ZMIANY W BIOCENOZACH:
* Zmiany kierunkowe = sukcesja
* Zmiany niekierunkowe w czasie (maja charakter cykliczny)
ZMIANY KIERUNKOWE = SUKCESJA
KLASYCZNY MODEL SUKCESJI - MODEL UŁATWIANIA
MODEL TOLERANCJI
MODEL KOLONIZACJI LOSOWEJ
KLASYCZNY MODEL SUKCESJI - MODEL UŁATWIANIA
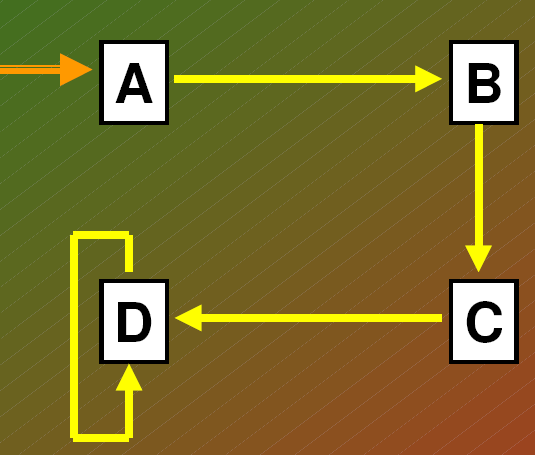
Pojedyncza strzałka - sekwencja gatunków
Podwójna strzałka - alternatywny punkt początkowy sukcesji po zakłóceniu w środowisku
strzałka zakrzywiona - wskazują, że nastąpiła
samowymiana gatunku
Sekwencje wymiany gatunków w procesie sukcesji
-Podstawowym założeniem jest to, że jeden gatunek zastępuje drugi, ponieważ w każdym stadium gatunki
tak przekształcają środowisko, że dla nich samych staje sie ono mniej odpowiednie, dla innych zaś
bardziej odpowiednie.
-Wymiana gatunków jest procesem uporzadkowanym i przewidywalnym oraz sprawia, że sukcesja jest
procesem kierunkowym.
-Sukcesja jest ułatwiana przez gatunki żyjace we wczesniejszych fazach sukcesji.
-Clements (1916, 1936) - hipoteza monoklimaksu
-Egler (1954) - sztafeta florystyczna
-Connel, Slatyer (1977) - model ułatwiania
MODEL HAMOWANIA (Connel, Slatyer 1977)

Pojedyncza strzałka - sekwencja gatunków
Podwójna strzałka - alternatywny punkt początkowy sukcesji po zakłóceniu w środowisku
- Wymiana gatunków jest hamowana przez wczesniejszych kolonizatorów, a szeregi sukcesyjne
zale_a od tego, jaki gatunek osiedlił sie pierwszy.
- Wymiana gatunków niekoniecznie jest uporzadkowana, ponieważ każdy gatunek eliminuje lub
hamuje rozwój nowych kolonistów.
- Sukcesja jest bardzo niejednorodna, ponieważ rozwój fito - zoocenozy w dowolnym miejscu zależy
od tego, jaki gatunek dotrze tam pierwszy.
MODEL TOLERANCJI (Connel, Slatyer 1977)
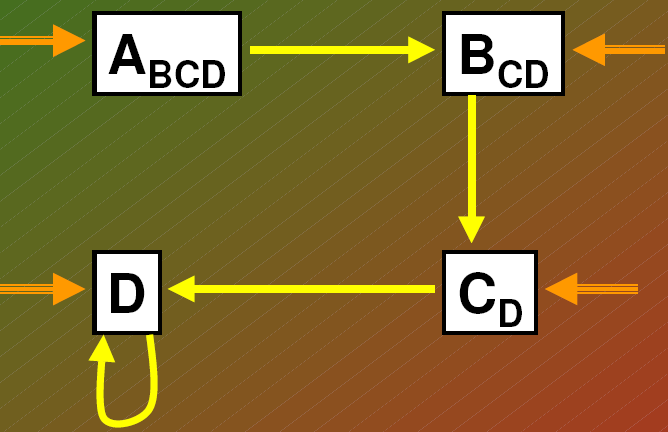
Pojedyncza strzałka - sekwencja gatunków
Podwójna strzałka - alternatywny punkt początkowy sukcesji po zakłóceniu w środowisku
strzałka zakrzywiona: wskazują, że nastąpiła samowymiana gatunku
małe litery (indeks dolny): gatunki, które są mniej
waznym składnikiem biocenozy albo są w stadium
nasienia czy zarodnika
* model pośredni
- Sukcesje może rozpoczac każdy gatunek. Obecnosc gatunków wczesnych stadiów sukcesji nie ma
zasadniczego znaczenia.
- Gatunki sa wymieniane przez inne, które lepiej toleruja działanie czynników ograniczajacych.
- Niektóre gatunki moga uzyskac przewage w konkurencji i własnie one beda dominowac w biocenozie klimaksowej.
Egler (1954) - sekwencja poczatkowego składu florystycznego
MODEL KOLONIZACJI LOSOWEJ (Lawton 1987)
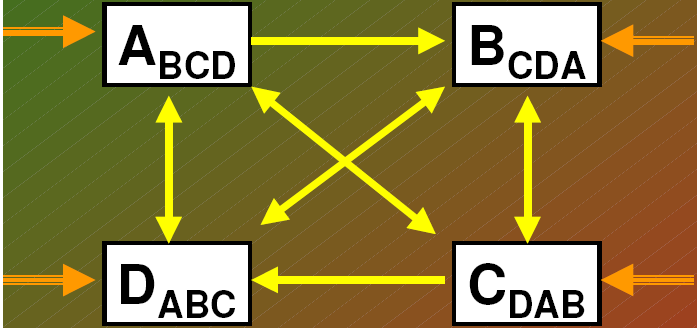
Pojedyncza strzałka - sekwencja gatunków
Podwójna strzałka - alternatywny punkt początkowy sukcesji po zakłóceniu w środowisku
strzałka zakrzywiona: wskazują, że nastąpiła samowymiana gatunku
małe litery (indeks dolny): gatunki, które są mniej
waznym składnikiem biocenozy albo są w stadium
nasienia czy zarodnika
* model zerowy
- Sukcesja polega jedynie na losowym przeżywaniu różnych gatunków i losowej kolonizacji przez nowe
gatunki.
- Nie ma żadnego „ułatwiania” i żadnej konkurencji międzygatunkowej.
Podobieństwa i różnice:
- Pierwsze trzy modele sukcesji przewidują pojawienie się w procesie sukcesji najpierw wielu gatunków
pionierskich
- Mechanizm osiedlania się kolejnych gatunków to najważniejszy element, który odróżnia 4 hipotezy
* Klasyczny model ułatwiania: wymiana gatunków jest ułatwiana przez wcześniejsze stadia
* Model hamowania: wymiana gatunków jest hamowana przez osiedlone osobniki do czasu, kiedy
zostaną uszkodzone lub zgina
* Modele tolerancji i kolonizacji losowej: osiedlone osobniki nie maja wpływu na wymianę gatunków
- Zwykle szeregi sukcesyjne są złożone i nie zmierzają w jednym kierunku do określonego punktu
końcowego
- Mamy przykłady, gdy niektóre fragmenty szeregu sukcesyjnego pasują do modelu klasycznego, inne
modelu hamowania lub tolerancji
Sukcesja na lodowcowej morenie: Klasyczny model ułatwiania
rośliny pionierskie
zmieniają właściwości gleby
nowe gatunki
zmieniają właściwości gleby
nowe gatunki
Sukcesja na piaszczystej wydmie (jezioro Michigan)
rośliny pionierskie
Model ułatwienia
dąb czarny
model hamowania
biocenoza klimaksowa I biocenoza klimaksowa II
biocenoza klimaksowa III biocenoza klimaksowa IV
Sukcesja na terenach porolnych (Karolina Północna)
palusznik
Przymiotno kanadyjskie
Aster Model hamowania
Palczatka Model tolerancji (sosna niezależna od
wcześniejszych stadiów, zasiedla pole,
gdy pojawia się jej nasiona)
Sosna
Model ułatwiania (dęby uzależnione od
zmian w glebie spowodowanych przez sosnę)
Dęby
Sukcesja wokół wulkanu na górze St. Helens
Organizmy pionierskie
Model ułatwienia
Organizmy pośredniego etapu
Model hamowania
Organizmy późnego etapu sukcesji
Obecnie próbujemy analizować sukcesje jako dynamiczny proces wynikający z równowagi miedzy
zdolnością kolonizowania nowych siedlisk przez niektóre gatunki i efektywnością w konkurencji
innych
STAN KLIMAKSU :
KLIMAKS - ostateczna, stabilna biocenoza; układ samopodtrzymujący się i znajdujący w stanie
równowagi ze środowiskiem fizycznym i biotycznym
Teoria monoklimaksu (Clements 1916, 1936)
Teoria poliklimaksu (Tansley (1939), Daubenmire (1966)
Teoria układu klimaksowego (Whittaker (1953)
Teoria monoklimaksu :
- Ka_demu rejonowi geograficznemu odpowiada tylko jedna biocenoza klimaksowa, do której da_a w
swoim rozwoju wszystkie biocenozy.
- Klimaks w danym rejonie jest wyłacznie funkcja klimatu tam panujacego.
* W przyrodzie obserwujemy biocenozy, które według teorii Clementsa nie sa klimaksowe, ale mimo to sa
w stanie równowagi.
Teoria poliklimaksu :
- W danym miejscu może być wyróżnionych wiele różnych biocenoz klimaksowych.
(k. klimatyczny k. edaficzny k. Topograficzny)
- Stan równowagi nie może być osiągnięty, bowiem biocenoza jest kształtowana przez zmienny klimat, a
nie stały.
* Rzeczywista różnica miedzy tymi szkołami jest związana ze skalą czasu, w jakiej mierzymy względna
stabilność.
* Czy powinniśmy rozpatrywać sukcesje w skali geologicznej, czy ekologicznej?
Klimat zmienia się w skali czasu ekologicznego, jak i geologicznego.
Stan równowagi nigdy nie może być osiągnięty, bowiem roślinność jest kształtowana przez klimat
zmienny.
Sukcesja jest procesem ciągłym, ponieważ mamy do czynienia ze zmienna roślinnością współgrającą ze zmiennym klimatem.
Teoria układu klimaksowego (Whittaker (1953)
Nie istnieje tylko jeden klimaks klimatyczny w danym rejonie (teoria monoklimaksu), czy te_ kilka klimaksów(teoria poliklimaksu).
Mamy do czynienia z kontinuum typów klimaksu, zmieniających się wzdłuż gradientów środowiskowych.
* Klimaks jest postrzegany jako biocenoza w stanie równowagi, a tworzace ja populacje znajduja sie w
dynamicznej równowadze z gradientami srodowiskowymi. Teoria układu klimaksowego (Whittaker
(1953)
* Nie mówimy o klimaksie klimatycznym, ale o dominujacych klimaksach, które sa koncowym rezultatem
oddziaływania:
klimatu, czynników biotycznych, topografii, gleby pożary, zasolenie, zdarzenia losowe i inne
Wyżyny w północno-zachodniej Szkocji: mozaika zbiorowisk, zmieniająca się pod wpływem zmian intensywności wypasania owiec i częstości pożarów.
* Mała presja wypasu owiec zadrzewienia z sosna i brzoza
i brak pożarów
* Większa presja wypasu i zespoły trawiaste
częste pożary
* Wypasanie umiarkowane, zbiorowiska z wrzosem i orlica
a pożary rzadkie
ZMIANY CYKLICZNE W BIOCENOZACH
Istnieje cała kategoria zmian w biocenozach, które nie sa sukcesja i maja charakter cykliczny.
Przykład: wrzosowisko z Calluna w Szkocji
1. Faza pionierska (6-10 lat)
- wkroczenie i wczesny etap wzrostu Calluna: otwarte płaty roślinności z wieloma gatunkami roślin
2. Faza budowania (7-15 lat)
- maksymalne pokrycie Calluna, intensywne kwitnienie, niewiele in. gatunków
3. Faza dojrzała (14-25 lat)
- powstawanie przerw w łanie Calluna i wkracza więcej gatunków
4. Faza degeneracyjna (20-30 lat)
- gałązki Calluna obumieraja, zaczynają dominować porosty i mszaki
Biocenozy klimaksowe maja charakter dynamiczny i mogą zmieniać się cyklicznie z powodu cyklu
życiowego gatunków dominujących.
„Stabilna” biocenoza może być mozaika roślinności w czterech fazach cyklicznych zmian lokalnych (faza pionierska, budowania, dojrzałości i degeneracyjna).
Podsumowanie:
BIOCENOZY W PRZYRODZIE NIE SA STABILNE przez długi okres z powodu:
- krótkoterminowych zmian klimatu lub innych czynników środowiskowych
- cyklicznych zmian procesów wzrostu i rozkładu
W przypadku większości biocenoz obserwujemy zmiany w czasie, ale nie znamy wszystkich
czynników, które je powodują. Trudno jest nam proponować, co zrobić, aby zmienić
kierunek niepożądanych zmian.
STRUKTURA I FUNKCJONOWANIE EKOSYSTEMU:
EKOSYSTEM - wszystkie rośliny i zwierzęta zasiedlające określony obszar wraz ze środowiskiem
fizycznym i chemicznym, w którym te organizmy żyją i z którym pozostają we
wzajemnych związkach - Tansley (1935)
EKOSYSTEM = BIOCENOZA + BIOTOP
Wymiana materii i energii miedzy elementami systemu!
EKOSYSTEM - dowolny fragment biosfery, w którym grupa organizmów realizuje procesy produkcji i
dekompozycji, przy chociaż częściowo zamkniętym obiegu materii, z wykorzystaniem
przepływającej przez ten system energii
Terminy używane zamiast słowa „ekosystem”
Mikrokosmos [Forbes 1887]
Holocen [Friedrichs 1930]
Biosystem [Thienemann 1939]
Biocenoza [Naumov 1955]
Pleocen [Trojan 1975]
Fizjocenoza [Wodziczko 1950]
Biogeocenoza [Sukacev i Dylis 1964]
Przepływ energii przez ekosystem
Przepływy energii w bioenergetyce ekologicznej
* C = konsumpcja * FU = odchody
R = rozproszone ciepło (respiracja)
P = energia w wyprodukowanej biomasie (produkcja)
A = tempo pobierania energii po odjęciu strat
w odchodach (asymilacja)
* C = konsumpcja * FU = odchody
R = respiracja P = produkcja
A = asymilacja
C = P + R + FU
A = P + R
Obieg materii w przyrodzie
Przepływ energii przez ekosystem
II prawo termodynamiki: każdej przemianie energetycznej towarzyszy strata wolnej energii w systemie
Zasada 10% [Lindeman] 5-22% Ułamek % do kilkudziesięciu % !
STRUKTURA BIOTYCZNA EKOSYSTEMU:
|
|
Łańcuchy i sieci pokarmowe, poziomy troficzne:
„KTO KOGO ZJADA” (zasada stanowiąca jeden z podstawowych elementów organizacji biocenozy)
Cechy wspólne sieci pokarmowych [Pimm i in. 1991]
1. Istnieje granica złożoności sieci
- Gęstość powiązań pozostaje stała, tzn. na każdy gatunek przypadają 2 interakcje troficzne, bez
względu na liczbę gatunków występujących w biocenozie
2. Łańcuchy troficzne są krótkie
- hipoteza energetyczna
- hipoteza dynamicznej równowagi
3. Istnieje niemal niezmienna proporcja miedzy liczbami gatunków szczytowych drapieżników, gatunków
pośrednich poziomów troficznych oraz gatunkami producentów, niezależnie od wielkości sieci
pokarmowej
4. W sieciach pokarmowych wszystkożerność jest rzadka
5. Zwierzęta kolejnych poziomów troficznych w łańcuchu są coraz większe (poza pasożytami)
6. Sieci troficzne ekosystemów w stałych warunkach środowiskowych maja dłuższe łańcuchy i wyższy
współczynnik konektancji.
Większość sieci pokarmowych jest opracowanych fragmentarycznie. Analiza złożonych sieci
pokarmowych obejmujących więcej ni_ 100 gatunków dopiero się rozpoczyna.
Poznanie struktury sieci pokarmowej pozwoli planować skuteczne strategie ochrony przyrody.
Ekosystem zbudowany z 10 000 elementów - Ile istnieje miedzy nimi relacji dwukierunkowych?
99 990 000
n elementów (n2 - n) zale_nosci
Sieci pokarmowe, łańcuchy troficzne NIEPRZEWIDYWALNOSC EKOSYSTEMU
Osobliwość burząca wszelkie prawa
Miejsce: zachodnie wybrzeże USA, dziuple drzew
Larwy komara Orzeski bakterie i in.
Aedes sierrensis Lambornella clarki mikroorganizmy
Gildie, gatunki kluczowe i gatunki dominujące:
GILDIE:
- grupy gatunków wykorzystujące wspólna pule zasobów w podobny sposób
* np. kolibry i inne ptaki, które żywią się nektarem w rejonach tropikalnych
* np. mrówki, gryzonie, ptaki, które żywią się nasionami
Zalety pojęcia gildii w badaniach organizacji biocenoz
1. Gildie pozwalają na porównanie biocenoz przez skoncentrowanie się na konkretnych grupach
funkcjonalnych.
2. Redukuje liczbę komponentów biocenozy.
3. Zwraca uwagę na fakt, że jednostki ekologiczne nie są jednostkami taksonomicznymi.
GATUNKI KLUCZOWE (ZWORNIKOWE):
Gatunki, których oddziaływanie określa strukturę biocenozy R.T. Paine (1969)
* Rozgwiazda - Pisaster ochraceous
- gatunek kluczowy biocenoz skalistych stref miedzypływowych z zachodniej części Ameryki
Północnej
SKUTKI USUNIECIA ROZGWIAZDY:
- Małż Mytilus californianus zmonopolizował przestrzeń i wyparł inne bezkręgowce oraz glony
- tam, gdzie występuje rozgwiazda, koegzystuje 15 gatunków mięczaków i pąkli
* Wydra morska - Enhydra lutris
- występowały licznie u zachodnich wybrzeży Ameryki Północnej, od Alaski po południowa
Kalifornie
- pod koniec XIX wieku zostały prawie całkowicie wytępione przez europejskich eksploratorów i
osadników
SKUTKI USUNIECIA WYDRY:
* jeżowce (główne ofiary wydr) gwałtownie zwiększyły liczebność i zaczęły konsumować
przybrzeżne wodorosty
* powstały „pustkowia jeżowcowe”
* zniknęły skorupiaki, głowonogi, ryby i in. organizmów
* wieloryby szare przestały się pojawiać u wybrzeży
Dużym wysiłkiem społecznym dokonano restytucji wydry
* wzrosła liczebność wydry
* spadła liczebność jeżowców
* lasy wodorostów wróciły do stanu pierwotnego bogactwa
* pojawiło się mnóstwo pomniejszych glonów, skorupiaków, głowonogów, ryb i in. organizmów
* wieloryby szare znów migrowały bliżej brzegu
* Fomes igniarius - pasożytniczy grzyb atakujący topole
brak grzyba:
- dzięcioł Sphyrapicus nuchalis nie potrafi wykuć dziupli w zdrowej topoli
- Dziuplaki, np. jaskółki drzewne (Tachycineta bicolor, T. thalassina) nie mogą skorzystać z dziupli
po dzięciołach
- Liczne owady trąca dostęp do słodkiego soku wypływającego z topoli
* Słoń afrykański (Loxodonta africana):
- gatunek kluczowy na sawannach
* sposób odwiania słoni powoduje niszczenie krzewów i młodych drzew
* zbiorowiska zaroślowe zamieniają się w otwarte tereny trawiaste
GATUNKI DOMINUJACE:
wyróżnia się na podstawie ich liczebności, biomasy lub wielkości produkcji, mogą wywierać ogromny wpływ na występowanie innych gatunków
Struktura dominacji jest ważnym, aczkolwiek słabo poznanym elementem organizacji
biocenozy
Gatunki dominujące mogą być centralnym punktem interakcji, które określają działanie innych
gatunków w biocenozie
Gatunki dominujące mogą mięć znaczenie dla stabilności samej biocenozy, jej organizacji
Gatunki dominujące wygrywają w konkurencji, a swoja pozycje zawdzięczają wypieraniu
konkurencyjnemu.
1. konkurencyjna hierarchia liniowa
![]()
2. układ zależności konkurencyjnych
Wyszukiwarka