MIKROBIOLOGIA-ZAGADNIENIA cz. I
Wielkość i kształt komórek bakteryjnych
Są jednymi z najmniejszych mikroorganizmów.
Najmniejsze z nich (tzw. ciałka elementarne form L bakterii mają rozmiar ok. 0,15-1,2 μm;
Największe ( bakterie purpurowe i siarkowe) osiągają rozmiar kilkunastu mikrometrów.
Drobne wymiary stwarzają konieczność uproszczenia w budowie wewnętrznej i mniej doskonałego rozdziału funkcji pomiędzy organelle komórkowe.
Kształt komórek bakteryjnych:
bakterie właściwe (Eubacteriae)
kuliste:
ziarniaki (Coccus)
dwoinki (Diplococcus)
paciorkowce (Streptococcus)
gronkowce (Staphylococcus)
pakietowce (Sarcina)
czworaczki (Tetracoccus)
cylindryczne:
pałeczki
laseczki (Bacillus) (wydłużone i ucięte na końcach)
maczugowce (Corynebacterium)
przecinkowce (Vibrio)
śrubowce (Spirillum)
krętki
skręcone: (ten typ morfologiczny był uwzględniony w instrukcji, prof. Kołwzan podawała tylko te 2 w/w.)
przecinkowce (Vibrio)
śrubowce (Spirillum)
krętki
Kształt i wielkość komórek bakteryjnych są w znacznym stopniu zależne od:
wieku i warunku hodowli;
temperatury inkubacji;
czasu trwania hodowli;
rodzaju podłoża
składu chemicznego podłoża odżywczego;
zmiany w obrębie materiału genetycznego.
Na podłożu standardowym i w stałych warunkach hodowli komórki danego gatunku mają określony kształt będący ich istotna cecha diagnostyczną. W okresie starzenia się bakterii mogą powstawać formy o zmiennych kształtach ( powstaje na skutek zaburzenia w mechanizmie wytworzenia błon podziałowych, nagromadzeniu się w środowisku metabolitów, niekorzystnych warunków rozwoju dla komórek np. natlenienie, temperatura pH środowiska itp.
Omówić budowę i funkcję form przetrwanych.
Budowa przetrwalnika:
rdzeń - cytoplazma otoczona błoną komórkową, czyli protoplast przetrwalnika zawiera wszystkie struktury potrzebne do syntezy białek oraz wytwarzania energii na drodze glikolizy;
ściana przetrwalnika - warstwa znajdująca się najbliżej na zewnątrz błony cytoplazmatycznej. Jest zbudowana z mureiny, po wykiełkowaniu endospory w komórkę wegetatywną staje się ścianą komórkową;
korteks - najgrubsza warstwa ochrony przetrwalnika, zbudowana z mureiny, o mniejszej liczbie mostków poprzecznych niż ściana komórkowa. Zawiera kwas dipikolinowy, wrażliwa na działanie lizozymu;
płaszcz - wewnętrzny i zewnętrzny zbudowany jest z białka kreatyno podobnego z wieloma mostkami dwusiarczkowymi. Jest nieprzepuszczalny zawiera dużą odporność na antybiotyki i środki dezynfekujące. Osłony te mogą stanowić do 50% objętości i 40-60% suchej masy;
egzosporgium - występuje tylko u niektórych gatunków Bacillus błona lipoproteinowa, zawierająca niektóre węglowodany;
Przetrwalniki zawierają o około 40% więcej białka i prawie 4-krotnie mniej węglowodanów niż komórki wegetatywne. Charakterystycznym dla endospor związkiem (występującym tylko w nich) jest kwas dipikolinowy Kwas ten wiąże jony wapnia w stosunku 1:1 tworząc dipikolinianwapnia, ( mogący stanowić do 15% suchej masy spory). Związek ten odgrywa znaczną rolę w ciepłooporności endospor. Przetrwalniki nie zawierają β-hydroksymaślanu, który w komórkach wegetatywnych stanowi prawie 30% suchej masy.
Funkcja:
Są to formy rozwoju umożliwiające przetrwanie warunków, które mogłyby być zabójcze dla normalnych postaci wegetatywnych. Zdolność do wytwarzania endospor (przetrwalników) ma tylko nieliczna grupa bakterii, są to względne i bezwzględne tlenowce z rodzaju Bacillus, Sporosarcina oraz bezwzględne beztlenowce z rodzaju Clostridium;
wysoka temperatura - bakterie giną po 10-minutowym ogrzewaniu w 80°C, natomiast endospory wytrzymują nawet wielogodzinne gotowanie i mogą kiełkować kiedy trafia na korzystne warunki środowiska (ciepłooporność jest proporcjonalna do zawartości kwasu dipikolinowego);
niska temperatura - odporna nawet na zamrażanie, głównie dzięki grubym ścianom i minimalnej zawartości wody;
promieniowanie - dzięki dużej liczbie mostków siarczkowych w zewnętrznych osłonkach białkowych;
wysuszenie - mogą przetrwać latami, a nawet wiekami bez wody;
czynniki chemiczne (wysokie i niskie pH, duże stężenie NaCl).
nie służą rozmnażaniu, a jedynie przetrwaniu, gdyż każda komórka może wytworzyć w swym wnętrzu jedynie jeden przetrwalnik.
Rodzaje przetrwalników:
Endospory - (powstające wewnątrz komórki macierzystej) charakteryzują się największą odpornością na ciepło, wysychanie, promieniowanie i czynniki chemiczne spośród wszystkich form przetrwanych;
Cysty - ( Azotobacter, Methylocystis) okrągłe, grubościenne komórki, są odporne na wysychanie, mechaniczne naprężenie, promieniowanie. Różnica między nimi, a endosporami polega na tym, że;
są to przekształcone całe komórki wegetatywne, a nie ich części;
nie są odporne na ciepło.
Mikrocysty - występują u bakterii śluzowych, twory kuliste lub w kształcie krótkich pałeczek, są oporne na wysuszenie względnie oporne na podwyższona temp. i odznaczające się zdolnością przetrwania niekorzystnych warunków.
Konidia (promieniowce) - często wytworzone na specjalnych nitkach sporonośnych, maja tendencje do wyrastania ku górze ponad podłoże, nie są tak wytrzymałe na temperaturę jak endospory, ale dość dobrze znoszą suszę.
Narządy ruchu mikroorganizmów i ich budowa:
U większości bakterii aktywny ruch wywołany jest rotacja rzęsek. Możliwy jest także ruch ślizgowy (bakterie śluzowe)
Rzęski - są to wyrostki o średnicy 12-18 nm i długości do 20 μm. Składają się z 3 części: włókna, ciałka podstawowego (ciałka bazalnego) i haka łączącego obie te części. Są zaczepione jednym końcem w wewnętrznej osłonie komórki - błonie cytoplazmatycznej i łączą się z jej treścią protoplazmatyczną. Mają kształt lekko skręconej spirali o równej grubości na całej długości i składają się z heliakalnie zwiniętych łańcuchów kurczliwego białka zwanego flagelliną. Umiejscowienie i liczba rzęsek jest dla bakterii cechą charakterystyczną i ma znaczenie taksonomiczne. Rzęska działa jak śruba napędowa a jej ruchy mogą odbywać się zgodnie lub niezgodnie ze wskazówkami zegara.
Organellum ruchu niektórych krętków jest włókno osiowe, składa się ono z pęczka włókienek. Swą budową przypomina typowe rzęski prokariotyczne, wyrastają one z bieguna komórki i spiralnie ja oplatają, dzięki takiej lokalizacji możliwy jest wężykowaty ruch bakterii w środowisku o dużej lepkości.
Fimbrie i pili (koniugacja)
Charakterystyczne struktury występujące wyłącznie u bakterii to fimbrie - zakotwiczone w cytoplazmie nitkowate wyrostki. Są to struktury podobne do rzęsek bakteryjnych, przy czym są one znacznie mniejsze, sztywniejsze i delikatniejsze. Osiągają średnicę 3-25 nm, długość do 12 μm. Zbudowane są z białka zwanego piliną, syntezowanego w cytoplazmie w postaci prebiałka. Cienkie fimbrie ułatwiają przyczepianie się komórek do podłoża oraz między sobą(wytwarzanie nalotu).
Grubsze wyrostki, zwane też pilami płciowymi (pilami typu F),służą podczas koniugacji do przenoszenia DNA (pilusy - organ za pomocą którego, komórki męskie rozpoznają komórki żeńskie i łączą się w procesie koniugacji). Są to puste rurki białkowe o długości do 10 μm.
Koniugacja - przenoszenie dziedzicznych cech dawcy na szczep biorcy przez bezpośredni kontakt w parach. Połączenie się 2 bakterii zachodzi poprzez mostek cytoplazmatyczny utworzony przez pili płciowe. Koniugują tylko bakterie F+ (dawca, zawiera czynnik płciowy F i może wytwarzać pili płciowe) z bakterią F- (biorca, nie ma czynnika F). Podczas koniugacji biorca może uzyskać czynnik F i zacznie wytwarzać pili płciowe, stanie się dawcą.
Otoczki i śluzy międzykomórkowe
Są to mniej lub bardziej grube warstwy substancji zawierającej dużą ilość wody.
Otoczki - mogą być polisacharydowe (głównie z glukozy lub dekstranu), polipeptydowe(gł. z kwasu poliglutaminowego) oraz mieszane cukrowo-lipidowe lub białkowo-cukrowe. Otoczki można uwidocznić pod mikroskopem świetlnym stosując barwniki nie przechodzące przez materiał otoczkowy, np. nigrozynę, czerwień Kongo, tusz chiński. W technice barwienia negatywnego na ciemnym tle preparatu pojawiają się jasne otoczki. Cieńsze otoczki barwi się z wykorzystaniem swoistych przeciwciał.
Otoczki chronią bakterie przed wyschnięciem;
ułatwiają adhezję;
chronią przed reakcją obronną zakażanego organizmu.
Wykazano też, że niektóre bakterie syntetyzujące otoczki są bardziej oporne na działanie antybiotyków, gdyż ich przenikanie do komórki jest utrudnione. U bakterii chorobotwórczych otoczki mogą być czynnikiem zjadliwości, tzn. bakterie otoczkowe są bardziej zjadliwe.
Śluzy otoczkowe - są to liczne substancji otoczkowe wydzielane do środowiska, można je oddzielić od powierzchni komórek przez wstrząsanie. Jest wynikiem gromadzenia się na powierzchni komórek polimerów, polisacharydów i polipeptydów.
ułatwiają bakteriom przetrwanie okresu suszy;
chronią przed szkodliwymi czynnikami zewnętrznymi;
chronią bakterie przed fagocytozą (pochłonięcie i strawienie przez komórki żerne krwi i tkanek).
Ściana komórkowa bakterii gram + i gram -, różnice
Ściana komórkowa jest elastyczną strukturą nadającą komórce określony kształt;
stanowi barierę ochronną przed czynnikami zewnętrznymi fizycznymi i chemicznymi, a także przed innymi mikroorganizmami;
jest przepuszczalna dla licznych substancji i soli mineralnych;
szkielet ściany składa się z polimeru -peptydoglikanu, zwanego mureiną.
Ściana komórkowa bakterii gram dodatnich
Mureina stanowi 30-70% suchej masy ściany komórkowej, składa się z około 40 warstw.
Do murein przyłączone są też różne kwasy tejchojowe (łańcuchy złożone z 8-50 cząsteczek glicerolu lub rybitolu połączonych mostkami fosfoestrowymi).
Ściana komórkowa bakterii gram ujemnych
Sieć mureiny jest jednowarstwowa i stanowi mniej niż 10% suchej masy ściany komórkowej.
W ścianie bakterii gramujemnych nie wykryto kwasów tejchojowych.
Część dominującą ściany stanowi warstwa plastyczna, którą tworzy błona zewnętrzna zbudowana z fosfolipidów, białek, lipoproteidu Brauna, lipopolisacharydu/LPS/.
Białka o funkcji transportowej (poryny) tworzą w błonie kanały wypełnione wodą, które pozwalają na wniknięcie do komórki niskocząsteczkowych substancji hydrofilowych.
Między błoną zewnętrzną a mureiną znajduje się tzw. przestrzeń peryplazmatyczna. Zawiera ona znaczną ilość enzymów biorących udział m.in. w rozkładzie substratów(metanol, glukoza),wykorzystywaniu związków nieorganicznych(siarczany, azotany),rozkładzie białek, polisacharydów, kwasów nukleinowych.
Lipopolisacharydy (LPS) mają duże znaczenie w diagnostyce bakteriologicznej i epidemiologii. Różne szczepy Salmonella typ himurium oraz Shigella desynteriae odpowiedzialne za infekcje jelitowe różnicuje się dzięki O-swoistem ubocznemu łańcuchowi lipopolisacharydów występujących w ich błonie zewnętrznej. Różnice wykrywa się metodami immunologicznymi i serologicznymi, które pozwalają na identyfikacje szczepu i zlokalizowanie źródła infekcji.
RÓŻNICE
GRAM + |
GRAM - |
gruby peptydoglikan |
cienki peptydoglikan |
kwasy tejchojowe: przepływ jonów, ochrona, specyficzność antygenowa |
brak kwasów tejchojowych |
wrażliwe na lizozym |
odporne na działanie lizozymu |
są wrażliwe na penycilynę |
mało wrażliwe na penycilinę |
bardzo wrażliwe na detergenty |
mało wrażliwe na detergenty |
duża wrażliwość na barwniki anilinowe |
mała wrażliwość na barwniki anilinowe |
wrażliwe na eozynę i błękit metylowy |
wrażliwe na safraninę |
mała zawartość poliamin |
duża zawartość poliamin |
duża zapas wolnych aminokwasów |
mały zapas wolnych aminokwasów |
Barwienie metoda Grama - jest to barwienie złożone, pozytywne, w którym stosuje się kolejne dwa kontrastowe barwniki zasadowe: fiolet krystaliczny i fuksynę. Właściwy proce poprzedzony jest termicznym utrwaleniem preparatu. Bakterie gramdodatnie barwią się na fioletowo, gramujemne na czerwono
Budowa i funkcja rybosomów
Rybosomy są drobnymi tworami zbudowanymi z dwóch podjednostek
małej 30S, zawiera 16S rRNA oraz od 1 do 21 specyficznych białek
dużej 50S, zawiera 23S rRNA oraz od i do 32 specyficznych białek
U Prokaryota są mniejsze niż u Eukaryota, mają niższą masę cząsteczkową i stałą sedymentacji Svedberga (która określa sedymentacje w ultrawirówce), wynoszącą70S, w porównaniu do 80S u Eukaryota. Różnice między rybosomami mają ogromne znaczenie przy leczeniu infekcji, gdyż niektóre antybiotyki wybiórczo hamują syntezę białek na rybosomach 70S, nie wpływając na działanie rybosomów 80S.
Podjednostki rybosomów występują w cytoplazmie oddzielnie, łączą się ze sobą tylko po połączeniu z mRNA w czasie syntezy białek. Tworzą wtedy polirybosomy (polisomy) -skupienia wielu rybosomów połączonych nicią mRNA. Są centrami syntezy białek i jako takie odgrywają zasadnicza role w przemianie materii.
Biosynteza białka, zachodzący w żywych komórkach organizmu proces powstawania białka uwarunkowany przez zapisaną w DNA (kwasy nukleinowe) informację genetyczną (gen).
Pierwszym jego etapem jest transkrypcja odpowiedniego odcinka DNA, która polega na syntezie RNA na matrycy określonego odcinka DNA przy udziale polimerazy RNA. RNA powstały w wyniku transkrypcji, zawierający informacje dla syntezy białek, zwany jest mRNA. Przenosi on transkrybowaną informację genetyczną z jądra do cytoplazmy. Tutaj dochodzi do modyfikacji mRNA, tzn. do wycinania, z udziałem odpowiednich enzymów, sekwencji niekodujących - intronów i pozostawiania sekwencji kodujących - egzonów. Tak zmodyfikowane i skrócone cząsteczki mRNA wnikają pomiędzy dwie podjednostki rybosomów, gdzie odbywa się właściwe odczytywanie kodu genetycznego i przepisywanie go na sekwencję aminokwasową białka w procesie zwanym translacją.
Znajdujące się w cytoplazmie aminokwasy są przenoszone na rybosomy za pomocą tRNA. Cząsteczki tRNA z doczepionymi aminokwasami przedostają się do rybosomów i kolejno dopasowują się, na zasadzie komplementarności, swoimi antykodonami do odpowiednich kodonów mRNA. Translacja zaczyna się od kodonu startowego, zapewniającego dalsze odczytywanie mRNA we właściwej kolejności - najczęściej jest to kodon AUG lub GUG, a kończy się kodonem symbolizującym ostatni aminokwas (u prokariontów są to kodony nonsensowne - nie oznaczające żadnego aminokwasu). Po zakończeniu syntezy cząsteczki białka wędrują przez przestrzenie pomiędzy błonami reticulum endoplazmatycznego do aparatu Golgiego albo wydzielane są na zewnątrz komórki, lub pozostają przez jakiś czas związane z błonami ziarnistego (szorstkiego) reticulum endoplazmatycznego i wykorzystywane jako białka wewnątrzkomórkowe. Energia potrzebna do syntezowania wiązań peptydowych pochodzi z wysokoenergetycznych wiązań ATP.
Biosynteza białka może zachodzić również w mitochondriach i plastydach roślinnych, w których występuje DNA.
Budowa i funkcja błony cytoplazmatycznej
Wykazuje typową dla wszystkich błon elementarnych strukturę trójwarstwową i zbudowana jest z lipidów (20-35%) i białek (50-75%).
Składa się z podwójnej warstwy lipidowej. Hydrofobowe końce fosfolipidów i trój glicerydów skierowane są do środka, a hydrofilowe „główki” na zewnątrz.
W tę podwójną warstwę włączone są białka, niektóre przenikają przez całą grubość błony, inne tylko przez część(białka integralne) lub są przyłączone do zewnętrznej i wewnętrznej warstwy hydrofilowej(białka powierzchniowe).
Funkcje błony komórkowej:
•jako półprzepuszczalna stanowi barierę osmotyczną komórki i kontroluje wnikanie i usuwanie różnych substancji,
•jest miejscem zakotwiczenia enzymów biorących udział w przenoszeniu elektronów i w fosforylacji oksydatywnej, a więc w tworzeniu i magazynowaniu energii,
•uczestniczy w procesach syntezy ściany komórkowej, składników otoczki śluzowej, pili, fimbrii, a także wydziela enzymy zewnątrzkomórkowe,
•stanowi centrum replikacji DNA.
Rodzaje transportu:
DYFUZJA BIERNA - najprostszy sposób przenikania, ponieważ substancje przenikają wg. gradientu stężeń tzn. z obszaru o wyższym stężeniu do obszaru o niższym stężeniu.
TRANSPORT AKTYWNY - cząsteczki przechodzą do komórki, ale wymaga to zużycia energii. Takim związkiem energetyvznym jest ATP kwas adezynotrójfosforanowy.
DYFUZJA UŁATWIONA - substancja, która chce wejść do komórki łączy się w obrębi błony z inna substancją zwaną nośnikiem. Tworzy się kompleks nośnik-cząsteczka, który, po przejąciu przez błony rozpada się. Cząsteczka zostaje w komórce. Nośnik wraca na swoje miejsce w błonie i czeka na ponowne tworzenie kompleksu.
Białka transportowe - Dużą grupę białek stanowią białka transportujące, które przenoszą małe cząsteczki lub jony. Takimi białkami są, m.in.: przenosząca lipidy albumina, gromadząca i przenosząca żelazo transferyna oraz podstawowy nośnik tlenu molekularnego — hemoglobina. Hemoglobina jest białkiem występującym obficie w czerwonych krwinkach (erytrocytach) i to właśnie dzięki jej obecności możliwe jest zwiększenie stężenia tlenu molekularnego w płynach ustrojowych o prawie dwa rzędy wielkości w stosunku do ilości tlenu zawartego pod ciśnieniem atmosferycznym w wodzie.
Nukleotyd
Nukleotydy są cząsteczkami składającymi się z trzech grup: zasady azotowej, cukru i grup fosforanowych. W nukleotydach występują dwa różne cukry - ryboza i deoksyryboza, oba są pochodnymi pentozy, pięciowęglowej cząsteczki cyklicznej. Zasady azotowe spotykane w nukleotydach są pochodnymi puryny lub pirymidyny. Do zasad purynowych należą: adenina (A) i guanina (G), do pirymidynowych: uracyl (U), cytozyna (C) i tymina (T). Funkcje:
Nukleotydy są prekursorami DNA i RNA
Nukleotydy mogą być przenośnikami energii chemicznej
Nukleotydy posiadające kilka grup fosforanowych mogą pełnić rolę uniwersalnych przenośników energii. Takim związkiem jest ATP (adeninotrifosforan) posiadający trzy grupy fosforanowe. Przyłączenie ostatniej z nich wymaga dużego na kładu energetycznego. Ta energia uwalniana jest następnie przy przerywaniu tego wiązania. Tak więc cząsteczki ATP są w stanie magazynować energię lub przenosić ją w punkt odległy od miejsca jej syntezy. Nukleotydy adeninowe są składnikami trzech podstawowych koenzymów: NAD+, FAD i CoA Nukleotydy mogą pełnić w komórce rolę regulatorową. Replikacja DNA Replikacja DNA jest procesem zapewniającym przekazywanie informacji genetycznej z komórek rodzicielskich do komórek potomnych w sposób prawie doskonały za sprawą mechanizmu replikacji, a także dzięki systemom ochronnych, zdolnych do wykrywania i naprawy błędów. Podstawowa cecha replikacji jest jej semikonserwatywność. Znaczy to, że synteza nowych nici może zachodzić tylko z udziałem nici rodzicielskich, służących jako matryce. Mówiąc o replikacji należy wprowadzić pojecie replikonu. Replikon jest to jednostka replikacji, za która przyjęto uważać odcinek DNA, zawierający miejsce startu oraz przylegające sekwencje uczestniczące w kontroli tego procesu. W procesie replikacji można wyróżnić trzy zasadnicze etapy:
inicjacje,
elongacje,
terminacje.
Nukleoid - obszar komórki prokariotycznej będący odpowiednikiem jądra komórkowego u Eukaryota. W przeciwieństwie do jądra komórek eukariotycznych nukleoid nie jest oddzielony od cytoplazmy otoczką jądrową. Zawiera genofor (chromosom bakteryjny), czyli pojedynczą, kolistą cząsteczkę dwuniciowego DNA o długości do 200 nm (0,6 -13 mln par zasad).
Plazmidy- samo replikujące się zamknięte, koliste cząsteczki dwuniciowego DNA, które replikują się niezależnie od głównego genoforu. Ich funkcją jest przekazywanie innym bakteriom genów kodujących oporność na różnego rodzaju antybiotyki.
Nukleoid wraz z plazmidami zawiera pełną informację genetyczną komórki
Mezosom:
zbudowany z form błoniastych;
uwypuklenia błony komórkowej, będące efektem działań środków chemicznych używanych do mikroskopowania;
zakotwiczają nukleoid;
zawiera wiele enzymów (synteza ściany, oddychanie - cytochromy, procesy oksydoredukcyjne);
stanowi odpowiednik mitochondriów czyli jest centrum energetycznym(na ich powierzchni znajdują się przenośniki elektronowe), w którym wytwarzane jest ATP i zachodzi proces oddychania wewnątrzkomórkowego (tylko u bakterii tlenowych);
bierze udział w transporcie elektronów;
jest miejscem syntezy kwasów tłuszczowych.
Budowa i funkcja ciał chromatoforowych (fotosyntez bakteryjna)
błoniaste pęcherzyki lub rurki;
często zawierają warstwo ułożony system błon wewnętrznych (budowa lameralna -warstwowa;
zawierają chlorofil, barwniki karotenoidowe białka i lipidy
występują u bakterii fotosyntezujących;
są centrami procesów fotosyntezy.
Istnieją też bakterie fotoautotroficzne, które substancje do swojego odżywiania „produkują” w sobie dzięki procesom fotosyntezy. Przekształcają one zredukowane związki mineralne przy użyciu energii świetlnej (chemoautotrofy posykiwały ją z reakcji chemicznych utleniania różnych związków) w związki organiczne i nie wydzielany jest tlen w tej reakcji. (tym między innymi różni się fotosynteza bakteryjna od fotosyntezy roślinnej). Wydzielane zaś są utlenione związki organiczne oraz utlenione związki mineralne.
U bakterii fotoautotroficznych występuje bakteriochlorofil będący zmagazynowany w tworach nazywanych tylakoidami lub chromatoforami. Fotosynteza bakteryjna przebiega zaś w widmie światła czerwonego, przy braku tlenu. Bakterie jednak zyskują w takim środowisku, potrzebne im do reakcji fotosyntezy, elektrony redukujące z zredukowanych związków wodoru, siarki a także niektórych kwasów organicznych.
Fotosyntezę przeprowadzać mogą tez sinice, one pobierają dwutlenek węgla z powietrza a wodór z wody, i ni podobnie do roślin wydzielają cząsteczkę tlenu
Bakterie fotosyntezujące zawierają chlorofil (sinice) lub bakterio-chlorofil (pozostałe) oraz karotenoidy. Duża liczba tych ostatnich, o zabarwieniu czerwonym i brązowym, może prowadzić do zamaskowania zielonego bakteriochlorofilu, jak dzieje się to u bakterii purpurowych. Sinice przeprowadzają fotosyntezę w sposób podobny do roślin, tzn. z udziałem wody jako źródła atomów wodoru, a co za tym idzie - z wydzielaniem tlenu. U bakterii zielonych i bakterii purpurowych źródłem atomów wodoru mogą być rożne związki nieorganiczne lub organiczne. Często związkiem takim jest siarkowodoru (nieorganiczny, H2S), a produktem powstającym po odebraniu wodoru - czysta siarka.
Różnice pomiędzy fotosynteza bakteryjną i zachodzącą u Eucaryota :
U bakterii występuje inny barwnik zielony tzw. Bakteriochlorofil
U bakterii nie zachodzi proces fotofosforylacji niecyklicznej.
Występuje u nich tylko fotofosforylacja cykliczna.
Fotosynteza bakteryjna zachodzi w warunkach beztlenowych(anaerobowych)
W fotosyntezie bakteryjnej nie zachodzi fotoliza wody, źródłem(donorem) elektronów redukujących bakteriochlorofil są inne związki takie jak siarkowodór, siarka, tiosiarczany czy wodór.
6CO2 + 12H2S - C6H12O6 + 12S + 6H2O
6CO2 + 6H2O C6H12 O6 + 6O2
Materiał zapasowy w komórkach
Substancje, które można uważać za materiały zapasowe to:
polisacharydy,
tłuszcze,
polifosforany,
siarka.
Znajdują się one w komórce w postaci osmotycznie nieczynnej i są nierozpuszczalne w wodzie.
Do odkładania substancji zapasowych konieczne jest, aby w podłożu były obecne składniki potrzebne do ich syntezy. Po przywróceniu warunków sprzyjających wzrostowi substancje zapasowe mogą być ponownie włączone w metabolizm.
Występowanie polisacharydów;
ziarna glikogenu występują w komórkach E.coli, Salmonella, Bacillus, Micrococcus luteus.
Skrobię zawierają Acetobacter pasteurianus i wiele gatunków z rodzaju Neisseria.
granulozę (substancję podobną do skrobi) gromadzą bakterie z rodzaju Clostridium.
Substancje tłuszczowe występują u mikroorganizmów w postaci kropelek. U wielu bakterii te kropelki zawierają kwas poli-β-hydroksymasłowy ( PHB), poliester rozpuszczalny w chloroformie, nierozpuszczalny w eterze. Jest zbudowany z około 60 reszt kwasuβ-hydroksymasłowego. Może on stanowić aż 90% suchej masy komórki. Gromadzą go liczne bakterie tlenowe, sinice i beztlenowe bakterie fototroficzne. Produkowany jest w warunkach ograniczonego dostępu do tlenu, jako produkt fermentacji. Po przywróceniu warunków tlenowych PHB może zostać wykorzystany jako źródło węgla i energii w metabolizmie oksydacyjnym.
Wiele bakterii gromadzi w komórkach kwas fosforowy pod postacią ziaren polifosforanów, zwanych ziarnami wolutyny (po raz pierwszy opisano je u Spirillumvolutans).Fosforany zmagazynowane w ten sposób mogą być wykorzystane w przypadku ich braku w podłożu, umożliwiając komórce nawet kilkakrotne podziały lub wytworzenie przetrwalnika.
Siarkę w postaci silnie załamujących światło kuleczek gromadzą bakterie utleniające siarkowodór i siarczki do siarczanów .Ilość nagromadzonej siarki zależy od stężenia siarkowodoru. Przy jego braku nagromadzona siarka jest utleniana do siarczanu. Dla bakterii tlenowych siarka może być źródłem energii, dla beztlenowych donorem elektronów.
Porównać komórkę pro i eukariotyczną
Skład chemiczny komórek
skład chemiczny:
źródło węgla, które musi znajdować się w podłożu;
woda 70-86% suchej masy;
sucha masa: C:N:P 100:10:1
fosfor,
węgiel,
azot.
mikroelementy:
siarka,
cynk,
magnez,
żelazo,
mangan.
skład organiczny:
białka (42- 63% suchej masy, wychodzą w skład organelli - fimbrii, rzęsek, rybosomów,
cukry - stanowią do 10% suchej masy, u bakterii przeważnie cukrowce spotykane są w postaci polimerów, element ściany komórkowej u bakterii Gram -, element błony cytoplazmatycznej i białek strukturalnych, pełni funkcje zapasowe, buduje woski u gruźlicy.
wielocukry, stanowią do 10% suchej masy, odgrywają rolę materiałów zapasowych komórki (skrobia, glikogen)oraz budulca, z jakiego zbudowane są sztywne ściany komórkowe bakterii i śluzowe substancje otoczkowe
tłuszcze - ich zawartość zależy od typu bakterii, ich skład odmienny niż u innych org.
kwasy nukleinowe - zbudowane z łańcuchów nukleotydowych (RNA - 15% suchej masy, DNA)
witaminy i czynniki wzrostowe ilościowo stanowią małą frakcję masy komórkowej ale jako niebiałkowe składowe wielu enzymów odgrywają rolę bardzo dużą.
składniki popiołowe - ich ilość zależy od gatunku ale przede wszystkim od warunków hodowli i składu podłoża
Sposób pobierania pokarmu przez bakterie.
całą powierzchnią błony cytoplazmatycznej
DYFUZJA BIERNA - substancje przenikają zgodnie z gradientem stężeń tzn. z obszaru o wyższym stężeniu do obszaru o niższym stężeniu.
TRANSPORT AKTYWNY - cząsteczki przechodzą do komórki, ale wymaga to zużycia energii. Takim związkiem energetycznym jest ATP kwas adenozyno trójfosforanowy.
TRANSPORT UŁATWIONY (wbrew gradientowi stężeń) substancja, która chce wejść do komórki łączy się w obrębi błony z inna substancją zwaną nośnikiem. Tworzy się kompleks nośnik-cząsteczka, który, po przejąciu przez błony rozpada się. Cząsteczka zostaje w komórce. Nośnik wraca na swoje miejsce w błonie i czeka na ponowne tworzenie kompleksu.
Wielkocząsteczkowego - bakterie pobierają pokarm całą powierzchnią ciała okrytą ścianą komórkową i błoną cytoplazmatyczną. Bakterie nie potrafią wchłaniać cząstek w postaci cząstek czy kropelek, jak czynią to komórki eukariotyczne. Pobieranie pokarmu jest jednoznaczne z przeprowadzeniem go przez osłony komórkowe. Pobieranie musi być poprzedzone rozbiciem makrocząsteczek na pochodne związki drobnocząsteczkowe. Proces ten nazywamy trawieniem lub rozkładem i ma on najczęściej charakter hydrolizy i przebiega z udziałem enzymów. Bakterie muszą najpierw rozłożyć pokarm na zewnątrz komórki.
Wchłanianie zw. drobnocząsteczkowe- Jest on pobierany całą powierzchnią ciała, ściana komórkowa działa na zasadzie sita - przepuszcza cząstki drobnocząsteczkowe, a nie przepuszcza wielkocząsteczkowych, działa wybiórczo. Właściwym organem pobierającym pokarm jest błona cytoplazmatyczna - działa ona wybiórczo - wybiera to co jest jej potrzebne.
Pobieranie pokarmu nie jest procesem biernym. Jednym z mechanizmów jest przenikanie na zasadzie wybiórczej rozpuszczalności w tłuszczach. Największe znaczenie - odżywianie przez przenośniki, czyli bakteria rozpoznaje związek, wiąże go za pomocą białek - białka wiążące (wiązanie następuje w błonie, przekaźniki przekazują pokarm do cytoplazmy, przekaźniki to peptydy cykliczne lub łańcuchowe).Źródła pierwiastków podstawowych
węgiel:
autotrofy- z dwutlenku węgla
heterotrofy - ze związków organicznych
azot:
sole amonowe, azotany/azotyny, (sole mineralne) - rośliny
azot atmosferyczny - bakterie symbiotyczne (np. brodawkowe z rodzaju Rhizobium); żyjące w warunkach tlenowych Azothobacter, beztlenowych Colstridium; sinice i grzyby.
aminokwasy (organiczne):
auksotrofy - maja dość duże wymagania odżywcze (dostarczenie gotowych składników).
fosfor
spotykamy go pod postacią nieorganiczną w formie jonu fosforanowego
siarka
chemoautotrofy - jon siarczanowe, siarczany tiosiarczany
auksotrofy - aminokwasy -cysteina i metionina; witaminy-biotyna
Czy różnią się prototrofy od auksotrofów
Źródła fosforu i siarki
fosfor - jest czerpany przez bakterie w postaci, w której spotkamy go w związkach organicznych, a jako jon fosforanowych, który zwykle w pokarmie może mieć postać fosforanu nieorganicznego
siarka - większość bakterii czerpie siarkę z jonów siarczanowych, w związkach biogennych występuje w postaci zredukowanych grup sulfhydrylowych; siarczyny; gotowe związki, które zawierają siarkę; siarka zredukowana -siarczyny, tiosiarczany; związki organiczne zawierające siarkę - cysteina, metionina,biotyna
węgiel - heterotrofy i autotrofy
Enzymy Enzymy są to biologicznie czynne białka, które regulują szybkość reakcji biochemicznych; przyspieszają reakcje, ale tylko te, które są termodynamiczne możliwe tzn. te, w który przebiegu wyzwala się energia, a produkty ostateczne zawierają mniej energii, niż wyjściowe. Obniżają energię aktywacji, same jednak nie ulegają przemianie, dlatego nie zużywają się bezpośrednio w wyniku reakcji. Enzymy są wysoce swoiste, tzn. działają tylko na określony, sobie właściwy substrat i katalizują tylko dana reakcję.. Większość enzymów składa się z: - części białkowej, czyli apoenzymu, - części niebiałkowej, czyli grupy prostetycznej lub koenzymu. Reakcje enzymatyczne Na powierzchni cząsteczki enzymu znajduje się zagłębienie będące miejscem wiązania substratu, nazwane centrum aktywnym. Przestrzenne dopasowanie substratu do centrum aktywnego enzymu umożliwia ich związanie i wytworzenie kompleksu enzym - substrat (E-S), a w efekcie obniżenie energii aktywacji reakcji, której następnie ulegnie substrat (energia aktywacji to taka ilość energii, która jest niezbędna do zapoczątkowania reakcji chemicznej). Ogólne równanie reakcji enzymatycznej katalizowanej przez enzym można zapisać w następujący sposób: ENZYM + SUBSTRAT ↔ KOMPLEKS E-S ↔ ENZYM + PRODUKT W ustaleniu odpowiedniej konformacji centrum aktywnego umożliwiającej utworzenie kompleksu E-S często biorą udział aktywatory, których role pełnią jony metali lub koenzymy. Na aktywność enzymów wpływają różne czynniki, wśród nich wymienić można temperaturę i pH. Wraz ze wzrostem temperatury rośnie szybkość reakcji enzymatycznych. Jednak po przekroczeniu pewnej wartości szybkość ta nagle spada, co jest wynikiem cieplnej denaturacji enzymu. Optymalna temperatura to taka, przy której szybkość katalizowanej reakcji jest największa. Nie można wyznaczyć takiej reguły w przypadku wpływu pH na aktywność enzymatyczną, ponieważ enzymy wykazują dużą różnorodność w zależności od środowiska, w jakim działają. Przykładem mogą być enzymy trawienne, spośród których amylaza ślinowa wykazuje optimum w środowisku obojętnym, pepsyna w kwaśnym, a trypsyna w zasadowym.
oksydoreduktazy - (dehydrogenazy, oksydazy) - Przenosza elektrony i wodory z substratu na jakiś akceptor
transferazy - transaminazy, przenoszą z jednego związku na drugi określona grupę chemiczną aminową, acetylową itp.
hydrolazy - rozkładają substrat na drodze hydrolizy, należą tu enzymy rozszczepiające białka- proteazy, celulozę-celulazy.
liazy - odszczepiają pewne grupy bez udziału hydrolizy
Izomerazy- enzym przebudowujący strukturę cząsteczki bez jej rozkładu
ligazy (syntetazy) - katalizują łączenie się 2 cząsteczek
Wyjaśnić pojęcia:
dostarczenie substancji konstrukcyjnych
wytworzeni odpowiedniej ilości energii
stworzenie odpowiedniego potencjału redukcyjnego
zabezpieczenie organizmu przed raptownymi zmianami warunków środowiska
pokarmu - heterotrofy
energii słonecznej - fotoautotrofy
energii utleniania prostych związków nieorganicznych - chemoautotrofy
regulacja hormonalna i nerwowa
wytwarzanie enzymów pod kontrola genetyczną.
Wyjaśnić pojęcia:
bierze udział w regulacji ciśnienia krwi oddziałując na receptory,
pełni funkcję akumulatora, jest przenośnikiem energii swobodnej (pełni rolę w wymianie, transporcie, magazynowaniu energii)
uczestniczy w wielu procesach oksydacyjno - redukcyjnych związanych z katabolizmem i uwalnianiem energii
główny akceptor elektronów w reakcjach utleniania substratów w łańcuchu oddechowym,
przenosi elektrony
bierze udział w reakcjach oksydoredukcyjnych
jest używany w reakcjach katabolicznych
używany w reakcjach anabolicznych
przenosi elektrony
bierze udział w reakcjach oksydoredukcyjnych
Auto i heterotrofia
energia świetlna, a organizmy które ją wykorzystują to fotoautotrofy. Należą do nich wszystkie rośliny zielone, glony, sinice i niektóre bakterie fotosyntezujące, bakterie purpurowe siarkowe (znajdując się w ciemności i w warunkach tlenowych mogą odżywiać się heterotroficznie, związkami organicznymi), niektóre pierwotniaki, glony, rośliny naczyniowe
energia uwalniana w czasie reakcji chemicznych. Energię uwalniana utlenianiu związków mineralnych wykorzystują chemolitotrofy, a typ odżywiania zwane jest chmosyntezą -bakterie nitryfikacyjne, siarkowe, żelazowe, wodorowe).
biofagi, tj. zjadające inne żywe organizmy lub ich tkanki,
się fitofagi (zjadające rośliny), czyli roślinożerców,
zoofagi (zjadające zwierzęta) - inaczej drapieżniki.
saprobionty żyjące w środowiskach zawierających rozkładające się szczątki różnych organizmów i odżywiające się martwą materią organiczną głównie pochodzenia roślinnego lub zwierzęcego Saprobionty dzieli się na:
mikrokonsumentów (bakterie, pierwotniaki, grzyby) nazywanych także reducentami, saprofitami lub osmotrofami,
makrokonsumentów (zwierzęta ), czyli saprofagi lub detrytofagi.
ksylofagi są to organizmy odżywiające się drewnem,
korpofagi - odżywiające się grzybami.
Typy procesów oddechowych u bakterii
układy oksydacyjno - redukcyjne - zdolność do pobierania i oddawania elektronów wyrażam tak zwanym potencjałem oksydoredukcyjnym, związki które łatwo oddają elektrony, a tym samym łatwo ulegają utlenieniu nazywamy związkami redukcyjnymi, natomiast te, które mają większą zdolność pobierania określane są jako związki utleniające. Związki można ułożyć zależnie od ich potencjału redukcyjnego i tak te o najniższym będą oddawały sąsiedniemu o wyższym potencjale itd. aż do ostatniego związku, którym jest tlen. Związki, które przekazują elektrony od najbardziej zredukowanego dawcy do najbardziej utlenionego biorcy elektronów, nazywamy przenośnikami elektronów. Chemolitotrofy mogą jako dawców wykorzystywać nieorganiczne związki zredukowane.
łańcuch oddechowy- elektrony oderwane od substratu oddechowego wędrują przez kolejne przenośniki aż na ostateczny akceptor, którym w oddychaniu tlenowym jest tlen. Łańcuch przenoszenia elektronów nazywamy łańcuchem oddechowym, a kierunek przepływu elektronów wyznaczają wielkości potencjału redukcyjnego
dehydrogenazy - enzymy katalizujące utlenianie substratu w procesach oddechowych, odłączają od substratu po 2 elektrony i łącznie z nimi 2 protony, są wysoce specyficzne
Omówić rozkład heksoz do stadium glikolizy
utlenianie glukozo- 6-fosforanu do rybulozo- 5- fosforanu z jednoczesnym wytworzeniem dwóch cząsteczek NADPH
przejście rybulozo-5-fosforanu do rybozo-5-fosforanu
Powiązanie szlaku pentozofosforanowego z glikolizą
Jeśli zapotrzebowanie w komórce na NADPH jest większe niż na rybozo-5-fosforan, zostaje on przekształcony we fruktozo-6-fosforan i aldehyd 3-fosfoglicerynowy, które są metabolitami glikolizy, zgodnie przedstawionymi reakcjami.
Jeśli natomiast zapotrzebowanie na rybozo-5-fosforan jest znacznie większe niż na NADPH, transketolaza i transaldolaza przekształcają fruktozo-6-fosforan i aldehyd 3-fosfoglicerynowy, pobrane z glikolizy, w rybozo-5-fosforan.
Rodzaje fermentacji
beztlenowa (właściwa) - beztlenowy rozkład substratu pokarmowego (cukru) w celu uzyskania energii (czyli rodzaj oddychania beztlenowego):
kwas octowy,
kwas cytrynowy,
kwas fumarowy
tlenowa - niepełny utlenianie substratu
kwas mlekowy;
kwas propionowy
homofermentatywne
heterofermentatywne
Czym się różni homofermentacja od hetero fermentacji
fermentacja mlekowa właściwa - jedyny powstającym produktem jest kwas mlekowy
fermentacja alkoholowa - (Saccharomyces) - powstaje etanol
fermentacja mlekowa - produktami tej fermentacji są; kwas mlekowy, etanol i CO2, powodują ja niektóre bakterie mlekowe, np. Lactobacillus plantarum, L. brevis
f. masłowa - Clostridium
propionowa - Propionibacterium, powstaje kwas propionowy, octowy, bursztynowy
f. acetonowa - wywołana przez niektóre Colstridia -aceton, butanol, CO2, H2
Oddychanie beztlenowe z redukcja związków mineralnych
Przekształcenie pirogronianu do ostatniego produktów w oddychaniu tlenowym.
Łańcuch oddechowy w oddychaniu tlenowym
Rozmnażanie bakterii (więcej informacji na ten temat w zeszycie Darii)
podział poprzeczny - z jednej komórki powstaja dwie identyczne, jedno pokolenie
pączkowanie (Hyphomicrobium sp, Rhodomicrobium sp)
fragmentacja nitek (promieniowce -bakterie przypominające grzyby, tworzą pseudogrzybnię, np. Nocardia sp. -jej fragmentacja w osadzie czynnym powoduje spienienie, wytwarza środki powierzchniowo czynne
pseudokonidia (promieniowce np. Stereptomyces sp. - wytwarza antybiotyki
rozpad nitek na komórki (pływki) - bakterie nitkowe opatrzone pochewkami np. Sphaerotilus natans - występuja w osadzie czynnym, odpowiedzialne za puchniecie osadu
tworzenie komórek gonidialnych (pseudokonidiów) przez końcową komórkę nitki (bakterie nitkowate opatrzone pochewkami np. Crenothrix sp.
tworzenie ciałek elementarnych
Chlamydie - bakterie bardzo małe
mikoplazmy- nie mają ściany komórkowej, najmniejsze tworzą formy przetrwalnikowe - ciałka elementarne
rozpad ciał olbrzymich (mikoplazmy)
Mikroflora autochtoniczna i allochtoniczna
Przedostają się do zbiorników wodnych podczas silnych opadów wraz ze spływem powierzchniowym.
Źródłem bakterii chorobotwórczych (patogennych) są głównie ścieki miejskie. Do wód przedostają się także na drodze infiltracji i spływów powierzchniowych bakterie patogenne pochodzące z gleby.
Rola powietrza w zakażeniu wody jest istotna w zasięgu gęsto zaludnionych miast i okręgów przemysłowych. W zakażeniu główną rolę odgrywają bakterie zawieszone w powietrzu w postaci bioaerozoli lub pyłu bakteryjnego. Wraz z opadami przedostają się one do wód powierzchniowych.
Bakterie chorobotwórcze przenoszone drogą wodną
pałeczki duru brzusznego, czyli pałeczki z gatunku Salmonella typhi,
a także inne Gram-ujemne bakterie z rodzaju Salmonella, które są przyczyną różnorodnych zakażeń przewodu pokarmowego, objawiających się wymiotami i biegunką.
Nieco rzadziej w zanieczyszczonych zbiornikach wodnych występują Gram-ujemne pałeczki z rodzaju, Shigella, które powodują czerwonkę bakteryjną.
Wirusy i pierwotniaki przenoszone droga wodną
Omówić organizmy wskaźnikowe
Mikroflora autochtoniczna i zymogeniczna w glebie
Od czego zależy przeżywalność mikroorganizmów w glebie
zawartości w glebie wody,
ciśnienia osmotycznego,
potencjału oksydacyjno redukcyjnego,
ph gleby,
temperatury,
natlenienia,
zawartości składników pokarmowych,
związków toksycznych,
światła.
Drobnoustroje występujące w glebie
pałeczki duru brzusznego z rodzaju Salmonella
pałeczki czerwonki - Shigella.,
beztlenowe laseczki wywołujące tężec - Clostridium tetani,
zgorzel gazową - Clostridium perfringens,
zatrucia pokarmowe - Clostridium botulinum
oraz tlenowe laseczki wąglika - Bacillus anthracis,
a także prątki - Mycobacterium tuberculosis.
Analiza helimitologiczna i mikrobiologiczna gleby
Oznaczenie obecności w glebie bakterii chorobotwórczych z rodzaju Salmonella. W glebie przeznaczonej do celów rolniczych nie mogą się znajdować bakterie należących do tego rodzaju.
Badania helmintologiczne polegające na określeniu liczby żywych jaj robaków jelitowych: glisty ludzkiej (Ascaris lumbricoides), glisty psiej (Toxocara sp.), włosogłówki (Trichuris trichiura), określeniu stadium ich rozwoju oraz stopnia odporności na różne warunki mikroklimatyczne.
nie zanieczyszczoną - nie wykrycie jaj,
słabo zanieczyszczoną - wykrycie pojedynczych jaj (1-3),
średnio zanieczyszczoną - wykrycie do 10 jaj,
silnie zanieczyszczoną - wykrycie 10 jaj i więcej
ogólnej liczby bakterii - wykonywane na podłożu agarowym, temperatura - 280C, czas inkubacji -48 h,
bakterii przetrwalnikujących (sporowych) - podłoże agarowe, temperatura 280C, czas inkubacji-48 h,
bakterii termofilnych - podłoże agarowe, temperatura 550C, czas inkubacji-24 h.
liczebności bakterii wskaźnikowych:
pałeczek grupy coli,
pałeczek grupy coli typu fekalnego (termotolerancyjnych), głównie pałeczki okrężnicy (Escherichia coli),
laseczek Clostridium perfringens,
Miano - najmniejsza ilość badanej gleby wyrażona w gramach, w której stwierdza się jeszcze obecność badanych bakterii.
Bakterie grupy coli - Gram-ujemne pałeczki niewytwarzające przetrwalników, rozwijające się w warunkach względnie beztlenowych, mające zdolność fermentowania laktozy z wytworzeniem kwasu i gazu w ciągu 48 h hodowania w temperaturze 35-37OC. Należą one do rodzaju Escherichia, Citrobacter i Enterobacter w obrębie rodziny Enterobacteriaceae.
Bakterie coli typu fekalnego (termotolerancyjne) pałeczki okrężnicy (Escherichia coli) - są to pałeczki z grupy coli, które poza wszystkimi właściwościami tej grupy (pałeczki Gram-ujemne, oksydazoujemne, niewytwarzające przetrwalników, względnie beztlenowe, fermentujące laktozę z wytworzeniem kwasu i gazu w temperaturze 370C w ciągu 48 h oraz dające charakterystyczne kolonie na pożywce Endo) mają ponadto zdolność wzrostu oraz przeprowadzania niektórych procesów biochemicznych, a w szczególności fermentowania laktozy z wytworzeniem kwasu i gazu również w temperaturze 440C
Bakterie Clostridium perfringens - są to nieruchliwe, beztlenowe laseczki Gram-dodatnie wytwarzające otoczki i wykazujące zdolność do fermentacji glukozy, laktozy, rozkładu żelatyny, a także redukcji siarczynów do H2S.
Bakterie z rodzaju Salmonella - chorobotwórcze pałeczki przewodu pokarmowego. Wiele gatunków bakterii tego samego rodzaju wywołuje salmonellozy, czyli zatrucia pokarmowe. Salmonella typhi wywołuje chorobę zwaną durem brzusznym.
Bakterie wytwarzające przetrwalniki (bakterie sporowe) - Gram-dodatnie laseczki (tlenowe i beztlenowe) wytwarzające przetrwalniki (zwane również endosporami). Należą głównie do rodzaju Bacillus i Clostridium.
Bakterie termofilne - to bakterie rozwijające się w wyższych temperaturach, 45-600C (optimum - 550C), głównie w oborniku i kompoście, stosowanych do nawożenia i mogą być wskaźnikiem ich obecności w glebach uprawnych.
„Ogólna” liczba bakterii - to wskaźnik zawartości związków organicznych w glebie.
W końcowym etapie procesu mineralizacji, w miarę wyczerpywania się łatwo rozkładalnych źródeł pokarmowych wzrasta ilość spor bakteryjnych Stosunek ogólnej liczby bakterii do liczby bakterii sporowych - świadczy o stopniu rozkładu w glebie związków organicznych.
Badania helmintologiczne - służą wykrywaniu robaków jelitowych w glebie, a głównie ich jaj (tzw. jaj „geohelmintów”):
glisty ludzkiej (Ascaris lumbricoides),
glisty psiej (Toxocara sp.),
włosogłówki (Trichuris trichiura).
Stadium tworzenia się larwy w jaju i okresy zdolności życiowych robaków (helmintów) w glebie, mogą być użyte do oceny, od jak dawna gleba jest zanieczyszczona.
Uzdatnianie wody:
sedymentacja (+)
obniżenie ogólnej liczby bakterii o 75-99%
promieniowanie słoneczne
flokulacja sedymentacyjna (zbijanie zanieczyszczeń w kłaczki)
drapieżnictwo
współzawodnictwo o pożywnie
pasożyty
(-)
rozwój form planktonowych (sinice, okrzemki, zielenice)
opad pyłu i mikroorganizmów
zanieczyszczenie przez ptactwo wodne
filtracja powolna (przez złoża pisaku o dużych średnicach)
(+) wytwarza się błona biologiczna (glony bakterie auto i heterotroficzne)
adsorpcja
wytrącanie
rozkład
kumulacja zanieczyszczeń
usuwanie bakterii allochtonicznych
(-) nadmierny rozwój glonów
zakwity
produkcja toksycznych metabolitów
odtlenienie zbiorników
koagulacja
siarczan glinu + wapń ->Co2, zakwit glonów (chlorella,Chlamydomans)
filtracja pospieszna - (przez bardzo drobny piasek)
(+) mało bakterii
cienka błona biologiczna
czasami konieczna inkubacja
(-)
przy dużej ilości materii organicznej rozmnaża się niepożądana mikroflora
duża przepuszczalność filtrów, przechodzi 60% drobnoustrojów z wody surowej
filtracja przez węgiel aktywny
(+)
adsorpcja zanieczyszczeń organicznych
biodegradacja
biokumulacja
nagromadzenie i namnażanie mikroorganizmów
(-)
przedostawanie się mikroorganizmów wtórnie do wody uzdatnionej
konieczność chlorowania i wtórna selekcja mikroorganizmów
dezynfekcja (ostatni etap)
ozonowanie - ozon jest najskuteczniejszym środkiem dezynfekującym, jego skuteczność zależy od czasu działania, zawartości węglanów, temperatury, zawartości związków organicznych
(+) wymaga znacznie mniejszych dawek niż chlor
utlenia substancje organiczne odpowiedzialne za problemy z zapachem, smakiem i barwa wody
(-) produkty uboczne ozonowania są niebezpieczne dla zdrowia
w czasie ozonowania powstają związki łatwo asymilowane przez bakterie
związki nietoksyczne mogą stać się toksyczne
po ozonowaniu trzeba zastosować jeszcze jakiś środek dezynfekujący
chlorowanie
może prowadzić do tworzenia szkodliwych produktów ubocznych
kwas podchlorawy wchodzi w reakcje ze związkami i powoduje, że są szkodliwe (THM - związki rakotwórcze)
filtracja
temperatura - autoklaw
ultradźwięk
ultrafiolet
dimeryzacja
Naturalne metody oczyszczania ścieków.
oczyszczanie w gruncie
pola nawadniane - ścieki wykorzystywane są do celów rolniczych, plon zwiększa się o ok. 20 %, roczna dawka - nie więcej niż 600 mm/ar.
Pola irygacyjne - różnica miedzy nimi a polami nawadnianymi polega na tym, że najważniejszym celem jest oczyszczenie ścieków a nie korzyści rolnicze. Dawka do 1500 mm/ar
\ Filtry gruntowe - do 3000 mm/ar, nie wykorzystuje się tych pól do celów rolniczych
stawy ściekowe - zbiorniki ziemne (najczęściej zwykłe stawy). Stawy są dużo bardziej wrażliwe na obciążenie ściekami. Przy większych obciążeniach można stosować natlenianie. Najczęściej łączy się kilka stawów ze sobą:
bakteryjny
glonowy
skorupiakowy
eukariota |
składnik/ cecha |
prokariota |
+ |
błona komórkowa |
+ |
+ |
cytoplazma |
+ |
+ |
Rekombinacja genetyczna |
+ |
mitochondrium |
= |
mezosom |
rybosom |
80S/70S |
rybosom bakteryjny |
+ |
jądro komórkowe |
- |
+ |
retikulum endoplazma tyczne |
- |
+ |
lizozym |
- |
+ |
mikrotubule |
- |
+ |
otoczka jądrowa |
- |
+ |
cytoszkielet |
- |
+ |
chloroplasty |
- |
+ |
aparat Gologiego |
- |
+ |
ruchy cytoplazmy |
- |
- |
wić |
+ |
- |
plazmid |
+ |
- |
genofor |
+ |
+/- |
ściana komórkowa |
+ |
prokariota |
eukariota |
-nie mają wyodrębnionego jądra otoczonego błoną, DNA występuje w cytoplazmie jako koliście zamknięta cząsteczka -twór zwany nukleoidem tworzony przez splątek bardzo długiej (do 1000 razy dłuższej niż cały organizm),podwójnej helisy kwasy deoksyrybonukleinowego, zanurzony w cytoplazmie i bezpośrednio stykający się z jej składnikami |
- wyraźnie wyodrębnione z cytoplazmy jądro komórkowe zawierające błonę jądrową, sok jądrowy, chromosomy i jąderko (jądro zawiera większą część genomu umieszczonego w zespole chromosomów) |
-brak centrometru i centrioli, wrzeciona mitotycznego, brak mitochondriów, plastydów, lizosomów (ich funkcje spełniają inaczej i prościej ukszt. cząstki cytoplazmatyczne lub funkcje te są rozdzielone pomiędzy pozostałe układy komórkowe) -brak retikulum endoplazmatycznego -błona komórkowa inaczej zbudowana,inny układ ściany komórkowej u form ją posiadających |
-chromosomy są replikowane w procesie zwanym mitozą, w chromosomach DNA jest poł. z zasadowymi białkami zwanymi histonami
-w komórce eukariotycznej znajdują się też organelle, jak mitochondria i chloroplasty (w roślinach)
-stosunkowo duże rybosomy eukariotyczne (80S) |
-podział wnętrza na przedziały jest mniej wyraźny niż u eukariotów -rybosomy stosunkowo małe (70S) -stosunkowo słabo zróżnicowane morfologicznie, można wyróżnić jedynie kilka podstawowych kształtów, będących pochodnymi kuli i prostych lub zakrzywionych cylindrów |
|
-różnorodność i zmienność właściwości metabolicznych -istnieją grupy prokariotów, które mogą żyć beztlenowo, inne grupy zdolne do wykorzystania energii słonecznej i zdolne do syntetyzowania swojej materii komórkowej ze zw. organicznych lub CO2 |
|
Pobór pokarmu zależy w pierwszym rzędzie od zdolności bakterii do samodzielnej syntezy składników komórkowych. Im więcej takich składników bakteria syntezuje, tym mniej ich potrzebuje jako niezbędnych w pokarmie. Dobór pokarmu zależy także od jego dostępności.
Pobieranie pokarmu możemy podzielić na wykorzystanie pokarmu wielkocząsteczkowego i drobnocząsteczkowego:
Auksotrofy drobnoustroje, które do wzrostu muszą pobierać z zewnątrz jakąś substancję wzrostową której same nie umieją wytworzyć np. aminokwas, witaminy czy zasady purynowe.
Mogą powstawać w wyniku mutacji szczepów drobnoustrojów mających zdolność do samodzielnego produkowania wyżej wymienionych czynników wzrostowych (prototrofy). Należą do nich salmonella, gronkowiec złocisty (Staphylococcus aureus). Są to organizmy, które wymagają więcej niż jednego związku.
Prototrofy - bardzo pierwotne organizmy prokariotyczne, heterotroficzne, pobierające z otoczenia proste związki chemiczne, od których dostępności zależy ich przetrwanie w danym środowisku. Są to np. pałeczki gram ujemne, laseczki tlenowe (Bacillus), laseczki beztlenowe (Clostridium), związkiem prostym mogą to być kwasy nukleinowe, aldehydy, ketony, alkohol, białka. W przeciwieństwie do auksotrofów wymagają tylko jednego związku.
Różnice:
Auksotrofy |
Prototrofy |
Skomplikowane, gotowe już związki chemiczne |
Proste związki chemiczne |
Wymagają więcej niż jednego związku |
Wymagają tylko jednego związku |
Enzymy dzielimy na grupy w zależności od typu przemian chemicznych przez nie wywołanych:
Metabolizm- ogół procesów fizycznych i chemicznych oraz towarzyszące im przemiany energii zachodzące w żywych komórkach i stanowiące podstawę życia. Do podstawowych zadań metabolizmu należą:
Źródłem energii jest energia zawarta w składnikach żywej komórki i odnawiane przez przyjmowanie;
Proces metabolizmu musi być stale kontrolowany, warunek ten jest spełniony przez liczne mechanizmy regulacyjne;
Metabolizm jest integralnie związane z organizmem żywym, ustaje dopiero w chwili śmierci. Dla podtrzymania metabolizmu każda komórka musi być układem otwartym, przez co w sposób ciągły przepływa materia i energia
Katabolizm - związany z rozkładem i pozyskiwaniem energii, rekcja egzoergiczna prowadząca do uwalniania energii. Powstające produkty mają niższy poziom energetyczny niż zużywane substraty, dochodzi do degradacji energetycznej.
Anabolizm - związany z syntezą i wzrostem komórki, stanowi reakcję syntezy złożonych związków organicznych z prostych związków. Reakcje te są endoergiczne tzn. aby zaszły potrzebna jest energia. Dzięki temu powstające produkty maja wyższy poziom energetyczny niż wykorzystywane substraty.
utlenianie biologiczne, oddychanie tkankowe, oddychanie komórkowe-
zespół reakcji biochemicznych zachodzących w żywych komórkach, dostarczających im niezbędnej energii; komórka pobiera tlen, a wydala dwutlenek węgla oraz wydziela ciepło; u większości organizmów odbywa się poprzez rozkład cukrów i tłuszczów oraz w cyklu Krebsa; z oddychaniem komórkowym sprzężone jest magazynowanie energii w postaci tzw. ATP (adenozynotrifosforanów), stanowiących centra gospodarki energetycznej komórki; oba te procesy zachodzą w każdej komórce, gł. w wewn. błonie mitochondrialnej.
Procesy utleniania katalizowane są przez enzymy oksydoredukcyjne przejmujące elektrony z utlenianych substratów organicznych na współdziałające z nimi koenzymy (np. FAD, NAD), za pośrednictwem których elektrony trafiają na ostateczne ich akceptory; akceptorami tymi mogą być inne związki organiczne lub mitochondrialny łańcuch oddechowy przenoszący elektrony na cząsteczki tlenu i generujący siłę protonomotoryczną
C6H12O6 + 6O2 → 6CO2 + 6H2O + energia (38 ATP)
ATP, adenozynotrójfosforan, nukleotyd zbudowany z cukru-rybozy, zasady azotowej-adeniny i 3reszt kwasu fosforowego. ATP wchodzi w reakcje tylko w obecności kationów metali dwuwartościowych (np. Mn2+,, Mg2+), z którymi tworzy kompleksy. ATP powstaje z ADP w wyniku fosforylacji substratowej (fosforylacja) oraz w procesie chemiosmozy (fosforylacja oksydacyjna i fosforylacja fotosyntetyczna).
Dinukleotyd nikotynoamidoadeninowy NAD - nukleotyd pełniący istotną rolę w procesach oddychania komórkowego, pierwszy przenośnik odbierający elektron od dehydrogenaz ,.
NAD+ -(dinukleotyd nikotynoamidoadenininowy) jest koenzymem mającym strukturę opartą na adeninie i 2 cząsteczkach rybozy powiązanych grupami fosforanowymi i pierścieniem forma utleniona dinukleotydu , wiąże jeden proton i dwa elektrony .
NADP+ -) jest koenzymem mającym strukturę opartą na adeninie i 2 cząsteczkach rybozy powiązanych grupami fosforanowymi i pierścieniem. Cząsteczka NADP+ różni się do NAD+ obecnością reszty fosforanowej przy węglu 2' rybozy nukleotydu adeninowego.
Dinukleotyd flawinoadeninowy, FAD -jest syntetyzowany z ryboflawiny i 2 cząsteczek ATP. Jego reaktywna częścią jest pierścień izoalaksazynowy. Wraz z FMN jest pierwszym akceptorem wodoru w łańcuchu oddechowym. Może przyjmować 2 elektrony. Pełni funkcję przenośnika elektronów i protonów (kationów wodorowych). Przenosi dwa protony i dwa elektrony, w efekcie czego utleniona forma FAD przechodzi odwracalnie w formę zredukowaną FADH2. Mononukleotyd flawinowy, FMN, jest tak jak FAD syntetyzowany z ryboflawiny i 2 cząsteczek ATP. (ryboflawino-5'-fosforan). Wraz z FAD jest pierwszym akceptorem wodoru w łańcuchu oddechowym. Może przyjmować jeden elektron.
Autotrofy, organizmy samożywne - zdolne do syntetyzowania związków organicznych z prostych związków nieorganicznych, syntezują one cukrowce i złożone związki azotowe z dwutlenku węgla, wody i soli mineralnych. Pobierają w pokarmie jedynie utlenione związki węgla (kwas węglowy lub CO2) i przy przetwarzaniu w związki organiczne musi je zredukować. Proces redukcji związków powstających przy odżywianiu się CO2 pochłania energie i wymaga jej dopływu z zewnątrz. Źródłem tej energii może być:
Heterotrofy organizmy cudzożywne, dla których źródłem energii wykorzystywanej do procesów życiowych jest pokarm w postaci gotowej materii organicznej. Należą do nich prototrofy i auksotrofy. Dla tych pierwszych niezbędny jest tylko jeden związek organiczny w pokarmie. Auksotrofy potrzebują poza tym jednym prostym związkiem organicznym jeszcze jakiegoś jednego lub wielu skomplikowanych związków organicznych np. witaminy albo aminokwasu. Prototrofy znajdziemy w środowisku ubogim w pokarm, jak gleba, czy woda. Heterotrofy dzieli się na
Te podstawowe grupy troficzne są dzielone następnie na podgrupy ze względu na rodzaj pobieranego pokarmu:
Heterotrofami są także wszystkie pasożyty odżywiające się kosztem roślin, roślinożerców i drapieżników. Również wśród roślin jest wiele heterotrofów, które nie mają chlorofilu i nie muszą syntetyzować materii organicznej, np. pasożyty roślinne i saprofity.
Autotrofy budują w swych komórkach złożone cząsteczki z prostych cząstek, takich jak dwutlenek węgla i woda. Heterotrofy z kolei potrzebują do ich produkcji substancji bardziej złożonych - monosacharydów czy aminokwasów.
oddychanie tlenowe - jest złożonym procesem, którego istotą jest oderwanie pary elektronów i odwodorowanie substratu przez specyficzne enzymy, dehydrogenazę i stopniowe przenoszenie elektronów przez szereg przenośników aż na tlen (ostatni biorca), przy czym każdemu etapowi przenoszenia elektronów towarzyszy wyzwolenie pewnej ilości energii
Oddychanie tlenowe (aerobowe) (wzięte z tego co opracowali ludzie) jest wielostopniowym biochemicznym procesem utleniania związków organicznych związanym z wytwarzaniem energii użytecznej metabolicznie. Oddychanie przebiega w każdej żywej komórce w sposób stały. Zachodzi ono nawet wtedy, gdy inne procesy metaboliczne zostaną zahamowane. Chociaż istnieją różnice w przebiegu procesu oddychania u poszczególnych grup organizmów, to zestaw enzymów katalizujących poszczególne reakcje składające się na oddychanie jest zbliżony u wszystkich organizmów żywych. Energia uwolniona w procesie utleniania związków organicznych pojawia się częściowo w postaci związku wysokoenergetycznego - ATP
C6H12O6 + 6 H2O => 6CO2 + 6 H2O + energia
Podstawowy typ oddychania, przebiegający w normalnych warunkach we wszystkich komórkach żywych organizmów
Zasadnicze etapy oddychania komórkowego u organizmów eukariotycznych przebiegają w mitochondriach
Mitochondria - organelle występujące w cytoplazmie komórek eukariotycznych oddychających tlenowo
Mitochondria zbudowane są z dwóch błon: zewnętrznej i wewnętrznej oraz amorficznej macierzy (matriks) otoczonej wewnętrzną błoną mitochondrialną.
Liczba grzebieni mitochondrialnych (fałdów wewnętrznej błony mitochondrium) odzwierciedla stopień aktywności metabolicznej mitochondrium
Etapy oddychania tlenowego:
1. Glikoliza
przekształcenie glukozy w dwie trójwęglowe cząsteczki pirogronianiu
utworzenie ATP i NADH
2. Tworzenie acetylo-CoA
utlenianie pirogronianu do dwuwęglowej cząsteczki octanu, który łączy się z koenzymem A
uwolnienie CO2 i NADH
3. Cykl kwasu cytrynowego
przemiany acetylo-CoA prowadzące do uwolnienia CO2, ATP, NADH i FADH2
4. Łańcuch oddechowy
utworzenie ATP i wody
Oddychanie beztlenowe - Niektóre organizmy żywe czerpią energię wyłącznie z beztlenowego rozkładu związków organicznych, czyli fermentacji lub rozkładu związków nieorganicznych, na przykład azotanów lub siarczanów (oddychanie azotowe, siarczanowe. Takie organizmy mogą żyć w środowisku całkowicie pozbawionym tlenu. Przykładem jest żyjąca w ludzkim przewodzie pokarmowym Escherichia coli oddychająca (uzyskująca energię) poprzez redukcję azotanów do azotynów; inna bakteria, z rodzaju Desulfovibrio, redukuje siarczany do siarkowodoru. Wobec tego oddychanie beztlenowe jest spotykane tylko u bakterii, które wykorzystują utlenione związki mineralne jako akceptory elektronów.
Redukcja azotanów (denitryfikacja) DENITRYFIKACJA - redukcja azotanów do amoniaku (denitryfikacja częściowa) lub azotu cząsteczkowego (denitryfikacja całkowita) zachodząca w warunkach beztlenowych pod wpływem niektórych gat. bakterii glebowych i wodnych (zw. denitryfikatorami), np. Micrococcus denitrificans, Thiobacillus denitrificans; końcowy etap oddychania beztlenowego, w którym azotany są akceptorem atomów wodoru; denitryfikacja całkowita jest jednym z ogniw obiegu azotu w przyrodzie; nie jest korzystna dla rolnictwa, gdyż zuboża glebę w przyswajalny dla roślin azot; użyteczna przy biol. oczyszczaniu ścieków o dużej zawartości azotanów.
Paracoccus dentrificans:
2 NO32- + 4 H+=> 2 NO22- + 2 H2O
2 NO22- + 4 H+ =>2 NO + 2 H2O
2 NO + 2 H+ =>N2O + H20
N2O + 2H+ => N2 + H2O
Redukcja siarczanów
Desulfovibrio sp bakterie bezwzględnie beztlenowe, występują w osadach dennych zbiorników wodnych, w odach podziemnych, zwłaszcza w pobliżu złóż ropy naftowej. Substratem są kwasy organiczne, powstają duże ilości kwasu siarkowego:
SO44- + 8 H+ => H2S + H2O + 2 OH2-
Redukcja węglanów i CO2
Methanobacterium sp. (bakterie metanogenne) -akceptorem jest CO2 lub weglany, powstającym związkiem metan
8 H+ + CO2 =>CH4 + 2 H2O
Oddychanie beztlenowe występuje u prokariotów, które nie są zdolne do wykorzystywania tlenu jako końcowego akceptora elektronów. Takie organizmy zwane zwane są bezwzględnymi beztlenowcami. Inne mikroorganizmy prokariotyczne mogą wykorzystywać oddychanie beztlenowe , jeśli akurat nie ma tlenu w środowisku. W czasie oddychania beztlenowego powstaje mniej energii. Alternatywnymi akceptorami elektronów mogą być azotany, siarczany czy dwutlenek węgla.
Jednym z najważniejszych procesów rozkładu heksoz jest glikoliza, gdzie w wyniku ciągu reakcji jedna cząsteczka glukozy zostaje przekształcona w dwie cząsteczki pirogronianu. GLIKOLIZA- Proces glikolizy zachodzi z udziałem jedenastu enzymów. Enzymy oraz wszystkie substraty dostarczane są do cytoplazmy komórki, gdzie proces ten się odbywa. Glikoliza jest przemiana beztlenową lecz może zachodzić również w warunkach tlenowych. Proces ten podzielony jest na dwa etapy.
I etap - dochodzi do ufosforylowania glukozy lub innego cukru będącego substratem glikolizy, np. fruktozy, sacharozy, glikogenu, skrobi. Do procesu tego zużywany jest ATP a produktem reakcji jest aldehyd 3-fosfoglicerynowy.
II etap - wyniku reakcji redukcji i utleniania powstaje kwas pirogronowy. W reakcji tej bierze udział dinukleotyd nikotynamidoadeninowy ( NAD+ ) a powstała energia kumulowana jest w cząsteczkach ATP.
glukoza + 2 Pi + 2 ADP + 2 NAD+ → 2 cząsteczki pirogronianu + 2 ATP + 2 NADH + 2 H+ + 2 H2O
Zysk reakcji glikolizy wynosi: 2 cząsteczki ATP oraz powstanie pirogronianu
Etapy glikolizy:
Szlak Entnera - Doudoroffa - zachodzi bez glikolizy. Szlak ten funkcjonuje u wielu bakterii (Pseudomanas, Acetobacter). Rozpoczyna się od degradacji glukozy pod wpływem heksokinazy i powstaje utleniony glukozo - 6 - fosforan; dehydrogenaza przekształca go w 6- fosfoglukonian . Na skutek dehyroatazy powstaje 2-keto-3deoksy-6-fosfoglukonian (KDPG). W skutek rozszczepienia KDPG powstaje aldehyd 3-fosfoglicerynowy i pirogronian. Szlak ten dostarcza komórce prekursorów biosyntetycznych. Bilans to 2 cząsteczki pirogronianu i 1 cząsteczka ATP. Róznica między tym szlakiem, a glikolizą jest taka, iż oprócz związków 3-węglowych, mogą także powstawać 4, 5 i 7 węglowe, ponad to ułatwiają aktywacje własnych składników komórkowych.
Szlaku heksozomonofosforanowym (HMP), nazywanym też szlakiem pentozowym - bezpośrednio łączy się z glikolizą. Podstawowym zadaniem reakcji jest wytwarzanie NADPH i utlenianie glukozo-6-fosforanu do rybulozo-5-fosforanu. Utlenianie cząsteczki glukozy daje 36 cząsteczek ATP
Fermentacja mlekowa - homofermentacja, jedynym produktem jest kwas mlekowy. Fermentacja mlekowa jest procesem przemiany cukrów do produktów końcowych, wśród których dominuje kwas mlekowy. Pozostałe produktu to: kwas octowy, aldehyd octowy, etanol, CO2, acetoina, diacetyl, butandiol. Ze względu na szlaki przemian cukrów klasyfikuje bakterie mlekowe na:
Przeprowadzają ja bakterie mlekowe:
-Leuconostoc - paciorkowce heterofermentatywne (Leuconostoc citrovorum - bywa używany jako dodatek do zakwasów przy wyrobie masła) - Lactobacillus - pałeczki homo- i heterofermentatywne (Lactobacillus bulgaricus - pałeczka bułgarska występująca w jogurcie, Lactobacillus viridescens - powoduje zielenienie mięsa peklowanego i surowych kiełbas). - Streptococcus - paciorkowce homofermentatywne (Streptococcus lactis paciorkowiec mlekowy, Streptococcus cremoris - paciorkowiec śmietanowy)
Równanie sumaryczne właściwej fermentacji mlekowej
C6H12O6 + bakterie mlekowe => 2CH3CHOHCOOH + 22,5 kcal
(cukier prosty -> kwas mlekowy + energia) Fermentacja propionowa - fermentacja wywoływana przez bakterie propionowe - należą do rodzaju Propionibacterium, - mają kształt pałeczek, - są względnymi beztlenowcami, - nie wytwarzają przetrwalników, - mają zastosowanie przy produkcji serów dojrzewających, np. sera edamskiego (wytwarzający się kwas octowy i propionowy nadają serom charakterystyczny, nieco ostry smak, a dwutlenek węgla powoduje powstawanie dziurek w serze), - bakterie te towarzyszą bakteriom mlekowym w zakwasach chlebowych, a niekiedy też w kiszonkach. Równanie sumaryczne fermentacji propionowej
CH3CHOHCOOH + bakterie propionowe => 2CH3CH2COOH + CH3COOH + CO2 + H2O + kcal
(kwas propionowy + bakterie → kwas octowy + energia)
Fermentacja octowa, proces utleniania alkoholu etylowego i innych alkoholi do kwasu octowego, katalizowany przez enzymy bakteryjne wytwarzane przez rodzaj Acetobacter:
Fermentacja masłowa - wywołują ja bezwzględne beztlenowce, przetrwalnikujące, gramdodatnie laseczki, Clostridium butrycium. Substratem obok cukrów prostych i dwucukrów jest także skrobia, pektyny, celuloza. Podczas heterofermentacji oprócz kwasu masłowego powstaje także : kwas octowy, alkohol etylowy i metan.
C6H12O6 -> CH3CH2CH2COOH + 2C02 + 2H2 + energia
Homofermentacja - powstaje jeden produkt:
Heterofermentacja - powstają więcej niż dwa produkty :
Oddychanie beztlenowe - ostatecznym biorcą elektronów jest zewnątrzpochodny związek organiczny (fumaran) lub utleniony związek nieorganiczny (azotan, siarczan, węglan). ATP powstaje w wyniku fosforylacji oksydatywnej. Łańcuch oddechowy krótszy niż przy oddychaniu tlenowym. Proces ten jest bardziej wydajny niż fermentacja, mniej wydajny w porównaniu z oddychaniem tlenowym.
-Redukcja azotanów do azotynów, NO, N2O, N2 lub NH3 Gazowe produkty opuszczają środowisko -- deniryfikacja - redukcja azotanów do amoniaku (denitryfikacja częściowa) lub azotu cząsteczkowego (denitryfikacja całkowita) zachodząca w warunkach beztlenowych pod wpływem niektórych gat. bakterii glebowych i wodnych (zw. denitryfikatorami), np. Micrococcus denitrificans, Thiobacillus denitrificans; końcowy etap oddychania beztlenowego, w którym azotany są akceptorem atomów wodoru; denitryfikacja całkowita jest jednym z ogniw obiegu azotu w przyrodzie; nie jest korzystna dla rolnictwa, gdyż zuboża glebę w przyswajalny dla roślin azot; użyteczna przy biol. oczyszczaniu ścieków o dużej zawartości azotanów. desulfurykacja -(MIkrococcus, Pseudomonas) NH3,NO2-, N2 redukcja węglanów Methanobacterium sp. (bakterie metanogenne) -akceptorem jest CO2 lub weglany, powstającym związkiem metan
Cykl Krebsa.
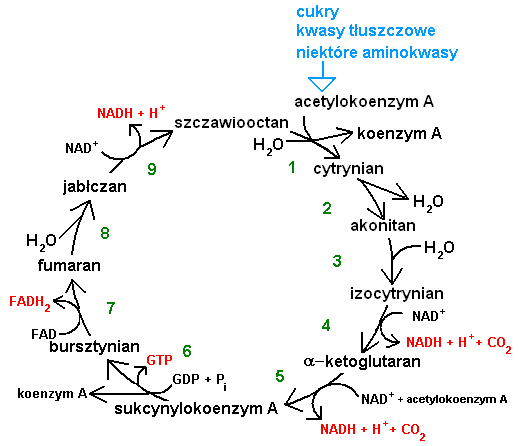
Cykl Krebsa, czyli cykl kwasu cytrynowego to cykl przemian metabolicznych, który przebiega w komórkach wszystkich organizmów oddychających tlenem. Został on odkryty w 1937 roku przez Hansa Krebsa i to od nazwiska tego biochemika bierze się jego nazwa.
Cykl Krebsa u eukariontów zlokalizowany jest wewnątrz mitochondriów - ważnych organelli komórkowych (u prokariontów przebiega w cytoplazmie). Składa się on z 9 etapów, katalizowanych przez 8 odrębnych enzymów (dwa etapy katalizuje ten sam enzym).
Zadaniem cyklu Krebsa jest utlenić związek o nazwie: acetylokoenzym A (acetylo-CoA) do 2 cząsteczek dwutlenku węgla (CO2), a pozyskaną w tym procesie energię ulokować w chemicznych nośnikach energii: GTP, NADH i FADH2.
Sumaryczny wzór cyklu Krebsa to:
acetylo-CoA + GDP + Pi + 3NAD+ + FAD + 2H20 → koenzym-A + GTP + 3NADH + 3H+ + FADH2 + 2CO2
Podczas jednego, pełnego obrotu cyklu Krebsa powstają 3 cząsteczki NADH, jedna cząsteczka FADH2 i jedna cząsteczka GTP. Najbardziej wszechstronnym nośnikiem energii w komórce jest ATP. GTP jest łatwo zamieniany na ATP przez odpowiedni enzym. Natomiast NADH i FADH2 biorą udział w mitochondrialnym łańcuchu oddechowym - przemianie, która zamienia energię tych zredukowanych związków na energię wiązań ATP. Niezbędnym uczestnikiem łańcucha oddechowego jest tlen.
Jedna cząsteczka NADH pozwala wyprodukować 3 cząsteczki ATP, a jedna cząsteczka FADH2 - 2 cząsteczki ATP. Nietrudno więc obliczyć, że jeden pełny obrót cyklu Krebsa pozwala wytworzyć 12 cząsteczek ATP - uniwersalnego nośnika energii dla komórki (3 x 3 + 1 x 2 + 1 = 12).
Acetylokoenzym A jest cząsteczką centralną dla metabolizmu tlenowców. Jest on produktem deaminacji wielu aminokwasów, cząsteczką końcową tzw. beta-oksydacji kwasów tłuszczowych, a także związkiem, w który przeprowadzany jest pirogronian - produkt glikolizy monocukrów.
A więc to dzięki tej "zbiorczej" cząsteczce, komórka może uzyskiwać energię zarówno z aminokwasów (składniki białek), jak i z tłuszczów i cukrów.
Zauważmy, że drugim substratem, do którego przyłączany jest acetylo-CoA na początku cyklu Krebsa, jest szczawiooctan. Ale mamy tu do czynienia z cyklem, czyli szczawiooctan jest także jednym z produktów końcowych. Cały cykl bierze zatem udział w utlenianiu acetylokoenzymu A, ale pozostaje on niezmienny dzięki swojej cykliczności (koniec przechodzi w początek). Jeśli coś uczestniczy w reakcji, a mimo to nie zmienia się, to jest to katalizator. A więc możemy traktować cały cykl Krebsa jak jeden, złożony katalizator.
W przypadku oddychania tlenowego na wskutek utleniania biologicznego z utlenionego substratu za pośrednictwem enzymu dehydrogenazy. Przy reakcjach utlenienia substancji organicznych najczęściej dochodzi do oderwania dwóch atomów wodoru od utlenianego związku, co nosi nazwę reakcji odwodorowania. Poszczególne procesy utleniania są tylko składowymi ogólnie pojętej reakcjo oksydoredukcyjnej. Przy utlenienie jednego związku zachodzi jednocześnie redukcja innego, gdyż oderwane od ultenianego związku elektrony są przyłączane do związku podlegającego redukcji. Wielkością określającą zdolność do przyjmowania i oddawania elektronów jest potencjał oksydoredukcyjny. Związek oddający elektrony jest określany jako reduktor, a związek przyjmujący elektrony jest utleniaczem. Jak wcześniej wspomniano enzymem katalizującym oderwanie dwóch elektronów i dwóch protonów wodorowych jest dehydrogenaza, odznaczająca się wysokim stopniem specyficzności, dzięki czemu katalizuje reakcje utlenienia tylko specyficznego dla niej substratu. Oderwane elektrony są przenoszone za pośrednictwem odpowiednich związków określanych przenośnikami elektronowymi na atom tlenu, ulegający w ten sposób redukcji i w postaci cząsteczki wydzielany jest do atmosfery.
W reakcji przeniesienia elektronów i protonów wodorowych na atom tlenu wydziela się duża porcja energii. Dlatego też cały proces transportu elektronów na tlen odbywa się za pośrednictwem związków takich jak koenzymy NAD, FAD oraz szeregu cytochromów, które razem tworzą tzw. łańcuch oddechowy. Na poszczególnych etapach transportu elektronów na cząsteczkę tlenu dochodzi to stopniowego uwalniania energii, która następnie posłuży przy reakcjach syntezy komórkowej. Wydzielana energia magazynowana jest w wysokoenergetycznych i łatwo dostępnych cząsteczkach ATP, które zużywane są trakcie kolejnych przemian. Niewielka ilość energii wydzielana jest w postaci ciepła.
Oddychanie (nie mylić z wymianą gazową!) jest procesem uwalniania energii zawartej w związkach organicznych. Może odbywać się przy pomocy tlenu z wydzielaniem CO2 i wody:
C6H12O6 + 6O2 → 6CO2 + 6H2O + energia
W taki sposób oddychają bezwzględne tlenowce (aeroby), które żyją tylko w obecności tego pierwiastka. Względne beztlenowce (anaeroby) są w stanie istnieć w przy niewielkich stężeniach tlenu, jednak energie czerpią z oddychania beztlenowego poprzez proces fermentacji bądź redukcji.
Mikroflora autochtoniczna wód - drobnoustroje naturalnie bytujące i rozmnażające się w środowisku wodnym.
Bakterie autochtoniczne
Wyróżniamy wśród nich: fotoautotrofy, chemoautotrofy i chemoorganotrofy.
- Bakterie fotosyntetyzujące (fotoautotrofy)
Wśród autotrofów fotosyntetyzujacych znajdują się bakterie purpurowe i zielone. Bakterie
purpurowe z uwagi na metabolizm dzielimy na następujące grupy:
● Bakterie zielone nitkowate (Chloroflexaceae),
● Bakterie zielone siarkowe (Chlorobiaceae),
● Bakterie purpurowe siarkowe (Chromatiaceae i Ectothiorhodaceae),
● Bakterie purpurowe niesiarkowe (Rhodospirillaceae),
● Heliobakterie (Heliobacteriaceae).
Fotosynteza bakteryjna przebiega nieco odmienne w porównaniu do fotosyntezy roślinnej.
Jest to przede wszystkim proces beztlenowy i wymaga obecności zredukowanych związków mineralnych oraz nie towarzyszy jej wydzielanie tlenu, lecz powstawanie utlenionych związków nieorganicznych lub organicznych. Barwniki asymilacyjne bakterii odznaczają się zdolnością do absorpcji światła w zakresie fal długich (podczerwieni) niepochłanianych przez rośliny zielone. Działalność fotosyntetyczną w wodach powierzchniowych prowadzą głównie glony i rośliny wyższe
natomiast fotosynteza bakteryjna ma mniejsze znaczenie.
- Bakterie chemosyntezujące (chemoautotrofy)
Chemoautotrofy czerpią energię z procesu utleniania związków nieorganicznych. W zależności od natury utlenianego substratu wyróżniamy: bakterie nitryfikacyjne, żelaziste, siarkowe i wodorowe.
● Rola bakterii nitryfikacyjnych w wodach polega głównie na utlenianiu jonów amonowych i azotynów a tym samym ich usuwaniu. Związki te w większych ilościach mogą być szkodliwe dla organizmów wodnych oraz zdrowia człowieka (w przypadku pobierania takiej wody do celów wodociągowych). Ponadto, wytwarzanie przez te bakterie azotanów jest podstawowym procesem dostarczającym roślinom wodnym najkorzystniejszego dla nich źródła azotu.
● Bakterie żelaziste rozwijają się w wodach o zawartości żelaza dwuwartościowego w zakresie 0,15-8,5 mg/dm3, a ich niekorzystny wpływ może wynikać z „obrastania” przez nie różnych przedmiotów znajdujących się w wodzie (kamienie, liście i łodygi roślin podwodnych, powierzchnie skrzeli ryb i narybku). Ujemna ich rola polega także na tym, że przyczyniają się do korozji i zarastania rur kanalizacyjnych czy wodociągowych oraz różnych konstrukcji metalowych. Najbardziej rozpowszechnione bakterie żelaziste Leptothrix ochracea i Crenothrix polyspora należą do bakterii
nitkowatych, które charakteryzują się tym, że pojedyncze komórki tworzą nitki, otoczone cieńszą lub grubszą pochewką utworzoną z galaretowatej substancji. Odkładane w komórkach związki żelaza zabarwiają nitki na kolor żółty lub ciemnobrązowy. Bakterie żelaziste w wodach słodkich występują bardzo licznie. Szczególnie w wodzie studziennej i źródlanej, gdzie dostrzec można większe ich skupienia. Masowo występują niekiedy w błotnistych potokach, moczarach i stawach.
● Bakterie siarkowe występują głównie w wodach zawierających siarkowodór, który dla większości drobnoustrojów jest toksyczny a dla nich stanowi warunek egzystencji. Spotyka się je w źródłach mineralnych zawierających siarkowodór pochodzenia geologicznego oraz w wodach silnie zanieczyszczonych, gdzie powstaje on jako produkt beztlenowego rozkładu białek. Typowymi przedstawicielami bakterii siarkowych są: bakteria poruszające się ruchem ślizgowym Beggiatoa alba
oraz przytwierdzona do podłoża Thiothrix nivea. Formy pojedyncze bakterii siarkowych to:
▪ Thiobacillus thioparus - odkłada siarkę pochodzącą z utlenienia tiosiarczanów,
▪ Thiobacillus thiooxidans - rośnie w środowisku kwaśnym przy pH 1,0-4,0,
▪ Thiobacillus ferroxidans - oprócz tiosiarczanów i tetrationianów ma zdolność do
utleniania soli żelazawych,
▪ Thiobacillus denitrificans - jest względnym beztlenowcem i wykazuje zdolność
wykorzystywania azotanów jako elektronobiorców przy utlenianiu siarkowodoru.
W warunkach tlenowych funkcje tę spełnia tlen.
● Bakterie wodorowe posiadają zdolność utleniania wodoru z wykorzystaniem tlenu cząsteczkowego jako ostatecznego akceptora elektronów. Najczęściej odżywiają się jednak heterotroficznie, na samożywny tryb życia przechodzą dopiero, gdy w środowisku pojawi się wodór. Najbardziej rozpowszechnione są gatunki należące do rodzaju Hydrogenomonas. Do bakterii wodorowych zaliczyć można Micrococcus denitrificans przeprowadzający utlenianie wodoru przy równoczesnej redukcji azotanu do azotu cząsteczkowego a także Desulfovibrio desulfuricans przeprowadzające
utlenianie wodoru przy równoczesnej redukcji siarczanu do siarkowodoru.
Bakterie heterotroficzne (chemoorganotrofy)
Przeważająca część bakterii autochtonicznych występujących w zbiornikach wodnych to bakterie chemoorganotroficzne należące do grupy saprofitów, odżywiających się martwą materią pochodzenia roślinnego i zwierzęcego. Do typowych bakterii utrzymujących się w całej masie wody, to jest w postaci bakterioplanktonu, należą urzęsione pałeczki Gram-ujemne, reprezentujące takie rodzaje jak: Pseudomonas, Achromobacter, Alcaligenes, Vibrio i Aeromonas, a także Gramdodatnie ziarniaki z rodzaju Micrococcus, krętki oraz bakterie spiralne z rodzaju Spirillum Na podwodnych częściach roślin wyższych, a także na podwodnych cząstkach stałych osiedlają się liczne bakterie stylikowe (np. Caulobacter), pochewkowe, nitkowate, a także pączkujące (np. Hyphomicrobium), które tworzą perifiton. W osadach dennych rozwijają się zazwyczaj beztlenowe bakterie gnilne, beztlenowe. bakterie celulolityczne, oraz liczne chemoorganotrofy beztlenowe takie jak np. bakterie z rodzaju Desulfovibrio, które redukują siarczany do siarkowodoru, oraz nie mniej liczne - beztlenowe bakterie metanogenne, które redukują różne związki organiczne do metanu.
Bakterie allochtoniczne - pochodzące z zewnątrz
Wody o dużej żyzności, a także silnie zanieczyszczone wody powierzchniowe obfitują w pochodzące z zewnątrz bakterie saprofityczne i pasożytnicze wśród których najliczniej reprezentowaną grupą są Gram-ujemne pałeczki jelitowe Escherichia coli oraz bakterie z rodzaju Proteus, Klebsiella i Enterobacter, a także pałeczki z gatunku Pseudomonas aeruginosa oraz rodzaju Arthrobacter czy Corynebacterium. Ponadto, do allochtonicznych bakterii wodnych należy zaliczyć Gram-dodatnie laseczki z rodzaju Bacillus i Clostridium, które są wypłukiwane z gleby.
Do najbardziej typowych bakterii bezwzględnie chorobotwórczych pojawiających się
w zanieczyszczonych wodach powierzchniowych należą
W krajach tropikalnych, w wodach powierzchniowych często spotyka się bakterie z gatunku, Vibrio cholerae, czyli przecinkowce cholery.
Ponadto w wodach zanieczyszczonych spotyka się prątki gruźlicy Mycobacterium tuberculosis oraz krętki z rodzaju, Leptospira. Te ostatnie bakterie wywołują żółtaczkę bakteryjną.
Poza wymienionymi bakteriami bezwzględnie chorobotwórczymi w wodach powierzchniowych są liczne bakterie Gram-ujemne, które określa się mianem drobnoustrojów oportunistycznych, czyli warunkowo chorobotwórczych. Należą one do rodzajów Pseudomonas, Aeromonas, Klebsiella, Flavobacterium, Enterobacter, Citrobacter, Serratia, Acinetobacter, Proteus i Providencia. Wszystkie te pałeczki wchodzą w skład normalnej flory jelitowej i nie są w zasadzie chorobotwórcze, o ile bytują one w przewodzie pokarmowym ludzi i zwierząt. W pewnych przypadkach bakterie te przedostają się jednak z przewodu pokarmowego do innych narządów i wówczas mogą stać się przyczyną wielu chorób np. zapalenia układu moczowego, zapalenia dróg oddechowych, a także posocznicy, czyli uogólnionego zakażenia wszystkich narządów wewnętrznych.
Wirusy
Oprócz bakterii chorobotwórczych wody powierzchniowe, do których odprowadzane są
ścieki bytowo-gospodarcze zawierają zawsze znaczne ilości innych drobnoustrojów
chorobotwórczych np. wirusa polio, który powoduje porażenie dziecięce, czyli chorobę
Heinego-Medina. Nawet w nieznacznie zanieczyszczonej wodzie rzecznej występują
enterowirusy, które wywołują schorzenia jelit.
Wirusy jelitowe, które mogą być przenoszone przez wodę
i choroby przez nie wywoływane
Wirusy |
Liczba typów |
Postacie i zespoły chorobowe
|
Poliovirus |
3 |
Porażenia, zapalenie opon mózgowych, gorączka |
ECHO |
34 |
Zapalenie opon mózgowych, choroby układu oddechowego, wysypka, biegunka, gorączka |
Coxsackie A |
23 |
Herpangina, choroby układu oddechowego, zapalenie opon mózgowych, gorączka |
Coxsackie B |
6 |
Zapalenia mięśnia sercowego, wrodzone wady serca, wysypka, gorączka, zapalenie opon mózgowych, choroby układu oddechowego, pleurodynia |
Enterowirusy |
4 |
Zapalenie opon mózgowych i mózgu, choroby układu oddechowego, ostre krwotoczne zapalenie spojówek, gorączka |
Wirus wzw A |
1 |
Wirusowe zapalenie wątroby |
Wirus Norwalk |
1 |
Epidemiczne biegunki, gorączka |
Parwowirusy |
3 |
Towarzyszą chorobom układu oddechowego |
Adenowirusy |
41 |
Choroby układu oddechowego, zakażenia oczu, biegunki |
Rotawirusy |
4 |
Epidemiczne biegunki (głownie u dzieci) |
Reowirusy |
3 |
Choroby dróg oddechowych |
Pierwotniaki
Zanieczyszczona woda może być przyczyną schorzeń przewodu pokarmowego
spowodowanych przez pierwotniaki. Większość pasożytniczych pierwotniaków produkuje cysty, które są w stanie przetrwać poza organizmem gospodarza w niekorzystnych warunkach środowiskowych. Gdy warunki te ulegają poprawie z cyst rozwijają się tzw. trofozoity, postacie wegetatywne występujące u człowieka.
Pierwotniak chorobotwórczy |
Choroba |
Objawy
|
Giardia lamblia(wiciowiec)
|
giardioza |
Chroniczne biegunki, skurcze brzucha, wzdęcia, ubytek masy ciała, zmęczenie |
Cryptosporidium parvum (sporowiec)
|
kryptosporidioza |
Bóle brzucha, brak łaknienia, wodniste biegunki, ubytek masy ciała |
Entamoeba histolytica (korzenionóżka)
|
amebioza |
Od łagodnej biegunki z krwią i śluzem do postaci ostrej z biegunką, gorączką i dreszczami |
Acanthamoeba castellani (korzenionóżka)
|
Pełzakowate zapalenie opon mózgowych i mózgu
|
Objawy ze strony centralnego układu nerwowego
|
Naegleria gruberi (korzenionóżka)
|
Pełzakowate zapalenie opon mózgowych i mózgu
|
Przedostaje się przez nos do mózgu ludzi kąpiących się, wywołuje ostre objawy zapalenia mózgu i opon mózgowych, kończące się śmiercią zakażonego osobnika |
Balantidium coli (orzęsek)
|
Czerwonka balantydiowa
|
Krwawa biegunka z powodu owrzodzenia ścian jelita grubego, w które wwierca się orzęsek
|
Obowiązujące normy oparte są na pośrednim wnioskowaniu o obecności mikroorganizmów chorobotwórczych na podstawie liczebności w wodzie bakterii wskaźnikowych, które stale żyją jako saprofity w przewodzie pokarmowym człowieka i zwierząt wyższych. Ich obecność w wodzie świadczy o jej zanieczyszczeniu fekalnym, a zatem również o niebezpieczeństwie zakażenia wody mikroorganizmami chorobotwórczymi. Bakterie pełniące rolę wskaźników sanitarnych powinny spełniać następujące warunki:
h) muszą być stale obecne w przewodzie pokarmowym człowieka, co pozwala zawsze na
wykrycie kałowego zanieczyszczenia wody,
i) do grupy organizmów wskaźnikowych powinny także należeć formy nie
przetrwalnikujące, co umożliwia wykrycie świeżego fekalnego zanieczyszczenia wody,
j) ich identyfikacja musi być możliwa przy użyciu łatwo dostępnych metod,
k) długość życia bakterii wskaźnikowych w środowisku zewnętrznym musi być większa
niż długość życia gatunków chorobotwórczych,
l) liczebność bakterii wskaźnikowych w jelicie człowieka i kale powinna być duża;
m) nie powinny się one rozmnażać w środowisku wodnym.
Jakie bakterie wskaźnikowe wykorzystywane są do oceny jakości zdrowotnej wody?
W rutynowej pracy laboratoriów, prowadzących nadzór sanitarno-epidemiologiczny,
niemożliwe jest stałe badanie wody w kierunku wykrywania wszystkich drobnoustrojów
chorobotwórczych i potencjalnie chorobotwórczych, które mogą w niej występować.
Dlatego też badania rutynowe koncentrują się przede wszystkim na wykrywaniu bakterii
wskazujących na kałowe zanieczyszczenie wody. Do oceny jakości sanitarnej wody
wykorzystywana jest mikroflora saprofityczna zasiedlająca jelito grube człowieka.
Przyjęto następujące wskaźniki fekalnego zanieczyszczenia wody:
● bakterie grupy coli,
● bakterie grupy coli typu fekalnego,
● paciorkowce kałowe,
● laseczki z rodzaju Clostridium, redukujące siarczyny
oraz w niektórych przypadkach
● gronkowce koagulazo-dodatnie,
● Pseudomonas aeruginosa.
Bakterie z grupy coli
Bakterie grupy coli to przede wszystkim szczepy Escherichia coli oraz drobnoustroje z
rodzaju Enterobacter, Citrobacter i Klebsiella. Wykrywane są one na podłożach z laktozą
po inkubacji w temperaturze 37 oC. i tylko te nieliczne szczepy z rodzajów Enterobacter, Citrobacter i Klebsiella, które mają
zdolność fermentacji laktozy w temperaturze 44 oC.
Obecność w badanej próbce wody bakterii grupy coli lub bakterii grupy coli typu
kałowego świadczy o stosunkowo świeżym zanieczyszczeniu wody kałem, ściekami,
glebą lub gnijącym materiałem roślinnym. W zasadzie dla większości rodzajów wód
zalecane jest oznaczanie liczby bakterii obu grup coli.
Paciorkowce kałowe
Paciorkowce kałowe charakteryzują się nieco dłuższym okresem przeżywalności
w środowisku wodnym i większą opornością na środki dezynfekcyjne, niż bakterie grupy
coli. Termin ten obejmuje drobnoustroje z rodzajów Enterococcus i Streptococcus
należące do grupy serologicznej Lancefield D.
Stwierdzenie obecności paciorkowców kałowych w badanej próbce wody w liczbie
znacznie przewyższającej liczbę bakterii grupy coli, sugerować może zanieczyszczenie
wody kałem zwierzęcym lub ściekami, pochodzącymi z ferm hodowlanych.
Laseczki z rodzaju Clostridium
O starym, odległym w czasie zanieczyszczeniu kałowym może świadczyć wykrycie
w badanej próbce wody bakterii redukujących siarczyny (głównie szczepy Clostridium
perfringens); ich przetrwalniki mogą zachować żywotność przez wiele lat
w niesprzyjających warunkach. Clostridia redukujące siarczyny są też bardzo dobrym
wskaźnikiem prawidłowości prowadzonych procesów uzdatniania wody, takich jak
koagulacja, sedymentacja i filtracja.
Przetrwalniki tych bakterii, a wraz z nimi również cysty pasożytniczych pierwotniaków
(Cryptosporidium parvum, Giardia lamblia) powinny być wyeliminowane właśnie w tych
etapach uzdatniania wody, gdyż są one bardzo oporne na działanie środków
dezynfekcyjnych. Wykrycie bakterii z rodzaju Clostridium jest technicznie znacznie
bardziej proste od poszukiwania pierwotniaków pasożytniczych i daje dużą pewność,
że woda uzdatniona jest wolna od pierwotniaków i jaj robaków chorobotwórczych
(helmintów).
Gronkowce
Gronkowce wykorzystywane są głównie przy ocenie jakości sanitarnej kąpielisk.
Kąpieliska bowiem są przyczyną szeregu zakażeń związanych z infekcjami dróg
oddechowych, skóry i oczu. W tym przypadku analiza oparta o standardowe indykatory
(Escherichia coli) jest niewystarczająca.
Często spotykane infekcje skóry powodowane są przez Staphylococcus aureus. W związku
z tym, że obecność tego mikroorganizmu w wodzie wynika z aktywności człowieka, został
on wybrany jako wskaźnikowy dla kąpielisk.
Pseudomonas aeruginosa
Obok wymienionych elementów analizy sanitarnej proponuje się obecnie dodatkowo
wykrywanie bakterii z gatunku Pseudomonas aeruginosa w wodzie do picia i na potrzeby
gospodarcze a także w wodzie dla zakładów kąpielowych oraz w wodach powierzchniowych. Przedstawicieli tego gatunku wyizolowano z kału ludzkiego oraz
w przypadkach zakażeń organizmu - z dróg moczowych, ucha środkowego, ropiejących
ran itp. Bakterie te stanowią potencjalny czynnik chorobotwórczy dla ludzi i zwierząt. Poza tym występują one powszechnie w wodach powierzchniowych i glebie. Warto także podkreślić, że mogą one bytować w chlorowanej wodzie, gdyż odznaczają się znaczną odpornością na zabiegi dezynfekcyjne.
Ogólna liczebność bakterii
W badaniach rutynowych określa się również ogólną liczbę bakterii (liczbę jednostek
tworzących kolonie, w skrócie jtk. lub cfu, z ang. colony forming units) obecnych w 1 ml
wody, po wykonaniu posiewu próbki na agar odżywczy i inkubacji w temperaturach 22oC
przez 72 godz. (psychrofile), oraz w 37oC przez 24 godz. (mezofile).
Ogólna liczebność bakterii psychrofilnych
W niższej temperaturze rosną przede wszystkim nie chorobotwórcze bakterie wodne.
Należy jednak mieć na uwadze fakt, że Gram-ujemne bakterie wodne wytwarzają
lipopolisachardy ściany komórkowej mogące działać toksycznie - tak jak endotoksyny
bakterii chorobotwórczych. Z tego powodu ich liczba powinna być także monitorowana.
Ponadnormatywny wzrost ich liczebności świadczyć może między innymi, o obecności
w wodzie łatwo przyswajalnych związków organicznych. Teoretycznie, obecność w wodzie
0,1 mg węgla organicznego może spowodować wzrost liczby bakterii w 1 ml do 108 jtk
jednostek tworzących kolonie - (ang. cfu). Również fosfor jest czynnikiem stymulującym
wzrost drobnoustrojów. Dodanie niewielkiej ilości tego pierwiastka (<50mg/l) powoduje
nawet 10-krotne przyspieszanie rozwoju bakterii w wodociągach.
Ogólna liczebność bakterii mezofilnych
Ze względów zdrowotnych, bardziej niebezpieczna jest ponadnormatywna liczba bakterii
rosnących w temperaturze 37oC, ponieważ mogą wśród nich być również bakterie
chorobotwórcze. Duża ich liczba w badanej próbce wody, może świadczyć, między innymi, o źle przebiegających procesach uzdatniania lub zasysaniu zanieczyszczonej wody. Zwiększenie ogólnej liczby bakterii obecnych w próbce wody może świadczyć także
o namnażaniu się drobnoustrojów na wewnętrznych powierzchniach instalacji wodnych, szczególnie na złączach rur i uszczelkach oraz tworzeniu się warstwy tzw. biofilmu.
Przekroczenie dopuszczanego poziomu ogólnej liczby bakterii powinno być zawsze
sygnałem do znalezienia przyczyny zanieczyszczenia i do podjęcia odpowiedniego
postępowania. Niekiedy może istnieć konieczność dodatkowego chlorowania, np. wody do
picia, powyżej 0,2 mg Cl2/l. W niektórych przypadkach dopiero zmiany konstrukcyjne
sieci wodociągowej i usunięcie biofilmu są w stanie skutecznie zabezpieczyć odbiorcę
przed wzrostem liczebności drobnoustrojów w wodzie.
Kryteria jakości sanitarnej wody do picia w Polsce.
Jakość bakteriologiczna wody do picia w Polsce oceniana jest na podstawie liczebności dwóch grup bakterii wskaźnikowych: Escherichia coli i enterokoków (tabela 1). Dodatkowo dokonuje się analizy bakterii z grupy coli oraz klostridiów redukujących siarczyny (tabela 2).Według stosowanych w Polsce kryteriów w 100 ml wody podawanej do sieci wodociągowej nie może być ani jednej komórki bakterii uznanych za wskaźnikowe. Podobne normy jakości wody obowiązują w Unii Europejskiej. Dodatkowym ważnym kryterium jakości wody do picia jest także ogólna liczebność bakterii psychrofilnych i mezofilnych w 1 ml wody. Według stosowanych w Polsce kryteriów w wodzie do spożycia przez ludzi liczebność bakterii psychrofilnych nie powinna przekraczać 100 komórek w 1ml, natomiast bakterii mezofilnych 50 komórek w 1 ml wody (tabela 2).
Osobne wymagania dotyczą wód wprowadzanych do jednostkowych opakowań (tabela 3) oraz wód w cysternach, zbiornikach magazynujących wodę w środkach transportu lądowego, powietrznego lub wodnego (tabela 4). Jakość wody ciepłej ocenia się na podstawie liczebności bakterii z rodzaju Legionella (tabela 5).
Mikroflora autochtoniczna
Mikroflorę gleby dzieli się na autochtoniczną i zymogeniczną. Drobnoustroje autochtoniczne czerpią energię z rozkładu materii organicznej powodując jej mineralizację. Przedstawicielami tej grupy w glebie są najczęściej drobnoustroje, dla których źródło węgla i energii stanowią substancje humusowe, tj. związki powstałe z rozkładu i przetworzenia materii organicznej pochodzenia roślinnego i zwierzęcego. Przedstawicielami tej grupy są najczęściej tlenowe nieprzetrwalnikujące bakterie z rodzaju Arthrobacter oraz prątki z rodzaju Mycobacterium. Stałymi mieszkańcami gleby są także promieniowce z rodzaju Streptomyces i Nocardia oraz bakterie śluzowe (Myxobacteriales). Dla gleby, choć nie tylko dla niej, charakterystyczne są bakterie wiążące azot atmosferyczny, bakterie nitryfikacyjne. Pospolite są także tlenowe laseczki przetrwalnikujące z rodzaju Bacillus i beztlenowe z rodzaju Clostridium.
Mikroflora zymogeniczna
Drobnoustroje zymogeniczne wprowadzane są do gleb okresowo. Pochodzą one z wprowadzanych do gleb ścieków, odpadów komunalnych i przemysłowych oraz z odpadów powstających w hodowli zwierząt (np.obornik i gnojówka), Są to organizmy o dużych wymaganiach odżywczych, których rozwój uzależniony jest od dopływu świeżej, łatwo przyswajalnej materii organicznej. Bakterie zymogeniczne bytujące w glebie to głównie gramujemne pałeczki z rodziny Enterobacteriaceae oraz z rodzaju Pseudomonas, Achromobacter, Flavobacterium.
W glebie zanieczyszczonej odchodami czy szczątkami zwierząt i roślin mogą występować drobnoustroje chorobotwórcze (patogenne). Spośród bakterii szczególnie niebezpieczne dla człowieka są pałeczki duru brzusznego z rodzaju Salmonella i pałeczki czerwonki - Shigella., beztlenowe laseczki wywołujące tężec - Clostridium tetani, zgorzel gazową - Clostridium perfringens, zatrucia pokarmowe - Clostridium botulinum oraz tlenowe laseczki wąglika - Bacillus anthracis, a także prątki - Mycobacterium tuberculosis.
Ponadto do bakterii chorobotwórczych występujących w glebie zalicza się niektóre promieniowce wywołujące tzw. promienicę. Grzyby są przyczyną zakażeń skóry i tkanek wewnętrznych, a organizmy zwierzęce jak pierwotniaki i robaki - chorób głównie przewodu pokarmowego. Szczególną uwagę należy zwrócić na możliwość występowania w glebie jaj robaków pasożytniczych np. glisty ludzkiej. Jej jaja przechodzą w glebie okres dojrzewania i wykazują dużą odporność na działanie czynników zewnętrznych. Podobnie jak endospory bakterii, zarodniki grzybów i cysty pierwotniaków, mogą przez długi okres czasu bytować w glebie i stanowić zagrożenie dla człowieka.
Przeżywalność mikroorganizmów zależy od czynników fizycznych i chemicznych:
Woda
● Wszystkie drobnoustroje wymagają dla prawidłowego rozwoju środowiska
zawierającego wodę.
● Woda umożliwia migracje mikroorganizmów w gruncie, dyfuzję substratu i związków odżywczych do wnętrza komórki oraz usunięcie z niej produktów metabolizmu. Wpływa jednocześnie na utrzymanie w komórce odpowiedniego ciśnienia osmotycznego i odczynu.
● Zarówno zbyt duże zagęszczenie składników odżywczych, jak i nadmierne uwodnienie hamuje lub w ogóle uniemożliwia wzrost mikroorganizmów.
● Nadmiar wody w glebie obniża dyfuzję tlenu i azotu oraz sprzyja rozwojowi drapieżników żywiących się bakteriami. Zbyt mała ilość wody może uniemożliwić
drapieżnym pierwotniakom przemieszczanie się, a przez to sprzyjać rozwojowi
populacji bakterii.
Ciśnienie osmotyczne
● Na rozwój drobnoustrojów duży wpływ wywiera ciśnienie osmotyczne roztworu
glebowego związane ściśle z nawilgoceniem gleby, zwiększające się w miarę jej
przesychania.
● W glebach średnio wilgotnych niezasobnych ciśnienie roztworu waha się w granicach
0,5 -5 atm. W glebach zasolonych może dochodzić do 100 atm. W komórkach
drobnoustrojów wynosi ono 3-6 atm.
● Ciśnienie osmotyczne wyższe w roztworze glebowym niż w komórkach, zakłóca
proces wchłaniania wody przez komórki drobnoustrojów, co wpływa na zahamowanie
ich wzrostu.
Potencjał oksydacyjno-redukcyjny
● Potencjał oksydacyjno-redukcyjny odzwierciedla tendencję substancji do pozyskiwania lub utraty elektronów. Ma on istotne znaczenie dla przebiegu zjawisk chemicznych i biologicznych w glebie.
● W roztworach glebowych rozpuszczają się sole pierwiastków o zmiennej wartościowości np. Fe, Mn, S. Od stosunku utlenionych i zredukowanych związków tych pierwiastków (Fe3+/Fe2+, MnO2/Mn2+, SO42-/S2-), a także od zaopatrzenia gleby w tlen, zależą procesy tlenowe i beztlenowe w glebie.
● Pierwiastki w formie utlenionej lub zredukowanej tworzą układy red-ox, od których z kolei zależy kierunek i charakter przemian metabolicznych.
● Na skutek procesów dysocjacji, woda gleby ma wpływ na wartość potencjału red-ox, co z kolei oddziałuje wybiórczo na rozwój i skład drobnoustrojów glebowych.
● Przesuszenie gleby i związane z tym lepsze napowietrzenie, wzmaga procesy utleniania i mineralizacji.
● Przeciwnie, nadmierne jej nawilgocenie powoduje brak tlenu, przez co środowisko opanowują mikroaerofile i beztlenowce; obniżają one potencjał re-dox gleby, co stymuluje procesy redukcji i fermentacji.
pH gleby
● Przebieg procesów mikrobiologicznych w glebie zależy w dużej mierze od jej odczynu, warunkuje on, bowiem aktywność enzymów oraz procesy transportu.
● Roztwór glebowy odznacza się właściwościami buforowymi, czyli zdolnością przeciwstawiania się zmianom odczynu; jednakże w ograniczonym zakresie.
● Odczyn gleby wpływa na rozpuszczalność i przyswajalność składników pokarmowych. Żelazo i mangan są dostępne tylko w warunkach niskiego odczynu, natomiast molibden w wysokim pH.
● Wartość pH gleby zależy od jej składu chemicznego, ale w czasie procesu biologicznego rozkładu materii organicznej mogą zachodzić zmiany pH wynikające z charakteru metabolizmu i fizjologii drobnoustrojów.
● Kwasowość gleby może się zwiększać pod wpływem np. kwaśnych deszczów, nawożenia, zasiedlania bakteriami utleniającymi siarkę itp., a to wpływa na przebieg szeregu transformacji metabolicznych.
● Jedną z najbardziej wrażliwych na pH reakcji w glebie jest nitryfikacja, czyli przemiana NH4+ do NO3-. Jony te istotnie też wpływają na pH gleby. Pobieranie przez drobnoustroje jonów amonowych (NH4+) ze środowiska przyczynia się do zakwaszenia gleby, natomiast przyswajanie azotanów (NO3-) - do jej alkalizacji. Te z kolei zmiany wpływają na rozpuszczalność innych soli i ich przyswajalność- przez mikroorganizmy.
● Wiele spośród znanych gatunków bakterii może rozwijać się w zakresie pH od 4 do 9. Optymalne dla bakterii warunki wzrostu istnieją przy pH 6,5 do 8,0.
● Drobnoustroje kwasolubne mogą rosnąć w zakresie pH od 1 do 6, ekstremalne acidofile rosną chętnie przy wartościach od 1 do 3. Wśród nich są niektóre gatunki Thiobacillus, Thermophilus i Sulfolobus, utleniające siarczki mineralne do kwasu siarkowego.
● Większość grzybów preferuje kwaśny odczyn środowiska. Grzyby jako grupa są
umiarkowanymi acidofilami (optymalne dla wzrostu pH wynosi od 4 do 6).
● Do umiarkowanych drobnoustrojów zasadolubnych (alkalofilów) należą bakterie z rodzaju Nitrosomonas - optymalne pH ich aktywności waha się od 7,3 do 9,6.
Temperatura
Mikroorganizmy glebowe różnią się termotolerancją; różne są optymalne temperatury
rozwoju. Uwzględniając wrażliwość drobnoustrojów na temperaturę można wśród nich
wyróżnić:
- psychrofile
- mezofile
- termofile
● Dla psychrofili temperatury wzrostu obejmują zakres od minus 5 do +25oC, dla
mezofili 15-45oC, dla termofili 40-70oC.
● Mimo, że pojedyncze gatunki mogą mieć szerszy lub węższy zakres tolerancji na
temperaturę, to jednak większość gatunków należy do mezofili i toleruje temperaturę
około 30oC.
● Organizmy rosnące w niskich temperaturach w pobliżu 0oC zawierają w błonie
komórkowej specyficzne lipidy utrzymujące jej półpłynność. Termofile zaś zawierają
lipidy, których punkt topnienia jest wysoki.
● Nadmierny wzrost temperatury powoduje poważne obniżenie efektywności procesu
biosyntezy na skutek zwiększonego zużycia źródła energii na oddychanie, spadku
wydajności produkcji, powstawania skutków ubocznych. Poniżej 6oC drobnoustroje
ograniczają procesy życiowe przechodząc w stan życia utajonego (anabiozy) bądź
w formy przetrwalne.
● W glebie temperatura na powierzchni może osiągać w południe nawet 70oC i
wykazywać dobową fluktuację ok.50o C. Zmiany temperatury na powierzchni w ciągu
doby nie mają wpływu na temperaturę w głębszych poziomach profilu glebowego.
Natlenienie
● Tlen należy do kluczowych czynników kształtujących warunki rozwoju
drobnoustrojów, decyduje o możliwości występowania lub braku ich wzrostu, wpływa
na szybkość wzrostu i przyrost biomasy oraz na fizjologię komórek, a więc na rodzaj,
wydajność i szybkość produkcji określonych metabolitów.
● Zapotrzebowanie na tlen drobnoustrojów zależy od charakteru ich metabolizmu,
rodzaju i stężenia źródła węgla i energii, fazy rozwoju populacji i stanu fizjologicznego
komórek. Tylko bakterie mogą przeżywać dłuższy okres czasu w warunkach
beztlenowych.
● Tlen zawarty w glebie zużywany jest w 70% przez drobnoustroje, a w 30% przez
korzenie roślin, natomiast procesy chemiczne wykorzystują tylko śladowe ilości tlenu.
● Warunki beztlenowe występują w glebach, w których zawartość tlenu w powietrzu
glebowym jest niższa od 1%.
● Jeżeli niemożliwe jest dostarczenie do gleby tlenu, proces biologicznego rozkładu materii organicznej prowadzony jest przez bakterie beztlenowe, wykorzystujące do procesów utleniania komórkowego tlen pochodzący z takich związków, jak siarczany lub azotany. W warunkach beztlenowych procesy dekompozycji materii organicznej zachodzą wolniej i są mniej wydajne energetycznie.
Zawartość składników pokarmowych
● Do budowy biomasy mikroorganizmów obok związków węgla niezbędne są także
inne składniki pokarmowe takie jak: azot, fosfor, siarka, wapń, magnez czy potas.
Do szczególnie ważnych pierwiastków należą azot i fosfor będące pierwiastkami
niezbędnymi do produkcji białek i kwasów nukleinowych.
● Żyzne gleby zawierają wszystkie niezbędne składniki w odpowiednich ilościach
natomiast w glebach zanieczyszczonych proporcje między poszczególnymi
pierwiastkami są zakłócone. Powszechnie uważa się, że stosunek wagowy węgla do
azotu i fosforu w glebie powinien kształtować się na poziomie 10:1:0,1
● Wapń poprawia właściwości fizyczne i chemiczne gleby oraz jej strukturę.
Związki toksyczne
● Obecność związków toksycznych jest czynnikiem opóźniającym bądź całkowicie
hamującym procesy mikrobiologiczne w glebie.
● Do szczególnie toksycznych związków należą pestycydy, węglowodory alifatyczne
i aromatyczne, formaldehyd, związki chloroorganiczne, metale ciężkie oraz sole
występujące w dużych stężeniach.
Światło
● Światło penetruje zaledwie kilka cm w głąb ziemi.
● Intensywność naświetlenia powierzchni gleby zależy od rodzaju i zagęszczenia
rosnących na niej roślin.
● Światło jest niezbędne jedynie dla przeprowadzających proces fotosyntezy glonów
zasiedlających glebę.
● Naświetlenie ma wpływ na aktywność dżdżownic - przemieszczają się one na powierzchnię gleby w nocy w poszukiwaniu żywności i w celach rozrodczych.
W glebie zanieczyszczonej odchodami czy szczątkami zwierząt i roślin mogą występować drobnoustroje chorobotwórcze (patogenne). Spośród bakterii szczególnie niebezpieczne dla człowieka są:
Ponadto do bakterii chorobotwórczych występujących w glebie zalicza się niektóre promieniowce wywołujące tzw. promienicę. Grzyby są przyczyną zakażeń skóry i tkanek wewnętrznych, a organizmy zwierzęce jak pierwotniaki i robaki - chorób głównie przewodu pokarmowego. Szczególną uwagę należy zwrócić na możliwość występowania w glebie jaj robaków pasożytniczych np. glisty ludzkiej. Jej jaja przechodzą w glebie okres dojrzewania i wykazują dużą odporność na działanie czynników zewnętrznych. Podobnie jak endospory bakterii, zarodniki grzybów i cysty pierwotniaków, mogą przez długi okres czasu bytować w glebie i stanowić zagrożenie dla człowieka.
Zakres analizy mikrobiologicznej gleby pod względem sanitarnym powinien obejmować:
Stopień zanieczyszczenia gleby jajami robaków (helmintów) określa się w odniesieniu do 100 g gleby, uznając glebę za:
Ponadto badania uzupełnić można o oznaczenia:
Określenia stosowane w analizie mikrobiologiczno-helmintologicznej gleby:
Alternatywne metody dezynfekcji:
Wyróżnia się trzy zasadnicze metody oczyszczania ścieków : mechaniczne, chemiczne i biologiczne. W konkretnej oczyszczalni mogą być wykorzystane wszystkie wymienione sposoby lub niektóre z nich. W oczyszczalniach biologicznych zanim ścieki zostaną poddane działaniu mikroorganizmów, są wstępnie oczyszczane na drodze mechanicznej np. na kratach, w osadnikach czy piaskownikach. Istnieją cztery stopnie oczyszczania ścieków :
I stopień - oczyszczanie mechaniczne, polegające na usuwaniu zanieczyszczeń stałych, do zawiesin opadających włącznie,
II stopień - oczyszczanie biologiczne usuwające zanieczyszczenia rozpuszczone,
III stopień - usuwanie związków biogennych (głównie soli azotu i fosforu),
IV stopień - usuwanie resztkowych zanieczyszczeń trudno rozkładalnych, tzw. związków refrakcyjnych (ten stopień oczyszczania określany jest jako odnowa wody).
W procesie biologicznego oczyszczania ścieków zachodzą następujące zjawiska:
● rozkład substancji organicznych do CO2, H2O i NH3 (w zależności od pH),
● nitryfikacja (utlenianie NH3 za pomocą bakterii Nitrosomonas do azotynów),
a następnie za pomocą bakterii Nitrobacter do azotanów,
● denitryfikacja (przemiana azotanów do azotu gazowego N2).
Warunki, aby bakterie miały szansę się rozwijać - odpowiednie:
pH, natlenienie, dostarczenie zw. Biogennych (azot i fosfor)
Metody naturalne:
Bakterie i substancje organiczne zatrzymują się na gruzełkach i oczyszczone ścieki filtrują niżej.
oczyszczanie hydrobotaniczne (fitoremediacja)
w ekosystemach podmokłych, bagiennych
mikroorg. rozkładają materię organiczną
warunki meteorologiczne
stężenie zanieczyszczeń
pH, substancje biogenne
wiek plantacji
skład roślin zastosowanycj
rozkład substancji organicznej
usuwanie azotu
usuwanie fosforu
usuwanie pasożytów, bakterii, wirusów
gruntowo - roślinne - ścieki spływają przez system korzeniowy
zbiorniki płytkie z roślinnością bagienną
zbiorniki z roślinnością pływającą po powierzchni
Sztuczne metody oczyszczania ścieków- wykorzystują procesy samooczyszczania zachodzące w naturze ze zwielokrotnioną intensywnością
Biologiczne oczyszczanie ścieków - osad czynny.
hodowla czynna - doprowadza się ścieki i wylewa
hodowla statyczna - stale doprowadza i odprowadza
o. Wolnoprływające np. pantofelek
o. Pełzające po kłaczkach
o. Osiadłe np. Vorticella opercularia - bardzo dobra praca osadu
nieoskorupione - świadczy o niestabilności os.
oskorupione -> niewiele zw. org, dobre natlenienie, proces nitryfikacji
temperatura: 15-25 st. - optimum
ilość tlenu: (ograniczane przez temp.) natlenienie1-3 mgO2/litr
odczyn 7-7,5 - optimum; 6-9 - dopuszczalne
składniki odżywcze: węgiel, azot, fosfor
związki toksyczne - gł. w ściekach przemysłowych -> dezaktywacja
adaptacja - dozowanie ścieków toksycznych
inokulacja - preparaty
zsedymentowany osad jest zawracany do komory natlenienia
Biologiczne oczyszczanie gruntów
środowisko podlegające bioremediacji powinno zawierać mikroorganizmy u których zachodzą wymagane procesy kataboliczne,
mikroorganizmy wykorzystywane w procesie bioremediacji powinny być zdolne do przetwarzania związków chemicznych w odpowiednim tempie i obniżać ich koncentracje do poziomu nie przekraczającego norm,
metabolity powstające podczas mikrobiologicznego rozkładu nie mogą posiadać właściwości toksycznych, mutagennych czy rakotwórczych,
środowisko podlegające bioremediacji nie może zawierać związków chemicznych działających toksycznie i inhibitująco na drobnoustroje przeprowadzające proces biodegradacji,
dany związek bądź związki, które mają być usunięte w procesie bioremediacji muszą być dostępne dla mikroorganizmów a więc biodegradowalne,
warunki w miejscu przeprowadzania procesu lub w bioreaktorze powinny sprzyjać rozwojowi i działalności drobnoustrojów np. dostateczna ilość składników pokarmowych, tlen lub inny elektronobiorca, sprzyjająca wilgotność, odpowiednia temperatura oraz dodatkowe źródło węgla w sytuacji, gdy biodegradacja ma zachodzić w oparciu o zjawisko kometabolizmu,
koszt technologii powinien być tańszy lub porównywalny do innych technologii dotychczas opracowanych i stosowanych.
inokulacji gruntu mikroorganizmami aktywnie rozkładającymi zanieczyszczenia
zabiegi przewietrzania gleby strumieniem powietrza,
przemywania roztworem zawierającym substancje odżywcze czy kultury aktywnych mikroorganizmów
bioreaktory
laguny stosowane do oczyszczanie zaolejonych ścieków
reaktory półpłynne stosowane dooczyszczania zanieczyszczonej gleby, szlamów i osadów
bakteryjne
enzymatyczne
bakteryjno-enzymatyczne
Klasyczna metoda identyfikacji mikroorganizmów
Taksonomia sztuczna: - porządkuje mikroorganizmy na podstawie podobieństwa cech fenotypowych (morfologii, biochemii, serologii, fizjologii)
Taksonomia numeryczna - jest zobiektywizowaną formą taksonomii sztucznej i opiera się na zasadzie Adansona .
W klasyfikacji tej bierze się pod uwagę dużą liczbę cech (ponad 100) i formułuje je alternatywnie (+, -)
Każdą cechę szczepu porównuje się z taką samą cechą innego szczepu
Oblicza się współczynnik podobieństwa S
Gatunek znany: Salmonella enterica
Gatunek nieokreślony: Salmonella sp.
Wszystkie (różne) gatunki danego rodzaju: Salmonella spp.
Odmiany (varietas): pathovar, phagovar, serovar Salmonella enterica sv. Typhi (Salmonella Typhi)
Szczep typowy, wzorcowy lub referencyjny
wykazują mniej niż 3% różnic w sekwencji nukleotydowej 16S rRNA
wykazują mniej niż 3% różnic w udziale zasad G +C w genomowym DNA
charakteryzują się co najmniej 70% podobieństwem sekwencji DNA, mierzonym poziomem hybrydyzacji DNA/DNA
Różnią się od innych gatunków zespołem istotnych cech fenotypowych
rośliny przyswajają produkty rozkładu i tworzą swoją biomasę (trzcina, pałka)
Dobór roślin - stosunkowo szybko rosnące, dające dużą biomasę, ma zdolność
akumulacji, mało podatna na działanie szkodników.
Od czego zależy takie oczyszczanie:
Zadania oczyszczania hydrobotanicznego:
(proc. sanitacji ścieków)
3 rodzaje oczyszczania hydrobotanicznego:
Złoża biologiczne
Oczyszczanie ścieków na złożach biologicznych odbywa się w zbiornikach wypełnionych
luźno usypanym materiałem ziarnistym i porowatym). Ścieki za pomocą zraszaczy są rozpryskiwane na górną powierzchnię złoża i spływają następnie przez wypełniający go materiał. Na materiale stałym, z którego zbudowane jest złoże wytwarza się błona biologiczna stanowiąca śluzowatą warstewkę złożoną z mikroorganizmów takich jak: bakterie (głównie bakterie tlenowe), pierwotniaki, grzyby. Złoże stałe to gruboziarnisty materiał porowaty: tłuczeń, koks, żużel wielkopiecowy, tuf wulkaniczny, tworzywa sztuczne i inne materiały odporne na wpływy atmosferyczne. Biomasa, która bierze udział w procesie rozkładu związków organicznych jest biomasą osiadłą na podtrzymującym materiale porowatym. Praca złoża polega na stałym doprowadzaniu ścieków i ich przepływie przez złoże w kontakcie z błoną biologiczną, podczas którego zachodzi mineralizacja zanieczyszczeń
na skutek tlenowego rozkładu przez mikroorganizmy i odpływ ze złoża oczyszczonego ścieku. Błona biologiczna jest początkowo utworzona z bakterii zooglealnych produkujących śluzowate otoczki. Z czasem skład gatunkowy błony zmienia się w wyniku sukcesji. Obok bakterii pojawiają się grzyby, pierwotniaki, wrotki, pierścienice i larwy much. Procesy życiowe mikroorganizmów są uzależnione od dopływu tlenu, a skuteczność rozkładu biologicznego w błonie zależy od szybkości odprowadzania, ditlenku węgla jako produktu rozkładu.
Realizacja sprawnego procesu polega na:
● zapewnieniu kontaktu ścieków z błoną biologiczną,
● zapewnieniu odpowiedniego napowietrzenia.
W zależności od obciążenia ładunkiem organicznym wyróżniamy następujące rodzaje złóż
biologicznych:
● Niskoobciążone (nitryfikacyjne) - mogą być wypełnione materiałem naturalnym lub sztucznym. Ładunek organiczny doprowadzany na złoże jest mniejszy od 0,4 kg BZT5/m3·d. W złożach zraszanych błona jest dobrze rozwinięta i proces biologicznego rozkładu jest prawie zupełny. W końcowej fazie oczyszczania zachodzą intensywne procesy nitryfikacji, które powodują wzrost zawartości azotanów w odpływie kierowanym do osadników wtórnych (osadnik wtórny jest to urządzenie wykorzystujące proces sedymentacji, w celu oddzielenia osadu czynnego od ścieków oczyszczonych).
● Średnioobciążone - wypełnione materiałem naturalno-syntetycznym, pracują przy obciążeniu objętości ładunkiem w granicach 0,4-0,65 kg BZT5/m3·d. Często stosowana jest w tych złożach recyrkulacja części ścieków oczyszczonych w celu zapewnienia odpowiedniej intensywności zraszania złoża i zapewnienia odpowiedniego stężenia ścieków doprowadzanych. Redukcja związków organicznych na tych złożach jest zadawalająca, a procesy nitryfikacji przebiegają częściowo. Nie jest konieczne wprowadzanie dodatkowych procesów oczyszczania.
● Wysokoobciążone (spłukiwane) - są wypełnione materiałem naturalnosyntetycznym, obciążenie objętości złoża ładunkiem wynosi: 0,65-1,6 kg BZT5/m3·d. W złożach spłukiwanych intensywność przepływu ścieków jest większa, lecz słabiej rozwija się błona biologiczna złożona tu prawie wyłącznie z bakterii. Przepływająca ciecz wypłukuje ze złoża zużyty i martwy materiał biologiczny, wypłukiwany materiał unoszony jest w postaci kłaczkowatego osadu. Na tego typu złożach, zachodzi jedynie częściowa mineralizacja związków organicznych a proces nitryfikacji jest tu hamowany. O częściowej mineralizacji związków organicznych świadczy niska zawartość azotanów w odpływie ze złóż. W systemach złożonych, po tego rodzaju złożach, stosuje się
Osad czynny - jest także jedną z metod sztucznych opirea się na metodzie naturalnej (ulepszone stawy ściekowe
Osad czynny jest metodą oczyszczania, w której elementem oczyszczającym są bakterie zlepione w kłaczki zawieszone w środowisku płynnym (ścieki). Kłaczki osadu powstają podobnie jak błona biologiczna, w wyniku łączenia się bakterii zooglealnych wytwarzających lepki śluz. Proces tworzenia się kłaczków, zwany flokulacją, zachodzi spontanicznie w trakcie napowietrzania komory wypełnionej ściekami.
W oczyszczalniach pracujących metodą osadu czynnego ścieki, po oczyszczaniu mechanicznym w osadniku wstępnym, są kierowane do komory osadu czynnego, zwanej też komorą napowietrzania, gdzie odbywa się właściwe oczyszczanie biologiczne. Kłaczki osadu wchodzą tu w kontakt ze ściekami w warunkach sztucznego napowietrzania i intensywnego mieszania. Bakterie obecne w kłaczkach pochłaniają substancję organiczną ze ścieków i część jej zużywają jako paliwo (źródło energii), część natomiast przyswajają jako materię bu dulcową, co prowadzi do zwiększenia biomasy. Efektem tych procesów jest mineralizacja ścieków i wzrost masy osadu czynnego.
Wykorzystanie zanieczyszczeń przez bakterie osadu czynnego.
Po kilku godzinach przetrzymywania ścieków w komorze, są one przepuszczane do osadnika wtórnego, w którym zachodzi oddzielenie kłaczków od oczyszczonych ścieków. Kłaczki osadzają się na dnie osadnika i są recyrkulowane albo z powrotem do komory napowietrzania, by utrzymać stężenie biomasy (osad powrotny), albo do osadnika wstępnego i wraz z osadem wstępnym usuwane z obiegu oraz unieszkodliwiane (osad nadmierny). Układ technologiczny procesu osadu czynnego przedstawia rysunek:
Układ oczyszczalni ścieków z osadem czynnym:
1 - osadnik wstępny, 2 - komora osadu czynnego, 3 - osadnik wtórny, 4 - stacja pomp osadowych, Qść - ścieki, Qn - osad nadmierny, Qr - osad powrotny
Jako, że elementem oczyszczającym w tej metodzie są kłaczki zlepionych bakterii, dlatego ważne jest, aby proces flokulacji przebiegał prawidłowo i aby powstałe kłaczki miały odpowiednią wielkość, kształt i strukturę. Flokulacja zależy od stopnia obciążenia osadu ładunkiem zanieczyszczeń. Przy bardzo wysokim i bardzo niskim obciążeniu obserwuje się bowiem zjawisko deflokulacji, czyli rozpraszania się bakterii skupionych w kłaczki. Gdy osad jest nadmiernie obciążony, kłaczki są nie dotlenione, natomiast w przypadku niedociążenia bakterie giną z braku pokarmu.
Wielkość kłaczków jest istotną cechą wpływającą na efektywność oczyszczania. Zależą od niej dwa ważne procesy: biosorpcja (pochłanianie zanieczyszczeń ) i sedymentacja (opadanie kłaczków). Biosorpcja jest etapem poprzedzającym biodegradację i zależy od wielkości powierzchni kłaczków. Powierzchnia ta jest tym większa, im kłaczki są mniejsze. Jednak zbyt małe kłaczki, choć mają dużą powierzchnię sorpcyjną, źle opadają w osadniku wtórnym. Może to prowadzić do wtórnego zanieczyszczenia odbiornika, do którego odprowadza się oczyszczone, ale nie oddzielone od kłaczków ścieki. Natomiast kłaczki zbyt duże, choć świetnie sedymentują, to jednocześnie słabo oczyszczają ścieki. Wynika to z faktu, że bakterie umieszczone w centralnej części dużego kłaczka mają utrudniony dostęp do tlenu i zanieczyszczeń będących substratem pokarmowym. Kłaczki muszą więc wykazywać właściwy stan rozdrobnienia zapewniający odpowiednie warunki tlenowe i pokarmowe.
Kształt kłaczków jest cechą, która może wskazywać na pewne niekorzystne zjawiska w osadzie. Np. kształt gwiaździsty, pierzasty czy siatkowaty wiążą się zwykle z obecnością mikroorganizmów nitkowatych (głównie bakterii i grzybów), których obecność w osadzie jest niepożądana, gdyż powodują jego puchnięcie.
Struktura kłaczka może być spoista bądź luźna. W warunkach niedotlenienia lub braku pokarmu kłaczki przybierają formę zbitych, obłonionych tworów koloru szarego. Obecność takich kłaczków w osadzie powoduje spadek jego aktywności. Wiele różnych czynników powoduje rozluźnienie kłaczków. Należą do nich: rozwój form nitkowatych, nadmiar lub niedostatek tlenu, przeciążenie osadu lub warunki głodowe.
Niekorzystnym zjawiskiem obniżającym zdolności sedymentacyjne kłaczków jest puchnięcie osadu czynnego. Z puchnięciem mamy do czynienia, gdy zwiększa się objętość osadu przy zachowaniu tej samej masy. Liczbowo wyraża to indeks osadu, który przedstawia objętość (w cm3) jednego grama jego suchej masy. Indeks wyraża więc gęstość osadu, lub stopień uwodnienia. Jeden gram suchej masy dobrze pracującego osadu czynnego ma zwykle objętość 67 cm3 (indeks = 67 cm3/g). Indeks osadu spuchniętego mieści się w granicach od 100 - 400 cm3/g. Spuchnięty osad jest co prawda bardzo aktywny (duża powierzchnia biosorpcyjna), jednak z powodu obniżonej opadalności nie może być on oddzielony od oczyszczonych ścieków w osadniku wtórnym . Wiele jest przyczyn puchnięcia osadu. Bezpośrednią przyczyną tego zjawiska mogą być organizmy nitkowate, bakterie zooglealne, rozproszenie kłaczków lub wzrost lekkości osadu spowodowany wydzielaniem się pęcherzyków gazu. Organizmy nitkowate (głównie bakterie i grzyby) wywołują tzw. puchnięcie włókniste. Zwykle dochodzi do niego, gdy skład ścieków jest niewłaściwy (zbyt duże ilości węglowodanów w stosunku do azotu i fosforu), w sytuacji przeciążenia osadu, nagłych zmian obciążenia , niedotlenienia, zakwaszenia, zatrucia, obniżenia temperatury lub niedostatecznie długiego przetrzymywania osadu w komorze napowietrzania. Do najczęściej spotykanych mikroorganizmów nitkowatych należą bakterie: Sphaerotilus, Beggiatoa i promieniowce. Mniej poznanym zjawiskiem jest puchnięcie niewłókniste. Wywołane jest ono nadmierną produkcją pozakomórkowych substancji śluzowych przez pewne bakterie zooglealne. Śluzy te wiążą bardzo dużo wody, co sprawia, że osad jest czterokrotnie silniej uwodniony niż w przypadku spuchnięcia włóknistego. Zwiększenie indeksu osadu może też spowodować rozproszenie kłaczków w wyniku zbyt silnej turbulencji w komorze lub też wynoszenie osadu w osadniku wtórnym. Z tym drugim przypadkiem mamy do czynienia, gdy osad jest zbyt długo przetrzymywany w osadniku i dochodzi do rozwoju beztlenowych bakterii denitryfikacyjnych wydzielających pęcherzyki azotu cząsteczkowego.
Bakterie tworzące kłaczki nie są jedynymi organizmami występującymi w osadzie czynnym. Oprócz bakterii występują tu liczne pierwotniaki i wrotki, rzadziej robaki, pierścienice i grzyby. Biocenoza osadu czynnego ma charakter heterotroficzny.
Piramida przedstawiająca zależności troficzne w osadzie czynnym.
Bazą pokarmową całego zespołu organizmów jest materia organiczna dopływająca wraz ze ściekami. Pierwszy poziom troficzny tworzą bakterie heterotroficzne, grzyby i pierwotniaki saprotroficzne (korzenionóżki i wiciowce), które odżywiają się materią organiczną w postaci roztworów i zawiesin. Drugi poziom stanowią pierwotniaki holozoiczne (gr.holos - cały) odżywiające się całymi mikroorganizmami. Należą tu orzęski. Wreszcie, najwyższy poziom troficzny zajmują orzęski drapieżne (zwłaszcza osiadłe gatunki z grupy Suctoria, które zjadają pierwotniaki bakteriożerne) , a także bezkręgowce takie, jak wrotki, robaki z grupy nicieni i in. Organizmy obecne we wpracowanym osadzie czynnym tworzą zespół, który powstał w wyniku procesu sukcesji. Początkowo w ściekach obecne są głównie bakterie, wiciowce (Zooflagellata) i korzenionóżki (ameby) (Rhizopoda), a więc organizmy odżywiające się materią organiczną zawartą w ściekach. Z czasem pojawiają się orzęski (Ciliata) zjadające bakterie i drapieżne. Z orzęsków najwcześniej rozwijają się formy wolno pływające, pożerające bakterie, które nie zlepiły się jeszcze w kłaczki (nie sflokulowane). W następnym etapie, gdy proces flokulacji jest już zaawansowany, pojawiają się formy osiadłe orzęsków (przyczepione do kłaczków). W ostatnim etapie rozwijają się wrotki (Rotatoria). Opisane przemiany ilustruje wykres:
Sukcesja mikroorganizmów podczas hodowli osadu czynnego
Znajomość przemian sukcesyjnych w osadzie oraz właściwości biologicznych mikroorganizmów jest przydatna w ocenie pracy oczyszczalni. Służy do tego analiza mikroskopowa próbki osadu, obejmująca, oprócz opisu wyglądu kłaczków, również badanie składu mikrofauny. Stwierdzenie dużej liczebności wiciowców i ameb (korzenionóżek), a więc pierwotniaków pionierskich, świadczy o przeciążeniu osadu i jego niedotlenieniu (a więc o warunkach przypominających początkowe etapy rozwoju osadu). Występowanie orzęsków natomiast, zwykle świadczy o dobrej pracy osadu. Dotyczy to zwłaszcza form osiadłych. Orzęski bakteriożerne wywierają stałą presję na populacje bakteryjne, które pobudzane są przez to do ciągłych podziałów. Wynikiem tego jest odmładzanie się flory bakteryjnej i jej zwiększona aktywność metaboliczna. Poza tym orzęski przyczyniają się do klarowania ścieków zjadając nie sflokulowane bakterie. Obecność w osadzie wrotków zwykle świadczy o dobrym natlenieniu osadu i jego stabilizacji
Na powierzchni bakteri znajdują się enzymy hydrolityczne, dzięki którym wiążą się z substratami. Bakterie mają na zewnątrz ładunek ujemny, więc łączą się wszystkie substancje o ładunku dodatnim oraz dodatkowo przyklejają się do śluzu. Powstają konglomeraty - kłaczki (FLOKULACJA - kłaczkowanie).
Najlepiej flokulują te bakterie, które wydzielają duże ilości śluzu - Zooglea ramigera; orzęski, wrotki - przez otwór gębowy wydzielają saubstancje mające charakter zlepiający.
Sposób w jaki kłaczkuje dany osad decyduje o prawidłowości prowadzenia procesu.
Ważny jest stosunek objętości do masy kłaczka.
Deflokulacja - całkowity rozpad kłaczków, za duża ilość ścieków -> całkowite zmętnienie
za mała -> autoliza
Biocenoza
W ściekach gospodarczo-bytowych: Zooglea, Pseudomonas, Achromobacter, Flavobacterium - HETEROTROFY; AUTOTROFY: nitryfikatory, siarkowe
GRZYBY: puchnięcie osadu - wzrost objętości w stosunku do masy
PIERWOTNIAKI: wiciowce, orzęski, korzenionóżki - uczestniczą we flokulacji, wskaźnik dobrej pracy osadu, część oczyszcza, większość odżywia się bakteriami - klarują osad
wiciowce - 30 000 w 1 ml, skaźnik złej pracy osadu, niedobór tlenu, ścieki
zagniwające
orzęski - dobra praca osadu, odżywiają się bakteriami, glonami, zw. org; 90
gatunków; do 10 000/ml
- korzenionóżki - nibynóżki otaczają pokarm, czasami mają skorupę zewnętrzną;
do 50 000/ 1ml; duża ilość świadczy o niestabilności osadu
zachodzi dobrze
ZWIERZĘTA BEZKRĘGOWE
- wrotki - osad dobrze pracuje, stabilny, dobrze natleniony; od 90% BZT;
odżywiają się bakteriami i cz. Osadu, wydalają dość duże ilości śluzu
- nicienie - max 130 w 1 ml; zjadają biomasę osadu czynnego, dotleniają kłaczka,
zwiększają powierzchnię sorpcyjną, rozdrabniają osad, jak jest ich za
dużo mogą zjeść osad, sprzyjają flokulacji.
Parametry fizyko-chemiczne decydujące o pracy osadu:
N:BZT5 - 1:17
P:BZT5 - 1:90
Uzupełnia się np. wodą amoniakalną
enzymu, zmniejszony poziom tlenu. Przyrost biomasy
Bioremediacja to proces oczyszczania gruntów, w którym wykorzystywane są mikroorganizmy- rozkładają one substancje toksyczne do mniej toksycznych/całkiem nie toksycznych.
Kryteria decydujące o możliwości zastosowania bioremediacji do neutralizacji zanieczyszczeń:
Przeprowadzanie procesów oczyszczania gruntów:
- in situ likwidacja zanieczyszczeń bezpośrednio w miejscu ich występowania
- ex situ zanieczyszczony grunt wydobywa się i dopiero wówczas poddaje właściwym zabiegom regeneracyjnym (stosowane w przypadku, gdy gleba skażona jest szczególnie niebezpiecznymi substancjami)
Metody in situ
biostymulacji procesu biodegradacji ropopochodnych dokonuje się poprzez
-wzbogacanie gruntów w pierwiastki biogenne- nawożenie P i N, ewentualnie K (optymalny stosunek C:N:P 100:10:1)
-korektę odczynu- odkwaszanie poprzez wapnowanie
-napowietrzanie- tworzenie warunków tlenowych
-uprawienia roślin wspomagających oczyszczanie- trawy, rośliny motylkowe
głębiej zalegające zanieczyszczenia ww metody plus
Bioekstrakcja wymuszona infiltracji wody gruntowej (bioremediacja stymulowana wodą) z napowietrzaniem (biowentylacja) daje zwiększenie desorpcji zanieczyszczeń ropopochodnych.
Bioremediacja stymulowana wodą in situ ma na celu wymuszenie pionowego, a następnie
poziomego przepływu wody wraz z substancjami ropopochodnymi w środowisku
gruntowo-wodnym może być wspomagana obecnością
substancji powierzchniowoczynnych w środowisku gruntowo-wodnym.
Badania wykazały, że surfaktanty syntetyczne oraz biosurfaktanty mogą
przyspieszać proces biodegradacji produktów naftowych, w szczególności ciężkich
frakcji ropy naftowej.
Biowentylacja stosowana jako samodzielna technika oczyszczania prowadzona w celu maksymalizacji ulatniania się związków węglowodorowych o małej masie cząsteczkowej (np. produktów na bazie benzyny lekkiej lub rozpuszczalników)
Napowietrzanie prowadzi się w sposób bierny lub aktywny. W przypadku wentylacji biernej napowietrzanie odbywa się poprzez system rur perforowanych w sposób samoistny. Wentylacja aktywna polega na wytworzeniu podciśnienia (ekstrakcja powietrza gruntowego) lub nadciśnienia (iniekcja powietrza).
Metody ex situ
stosowane gdy istnieje niebezpieczeństwo migracji toksycznych zanieczyszczeń do wód gruntowych oraz proces detoksykacji musi być przeprowadzony w krótkim czasie.
Metoda bioreaktorowa
metoda kosztowna, przeprowadzana pod pełną i ciągłą kontrolą stosowana dla mniejszych ilości gleby
technologii opartych o metody ex situ:
Mikroorganizmy wykorzystywane w procesie bioremediacji
Proces bioremediacji może być prowadzony w oparciu o mikroflorę autochtoniczną,
naturalnie zasiedlającą skażony grunt lub w oparciu o szczepy pochodzące z innych
środowisk bądź kolekcji odznaczające się wysoką aktywnością degradacyjną. Konieczne jest stosowanie mieszanej kultury mikroorganizmów o zróżnicowanych możliwościach metabolicznych
Dobór szczepów:
● Warunkiem adaptacji inokulantów w gruncie jest brak antagonistycznych
oddziaływań z naturalną mikroflorą gleby.
● Biopreparaty powinny być całkowicie bezpieczne dla człowieka i środowiska. Oznacza
to, że nie mogą to być organizmy patogenne. Poza tym ważne jest, aby metabolity
powstające w procesie rozkładu ksenobiotyków nie posiadały właściwości toksycznych,
mutagennych czy rakotwórczych
Szczepy zawarte w biopreparatach są przechowywane w formie liofilizatów, zamrażane bądź umieszczane w specjalnej zawiesinie podział preparatów ze względu na skład:
Zaszczepienie gruntu poprzedza namnożenie mikroorganizmów w specjalnie przystosowanych bioreaktorach
Bioaugmentacja
Wzbogacenie zanieczyszczonego terenu w specjalnie wyselekcjonowane, o dużej zdolności biodegradacji zanieczyszczeń, bakterie stosuje się, gdy rodzima populacja bakterii na skażonym terenie nie występuje.
Taksonomia (gr. taxis - porządek) zajmuje się opisem organizmów, ich nazywaniem i klasyfikacją (porządkowaniem)
Taksonomia jest poddyscypliną systematyki - szerszej dyscypliny badającej różnorodność organizmów i związki pokrewieństwa między nimi
Taksonomia dzieli się na:
- klasyfikację,
- nomenklaturę (nazewnictwo)
- identyfikację
Rodzaje taksonomii (klasyfikacji) pożądanej aktywności w kierunku biodegradacji zanieczyszczeń.
Taksonomia numeryczna opiera się na założeniu, że wszystkie uchwytne cechy organizmu są w klasyfikacji tak samo ważne (zasada Adansona)
S = (a + d) / (a +b + c + d)
(a, d) - sumy cech wspólnych porównywanych szczepów,
(b, c) - sumy cech różnych
S = 1 oznacza 100% podobieństwo
S < 0,02 oznacza brak podobieństwa
Badane szczepy można uznać za należące do jednego gatunku, gdy ich S > 0,8.
Taksonomia naturalna (filogenetyczna) - porządkuje mikroorganizmy na podstawie pokrewieństwa (wspólnego pochodzenia). Taksonomia naturalna- Celem taksonomii naturalnej (naturalnego systemu klasyfikacji) jest określenie pokrewieństwa między klasyfikowanymi organizmami (taksonomia ma być odbiciem filogenezy).
Cel ten osiągany jest przez analizę kwasów nukleinowych, a szczególnie przez analizę sekwencji 16S rRNA (badania Carla Woese) oraz sekwencji innych konserwatywnych makromolekuł (np. cytochromy)
Zasady nazewnictwa mikroorganizmów
(ang. type strain): np. ATCC 1234T
(American Type Culture Collection + numer identyfikacyjny)
International Journal of Systematic and Evolutionary Microbiology (IJSEM)
45. Metody genetyczne w identyfikacji (taksonomii) mikroorganizmów
Koncepcja gatunku u prokariotów
Gatunek to grupa organizmów, które:
W „Bergey's manual of systematic bacteriology” (wyd. 2005 r.) opisanych jest 7029 gatunków prokariotycznych, z czego 6740 to bakterie, a 289 to archeony. Bakterie podzielone są na 25 typów, z których najlicznijszy jest typ Proteobacteria podzielony na 5 klas (α, β, γ, δ, ε) (grupujący większość bakterii gramujemnych)
32
Wyszukiwarka