Obieg azotu.
Proteoliza. Pierwszym etapem tych przemian jest proteoliza, czyli hydroliza białek do aminokwasów. Mikroorganizmy heterotroficzne bytujące w środowisku glebowym wytwarzają i wydzielają enzymy biorące udział w tym procesie. Enzymy te powodują hydrolizę białek i polipeptydów do wolnych aminokwasów. Proces ten prowadzony jest przez hydrolazy peptydowe - enzymy działające na wiązanie peptydowe. Dzielą się one na peptydazy oraz na endopeptydazy, które rozpoczynają rozkład białek.
Procesem odwrotnym do proteolizy jest biosynteza białek. Proces ten polega na łączeniu się aminokwasów w łańcuchy polipeptydowe o określonej sekwencji. Biosynteza białek jest procesem de novo, tzn. zachodzi w komórce przy wykorzystaniu dostępnych aminokwasów i jej pełnego potencjału biochemicznego!
Warunkiem katabolitycznego wykorzystania aminokwasów przez komórkę jest usunięcie z nich grupy aminowej w procesie nazywanym deaminacją.
Deaminacja - odłączenie jonu amonowego (NH4+). Z wykorzystaniem dehydrogenaz:
Powyższa reakcja, przebiegająca dwukierunkowo, jest typowa dla bakterii z rodzaju Bacillus.
Proces deaminacji aminokwasów u drobnoustrojów przebiega również z udziałem oksydazy L-aminokwasów (deaminującej, warunki wybitnie tlenowe):
U zdecydowanej większości organizmów żywych podobną rolę odgrywa dehydrogenaza glutaminianowa. Proces można nazwać transdeaminacją (pompa glutaminianowa):
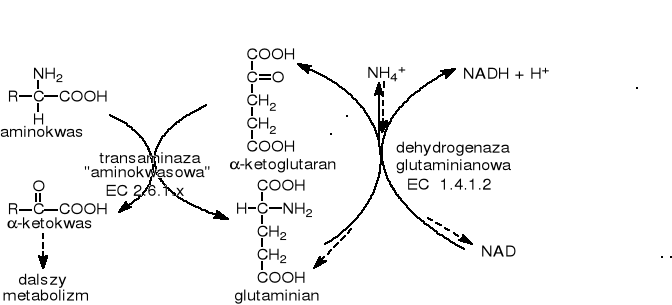
Inną drogą odłączenia od związku amoniaku (np. od aminokwasu) jest działanie amoniako-liaz. Produktem ubocznym jest amoniak, stąd przemiany te nazywane bywają ogólnie amonifikacją.
Procesem odwrotnym do deaminacji jest aminacja, czyli włączenie jonu NH4+ w obieg komórkowy, poprzez powstanie odpowiednich aminokwasów z α-ketokwasów.
Nitryfikacja. Prowadzi do przemiany NH3 w jony azotanowe V (NO3-). Jest to tlenowy proces dwuetapowy:
W etapie I następuje utlenienie jonów amonowych do azotynów:
NH4+ + 3/2 O2 → NO2- + 2H+ + H2O Nitrosomonas
Drugi etap procesu nitryfikacji polega na utlenieniu jonu azotynowego do jonu azotanowego. Reakcja jest odwracalna:
NO2-+ ½ O2 ↔ NO3- Nitrobacter
Nitrosomonas - w wodach słodkich, Nitrosocyjastis oceanus - w słonych.
Azotyny są toksyczne, o ich zawartości w wodzie świadczy obecność bakterii Nitrobacter.
Zbyt duże stężenie NH4+ jest toksyczne dla Nitrobacter (i nie tylko - dla Bacillus i większości drobnoustrojów toksyczne jest stężenie ok. 0,5%, azotanów ok. 1%) - stąd te bakterie pojawiają się jedynie wtedy, gdy Nitrosomonas zużyje większość jonów amonowych i jednocześnie w środowisku występują jony NO2-. Ale w tym samym czasie Nitrosomonas zaczynają znikać, gdyż nie mają już potencjalnego źródła energii, którym są jony NH4+. Kiedy w glebie lub w wodzie brakuje NH4+ i jednocześnie NO2- oba rodzaje bakterii zanikają.
Obecność pestycydów hamuje proces nitryfikacji.
Jest to proces wybitnie tlenowy.
pH: 6-8
temp.: 23-25 oC
Przemiany te mają duże znaczenie dla wzrostu większości roślin, które mają zdolność przyswajania azotanów, a nie amoniaku lub jonów amonowych, które są dla nich szkodliwe.
Większość roślin łatwiej przyswaja jony NO3-, jednakże są one łatwo wypłukiwane z gleby przez padające deszcze i stąd w terenach górzystych rozsądniej jest stosować nawozy zawierające sole amonowe, ponieważ jony NH4+ łatwo tworzą kompleksy ze składnikami gleby i stamtąd też są pobierane przez rośliny.
Proces nitryfikacji jest przykładem sukcesji ekologicznej.
Sukcesja ekologiczna - naturalne następstwo (po sobie) wybranych gatunków roślin i zwierząt, a także drobnoustrojów. Przykładem mogą być niektóre grupy mikroorganizmów występują kolejno po sobie prowadząc sukcesywną przemianę materii. Proces kolejnych dominacji drobnoustrojów z różnych grup powtarza się, aż do całkowitego rozkładu substratu. Sukcesja ekologiczna ma miejsce na przykład przy samooczyszczaniu wód.
Amonifikcja - asymilacja azotanów (NO3-)/?Nitrogeneza?. Dla roślin, wielu drobnoustrojów i niektórych grzybów jony NO3- (a także wyjątkowo NO2-) mogą być źródłem azotu. Jednakże jedyną drogą włączenia tego typu związków azotu w obieg komórkowy jest przekształcenie ich w jony NH4+.
hydroksyloamina
I II NH2OH NH3 amonifikacja
NO3- reduktaza azotanowa NO-2reduktaza azotynowa NO
N2O N2 denitryfikacja Proces amonifikacji NO3- do NH3 przebiega u wszystkich znanych organizmów dwustopniowo.
W I etapie jony NO3- są redukowane do jonów NO2-. Reakcja ta katalizowana jest przez reduktazę azotanową - jest to kluczowy enzym dla asymilacji i denitryfikacji azotanów u bakterii.
W II etapie jony NO2- są redukowane do NH3. Ta reakcja katalizowana może być przez trzy różne enzymy, choć efekt ich działanie jest identyczny: powstaje NH3.
Denitryfikacja. Jest to enzymatyczna redukcja azotanów do azotu cząsteczkowego. Proces ten zachodzi głównie w warunkach beztlenowych (np. Bacillus licheniformis)
Redukcja azotanów prowadzona jest w czterech etapach. W fazie pierwszej azotany redukują się do azotynów. W kolejnych etapach azotyny redukowane są do tlenku azotu, podtenku azotu i ten ostatni do azotu cząsteczkowego.
Nitrogeneza (wiązanie azotu). Kluczowym procesem przyswajania azotu z atmosfery jest przekształcenie gazowej jego formy w NH3. Pewien niewielki procent azotu trafia do organizmów żywych w formie jonów azotanowych NO3- powstających na skutek rozmaitych procesów geologicznych, atmosferyczych i antropogenicznych (głównie: nawożenie). Większość azotu z atmosfery trafia do biosfery poprzez wolno żyjące bakterie azotowe lub sinice. Organizmy te posiadają specjalny enzym - nitrogenazę, który katalizuje reakcję transformacji N2 do NH3.
Mo2+ jest aktywatorem
Pi - fosforan nieorganiczny
Proces głównie tlenowy.
Zachodzi wolno.
Organizmy wiążące azot: Azotobacter, glony (sinice - cjanobakterie), Clostridium (w warunkach beztlenowych - „wyjątek”), niektóre Bacillusy i Pseudomonasy, Rhizobium (każda roślina ma odpowiedni dla siebie szczep)
pH: ok. 7 ± 0,2
Nie może być NH3 i NO3-
Pestycydy hamują.
Bakterie Clostridium żyjące w symbiozie z bakteriami z rodzaju Azotobacter (pierwsze dostarczają kwas masłowy dla Azotobacter, a te z kolei zużywają tlen)
Z punktu widzenia ilości wiązanego azotu największą grupą są bakterie z grupy rizobiów (np. Rhizobium), jednakże zdolność do wiązania azotu u tych organizmów jest ściśle związana z symbiozą z roślinami motylkowymi. I występuje tu ścisła zależność (mutatualizm) gatunku Rhizobium od gatunku rośliny, przykładowo:
Rhizobium lupini - łubin, Rhizobium trifolii - koniczyna, Rhizobium phasoli - fasola,
Rhizobium japonicum - soja.
Aby doszło do symbiozy pomiędzy Rhizobium a rośliną muszą zostać spełnione pewne warunki:
właściwy szczep bakterii dla konkretnej rośliny,
pH~7,0,
obecność molibdenu w glebie,
niska zawartość NO2- i NO3-.
Obieg węgla.
Pierwiastek ten występuje w litosferze w ilości 123 000x1017 g.
W atmosferze pierwiastek ten występuje głównie w postaci CO2 w ilości 23x1017 g.
Schemat obiegu węgla w przyrodzie
1. - osadzanie, 2. - wietrzenie, 3. - fotosynteza, 4. - oddychanie, 5. - z rozkładu, 6. - spalanie
Nadmiar, który wędruje do atmosfery znajduje się w lignino celulozie (przyrost - 24t rocznie/osobę)
Biosacharydy
Błonnik roślinny - Ligninoceluloza
Błonnik roślinny to kompleks substancji ścian komórkowych roślin o budowie lignino-celulozowej nieprzyswajalnych przez nasz organizm. Jest substancją niejednorodną. Obejmuje on zarówno frakcje polisacharydów np. celulozę, hemicelulozę, pektyny, gumy i kleje roślinne, polisacharydy roślin morskich, skrobię oporną, jak i ligniny nie zaliczane do tej grupy związków chemicznych.
Budowa ligninocelulozy
- Elementarne fibryle celulozowe (mikrofibryle celulozy)
- Materiał wypełniający (hemicelulozy, pektyny i skrobia)
- Lignina
Skrobia.
Łatwo ulega biodegradacji.
Wiązania α-1,4-glikozydowe (glukoza) i α-1,6-glikozydowe (amyloza i amylopektyna)
Enzymy: amylolityczne, np. α-amylaza, dekstranaza (rozkładają do glukozy)
Pektyna.
Łatwo ulega biodegradacji.
Głównie kwasy uronowe - ?poligarakturonowe?.
Enzymy pektyno lityczne.
Celuloza
Trudno degradowana
Zbudowana z D-glukozy - wiązania -1,4-glikozydowe (okresowo występuje celobioza)
Enzymy: celulolityczne - celulaza („tnie” na pół), -glukozydaza (odszczepia po 1 glukozie od końca nieredukującego).
Rozkładają ją niektóre bakterie i zwierzęta (przeżuwacze, np. krowy)
W zależności od pochodzenia ilość cząsteczek glukozy waha się od 300 do ?ok.3000?
Hemiceluloza.
Zbudowana z wielu różnych monosacharydów, głównie pentoz (np. mannoza, arabinoza, itp.)
Występuje wiele odmian w zależności od pochodzenia.
Wiązania: -1,4-glikozydowe i 1,6 (jeżeli występuje glukoza)
Enzymy: rózne, w zależności od hemicelulozy - mannanazy, ksylaznazy
Budowa: liniowa lub rozgałęziona, jednorodna lub nie (przeróżne kombinacje), np.:
Hemicelulozy liniowe:
- jednorodne X - X - X - X - X - X - X - X - X - X
- niejednorodne X - Y - Z - X - Y - Z - X - Y - Z - X
Hemicelulozy z łańcuchami bocznymi:
- w bloku X - X - X - X - X - X -X - X - X -X
| | | | | |
Y Y Y Y Y Y
- rozmieszczenie przypadkowe Y Y Y
| | |
X - X - X - X - X - X - X - X - X - X
| | |
Y Y Y
- nieregularne X - X - X - X - X - X - X - X - X - X
| | |
Y Y Y
Hemicelulozy rozgałęzione:
X - X - X - X
/
X - X - X - X - X X - X - X - X - X - X - X - X
\ | |
X - X - X - X Y Y
| |
Y Y
|
Y
W zależności od składu łańcucha głównego hemicelulozy dzielą się na poszczególne grupy:
1. Ksylany:
- zbudowane z ksylozy
- wiązania -1,4
- występowanie: trawa, drzewa liściaste
2. Mannany:
- zbudowane z mannozy
- występują rozgałęzienia
- wiązania -1,4 pomiędzy glukozą i mannozą, co jakiś odstęp dołączona jest galaktoza poprzez wiązanie α-1,4
- występowanie: łupiny orzecha, ściana komórkowa drożdży, drzewa liściaste, pestki
Lignina (nie jest biosacharydem, tylko biopolimerem).
Bardzo trudno degradowana, tylko niektóre drobnoustroje są w stanie ją rozłożyć.
Skład: alkohole (głównie koniferylowy)
Biopolimer ten ma bardzo zwartą strukturę przestrzenną. Z tej zwartej struktury przestrzennej wynika wysoka odporność ligniny na biodegradację.
Włóknik - przetworzone fragmenty drewna, łodyg, itp.
Biodegradacja zdrewniałej ściany komórkowej roślin.
Grzyby rozkładające drewno podzielono ze względu na sposób oddziaływania na poszczególne elementy drewna. Wyróżniamy trzy grupy grzybów:
1. Grzyby tzw. ,,miękkiej zgnilizny”. Powodują one zmiękczenie zewnętrznej warstwy drewna. Tworzą cylindryczne otwory o stożkowatych zakończeniach. Grzyby te należą do klasy Ascomycetes i Deuteromycetes. Najważniejszymi przedstawicielami tej grupy są: Chaetomium i Humicola. Mikroorganizmy te najpierw atakują skrobię i pektyny, następnie bardzo wolno hemicelulozę. Nie posiadają zdolności degradacji ligniny.
2. Grzyby tzw. ,,brunatnej zgnilizny”
Obślizgłe,
charakterystyczny zapach,
brązowe - brunatnienie podłoża spowodowane rozkładem jasnej celulozy, a pozostawieniem brunatnej ligniny.
brak kory
Grzyby: Basidomycetes
Rozkładowi mulega celuloza i hemiceluloza
3. Grzyby ,,białej zgnilizny” - Phanerochaete chrysosporium.
Enzymy:
- laktaza - potrafi demetylować (odszczepiać metanol), narusza strukturę ligniny
- peroksydaza ligninowa - do jej działania niezbędny jest nadtlenek wodoru (bo peroksydaza) ?wytwarzany przez laktazę?
- peroksydaza manganozależna - mangan jest grupa prostetyczną, potrzebny nadtlenek wodoru.
- monoksygenazy - wbudowują tlen w pierścień przez co pęka wiązanie
- dioksygenazy - -||-
Obieg fosforu.
Nadmiar fosforanów w wodach może być dużym problemem - rosną groźne glony (Cjanophyta, itp.). Można je wytrącić chemicznie - opadają na dno, ale nie jest to korzystne. Można też biologicznie - poprzez mikroorganizmy, które kumulują fosforany, po czym są usuwane z uzyciem filtrów, np. Bacillus Cereus, Pseudomonasy, ale ostrożnie bo mogą zaszkodzić.
Najpierw stosuje się warunki tlenowe, a potem po asymilacji beztlenowe dzięki czemu bakterie padają.
Fosfor jest wypłukiwany w dużej części z gleb i spływa do jezior i innych zbiorników wodnych, co w konsekwencji powoduje ich eutrofizację.
Obieg siarki.
Z SO42- powstaje H2S i S.
Siarkowodór powstaje w warunkach beztlenowych, żeby się go pozbyć stosuje się natlenianie lub bakterie np. Thiobacillus
CO2
Wulkany
Skały osadowe
1.
2.
Drobnoustroje -
fotosynteza, chemosynteza
Rośliny
3.
3.
Zwierzęta
4.
4.
Martwa substancja organiczna
Drobnoustroje -
reducenci
5.
Złoża kopalne
Mineralizacja - bez O2
6.
Wyszukiwarka