Mięśnie zbudowane są z komórek mięśniowych zawierających liczne równolegle ułożone filamenty aktyny i miozyny
Filamenty aktynowe zawierają dwie skręcone cząsteczki aktyny
filamenty miozynowe- zbudowane są z wielu cząsteczek miozyny
podczas skurczu mięśnia filamenty aktynowe i miozynowe wślizgują się między siebie
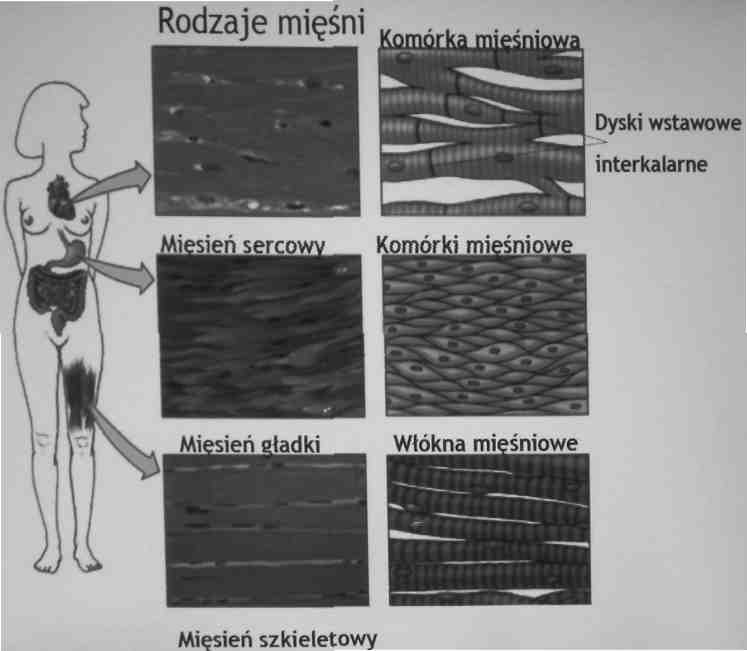
rozróżniamy trzy rodzaje mięśni: gładkie, sercowe i szkieletowe
Mięsnie gładkie-skurcze narządów wewnętrznych, kontrolowane przez autonomiczny układ nerwowy
komórki mięśni gładkich - strukturalnie najprostsze, posiadają jedno jądro komórkowe
mięśnie sercowe-regularne rozmieszczenie miozyny i aktyny (prążkowanie), wzmocnione dyskami wstawnymi (interkalamymi)
mięśnie szkieletowe (prążkowane)- ruchy dowolne, komórki mięśni szkieletowych zawierają wiele jąder
UKŁAD MIĘNIOWY CZŁOWIEKA
-składa się z 650 mięśni
-ich masa stanowi ok. 30-40% masy kobiet, 40-50%masy mężczyzn
BUDOWA ANATOMICZNA MIĘŚNIA
-tkanka łączna wytwarzana na powierzchni mięśnia zwana jest omięsną
-w 1 mm3 mięśnia znajduje się 2000 naczyń krwionośnych włosowatych
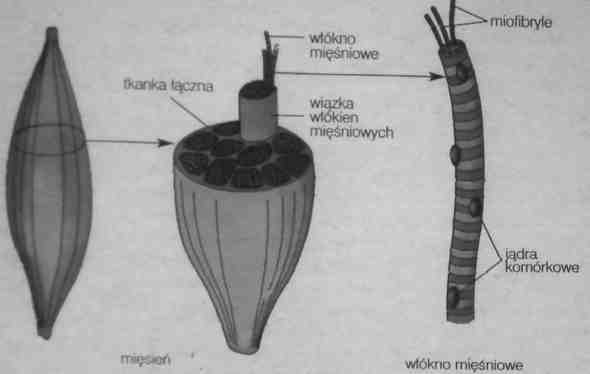
Mięśnie szkieletowe - budowa
1 Pojedyncze włókno mięśniowe to komórka mięśniowa.
Komórka mięśniowa składa się błony komórkowej (sarkolemmy) oraz wiązki miofibryli, zbudowanych z długich struktur (filamenty białkowe) miozyny i aktyny.
Przestrzeń między miofibrylami wypełnia sarkoplazma. Siateczka sarkoplazmatyczna gromadzi i uwalnia jony Ca2+.
2. Podstawową jednostka funkcjonalną miofibryli jest sarkomer, składający się z miozyny i aktyny.
3. Jednostka motoryczna składa się z neuronu ruchowego (motoneuronu), i wszystkich włókien mięśniowych przez
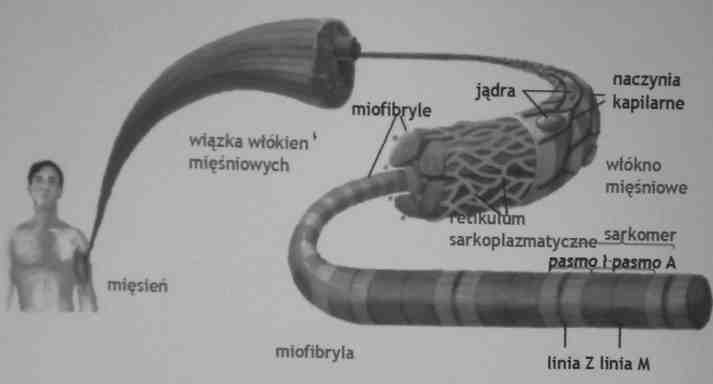
niego unerwionych.
Budowa mięśnia szkieletowego
Retikulm sarkoplazmatyczne: system płaskich membranowych pęcherzyków otaczający
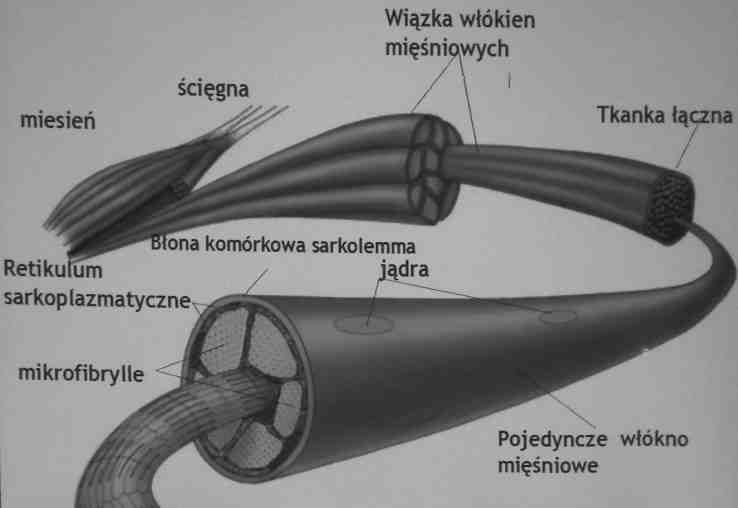
każdą miofibrylę.
podjednostka zdolna do skurczu, złożona z wiązek cienkich i grubych filamentów
Budowa sarkomeru
Każde włókno mięśniowe zbudowane jest z równolegle ułożonych wiązek wiązek miofibryli
miofibryle zawierają powtarzające się jednostki zwane
sarkomerami, które są ograniczone liniami Z
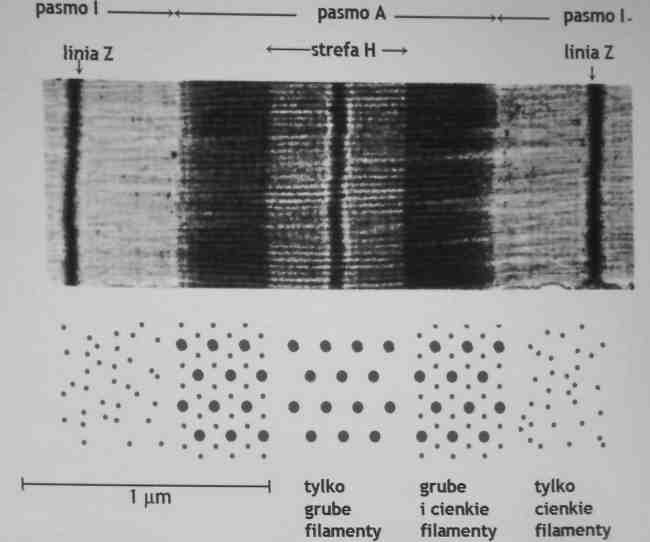
w obrębie sarkomeru występują naprzemiennie ciemne pasmo A i jasne pasmo I
Pasmo A zawiera grube filamenty zbudowane z miozyny i cienkie filamenty zbudowane z aktyny, troponiny i tropomizyny
w strefie H i paśmie I filamenty nie zachodzą na siebie dlatego są jaśniejsze
Linia M zawiera białka podtrzymujące filamenty miozynowe
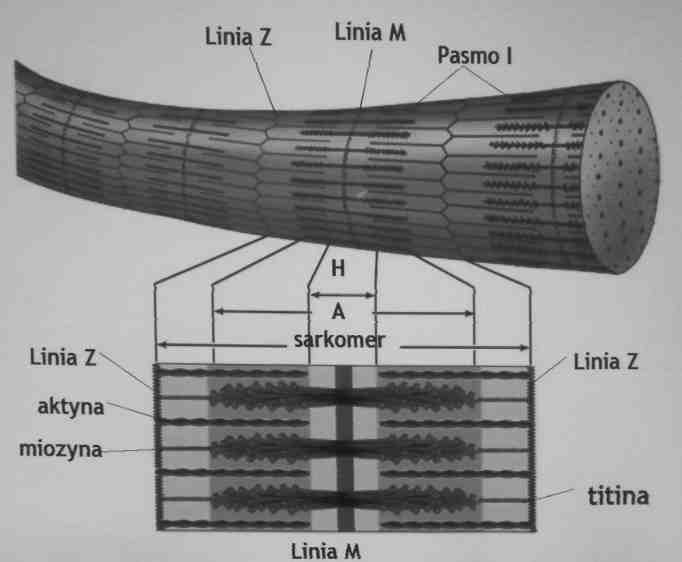
Budowa miofibryli
Wiązki filamentów miozynowych są utrzymywane w centralnej pozycji sarkomeru przez białko zwane
titiną
Każda cząsteczka titiny biegnie przez wiązkę miozyny od linii Z do Z, jest to bardzo elastyczne białko odporne na rozciąganie
Budowa sarkomeru
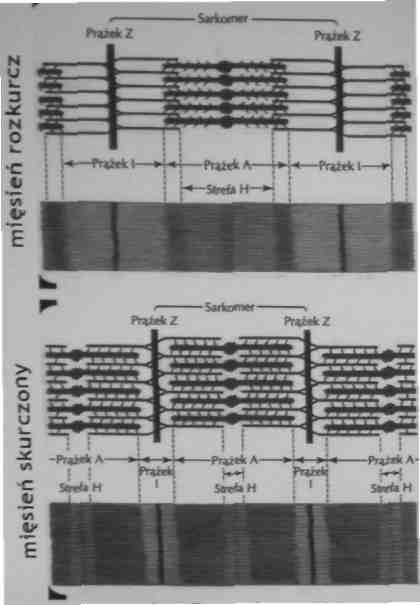
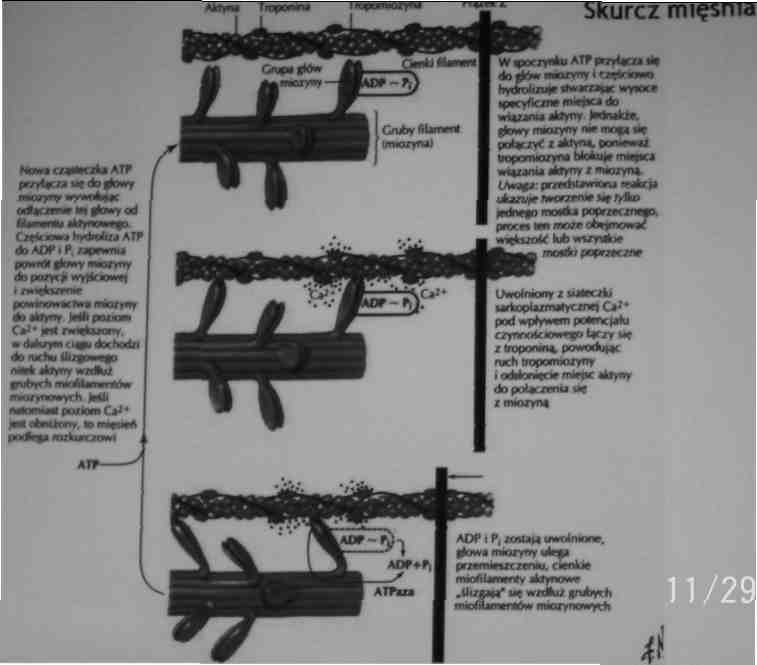
Mechanizm skurczu
Podczas skurczu mięśnia filamenty grube (miozyna) i cienkie (aktyna) wślizgują się miedzy siebie powodując zmniejszenie długości sarkomeru pasma H i I zwężają się a linie Z zbliżają się do pasma A - model wślizgiwania się filamentów
Skurcz i rozkurcz mięśniowy
Podczas skurczu mięśniowego cienkie filamenty wślizgują się pomiędzy grube, powodując zbliżenie pomiędzy prążkami Z i skracanie sarkomeru.
Prążki A pozostają tej samej długości, prążki I skracają się.
Strefy H ulegają zwężeniu lub znikają.
Skurcz mięśnia jest spowodowany ślizganiem się filamentow aktynowych po filamentach miozynowych.
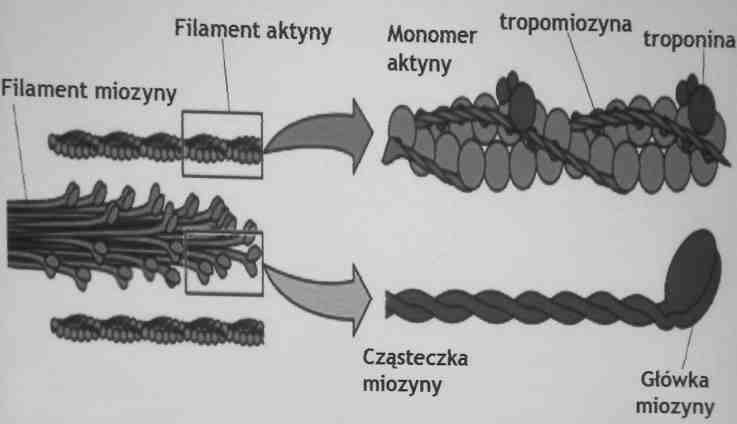
Budowa filamentów aktyny i miozyny
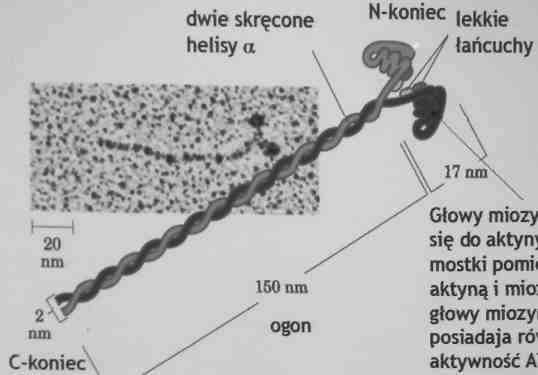
Miozyna- białko motoryczne
Białka motoryczne- działają przez przekształcanie zmian w konformacji białek w ukierunkowany ruch
Miozyna (M.cz. 540 000); 2 ciężkie łańcuch (M.cz. 220 000) i 4 lekkie łańcuchy (M.cz. 20 000)
Głowy miozyny wiążą się do aktyny i tworzą mostki pomiędzy aktyną i miozyna głowy miozyny posiadają również aktywność ATP-azawą
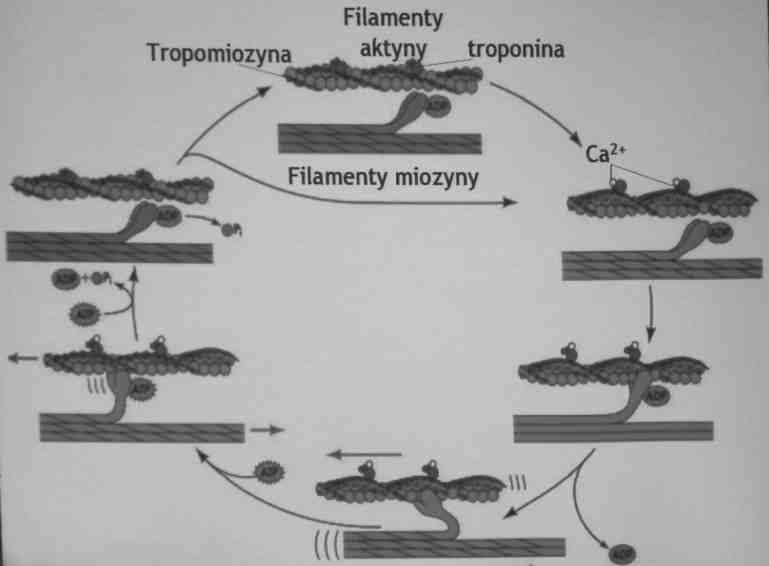
Uwolnienie Ca2+ powoduje skurcz mięśnia
Mięśnie szkieletowe - mechanizm skurczu
1.Czynnik zapoczątkowujący skurcz: neurotransmiter z motoneuronu (acetylocholina). Acetylocholina uwolniona przez neuron wiąże się z receptorem w komórce mięśniowej, co powoduje otwarcie kanału sodowego > depolaryzacja błony.
2. Depolaryzacja powoduje uwolnienie jonów Ca2+ z pęcherzyków siateczki sarkoplazmatycznej do sarkoplazmy.
3. Jony Ca2+ wiążą się do troponiny C znajdującej się na aktynie => odsłonięcie miejsc wiązania miozyny na filamencie aktyny.
4.Główki miozynowe mogą wiązać się z aktyną, następuje uwolnienie ADP
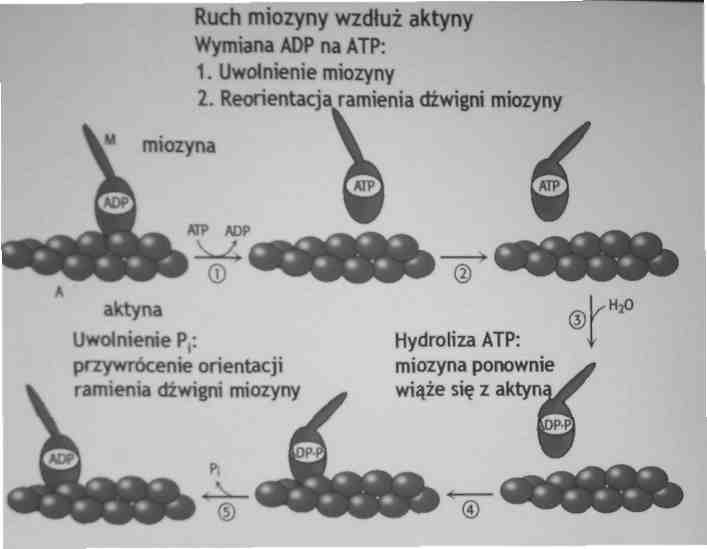
5. Skurcz mięśni szkieletowych występuje wskutek wnikania (ślizgania) cząsteczek aktyny między cząsteczki miozyny. Związanie ATP do miozyny, powoduje jej uwolnienie od aktyny. Hydroliza ATP do ADP powoduje zmianę konformacyjną miozyny i może ona ponownie łączyć się z aktyną powodując następny skurcz.
6. Skurcz mięśnia następuje tak długo, dopóki jony Ca2+ znajdujące się w sarkoplazmie nie zostaną wychwycone przez siateczkę sarkoplazmatyczną (za pośrednictwem pompy wapniowej zal. do ATP)
•ATP jest potrzebne aby zerwać wiązanie pomiędzy aktyną i miozyną
•energia jest potrzebna do zatrzymania skurczu mięśnia
•Brak ATP po śmierci powoduje sztywnienie mięśni
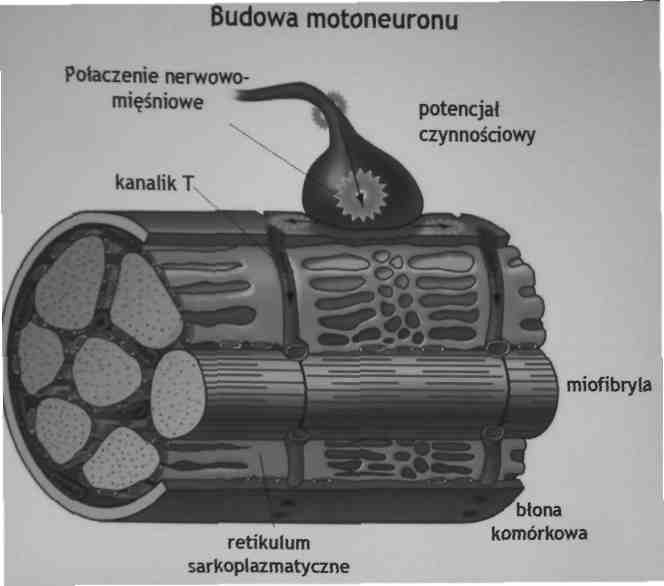
Potencjały czynnościowe napływające do złącza nerwowo-mięśniowego powodują uwalnianie acetylocholiny (ACh) z motoneuronów. Receptor dla acetylocholiny zlokalizowany w błonie komórkowej jest kanałem jonowym, który otwiera się po przyłączeniu ACh. Przyłączenie ACh wzmaga dokomórkowy prąd Na+, odpowiedzialny za depolaryzację błony komórkowej i powstanie potencjału czynnościowego.
Potencjał czynnościowy rozprzestrzenia się wzdłuż włókna mięśniowego do specjalnych wgłębień na błonie komórkowej, zwanych kanalikami transwersyjnymi (T).
Kanalik T jest zlokalizowany w sąsiedztwie cysterny brzeżnej siateczki sarkoplazmatycznej (SR).
Rozprzestrzeniające się fala depolaryzacji (wzdłuż kanalika T) I powoduje uwolnienie jonów Ca2+ z siateczki sarkoplazmatycznej (SR) do (I cytoplazmy.
Inne fragmenty SR, zawierające Ca2+-ATPazę, są odpowiedzialne za ponowne zgromadzenie jonów Ca2+, umożliwiając zakończenie skurczu.
W komórkach mięśnia gładkiego nie występują miofibryle. Filamenty aktynowe są zakotwiczone w Wonie komórkowej, a ponadto są przymocowane do ciałek gęstych w obrębie cytoplazmy.
Interakcja filamentów aktynowych z miozynowymi powoduje skurcz mięśniówki gładkiej.
Komórki mięśni gładkich nie zawierają kanalików T: ich odpowiednikiem są kaweole.
Kaweole są organellami, w których gromadzi się Ca2+napływający do komórki.
Mięśnie gładkie-mechanizm skurczu
• W mięśniach gładkich Ca2+ uwalniany z siateczki sarkoplazmatycznej do sarloplazmy łączy się z białkiem zwanym kalmoduliną
•Kompleks Ca-kalmodulina aktywuje kinazę miozynową, która fosforyluje główki miozyny.
•Chemicznie aktywowana miozyna rozpoczyna cykl wiązania i uwalniania aktyny, powodując skurcz mięśni.
•Fosfataza miozynową defosforyluje miozynę i zatrzymuje interakcję aktyna-miozyna
Sprzężenie pobudzeniowo- mechaniczne w mięśniach gładkich
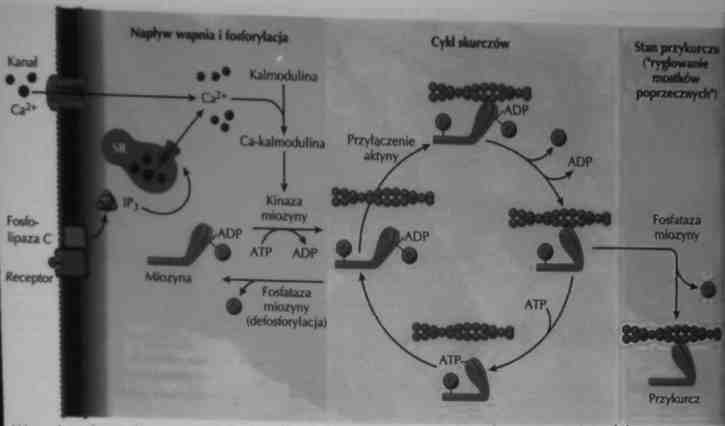
W reakcji kurczliwości mięśni gładkich uczestniczy zarówno Ca2+ zewnątrzkomórkowy,
jak i Ca2+ uwalniany z siateczki sarkoplazmatycznej.
Wysokie stężenie Ca2+wiązanie kalmoduliny => kinaza miozynowa fosforyluje miozynę =>
ślizganie się miozyny po aktynie.
Defosforylacja miozyny > spowolnienie cyklu skurczów => mechanizm „zatrzasku"
mostków poprzecznych (skurcz toniczny bez hydrolizy ATP).
Mięsień sercowy, podobnie jak mięsień szkieletowy, wykazuje poprzeczne prążkowanie (wynik regularnego rozmieszczenia aktyny i miozyny).
Błona komórkowa komórki mięśnia sercowego (kardiomiocyta) zawiera kanaliki transwersyjne (T), podobnie jak w mięśniach szkieletowych.
Różna jest budowa retikulum sarkoplazmatycznego: występują diady zamiast triad pomiędzy kanalikami T a siateczką sarkoplazmatyczną.
Skurcz mięśnia szkieletowego zależy od uwalniania Ca2+ z siateczki sarkoplazmatycznej, a zmiany w zewnątrzkomórkowym
stężeniu Ca2+ nie wpływają na siłę skurczu.
W kardiomiocycie, usunięcie Ca2+ z płynu zewnątrzkomórkowego zmniejsza siłę skurczu.
Sumacja: wzrost napięcia we włóknie mięśniowym wskutek wzrostu częstości pobudzeń
Bodziec
Wzrost częstości pobudzeń pojedynczego włókna mięśniowego powoduje wzrost napięcia w tym włóknie i prowadzi do sumacji, która jest wynikiem podwyższenia śródkomórkowego stężenia jonów Ca2+. Wytwarzane jest maksymalne napięcie zwane tężcowym.
Czas trwania skurczu tężcowego zależy od dostarczenia ATP.
Sumacja jest wynikiem podwyższenia śródkomórkowego stężenia jonów Ca2+, wynikającego ze zwiększonej częstości stymulacji tego włókna.
Rodzaje skurczów mięśni szkieletowych
Ze względu na częstotliwość pobudzeń:
- skurcz pojedynczy: pobudzenie (potencjał czynnościowy 5 ms) skurcz 7.5 - 120 ms
- skurcz tężcowy niezupełny: przerwa między bodźcami krótsza niż cały okres skurczu (pobudzenie następuje w momencie, kiedy mięsień zaczął się rozkurczać).
- skurcz tężcowy zupełny: przerwa między bodźcami krótsza niż okres kurczenia się mięśnia, a dłuższa niż okres jego refrakcji (następne pobudzenie na wstępującym ramieniu skurczu)
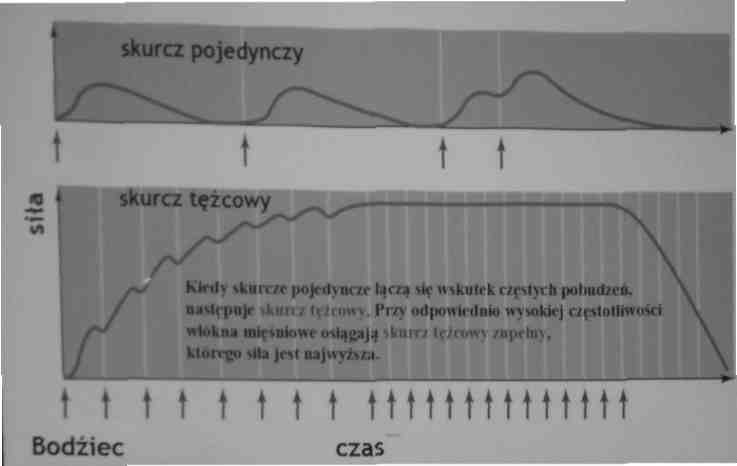
Najczęściej występującym rodzajem skurczów u człowieka jest skurcz tężcowy niezupełny
Rodzaje skurczów mięśni szkieletowych w zależności od częstości pobudzeń
Rodzaju skurczów e względu na rodzaj pracy:
•skurcz izotoniczny: napięcie mięśnia (tonus) nie zmienia się, natomiast długość mięśnia ulega zmianie:
skurcz koncentryczny: mięsień ulega skróceniu;
- skurcz ekscentryczny: mięsień ulega wydłużeniu;
•skurcz izometryczny: długość mięśnia nie zmienia się
•skurcz auksotoniczny: kilka rodzajów skurczów występujących jednocześnie, skurcze mieszane; najczęstsze skurcze podczas codziennej aktywności człowieka
Płynność ruchu zależy od asynchronicznej ( nie są pobudzane jednocześnie) działalności jednostek motorycznych
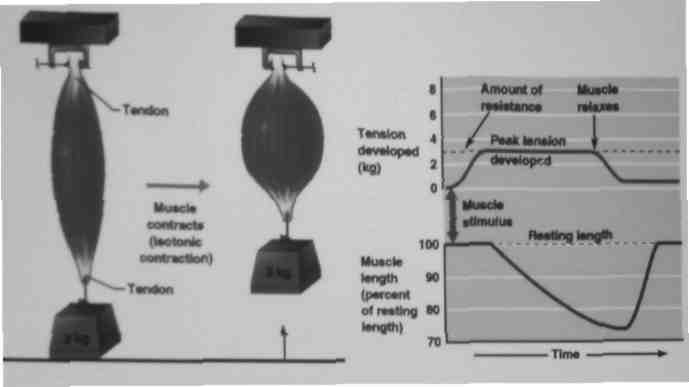
Skurcz izotoniczny- napięcie mięśnia nie zmienia się zmianie ulega długość mięśnia (skróceniu-koncentryczny lub wydłużeniu ekscentryczny)
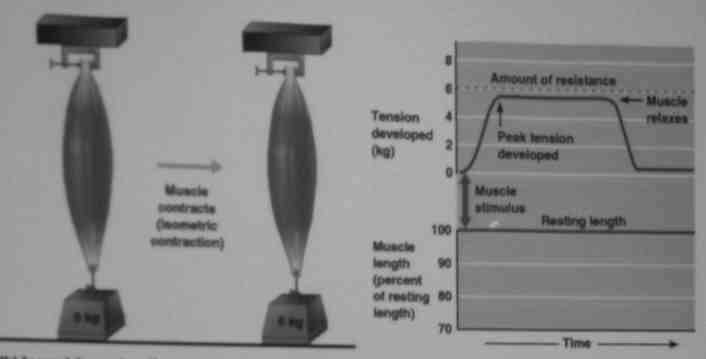
Skurcze izometryczne- mięsień rozwija siłę (napięcie) ale nie zmienia swoje, długości, energia uwalniana podczas skurczu zamienia się na ciepło
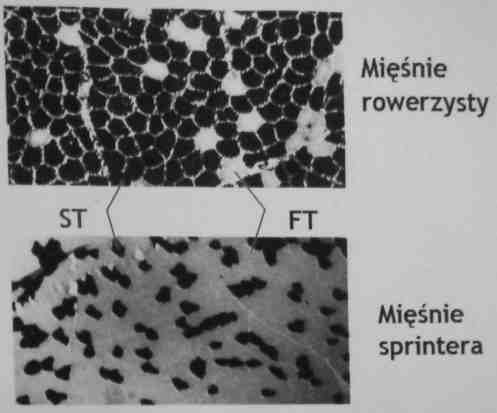
Jednostki motoryczne: FT i ST
PT (Fast Twitch): włókna szybkokurczliwe: kurczą się szybko, prędko się męczą. Mięsnie nie potrzebują dużo tlenu, bo mają wysoką aktywność beztlenową
ST (Slow Twitch): włókna wolnokurczliwe: kurczą się wolno, wytrzymałe na zmęczenie. Mięśnie zawierające włókna ST są czerwone, bo zawierają dużo naczyń krwionośnych.
Rodzaje włókien mięśniowych determinują siłę i
wytrzymałość
ST włókna wolnokurczliwe (czerwone)
• kurczą się wolno, wytrzymałe na zmęczenie
•zawierają dużo mitochondriów i białka wiążącego tlen mioglobiny w celu produkcji ATP
•mięśnie zawierające włókna ST są czerwone, bo zawierają dużo naczyń krwionośnych.
•paliwem energetycznym jest glikogen i tłuszcz
•wysoka wydolność (wytrzymałość) tlenowa (aerobowa), spalają substraty w obecności tlenu
•wykorzystywane w czasie wysiłków wytrzymałościowych o niskiej intensywności ( bieg maratoński, pływanie)
FT włókna szybkokurczliwe (białe)
•kurczą się szybko, prędko się męczą, mają bardzo mało mioglobiny i mitochondriow
•mięsnie nie potrzebują dużo tlenu, bo mają wysoką aktywność beztlenową
•Włókna szybkokurczliwe są wykorzystywane w wysiłkach krótkich wymagających dużej mocy (sprint, podnoszenie ciężarów)
Mięśnie człowieka zawierają włókna ST i FT, ich proporcje w mięśniach szkieletowych są determinowane genetycznie
|
|
|
•szybkość skurczu |
mała |
duża |
•zawartość glikogenu |
mata |
duża |
•zdolność do glikolizy |
mała |
duża |
•odporność na zmęczenie |
duża |
mała |
•typ oddychania |
aerobiczny |
anaerobiczny |
•naczynia kapilarne |
dużo |
mało |
• mitochondria |
dużo |
mało |
•mioglobina |
dużo |
mało |

Włókna wolnokurczliwe (ST) i szybkokurczliwe(FT)
Włókna wolnokurczliwe ST, spotykane w mięśniach odpowiedzialnych za postawę ciała, muszą się kurczyć przez długi czas bez zmęczenia, dlatego energię czerpią z tłuszczy
Włókna wolnokurczące (typu I lub SO) są czerwone
Włókna szybkokurczące mogą być:
-odporne na zmęczenie (tlenowo - glikolityczne, typu Iia (FOG))
-podatne na zmęczenie (glikolityczne, istnieją 2 typy: Ib. i Xii (FG))
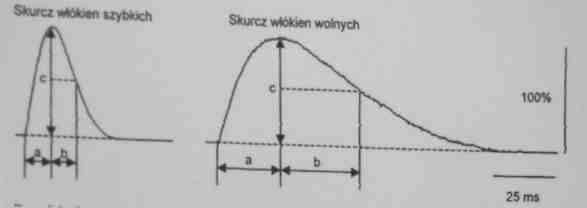
Włókna te są jasne
Włókna szybkokurczące kurczą się szybciej
Szybkość skurczu IIB>IIx>IIa
Zapotrzebowanie na energię: IIb>IIx>IIa
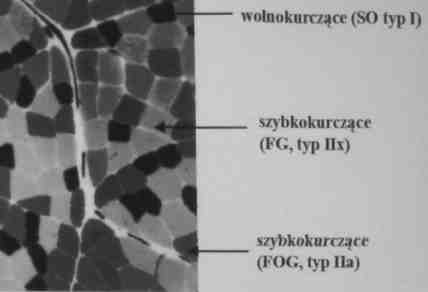
Większość mięśni jest heterogeniczna, czyli zawiera włókna mięśniowe różnych typów
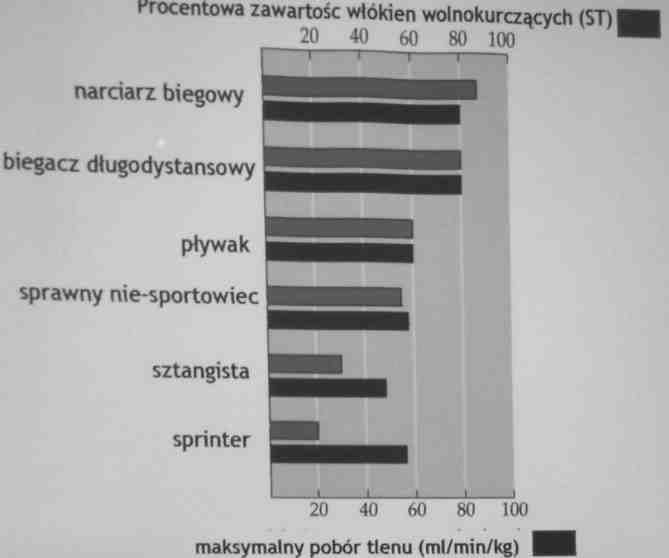
Ze względu na przewagę w danym mięśniu wolnych lub szybkich włókien mięśniowych mięśnie takie nazywane są wolnymi lub szybkimi.
Mięśnie, których czynność w ciągu doby trwa zwykle długo, zawierają przewagę włókien mięśniowych wolnokurczących się.
Osobnicy posiadający w mięśniach dużą ilość włókien wolnokurczących są biologiczne lepiej przygotowani do trwającej długo aktywności (w tym sportów wytrzymałościowych), niż osobnicy, u których jest więcej włókien szybkich.
ZAWARTOŚĆ WŁÓKIEN SZYBKOKURCZLIWYCH I WOLNOKURCZLIWYCH =>
Kenijczycy (wschodnia Afryka) mają dużo wolnokurczących (typ I) włókien w mięśniach, dlatego dominują w biegach długodystansowych.
Mieszkańcy zachodniej Afryki mają dużo szybkokurczących (typ llx i IIb) włókien w mięśniach.
Dlatego sportowcy z USA, gdzie mieszkają potomkowie mieszkańców Afryki Zachodniej, dominują w biegach krótkodystansowych.
Charakter włókien mięśniowych (szybkokurczące lub wolnokurczące) zależy od obecności izoform ciężkiego łańcucha miozyny.
Włókna wolnokurczące:
miozyna MYH7 (występuje w wolnokurczących
włóknach mięsni szkieletowych oraz w sercu)
Włókna szybkokurczące:
MYH1 (we włóknach Xii i FG)
MYH2 (we włóknach Iia i FOG)
MYH4 (we włóknach IIb i FOG)
MYH 3 (tylko u płodów)
W mięśniach gładkich występuje izoform MYH11
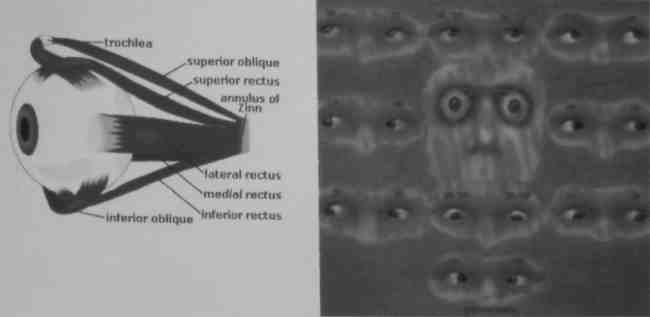
Wyjątkowy rodzaj mięśni stanowią mięśnie gałki ocznej, które muszą być bardzo szybkie i precyzyjne.
Występuje w nich miozyna MYH 13, która nie jest spokrewniona z innymi miozynami.
Są to włókna superszybkie.
Są jedynymi mięśniami, które nie ulegają uszkodzeniu w dystrofiach mięśniowych.
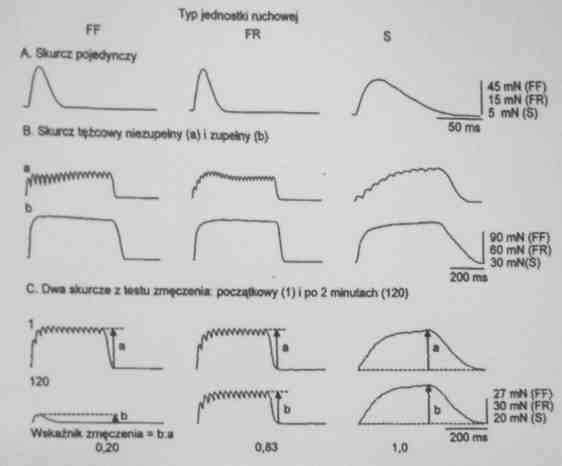
Włókna szybkokurczące można dodatkowo podzielić na odporne na zmęczenie (fast resistant, FR) i męczące się (fast fatigue, FF).
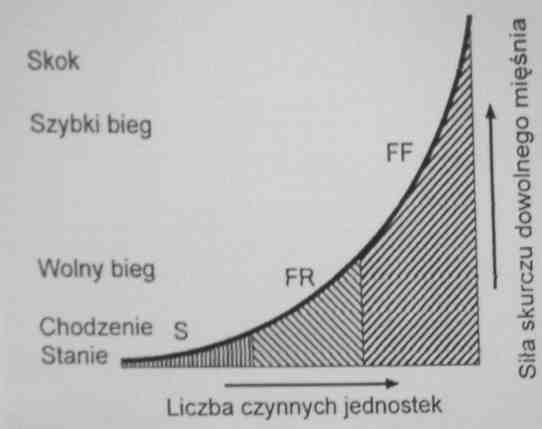
<= W czasie wykonywania pracy przez mięsień najpierw rekrutowane są włókna S, następnie FR, i w końcu FF.
Dlatego chodzenie w porównaniu z biegiem stosunkowo mało męczy.
Wielkość rozwijanej siły zależy od:
-przekroju fizjologicznego;
-początkowej długości mięśnia w czasie pobudzenia;
-liczby aktywnych jednostek motorycznych;
-typu aktywnych jednostek motorycznych (FT czy ST);
-wielkości mięśnia;
-kąta w stawie
-prędkości skracania mięśnia;
-częstotliwości pobudzeń.
Siłę skurczu wyraża się w niutonach (N)
1 niuton to siła z jaką trzeba działać na ciało o masie 1 kg, aby mu nadać przyspieszenie 1m/s2
Przekrój fizjologiczny jest prostopadły do przebiegu włókien
Przekrój fizjologiczny w decydujący sposób wpływa na wielkość siły rozwijanej przez mięsień: im większy przekrój poprzeczny, tym większa jest jego siła.
Na 1 cm przekroju poprzecznego mięsień rozwija 40-50 N.
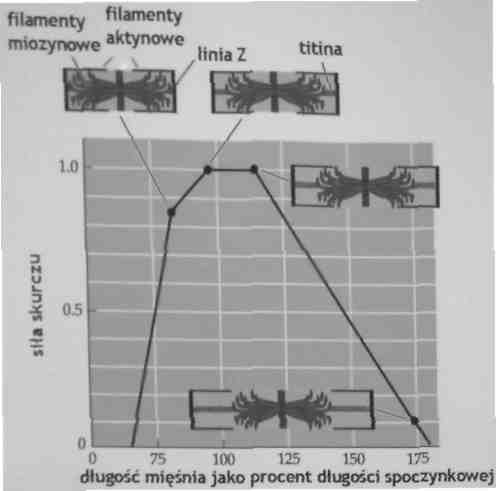
Mięsień rozwija maksymalną siłę, kiedy występuje maksymalna liczba połączeń główek miozynowych z centrami aktyny
Siła rozwijana przez sarkomer w zależności od jego długości
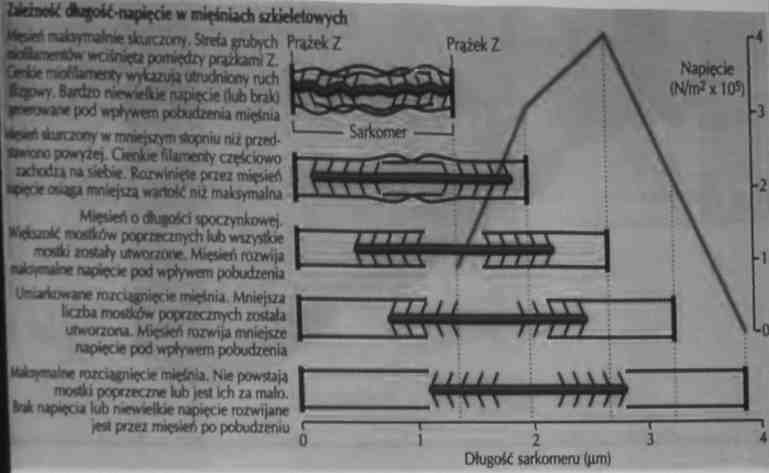
Stopień zachodzącej interakcji filamentów cienkich i grubych jest funkcją długości sarkomeru.
Wzrost siły możemy uzyskać przez wstępne rozciągnięcie mięśnia o 20%
Mięśnie i ich tkanka łączna mają elastyczne i sprężyste właściwości: kiedy są rozciągnięte, mają zdolność gromadzenia energii. W ruchu następującym po tym rozciągnięciu energia jest oddawana (uwalniana), w wyniku czego rośnie siła mięśnia.
Maksymalną siłę można uzyskać, kiedy mięsień zostanie wstępnie rozciągnięty o ok. 20% więcej od jego długości spoczynkowej. W tak rozciągniętym mięśniu kombinacja gromadzenia energii i siła skurczu mięśnia uzyskują optymalne warunki, w wyniku czego uzyskujemy maksymalną siłę.
Każdy staw ma optymalny kąt, w którym mięsień rozwija maksymalną siłę.
Gdy szybkość ruchu wzrasta, to siła maleje.
Zwiększenie częstotliwości pobudzeń zwiększa siłę mięśnia.
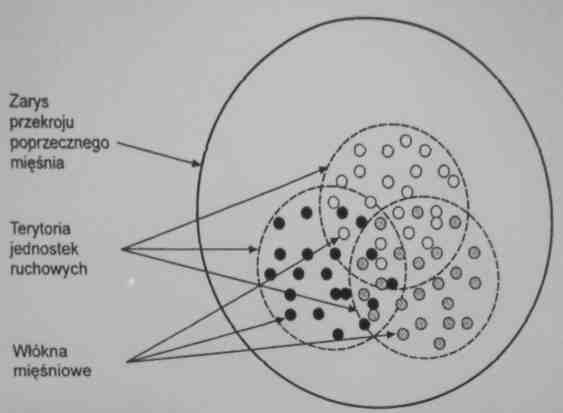
Mięsnie są strukturalnie podzielone na określoną lic/hę jednostek ruchowych (jednostek motorycznych).
Włókna mięśniowe unerwiane przez dany neuron ruchowy są rozproszone w obrębie tego samego mięśnia, tworząc tzw. terytoria jednostek ruchowych.
Podział mięśni ze względu funkcje (ruch)
mięśnie agonistyczne (protagonistyczne)- skurcz powoduje ruch, główny wykonawca danego ruchu np. mięsień dwugłosy ramienia
mięśnie antagonistyczne- przeciwstawiają się mięśniom agonistycznym np. mięsień trójgłowy ramienia
mięsnie synergistyczne- stabilizują staw, asystuje mięśniom agonistycznym np. mięsień ramienno- promieniowy
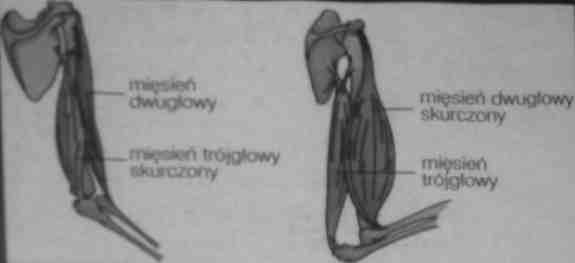
ANTAGONISTYCZNE DZIAŁANIE MIĘŚNI
-mięśnie mają zdolność do aktywnego kurczenia się
- ich rozkurcz jest aktem biernym -wymaga skurczu innego mięśnia
-wyróżniamy dwie grupy czynnościowe mięśni : zginacze (przywodziciele) i prostowniki (odwodziciele)
-mięśnie wykonujące przeciwstawną czynność nazywamy antagonistami
Energetyka skurczu mięśnia i wysiłku
W mięśniach następuje bezpośrednie przekształcenie energii chemicznej w mechaniczną
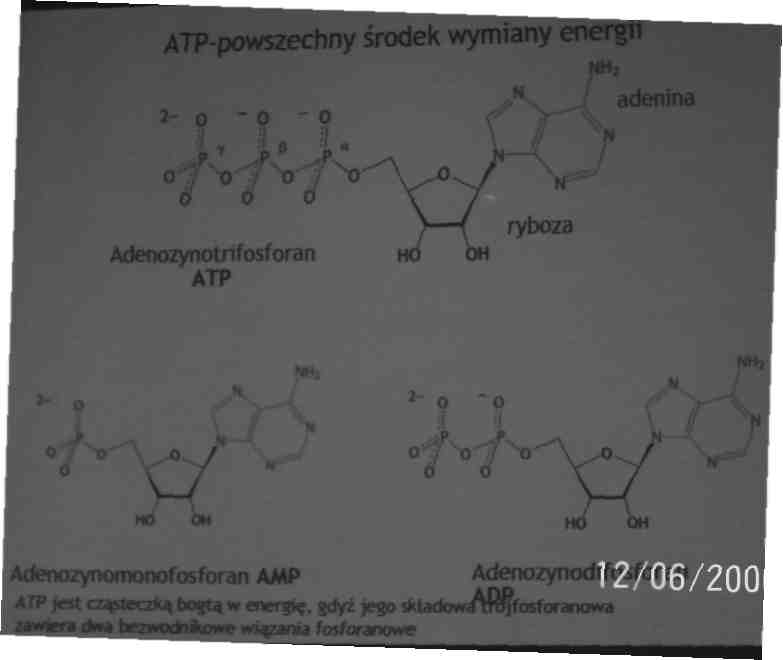
Energię czerpiemy bezpośrednio z ATP adenozynotrifosforanu
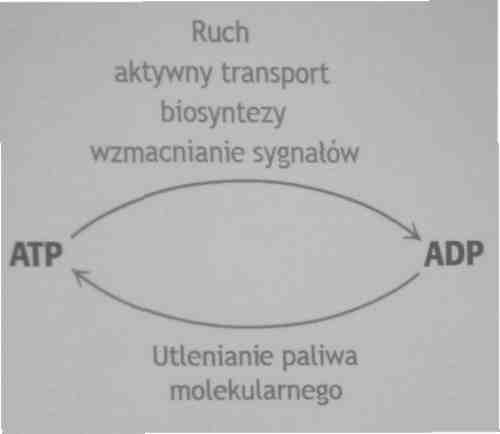
Cykl ATP-ADP jest podstawowym sposobem wymiany energii w układach biologicznych
Duża ilość energii jest wyzwalana gdy ATP ulega hydrolizie do adenozynomonofosforanu i ortofosforanu lub AMP i pirofosforanu. Tworzenie i zużywanie ATP są procesami ciągłymi
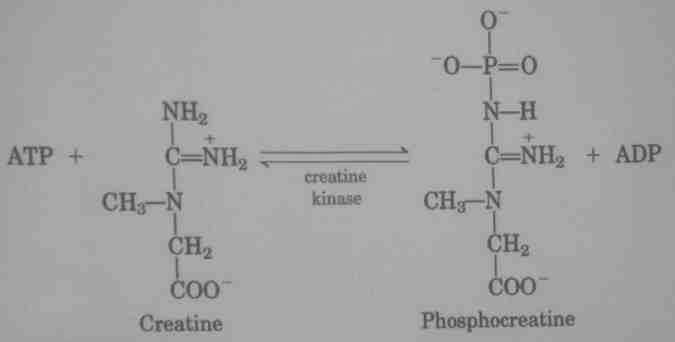
Energetyka skurczu mięśnia i wysiłku
1. Energię czerpiemy bezpośrednio z ATP.
2. ATP musi ulegać ciągłej resyntezie z szybkością dostosowaną do wielkości jego wykorzystywania.
3. Fosfokreatyna (PC) jest magazynem wysokonergetycznych fosforanów.
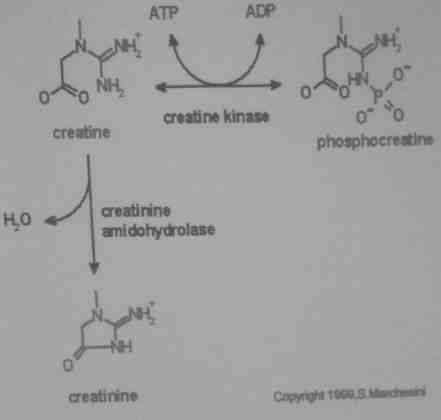
Produktem rozpadu kreatyny jest kreatynina. Kreatynina służy jako marker sprawności nerek.
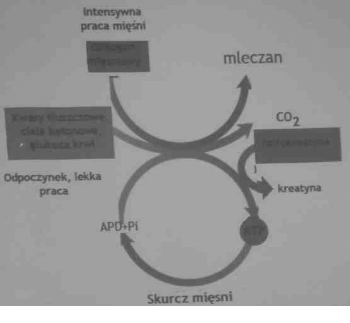
<= Źródłami energii podczas pracy mięśni są: ATP, fosfokreatyna, glukoza i glikogen, a także kwasy tłuszczowe i ciała ketonowe
Mięśnie posiadają trzy systemy otrzymywania ATP potrzebnego do skurczu:
•System natychmiastowy:przekształcanie ATP i fosfokreatyny (PC)( fosfageny)
•system glikolizy beztlenowej: przemiana węglowodanów na drodze przemian beztlenowych do pirogronianu i mleczanu
•system tlenowy-utleniania komórkowego-przemiana węglowodanów lub tłuszczy do H20 i C02
Czerpanie energii z tych trzech systemów zależy od intensywności i czasu trwania wysiłku
System natychmiastowy: wykorzystuje zapasy ATP (b. Małe ~90g) i fosfokreatyny (CP)
Fosfokreatyna (PC) jest magazynem wysokonergetycznych fosforanów i przekazuje je na ATP. Całkowita ilość energii dostępna w systemie natychmiastowym wynosi tylko 10 kalorii, ale jest dostępna natychmiast i umożliwia włóknom FS wytworzenie szybko siły
system ten wyczerpuje się w ciągu kilku sekund
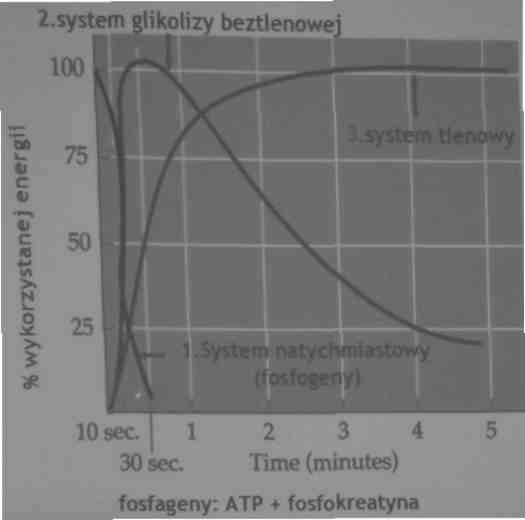
System glikolizy beztlenowej zostaje uruchomiony w ciągu kilku sekund:
•ATP jest wytarzane w procesie glikolizy w cytoplazmie włókien mięśniowych i jest natychmiast dostępne dla włókien miozynowych
•w następstwie glikolizy dochodzi do nagromadzenia kwasu mlekowego, co spowalnia proces
•System natychmiastowy i glikolityczny dostarczają energii na czas mniejszy niż jedna minuta
System tlenowy - może zostać uruchomiony w ciągu minuty i dostarcza on olbrzymia ilość ATP
system tlenowy - utlenianie komórkowe przebiega w wewnętrznej błonie mitochondrium z udziałem wielu
reakcji chemicznych, dzięki którym następuje synteza ATP
system tlenowy -jest wolniejszy niż
dwa omówione poprzednio (natychmiastowy i glikolizy ale dostarcza nieograniczoną ilość ATP)
Czerpanie energii z przemian beztlenowych lub tlenowych w zależności od intensywności i czasu wysiłku =>
Wysiłki beztlenowe trwają krótko i są bardzo intensywne
Kiedy rozpoczynamy ruch, niezależnie od jego prędkości, fosfageny wewnątrzmieśniowe natychmiast dostarczają energii.
Po kilku pierwszych sekundach ruchu coraz większa ilość energii do resyntezy ATP jest czerpana na drodze glikolizy beztlenowej.
Wysiłki trwające do 10 sekund: głównym źródłem energii są fosforany.
Wysiłki trwające do 2 minut: energia głównie z przemian mleczanowych: rozkład glikogenu do glukozy i glukozy do pirogronianu. Z powodu braku tlenu, pirogronian zostaje przekształcony w klas mlekowy.
Poziom kwasu mlekowego świadczy o udziale przemian beztlenowych.
Beztlenowa glikoliza może wyprodukować dużą ilość ATP, ale jest o połowę wolniejsza od układu fosfagenowego.
Podczas intensywnych ćwiczeń, szybkość powstawania pirogronianu w warunkach anaerobowych w szlaku glikolitycznym jest większa, niż szybkość jego utleniania w cyklu kwasu cytrynowego.
W wyniku redukcji pirogronianu powstaje mleczan (kwas mlekowy). Mleczan powoduje ból w mięśniach po dużym wysiłku.
Redukcja pirogronianu do mleczanu jest spowodowana również koniecznością odnowienia zapasów NAD+, który jest niezbędny do utleniania aldehydu 3-fosfoglicerynowego (czyli do podtrzymania glikolizy w pracującej tkance mięśniowej).
Tworząc mleczan mięśnie zyskują na czasie, a także przekazują innym narządom część obciążenia.
Cykl Corich: mięśnie dostarczają mleczanu do wątroby, która zużywa go do syntezy glukozy. Zarazem wątroba odbudowuje stężenie glukozy niezbędne komórkom mięśniowym, które czerpią ATP z jej glikolitycznego przekształcenia w mleczan.
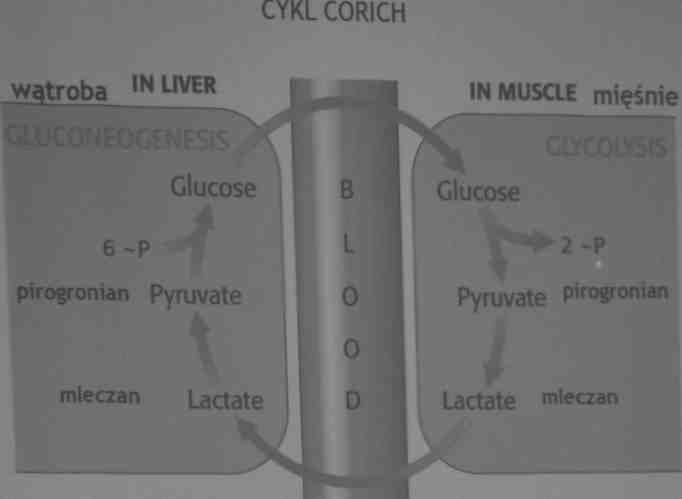
Mleczan powstający w pracującym mięśniu ulega w wątrobie przekształceniu w glukozę.
Opóźniona Bolesność Mięśni Szkieletowych (Delayed Onset Muscle Soreness, DOMS)
Jest skutkiem pracy ekscentrycznej mięśni szkieletowych.
Pojawia się ze znacznym opóźnieniem - zwykle po 24-48 godzinach od zakończenia wysiłku i przez wiele lat niesłusznie wiązano ją z teoria nagromadzenia kwasu mlekowego (tzw. zakwasów).
Objawia się m.in. osłabieniem maksymalnej siły skurczu dowolnego, ograniczeniem zakresu ruchu, obrzękami, bolesnością uciskową i dysfunkcjami nerwowo-mięśniowymi.
We krwi pojawiają się markery uszkodzenia tkankowego, a w biopsjach mięśniowych, wykonywanych bezpośrednio
po ćwiczeniach ekscentrycznych, wyraźnie widoczne jest uszkodzeni różnych struktur komórkowych.
Wysiłki tlenowe są wykonywane przez dłuższy czas z mniejszą intensywnością, i stanowią podstawę do treningu wytrzymałościowego
Procesy tlenowe: utlenianie glukozy, kwasów tłuszczowych i aminokwasów są utleniane z wytworzeniem ATP.
Udział procesów tlenowych zwiększa się wraz z wydłużeniem czasu wysiłku.
czasie długotrwałej pracy mięśniowej zapotrzebowanie energetyczne mięsni może być pokrywane w 95% przez procesy tlenowe.
W wysiłkach o wyjątkowo długim czasie trwania większa część energii pochodzi z rozpadu tłuszczów.
W wysiłkach trwających od 2 do 4 godzin około połowa energii może pochodzić z zasobów glikogenu, zanim ulegnie on całkowitemu wyczerpaniu.
Różnice w pozyskiwaniu energii między przemianami beztlenowymi i tlenowymi
Mechanizm tlenowy charakteryzuje się najmniejszą maksymalną szybkością generowania mocy. Natomiast zasoby energii dla mechanizmu tlenowego (tłuszcze) są praktycznie nieograniczone.
Mechanizm fosfagenowy jest wykorzystywany przez mięśnie dla rozwijania dużych mocy w czasie kilku sekund,
natomiast mechanizm tlenowy dla długo trwającej aktywności
fizycznej.
Między nimi sytuuje się mechanizm beztlenowego rozkładu glikogenu z wytworzeniem kwasu mlekowego (ważny w średnich dystansach).
Intensywność i czas trwania wysiłku warunkują wykorzystanie => substratów energetycznych
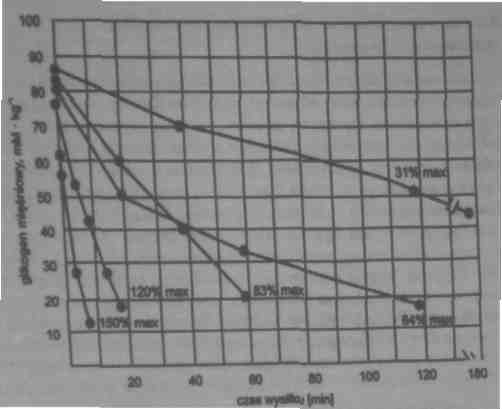
Wyczerpanie się glikogenu mięśniowego w zależności od intensywności (%max -procent mocy maksymalnej) i czasu trwania wysiłku
Wyczerpywanie glikogenu jest różne we włóknach ST (wolnokurczliwych) i FT (szybkokurczliych)
W wysiłkach o niewielkiej intensywności najpierw rekrutowane są włókna ST: dlatego w takich wysiłkach wyczerpanie glikogenu jest największe we włóknach ST.
Wraz ze wzrostem intensywności zwiększa się rekrutacja włókien FT, i dlatego w wysiłkach o dużej intensywności, szybkość wyczerpywania glikogenu jest największa we włóknach FT.
Dzieje się tak dlatego, że włókna FT wykorzystują w większym stopniu glikogen jako źródło energii.
Podczas wysiłku o wysokiej intensywności wszystkie włókna mięśniowe są rekrutowane.
Wysiłki długotrwałe są wykonywane przy znacznym wykorzystywaniu glikogenu i tłuszczów jako źródła energii
Zasoby energetyczne organizmu
Substrat |
Masa [g] |
Energia [kJ] |
Węglowodany: |
375 |
7800 |
- glikogen w wątrobie |
110 |
2300 |
glikogen w mięśniach |
250 |
5200 |
- glukoza w płynach ustrojowych |
15 |
300 |
Tłuszcze: |
10 000 |
420 000 |
- tkanka podskórna |
9800 |
411 600 |
- mięśnie |
200 |
8400 |
Podczas wysiłków wytrzymałościowych zasoby glikogenu zostają uszczuplone, i wtedy mięśnie zużywają głównie tłuszcze. Niemniej, preferowanym źródłem energii jest glikogen i glukoza.
Podział dyscyplin sportowych ze względu na źródła energetyczne
System fosfagenowy (prawie wyłącznie):
- 100 m sprint, skoki, podnoszenie ciężarów, skoki do wody
- biegi sprinterskie, piłka nożna, piłka siatkowa.
System fosfagenowy i glikogen-kwas mlekowy:
- bieg na 200 m, koszykówka, hokej na lodzie.
System glikogen - kwas mlekowy (głównie):
- bieg na 400 m, pływanie na 100 m, tenis.
System glikogen - kwas mlekowy i system tlenowy:
- bieg na 800 m, pływanie na 200 m, jazda na łyżwach na 1500 m, boks, wioślarstwo na 2000 m, bieg na 1500 m, pływanie na 400 m.
System tlenowy: jazda na łyżwach na 10 000 m, narciarstwo biegowe, bieg maratoński.
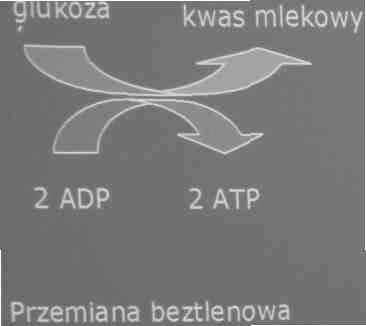
Źródła energii wykorzystywanej do pracy mięśniowej
2. Wysiłki trwające do 60sekund
-glukoza magazynowana jest w tkance mięśniowej w postaci glikogenu
-gromadzenie się kwasu mlekowego powoduje silne zakwaszanie środowiska tkanki mięśniowej (charakterystyczny skurcz lub ból). Działanie szlaku ustaje
-kwas mlekowy przenika do krwi i jest transportowany do wątroby, gdzie ulega przemianie w glukozę (glikoneogeneza).
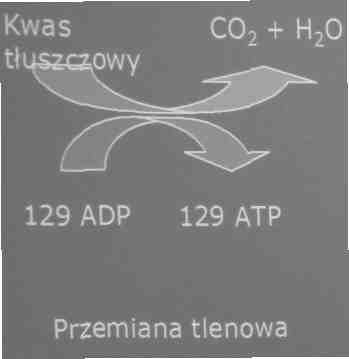
4. Wysiłki trwające ponad 60 minut
-zasoby kwasów tłuszczowych w organizmie są ogromne
-jest to najwolniejszy z przedstawionych szlaków metabolicznych. Czynnikiem ograniczającym tę przemianę jest szybkość transportu kwasów tłuszczowych z krwi do komórek mięśniowych
-czynnikiem ograniczającym długość pracy mięsni w tym trybie są inne układy niezdolne do długotrwałego funkcjonowania (np. układ nerwowy)
Wpływ wysiłku fizycznego na mięśnie szkieletowe
Człowiek jako gatunek powstał co najmniej 300 000 lat temu. Do zaspokojenia potrzeb życiowych potrzebował dużej aktywności ruchowej. Uważa się, że wynikiem tego jest stosunkowo niewielka liczba włókien mięśniowych typu FT (podtyp llx).
Ilość poszczególnych typów włókien w mięśniach dziedziczy się.
Tkanka mięśniowa może dostosowywać się do różnego stopnia obciążenia wysiłkiem fizycznym. Zdolność tę nazywamy adaptacją.
W procesie treningu dochodzi zarówno do zmian cech skurczu, jak i cech biochemicznych tkanki mięśniowej w takim kierunku, aby aktualnie wykonywana praca była realizowana w sposób optymalny. Trening
wytrzymałościowy siły mięśniowej szybkości
Czynniki wpływające na efektywność treningu:
• poziom siły rozwijanych skurczów
•czas trwania wysiłku
•częstotliwość wysiłku
• rodzaje skurczów (izometryczne lub izotoniczne, ekscentryczne lub koncentryczne).
W słabych skurczach udział biorą głównie włókna S, dlatego trening prowadzony na niskim poziomie siły skurczu indukuje zmiany adaptacyjne tylko we włóknach mięśniowych S i FR (I i IIa)
Skurcze o bardzo dużej sile oddziałują na włókna mięśniowe jednostek FF
Czas trwania jest również ważnym parametrem: na ogół czas mobilizacji jest najmniejszy w jednostkach najszybszych (typu llx). Ich niska odporność na zmęczenie jest więc objawem przystosowania do bardzo krótko trwającej aktywności.
Białka mięśniowe podlegają obrotowi metabolicznemu (tempo obrotu białek mięśniowych to ok. 50 g na dobę).
Tempo to wzrasta pod wpływem treningu siłowego.
Synteza nowych białek jest jednym z elementów adaptacji, ponieważ we włóknach mięśniowych pojawiają się miozyny charakterystyczne dla włókien mięśniowych innego typu.
Możliwa jest ekspresja dwóch rodzajów miozyny w jednym włóknie mięśniowym (są to tzw. włókna hybrydowe).
Czy możliwa jest transformacja włókien jednego typu w inny pod wpływem treningu?
Uważa się, że jednostki ruchowe osoby trenującej zmieniają cechy swojego skurczu, ale zmiany te nie są wystarczająco głębokie i transformacja włókien mięśniowych FT do ST (ani ST do FI nie następuje.
W wyniku regularnych ćwiczeń (treningu) nie zmienia się procentowy udział włókien ST w mięśniu.
Natomiast uważa się, że głębokie zmiany cech włókien FT, wraz ze znaczną zmianą typu występującej w nich miozyny, pozwalają na określenie tych zmian jako transformacji włókien mięśniowych szybkich.
Transformacja ta przebiega od włókien llx do Ha i odwrotnie.
Trening wytrzymałościowy powoduje wzrost liczby włókien mięśniowych Ma kosztem llx.
Trening wytrzymałości
Trening wytrzymałości jest realizowany na niższych poziomach siły skurczu, dlatego największe zmiany pojawiają się w włóknach typu I i Ha.
Trening ten zwiększa odporność na zmęczenie, co w odniesieniu do tkanki mięśniowej oznacza wzrost potencjału tlenowego.
Trening wytrzymałości
/ \
interwałowy ciągły
Trening interwałowy jest bardziej skuteczny, ale oba dają podobne zmiany w odniesieniu do progu przemian anaerobowych.
Trening wytrzymałościowy
Trening wytrzymałości prowadzi do podniesienia progu przemian anaerobowych, maksymalnego poboru tlenu (V02max) i progu wentylacyjnego.
Przyczynia się też do obniżenia stężenia mleczanu po obciążeniu submaksymalnym. Dzieje się tak dlatego, że dług tlenowy
zaciągany na początku wysiłku jest mniejszy, a ponadto trening fizyczny pobudza metabolizm (utylizację) powstałego mleczanu.
Trening wytrzymałości zwiększa gęstość kapilar w mięśniach, co polepsza ich ukrwienie i powoduje sprawniejsze dostarczanie tlenu.
Trening wytrzymałości prowadzi do:
• zwiększenia liczby, rozmiarów i gęstości mitochondriów
• wzrostu aktywności enzymów metabolizmu tlenowego co powoduje większą produkcję ATP
• wzrost stężenia mioglobiny
• zwiększenie zasobów glikogenu
• zwiększenia się w mięśniach procentowego udziału włókien typu Ha i zmniejszenia włókien typu lix
Trening wytrzymałości może spowodować niewielki wzrost masy mięśniowej.
U wybitnych biegaczy długodystansowych obserwuje się zmniejszenie masy mięśniowej, związane ze zmniejszeniem się średnicy włókien mięśniowych typu I i Ha (w cieńszych włóknach dyfuzja tlenu jest łatwiejsza).
Bardzo intensywny trening wytrzymałości może doprowadzić do atrofii części włókien mięśniowych
Trening wytrzymałości ma wpływ na szybkość skurczu:
•wzrost szybkości skurczu włókien ST
•obniżenie szybkości skurczu włókien FT
Trening fizyczny może powodować uszkodzenia włókien mięśniowych.
Skutkiem tych uszkodzeń jest pojawienie się zwiększonej aktywności enzymów wewnątrzkomórkowych w osoczu krwi (np. kinazy kreatynowej i dehydrogenazy mleczanowej).
Trening siły mięśniowej
•Trening siły powoduje znaczący wzrost średnicy włókien mięśniowych, a przez to wzrost masy mięśniowej i siły skurczu mięsni
•Wzrost średnicy włókien szybko kurczących się
•zwiększenie potencjału beztlenowego
•wzrost aktywności enzymów metabolizmu beztlenowego
•spadek poziomu mioglobiny
•wzmaga odporność na zmęczenie
• rozbudowuje unaczynienia kapilarne włókien mięśniowych.
Skurcze o charakterze wysiłku izometrycznego oraz ekscentrycznego powodują większy przyrost masy mięśniowej niż skurcze izotoniczne oraz koncentryczne
Kształtowanie siły mięśniowej poprzez wysiłki fizyczne, z przewagą skurczów izometrycznych, powinno rozpocząć się
dopiero po okresie pełnego ukształtowania kośćca, tzn. w wieku 20-30 lat
Fizjologia - mięśnie
1. Rodzaje mięśni, ich budowa.
2. Molekularne silniki: miozyna i aktyna.
3. Mechanizm skurczu mięśni.
4. Motoneurony, małe i duże jednostki motoryczne.
5. Mięsień sercowy i gładki.
6. Rodzaje skurczów mięśni.
7. Jednostki motoryczne szybkokurczące (FT) i wolnokurczące (ST).
8. Optymalna siła i kąt działania mięśnia.
9. Rola ATP, fosfokreatyny i glikogenu w skurczu mięśni.
10. System beztlenowy i tlenowy.
11. Udział poszczególnych systemów energetycznych podczas uprawiania różnych dyscyplin sportu.
12. Wpływ treningu na mięśnie szkieletowe.
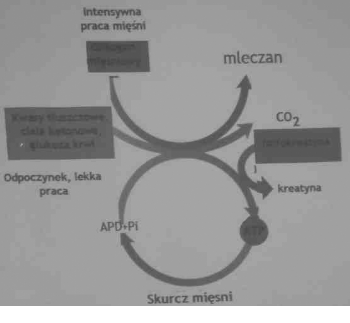
Glikogen mięśniowy
fosfokreatyna
Kwasy tłuszczowe,
ciała ketonowe,
glukoza krwi
glukoneogeneza
glikoliza
Czas wysiłku(min)
Glikogen mięśniowy mM-kg ?
Wyszukiwarka