Ewa Jankowska-Steifer
TKANKA MIĘŚNIOWA
(textus muscularis)
Możliwość poruszania się, czynność skurczowa serca, wydalanie płodu podczas porodu, przesuwanie treści jelitowej w przewodzie pokarmowym, przepływ krwi w naczyniach krwionośnych to nie jedyne przykłady przejawów ludzkiego życia uwarunkowane obecnością tkanki mięśniowej w organizmie.
Tkanka ta składa się z wydłużonych komórek (miocytów) zawierających białka kurczliwe odpowiadające za zdolność skurczu i rozkurczu. Pomiędzy komórkami mięśniowymi występują elementy tkanki łącznej, w obrębie których przebiegają naczynia krwionośne, naczynia włosowate oraz nerwy.
Tkanka mięśniowa jest pochodzenia mezodermalnego. Na podstawie morfologii oraz charakterystyki funkcjonalnej wyróżniamy trzy jej rodzaje:
tkankę mięśniową poprzecznie prążkowaną szkieletową
tkankę mięśniową poprzecznie prążkowaną serca
tkankę mięśniową gładką
W nazewnictwie elementów budujących komórki mięśniowe wymiennie stosuje się następujące określenia:
Cytoplazma = sarkoplazma, błona komórkowa = sarkolema, gładka siateczka śródplazmatyczna = siateczka sarkoplazmatyczna.
TKANKA MIĘŚNIOWA POPRZECZNIE PRĄŻKOWANA SZKIELETOWA (textus muscularis transversostriatus sceleti)
Tworzy zewnętrzną muskulaturę organizmu, występuje w obrębie jamy ustnej i oczodołów, w ścianie przełyku, odbytu i cewki moczowej. Jej czynność jest zależna od woli a tylko nieliczne mięśnie szkieletowe podlegają kontroli autonomicznego układu nerwowego.
Budowa komórki mięśniowej poprzecznie prążkowanej
Komórki mięśni szkieletowych określane są mianem włókien mięśniowych, wykazujących charakterystyczne poprzeczne prążkowanie. Długość komórek wynosi od kilku milimetrów do kilkudziesięciu centymetrów, co przeważnie odpowiada długości brzuśca mięśnia w którym są one zlokalizowane. Bezpośrednio pod błoną komórkową rozmieszczone są liczne jądra, o wydłużonym kształcie, ułożone równolegle do powierzchni sarkolemy. Głównym składnikiem włókna mięśniowego jest aparat kurczliwy utworzony z równolegle ułożonych włókienek mięśniowych, określanych miofibrylami. Ich długość odpowiada długości samej komórki. W cytoplazmie pomiędzy włókienkami występują mitochondria, specyficznie ukształtowana gładka siateczka śródplazmatyczna, ziarna glikogenu, krople lipidów oraz nieliczne lizosomy.
Miofibryle ułożone są wzdłuż długiej osi włókna mięśniowego. Tworzą je równolegle i gęsto ułożone dwa rodzaje miofilamentów. Miofilamenty cienkie (= aktynowe) zbudowane są z białka aktyny.
Łańcuch fibrylarnej aktyny F
utworzoy jest z globularnej
aktyny G, która posiada miejsce
wiązania miozyny. Dwa skręcone dookoła siebie łańcuchy aktyny F tworzą miofilament cienki.
Miofilamenty grube (= miozynowe) zbudowane są z miozyny II.
Cząsteczka miozyny II składa z
dwóch łańcuchów ciężkich,
które zwijają się dookoła
siebie tworząc helisę oraz z
łańcuchów lekkich, którymi są
globularne główki. Główki po
związaniu z aktyną
wykazują aktywność ATP-azy. Miozyna II układając się w pęczki, z główkami skierowanymi na zewnątrz , wykształca miofilament gruby.
Duże powiększenie mikroskopu optycznego umożliwia dostrzeżenie w komórkach naprzemiennie powtarzających się poprzecznych jasnych (I) i ciemnych (A) prążków, które są odzwierciedleniem regularnego układu miofilamentów wszystkich miofibryli. Podstawową jednostką strukturalną włókienka mięśniowego jest sarkomer. W jego obrębie wyróżnia się charakterystyczne odcinki określane prążkami.
Z
Budowę sarkomeru opisuje wzór: ½ I + A + H + M + H + A + ½ I.
W obrębie prążka I znajdują się tylko miofilamenty aktynowe, w prążku H tylko miofilamenty miozynowe, natomiast w prążku A współwystępują oba typy miofilamentów. W poprzek prążka H przebiega cienki prążek M zawierający dodatkowo białka M i C. Na granicy dwóch sąsiadujących sarkomerów znajduje się prążek Z, który jest miejscem zakotwiczenia miofilamentów aktynowych. Prążek Z zbudowany jest z białek: -aktyniny i desminy. W sąsiadujących miofibrylach sarkomery ułożone są równolegle względem siebie. Elementem spajającym włókienka mięśniowe są filamenty pośrednie desminowe znajdujące się między prążkami Z sąsiednich miofibryli.
Miofilamentom towarzyszą białka strukturalne sarkomeru. Jednym z nich jest nebulina, która owija się dookoła miofilamentów aktynowych utrzymując je we właściwym ułożeniu. Innym białkiem jest tityna, która w postaci długiego łańcucha występuje pomiędzy miofilamentami miozynowymi, w postaci spirali przechodzi przez prążek I i łączy się z prążkiem Z. Białko to stanowi elastyczny szkielet dla ruchu miofilamentów, wraz z -aktyniną uczestniczy w zakotwiczaniu aktyny w prążku Z.
W obrębie sarkomerów występują również białka regulatorowe niezbędne dla prawidłowego mechanizmu skurczu komórki mięśniowej. Są to troponina - troponina C wiąże wapń, - troponina I hamuje wiązanie aktyny do miozyny zaś - troponina T wiąże się z tropomiozyną. Tropomiozyna jest białkiem fibrylarnym złożonym z dwóch łańcuchów tworzących helisę. W czasie rozkurczu białko to zapobiega wiązaniu się główek miozynowych z miofilamentem aktynowym.
Błona komórkowa włókna mięśniowego wytwarza wpuklenia, zwane kanalikami T, biegnące prostopadle w głąb komórki. Kanaliki T otaczają pojedyncze miofibryle na granicy prążków I i A. Tak wymodelowana sarkolema umożliwia szybkie przenoszenie pobudzenia z powierzchni włókna do jego wnętrza. W błonie kanalików T znajdują się receptory dihydropirydynowe.
Poszczególnym miofibrylom towarzyszy charakterystyczny system błon gładkiej siateczki śródplazmatycznej, który otacza miofibrylę na kształt rękawa. W skład pojedynczego segmentu funkcjonalnego siateczki sarkoplazmatycznej wchodzą dwa spłaszczone zbiorniki, zwane cysternami brzeżnymi połączone systemem kanalików. Zbiorniki stanowią rezerwuar jonów Ca2+. W błonie cystern sąsiadującej z kanalikiem T znajdują się receptory rianodynowe.
Układ dwóch zbiorników gładkiej siateczki śródplazmatycznej oraz wciśniętego pomiędzy nimi kanalika T nazywamy triadą. Spełnia ona rolę przełącznika potencjału czynnościowego (depolaryzacja błony) docierającego poprzez kanaliki T w pobliże każdej miofibryli na sygnał do skurczu, którym jest uwalnianie wapnia ze zbiorników gładkiej siateczki sarkoplazmatycznej. Uwolnione jony Ca2+ wchodzą w czasowe połączenia z białkiem regulującym troponiną C i zapoczątkowują reakcję skurczu. W efekcie dochodzi do jednoczesnego skrócenia się sarkomerów wszystkich miofibryli jako wynik ślizgania się miofilamentów aktynowych po miofilamentach miozynowych. W czasie maksymalnego skurczu miofilamenty cienkie są wciągane w głąb prążka A, prążek I jest niewidoczny.
Najważniejsze etapy skurczu mięśnia szkieletowego:
Pobudzenie - rozchodzenie się potencjału czynnościowego poprzez kanaliki T, przekazanie sygnału do siateczki śródplazmatycznej poprzez oddziaływanie receptora dihydropirydynowego z rianodynowym z jednoczesnym uwolnieniem jonów wapnia z cystern brzeżnych.
Sprzężenie - czynnikiem sprzęgającym pobudzenie do skurczu są jony Ca2+. Wiążą się one z troponiną C indukując zmianę jej kształtu, w efekcie dochodzi do zmiany pozycji tropomiozyny umożliwiającej głowom miozyny wiązanie się z miofilamentem aktynowym.
Skurcz - przejściowa, powtarzająca się interakcja główek miozyny z miofilamentami cienkimi. Zmiany konformacyjne w cząsteczkach miozyny wynikające z wiązania i hydrolizowania cząsteczki ATP umożliwiają wędrówkę jej główek wzdłuż miofilamentu aktynowego. Niemal równocześnie pobudzenie wszystkich sarkomerów włókna mięśniowego powoduje szybki skurcz komórki.
Spoczynek - aktywne pobieranie jonów wapnia z sarkoplazmy do cystern siateczki sarkoplazmatycznej, powrót troponiny i tropomiozyny do pozycji spoczynkowej blokującej wiązanie aktyny z miozyną.
Powstawanie tkanki mięśniowej
W rozwoju zarodkowym włókna szkieletowe powstają jako wynik fuzji mezenchymatycznych komórek prekursorowych określanych mianem mioblastów. W wyniku tego procesu dochodzi do wykształcenia miotub, czyli wielojądrzastych tworów komórkowych, w obrębie których przebiega intensywna synteza białek kurczliwych. Część mioblastów nie podlega zjawisku fuzji i przylega do miotubul stając się komórkami satelitarnymi mięśni poprzecznie prążkowanych. Oba rodzaje komórek otacza wspólna błona podstawna. Komórki satelitarne są komórkami macierzystymi mięśni poprzecznie prążkowanych i uczestniczą one w naprawie uszkodzonych mięśni. Jednym z mechanizmów regeneracji jest wykształcanie nowych włókien mięśniowych. Innym jest przerost włókien już istniejących polegający na syntetyzowaniu białek i wykształcaniu dodatkowych miofibryli.
Budowa mięśnia szkieletowego
Mięśnie szkieletowe utworzone są z długich (do kilkudziesięciu cm) pęczków równolegle ułożonych komórek mięśniowych. W budowie mięśnia występują trzy zgrupowania tkanki łącznej właściwej. Przestrzenie pomiędzy pojedynczymi włóknami mięśniowymi wypełnia tkanka łączna luźna, zwana śródmięsną (endomysium), w niej przebiegają liczne naczynia włosowate. Włókna mięśniowe zorganizowane w pęczki otoczone są przez omięśną (perimysium), utworzoną z tkanki łącznej właściwej zbitej. Cały brzusiec mięśnia otacza namięsna (epimysium), czyli torebka zbudowana z tkanki łącznej właściwej o utkaniu zbitym. W obrębie tkanki łącznej przebiegają naczynia krwionośne, limfatyczne oraz nerwy.
Makroskopowa ocena tkanki mięśniowej uwidacznia różnice w naturalnym zabarwieniu mięśni. W zależności od stężenia mioglobiny w komórkach mięśniowych - czerwonego białka wiążącego tlen, włókna mogą być blade lub żywoczerwone. Różnice morfologiczne oraz biochemiczne stały się podstawą wyróżnienia różnych typów włókien mięśniowych:
włókna czerwone (= wolne) - bogate w mioglobinę, zawierają dużo mitochondriów a energia niezbędna do skurczu pochodzi z fosforylacji tlenowej, są to komórki kurczące się powoli o dużej wytrzymałości na zmęczenie.
włókna białe (= szybkie) - zawierają niewiele mioglobiny oraz mitochondriów, energia pozyskiwana jest ze spalania glukozy w procesie beztlenowej glikolizy, skurcz komórki jest szybki a wytrzymałość na zmęczenie niewielka.
włókna mieszane - posiadają cechy włókien białych i czerwonych
Większość mięśni zawiera mieszaną populację różnych włókien mięśniowych. Przy przewadze jednego z typów włókien mięśnie określa się jako czerwone lub białe.
Mięśnie szkieletowe charakteryzują się pewną plastycznością, przejawiającą się zdolnością włókien mięśniowych do zmiany rozmiaru i typu. Brak aktywności ruchowej mięśni spowodowany uszkodzeniem ich unerwienia doprowadza do ubytków masy mięśniowej oraz zaniku włókien wolnych. Częściowe odtworzenie włókien czerwonych można uzyskać stymulując mięśnie bodźcami elektrycznymi.
Przekształcanie się włókien mięśniowych, czyli ich konwersja dotyczy zarówno mięśni osób chorych jak i zdrowych. Odpowiedni, intensywny trening jest w stanie zmienić wzajemne proporcje włókien wolnych i szybkich (dla zainteresowanych: „Geny zamiast koksu”, J. L. Andersen i wsp., Świat nauki, sierpień 2001).
Synapsa nerwowo-mięśniowa
Potencjał czynnościowy dociera do komórki mięśniowej z neuronów ruchowych poprzez synapsę nerwowo-mięśniową. Końcowy odcinek aksonu ruchowego nerwu obwodowego w miejscu przylegania do komórki mięśniowej wykształca część presynaptyczną. W jej obrębie występują pęcherzyki z acetylocholiną. Element postsynaptyczny stanowi wyspecjalizowany, mocno pofałdowany fragment sarkolemmy, w którym zlokalizowane są receptory zdolne do odbioru sygnału przenoszonego przez acetylocholinę włókna nerwowego. Ten fragment włókna określany jest także płytką motoryczną. Zaburzenia transmisji nerwowo-mięśniowej klinicznie charakteryzują się nadmierną nużliwością mięśni. Najczęściej spotykanym schorzeniem jest miastenia rzekomoporaźna, będąca chorobą autoimmunologiczną. Organizm ludzki omyłkowo wytwarza przeciwciała przeciw swoim własnym receptorom acetylocholinowym. Przeciwciała wiążąc się z receptorami znajdującymi się na błonie postsynaptycznej inaktywują je, tym samym uniemożliwiając przekazanie pobudzenia z komórki nerwowej na mięśniową za pośrednictwem acetylocholiny uwalnianej z części presynaptycznej.
Zaburzenia w skomplikowanej budowie komórki mięśnia poprzecznie prążkowanego mogą doprowadzać do chorób zwyrodnieniowych mięśni. Z wewnętrzną powierzchnią sarkolemy łączą się kompleksy białkowe, w obrębie których występują m.in.: aktyna F, dystroglikan, sarkoglikan, laminina oraz dystrofina. Nieprawidłowa budowa białek tego kompleksu jest przyczyną różnych postaci dystrofii mięśniowych. Najczęstszą z nich jest dystrofia mięśniowa Duchenne'a, choroba recesywna sprzężona z chromosomem X - charakteryzująca się brakiem w obrębie komórki mięśniowej dystrofiny, co w efekcie doprowadza do martwicy i ubytku włókien mięśniowych. Jest to choroba postępująca, powodująca znaczne uszkodzenie mięśni i prowadząca do śmierci około 20 roku życia. Rzadziej występującą i o łagodniejszym przebiegu jest dystrofia Becket'a. W mięśniach osób chorych produkowana jest dystofina w ilości mniejszej niż u osób zdrowych, bądź też dystrofina o zmienionej budowie. Czas przeżycia pacjenta może być normalny lub nieznacznie skrócony.
TKANKA MIĘŚNIOWA POPRZECZNIE PRĄŻKOWANA SERCA
(textus muscularis transversostriatus cardiatus)
Występuje w ścianie przedsionków i komór serca tworząc najgrubszą warstwę ściany serca, czyli śródsierdzie (= mięsień sercowy, myocardium). Przylegające do siebie komórki mięśniowe są powiązane połączeniami przewodzącymi bodźce elektryczne. Wytwarzanie i rozprowadzanie impulsów sterujących rytmiczną pracą serca jest możliwe dzięki komórkom układu bodźcowo - przewodzącego.
Budowa komórki mięśniowej poprzecznie prążkowanej serca
Komórki mięśnia sercowego określa się mianem włókien mięśniowych sercowych lub kardiomiocytów. Kardiomiocyty posiadają jedno lub dwa jądra położone centralnie. Komórki są wydłużone z tępo zakończonymi końcami, którymi kontaktują się z sąsiadującymi włóknami wytwarzając w ten sposób szeregi komórek. Miejsce styku dwóch komórek nazywamy wstawką. W jej obrębie występują wyspecjalizowane połączenia międzykomórkowe:
obwódki przylegania, desmosomy oraz połączenia komunikujące neksus (= synapsa elektryczna - umożliwiająca szybki przepływ impulsów elektrycznych między komórkami).
Pomiędzy sąsiednimi szeregami występują połączenia utworzone przez boczne odgałęzienia końcowych fragmentów komórek. Przestrzeń między szeregami wypełnia tkanka łączna luźna. Komórki otacza błona podstawna.
Aparat kurczliwy włókna mięśniowego sercowego zbudowany jest z miofibryli utworzonych z typowych sarkomerów. W przeciwieństwie do mięśnia poprzecznie prążkowanego szkieletowego miofibryle mogą rozgałęziać się i łączyć z sąsiadującymi miofibrylami. Pomiędzy nimi w cytoplazmie znajdują się mitochondria, aparat Golgiego, gładka siateczka śródplazmatyczna, ziarna glikogenu, krople tłuszczu, lizosomy. Sarkoplazma komórki mięśnia sercowego wytwarza kanaliki T, które wpuklają się do wnętrza włókna na wysokości prążków Z. Siateczka sarkoplazmatyczna otacza miofibryle wytwarzając system kanalików i zbiorników. Kanalik T przylega do jednego zbiornika gładkiej siateczki śródplazmatycznej wytwarzając diadę.
Szczególnym typem komórki mięśnia sercowego łączącej w sobie cechy kardiomiocytu oraz komórki wydzielniczej są komórki mioendokrynowe. Ich lokalizacja ograniczona jest do ściany prawego przedsionka serca. W ziarnach wydzielniczych tych komórek występuje przedsionkowy czynnik natiuretyczny.
Czynność skurczowa kardiomiocytów jest niezależna od woli. Pobudzenie potrzebne do rytmicznych skurczów powstaje w układzie przewodzącym przedsionkowo-komorowym serca, którego końcowymi elementami są zmodyfikowane komórki mięśniowe, zwane włóknami Purkinjego.
W tkance poprzecznie prążkowanej serca brak jest komórek satelitarnych, uniemożliwia to regenerację mięśnia sercowego.
Mutacje w genach dla białek skurczowych lub im towarzyszących (aktyna, miozyna, troponina, tropomiozyna) są przyczyną miopatii mięśnia sercowego osłabiających jego funkcję.
TKANKA MIĘŚNIOWA GŁADKA
(textus muscularis nonstriatus s. glaber)
Tworzenie mocnych i elastycznych ścian w narządach wewnętrznych oraz czynność skurczowa są dwiema funkcjami tkanki mięśniowej gładkiej. W przypadku tej tkanki skurcz jest niezależny od woli. Czynnikami wywołującymi go mogą być: - pobudzenie ze strony autonomicznego układu nerwowego - hormony (np.: prostaglandyna, oksytocyna, angiotensyna), - spontanicznie wytwarzany potencjał czynnościowy sarkolemmy niektórych miocytów, - reakcja na rozciąganie.
Budowa komórki mięśniowej gładkiej
Komórki mięśniowe gładkie są wrzecionowatego kształtu z centralnie umieszczonym jądrem. Długość komórek jest różna, najkrótsze (ok.20 m) spotyka się w ścianach naczyń krwionośnych, najdłuższe zaś w ścianie ciężarnej macicy (ok. 200 m). Komórki otoczone są błona podstawną. Pomiędzy sąsiadującymi komórkami występują synapsy elektryczne (= nexus).
Na preparatach histologicznych tej tkanki nie obserwuje się poprzecznego prążkowania. Aparat kurczliwy wykazuje uporządkowaną budowę jednak odmienną od sarkomerowej. Miofilamenty biegną skośnie do długiej osi komórki w pęczkach po kilka lub kilkuset. Miejscem zakotwiczenia miofilamentów cienkich są: taśmy gęste znajdujące się po wewnętrznej stronie błony komórkowej oraz ciałka gęste zlokalizowane w cytoplazmie wewnątrz komórki. Zarówno taśmy oraz ciałka gęste zbudowane są z -aktyniny , filaminy i winkuliny. Ciałka gęste łączą się między sobą za pośrednictwem filamentów pośrednich desminowych, które stabilizują ich położenie wewnątrz komórki. Sarkolema komórki mięśniowej gładkiej wytwarza niewielkie wpuklenia w głąb cytoplazmy określane jamkami komórkowymi (caveolae). Ułatwiają one przeniesienie sygnału z powierzchni komórki w głąb cytoplazmy (brak kanalików T). W cytoplazmie komórek występują typowe organelle : mitochondria, szorstka siateczka śródplazmatyczna, aparat Golgiego, lizosomy. Rozbudowana siateczka sarkoplazmatyczna stanowi magazyn jonów wapniowych niezbędnych dla zainicjowania procesu skurczu w komórce.
Najważniejszymi zdarzeniami doprowadzającymi do skurczu komórki mięśniowej gładkiej jest: - depolaryzacja sarkolemmy powodująca otwarcie kanałów dla wapnia w błonie gładkiej siateczki śródplazmatycznej, - związanie wapnia przez białko kalmodulinę (brak troponiny), - zmiana konformacji kalmoduliny po połączeniu z wapniem umożliwiająca utworzenie kompleksu z kinazą łańcuchów lekkich miozyny II, która w formie aktywnej fosforyluje łańcuch lekki miozyny co umożliwia związanie głowy miozyny z aktyną. Energia potrzebna do zmiany położenia głowy miozyny względem miofilamentu aktynowego i tym samym do skurczu pochodzi z hydrolizy ATP pod wpływem ATP-azowej aktywności głowy miozyny.
W niektórych mięśniach gładkich znajdują się owalne lub gwiaździste miocyty zdolne do automatycznego wytwarzania impulsów do skurczu co jest wynikiem oscylacyjnego przepływu jonów Ca2+ z gładkiej siateczki śródplazmatycznej do cytoplazmy. Są to komórki śródmiąższowe, zwane komórkami Cajala.
Komórki mięśniowe mogą występować jako pojedyncze (np. w kosmkach jelitowych), mogą być skupione w pęczki (jak w gruczole krokowym), tworzyć błony mięśniowe (np. w naczyniach krwionośnych) lub też duże mięśnie (np. w macicy). Elementem spajającym poszczególne komórki jest tkanka łączna właściwa luźna oraz błony podstawne komórek. Komórki mięśni gładkich występują w ścianach wewnętrznych przewodów organizmu (przewód pokarmowy, drogi oddechowe, moczowe, przewody układu rozrodczego, naczynia krwionośne).
W narządach człowieka spotyka się komórki o cechach pośrednich: miocytów gładkich oraz komórek nabłonkowych lub fibroblastów. Są to odpowiednio komórki mioepitelialne i miofibroblasty.
jądro
triada
siateczka sarkoplazmatyczna
kanalik T
troponiny
helisa
główka
sarkolema
M
M
M
H
A
I
Z
Z
S A R K O M E R
tropomiozyna
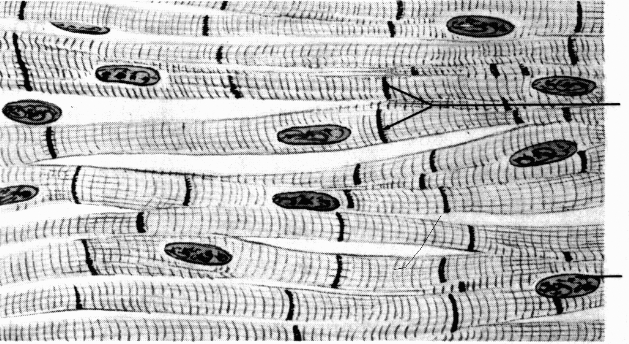
wstawka
jądro
sarkolema
miocyty
prążki A
prążki I
prążek Z
miofibryla
jądro
miofilament gruby
naczynia włosowate
G aktyna
F aktyna
Wyszukiwarka