekularnego współczesnych form należących do tych taksonów.
Zegar ten jest zegarem stochastycznym tzn., takim w którym prawdopodobieństwo zmian jest stałe, ale tempo zmian w krótkich odcinkach czasowych może być zmienne.
43. Zróżnicowanie wielkości genomu, poliploidyzacja
44. Mechanizmy duplikacji tandemowych
45. Różnicowanie się nowych genów po duplikacji, rodziny genów
Ewolucja nowego genu często zaczyna się od duplikacji, czyli podwojenia już istniejącego genu. Taka dodatkowa kopia może akumulować mutacje, podczas gdy oryginał funkcjonuje bez zmian. Z czasem ten proces doprowadza do wytworzenia nowego genu. Na przykładzie beta-globin . Opisane geny powstały prawdopodobnie w wyniku duplikacji genu kodującego mioglobinę. Kolejne duplikacje i zmiany w sekwencji doprowadziły do powstania alfa i beta - globin, uczestniczących w wytwarzaniu różnych form hemoglobiny w kolejnych stadiach rozwoju ssaków.
46. Nietandemowe powtórzenia w genomie, mechanizmy powstawania.
47. Hipoteza symbiotycznego pochodzenia organelli komórkowych
48. Mechanizmy specjacji, ewolucja izolacji rozrodczej
Izolacja rozrodcza zachodzi gdy populacje lokalne, w procesie przystosowania się do warunków miejscowych, zaczynają być dalece odmienne. Istnieją dwie grupy mechanizmów izolacji rozrodczej - prezygotyczna (zapobiega zapłodnieniu) i postzygtyczna. Mechanizmy izolacji prezygotycznej obejmują izolację sezonową, siedliskową, (ekologiczną), etologiczną, mechaniczną oraz izolację gametyczną. Mechanizmy izolacji postzygotycznej prowadzą do upośledzenia potencjału reprodukcyjnego.
Proces powstawania gatunku określa się mianem specjacji. Niezbędnym czynnikiem dla specjacji jest izolacja rozrodcza. Jeśli między osobnikami nie dochodzi do wymiany informacji genetycznej nawet w razie spotkania się możemy uznać, że specjacja się dokonała.
49. Specjacja allo- i sympatryczna.
Specjacja allopatryczna - specjacja będąca skutkiem geograficznego oddzielenia się jednej populacji od reszty gatunku i następującego potem procesu ewolucji, nazywana też specjacją geograficzną.
Populacja sympatryczna - gdy populacja tworzy nowy gatunek w obrębie wspólnego dla niej i populacji macierzystej obszaru geograficznego.