Epigenetyka
- nauka o dziedziczonych zmianach ekspresji genów, które nie są związane ze zmianami w sekwencji DNA (badanie dziedziczności pozagenowej).
Badanie oddziaływania pomiędzy genami i produktami tych genów, decydujące o powstaniu fenotypu.
Czynniki epigenetyczne
•Metylacja DNA - b.ważna rola w ekspresji genów, przyłączenie grupy metylowej do cytozyny powoduje zamknięcie danego obszaru chromatyny (wyciszanie genu).
•Acetylacja histonów - modyfikacja chromatyny. Euchromatyna posiada silnie acetylowane histony rdzeniowe, to powoduje jej rozluźnienie, DNA staje się dostępne dla czynników transkrypcyjnych i polimeraz. Heterochromatyna posiada niski poziom acetylacji - zamknięta konformacja chromatyny - wyciszona ekspresja genów.
•Konformacja struktury chromatyny
•Kolejność replikacji poszczególnych obszarów chromatyny
Acetylacja odgrywa kluczową rolę
w stymulacji ekspresji genów promujących rozwój stanu zapalnego w reumatoidalnym zapaleniu stawów.
Deacetylacja histonów jako nowy cel terapeutyczny w leczeniu tej choroby ?
Transpozony (skaczące geny) - 44% genomu
Barbara McClintock, nagroda Nobla w dziedzinie fizjologii i medycyny 1983
sekwencja DNA, która może przemieszczać się na inną pozycję
w genomie tej samej komórki w wyniku procesu zwanego transpozycją.
Klasy transpozonów:
•transpozony klasy I, retrotranspozony: zbudowane z RNA, wymagają przepisania na DNA z udziałem odwrotnej transkryptazy
•transpozony klasy II: przemieszczają się w genomie poprzez wycinanie z pierwotnego położenia i wklejanie w nowe miejsce
z udziałem enzymu transpozazy
•
Istnieje hipoteza, że transpozony pochodzą od wirusów (zdolność powielania się i przemieszczania), które utraciły geny odpowiedzialne za zjadliwość
Niekodujące RNA
- miRNA (mikroRNA) - jednoniciowe oligonukleotydy o długości 17 do 24 nukleotydów pełnią w komórce funkcje regulatorowe. Jako jeden ze składników kompleksu rybonukleoproteinowego specyficznie hamują translację mRNA blokując tym samym ekspresję genów
- nsiRNA (small interfering RNA) - dwuniciowe cząsteczki RNA o długości ok. 20-25 par zasad, które powodują wyciszanie ekspresji genów o homologicznej sekwencji.
Powstają przez pocięcie dwuniciowego RNA (np. wirusowego). Wiążą się z białkowym o aktywności rybonukleazy.
Kompleks ten wiąże się z komplementarną do siRNA cząsteczką mRNA i tnie ją na kawałki, uniemożliwiając powstanie kodowanego przez nią białka.
To, co jemy, wpływa na nasze geny
Materiał genetyczny roślin, które zjadamy, nie ulega całkowitej degradacji w naszym układzie pokarmowym a jego fragmenty mogą oddziaływać z naszymi genami.
Niewielkie cząsteczki materiału genetycznego zjedzonych roślin oddziałują z DNA człowieka, wpływając na funkcjonowanie jego genów.
Nauka zna przykłady, w których geny jednego organizmu zostały przechwycone i wykorzystane prze inne:
- ślimak z gatunku Elysia chlorotica, „kradnie” geny związane z procesem fotosyntezy od zielonych glonów, które stanowią jego pożywienie. Ślimak ten wytwarza białka, które nadają powierzchni jego ciała zieloną barwę
i umożliwiają pobieranie energii ze światła.
PRAWO HARDY'ego-WEINBERGA
Zespół osobników zwany populacją wykazuje rozkład właściwości dziedzicznych zależny od rozkładu alleli w populacji
.
Stosunki ilościowe alleli w populacji ulegają zmianom w czasie i przestrzeni w zależności od różnych czynników, takich jak:
wybiórcze kojarzenie się osobników,
mutacje,
selekcja,
migracja,
zmiany liczebności osobników
Częstość występowania różnych genotypów w populacji jest stała i zależna jedynie od częstości występowania alleli w populacji
Prawo jest prawdziwe przy założeniach:
1) populacja jest dostatecznie duża
2) krzyżowanie zachodzi losowo
3) dobór naturalny nie wywiera wpływu
4) populacja jest izolowana
5) brak mutacji
Po spełnieniu tych założeń nie zachodzi zmiana częstości alleli w badanej populacji
Równowaga Hardy'ego - Weinberga
proporcja alleli w kolejnych pokoleniach w stanie równowagi genetycznej jest taka sama (czyli to czego można się spodziewać, gdy populacja nie będzie
p2 + 2pq + q2 = 1
założenie p + q = 1
p2 =częstość homozygot prawidłowych w populacji AA (dominujących)
q2 =częstość homozygot nieprawidłowych w populacji aa (recesywnych)
2pq = częstość heterozygot (Aa)
p = częstość genów dominujących (allel prawidłowy)
q = częstość genów recesywnych (allel nieprawidłowy)
Nosicielem genu nie jest człowiek tylko chromosom
q = q2
ulegała ewolucji)
Ryzyko małżeństwa i ich dzieci związane z chorobą genetyczną na tle dziedziczenia autosomalnego recesywnego, na przykładzie fenyloketonurii
p2 + 2pq + q2 = 1 założenie p + q = 1
q2 = 1 : 10 000 (z obserwacji fenotypowych, chorzy)
q = q2 = 1/10 000 = 1/100 = 0.01
p = 1 - 0.01 = 0.99 ≈ 1
2pq = 2 x 1 x 0.01 = 0.02 ogólnopopulacyjne ryzyko bycia heterozygotą, nosicielem nieprawidłowego allela q = 2%
2/100 = 1/50 1/50 x 1/50 x ¼ = 1/10 000
Mukowiscydoza (Cystic Fibrosis, torbielowate zwłóknienie trzustki ) jest jedną z najczęściej występujących chorób uwarunkowanych genetycznie,
dziedziczoną autosomalnie recesywnie
•Częstość występowania 1:2300
• Do chwili obecnej znanych jest 1400 mutacji i 200 polimorfizmów genu CFTR
•Objawy mukowiscydozy związane są z upośledzeniem funkcjonowania białka CFTR, zwanego czasem transbłonowym regulatorem mukowiscydozy
•Przyczyną choroby jest wrodzony defekt powodujący zagęszczenie śluzu, co prowadzi do zalegania gęstej wydzieliny, głównie w obrębie układu oddechowego (płuca) i w przewodach trzustkowych (niewydolność wydzielnicza trzustki)
Mukowiscydoza w chwili obecnej jest chorobą nieuleczalną i prowadzi do przedwczesnego zgonu (30 lat)
Ryzyko małżeństwa i ich dzieci związane z chorobą genetyczną na tle dziedziczenia autosomalnego recesywnego, na przykładzie mukowiscydozy
p2 + 2pq + q2 = 1 założenie p + q = 1
q2 = 1 : 2 300 (z obserwacji fenotypowych, chorzy)
q = q2 = 1/2300 = 1/48
p = 1 - 1/48 = ≈ 1
2pq = 2 x 1 x 1/48 = 2/48 = 1/24 ogólnopopulacyjne ryzyko bycia heterozygotą, nosicielem nieprawidłowego allela q
1/24 x 1/24 x ¼ = 1/576 x ¼ = 1/2304
1: 2300
1: 144 !!!!
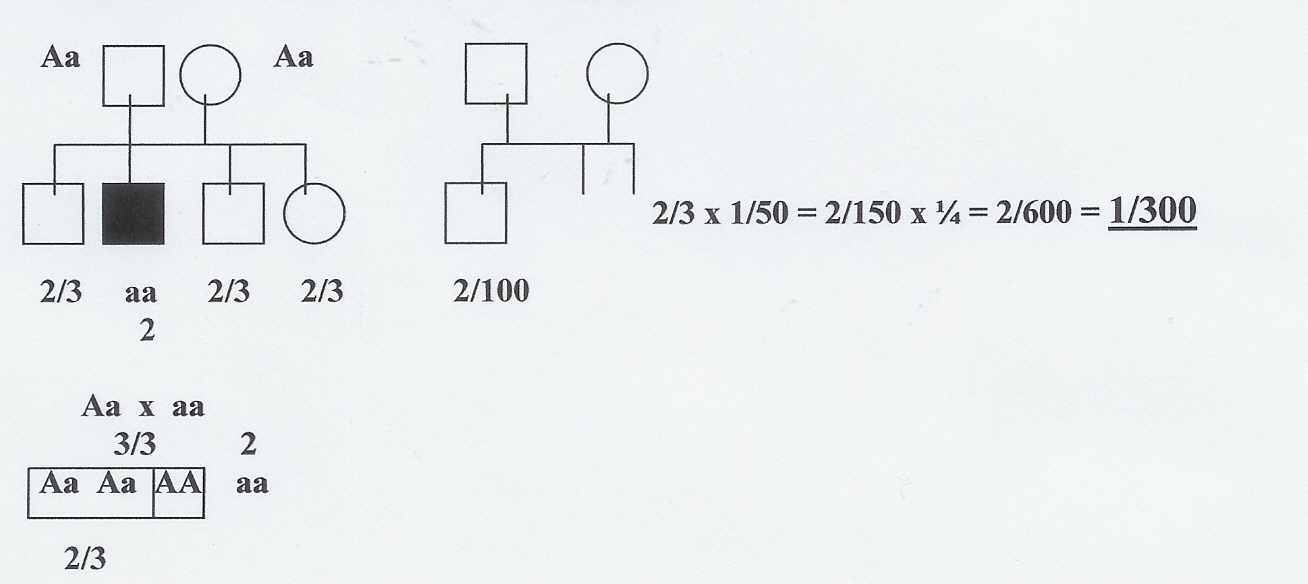
W przypadku obojga rodziców z historią choroby w rodzinie:
2/3 x 2/3 x ¼= 4/9 x ¼ = 4/36 = 1:9 !!!!!!!
Małżeństwa rodzinne, krewniacze
•Kościół do początków XIII wieku zakazywał kojarzenia się rodzinnego aż do siódmego pokolenia włącznie (ale dawał dyspensy). Sobór Laterański w 1215 roku złagodził dotychczasowe zasady kojarzenia się krewnych, udzielając zgody na związki już w piątym pokoleniu.
•Małżeństwa krewniacze pojawiły się już na początku naszej państwowości (związki małżeńskie w obrębie własnej dynstii). Większość książąt piastowskich była w jakiś sposób spokrewniona ze swoimi żonami. Na 160 książąt w związki małżeńskie wstąpiło 96 osób, z czego 47 w obrębie własnej dynastii.
•Konsekwencje - malała liczba dzieci, szczególnie po stronie męskiej, wymieranie całych linii dynastycznych, np. w szóstym pokoleniu wymarła linia wrocławska i raciborska
•Krewniacze związki prowadziły ponadto do rozpowszechniania takich chorób jak:
fenyloketonuria, schizofrenia (przydomki: Dziwaczny, Szalony), cukrzyca, padaczka, alergie, podagra, schorzenia nowotworowe, a także choroby o etiologii środowiskowej, których częstość zależy jednak od predyspozycji genetycznych np. gruźlica, choroba wrzodowa czy kamica.
•Być może na fenyloketonurię cierpiała Jadwiga, córka Henryka Pobożnego, którą ze względu na ograniczenia umysłowe oddano do klasztoru.
•Ponadto cechą charakterystyczną dla fenyloketonurii jest także niedobór melaniny, czego objawem są bardzo jasne włosy i jasna karnacja skóry. Być może od tego pochodzą przydomki Biały, np. Leszek Biały, Władysław Biały, Henryk III Biały, Konrad VII Biały, Konrad X Biały
Choroby autosomalne recesywne występują częściej u dzieci z małżeństw krewniaczych. Szacuje się, że każdy zdrowy człowiek jest nosicielem około dwóch do pięciu, specyficznych dla swojej rodziny mutacji recesywnych, które w układzie homozygotycznym prowadzą do ciężkich chorób. Gdy zaś dobierze sobie spokrewnionego partnera, gwałtownie wzrasta ryzyko, że oboje posiadają tę samą mutację.
W chorobach autosomalnych recesywnych praktycznie nie zdarzają się przypadki niepełnej penetracji i zmiennej ekspresji. Dla niektórych chorób autosomalnych recesywnych opracowano testy kliniczne, biochemiczne lub molekularne, umożliwiające wykrywanie heterozygotycznych nosicieli patologicznego genu.
Najbardziej znane przykłady chorób autosomalnych recesywnych stanowią: mukowiscydoza, fenyloketonuria, galaktozemia, niedokrwistość sierpowatokrwinkowa oraz generalnie choroby spowodowane mutacjami genów kodujących białka funkcjonalne (zwł. enzymy) - większość chorób metabolicznych.
Krewni - z względu na wspólnych przodków posiadają większą ilość wspólnych genów. Im bliższe pokrewieństwo tym więcej wspólnych genów. Zwiększa się ryzyko powstania wielokrotnych homozygot. Tutaj też ma zastosowanie prawo HARDY'ego-WEINBERGA
F = współczynnik wsobności, pokrewieństwa (określa udział wspólnych genów)
Zwiększa się ryzyko powstania wielokrotnych homozygot
R = q2 + Fpq
Związek wuja z bratanicą
F=1/2x1/4=1/8
wysoki współczynnik wsobności
Kuzyni I stopnia F=1/4x1/4=1/16
Kuzyni II stopnia F=1/8x1/8=1/64
brak przeciwskazań
Kuzyni, których rodzice są rodzeństwem, ale przyrodnim F= ¼ x 1/8 = 1/32
1/24
2/3 x 1/24 x ¼ = 2/72 x ¼ = 2/288 = 1/144