
Metabolizm
Metabolizm – ogół procesów fizycznych i chemicznych
związanych z wytwarzaniem energii:
pobieranie substancji z zewnątrz
przekształcanie, rozpad
częściowe przyswajanie
wydalanie substancji zbędnych
Anabolizm – procesy syntezy prowadzące do powstania
związków
organicznych o dużych cząsteczkach z pierwiastków i
związków
drobnocząsteczkowych
Katabolizm – procesy rozpadu związków
wielkocząsteczkowych

Podział drobnoustrojów wg
rodzaju:
źródła węgla: autotrofia; heterotrofia
źródła energii metabolicznej: fototrofia (światło);
chemotrofia (pierwiastki lub zw. chem.)
źródła elektronów: organotrof (zw. org.) litotrof (zw.
nieorg. lub pierwiastki)

Typ
pokarmow
y
Źródło
ener
gii
Źródło
elektr
onów
Źródło
węgla
przykłady
Fotolito(auto)t
rofy
światło
zw.
nieorg
.
CO
2
Sinice
fotoorganohete
rotrofy
światło
zw. org.
zw. org.
Bakterie
niesiarkowe
chemolitoautot
rofy
zw.
nieor
g.
zw.
nieorg
.
CO
2
Bakterie
nitryfikacyjne
chemoorgano
heterotrof
y
zw.
nieor
g.
zw. org.
zw. org.
Patogeny,
grzyby,
pierwotniaki,
zwierzęta,
wiele bakterii
Chemilitoheter
otrofy
zw.
nieor
g.
zw.
nieorg
.
zw. org.
Beggiatoa spp.

Typy procesów
oddechowych
Tlenowe
Beztlenowe:
Fermentacje
Oddychanie z redukcją związków
mineralnych

Przebieg procesów
związanych z pozyskiwaniem
energii
Wstępny rozkład wielocukrów za
pomocą zewnątrzkomórkowych
enzymów hydrolitycznych
Rozkład heksoz do stadium
pirogronianu
Przekształcenie pirogronianu

SZLAKI DEGRADACJI
CUKRÓW do stadium
pirogronianu
- szlak glikolityczny
- cykl pentozofosforanowy (HMP)
- cykl Endnera � Doudoroffa
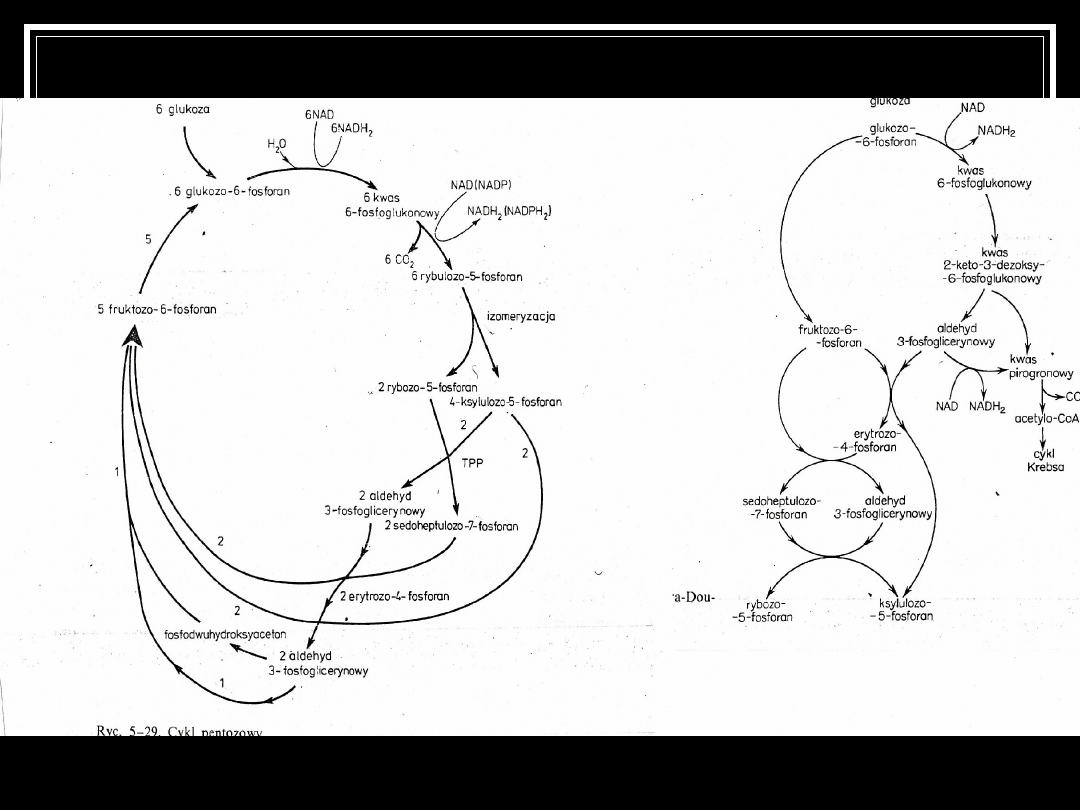

Szlak pentozofosforanowy
- tworzenie różnego rodzaju związków m.in. 6-
fosfofruktozy (substrat do glikolizy)
- produkcja pentoz niezbędna do budowy różnych
związków
- dostarczenie NADPH � niezbędnych potem do różnych
biosyntez
- utlenienie 1 mola glu: 12 NADPH (36 ATP) + 2 aldehydu
- istnienie tego szlaku umożliwia przyswajanie pentoz w
postaci ksylenu przez rośliny

cykl Endnera Doudoroffa
Pseudomonas, Glukonobacter
Szlak podobny do glikolizy. Z 1 glu -> pirogronian
(może wejść do cyklu Krebsa) + aldehyd-3-
fosfoglicerynowy (do cyklu HMP)
Erytrozo-4-fosforan do produkcji tyrozyny, fenyloalaniny
Pirogronian to źródło acetylo-CoA
Szczawiooctan jest źródłem: asparaginianu, tyrozyny,
metioniny

Głównym celem szlaków jest tworzenie energii w
postaci ATP (wysokoenergetyczne wiązania
bezwodnikowe)
ATP tworzone jest głównie w procesie fosforylacji
oksydatywnej w łańcuchu oddechowym
Dawca elektronów i H+ w łańcuchu oddechowym jest
NADH
Głównymi przenośnikami energii są: dehydrogenazy
(przenoszenie H+ i e), reduktazy (przenoszenie tylko
e)

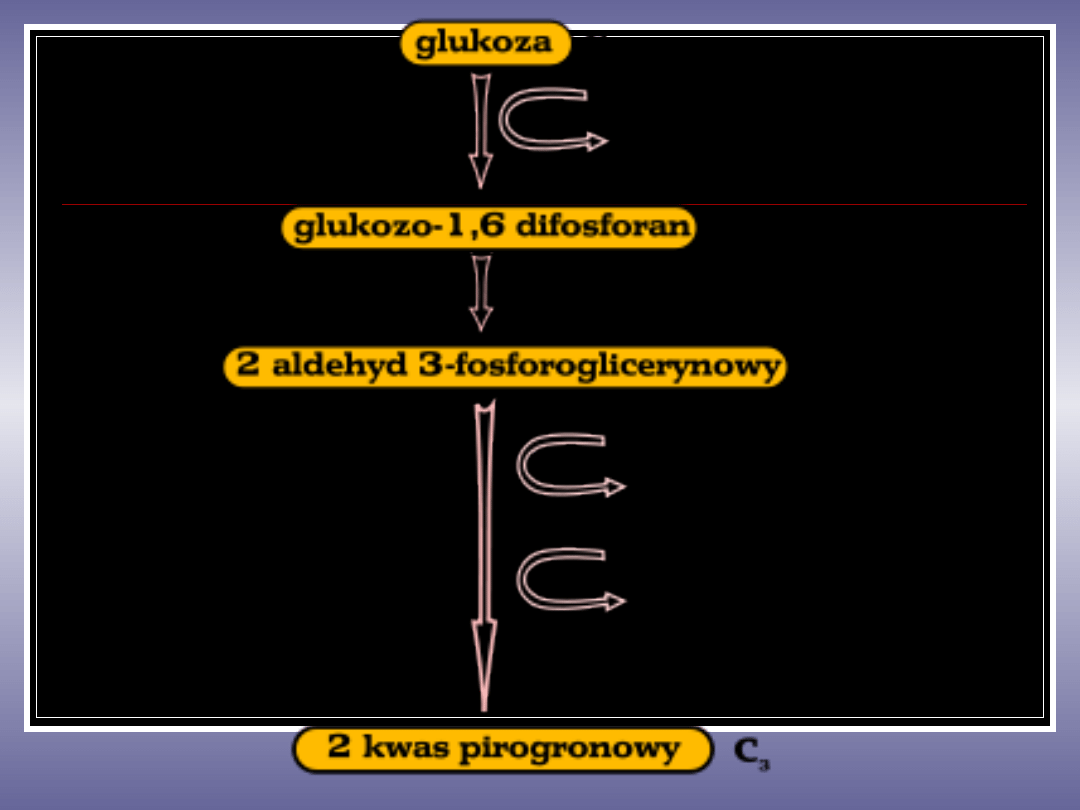
GLIKOLIZA (szlak Embdena – Meyerhofa-
Parnasa)
- proces
rozkładu cukrów do
, którego celem jest pozyskanie
energii pod postacią
.
Substratami dla procesu mogą być:
,
, galaktoza i glicerol. Proces glikolizy może
zachodzić zarówno w warunkach tlenowych, jak i
beztlenowych, uważa się jednak, że glikoliza jest
najstarszym ewolucyjnie procesem pozyskiwania
energii z cukru; prawdopodobnie wykształcił się on
jeszcze wtedy, gdy w atmosferze ziemskiej nie było
tlenu. Enzymy glikolityczne można znaleźć zarówno u
bakterii jak i u eukariotów.

W pierwszym etapie następuje fosforylacja (kosztem
ATP) różnych sacharydów: heksoz, glikogenu, skrobi
i ich rozkład z wytworzeniem aldehydu-3-
fosfoglicerynowego.
W drugim etapie zachodzą reakcje oksydo-
redukcyjne
(z udziałem dinukleotydu nikotynamidoadeninowego
NAD) dostarczające energii, która jest częściowo
magazynowana w cząsteczkach powstającego ATP
oraz następuje wytworzenie kwasu pirogronowego.
Przebieg I i II etapu glikolizy jest identyczny jak w
fermentacji alkoholowej. Powstały kwas
pirogronowy może ulegać różnym przemianom.
W warunkach beztlenowych, np. podczas pracy
mięśni, gdy następuje spadek stężenia tlenu w
tkankach, zachodzi trzeci etap glikolizy: kwas
pirogronowy ulega redukcji (przy udziale NADH) do
kwasu mlekowego. NADH utleniony ponownie do
NAD+ może ponownie brać udział w przemianie
następnej cząstki heksozy w drugim etapie glikolizy.

W trakcie glikolizy
zachodzi:
1. degradacja wielocukrów do heksoz
2. heksoza hydrolizowana przez enzymy wydzielane na zew. a-amylaza,
glukoamylaza
3. w 1-szym etapie szlaku następuje 2-krotna fosforylacja
4. rozpad do zw. 3-weglowych
5. do pirogronianu (2 cząsteczki)
Zysk glikolizy: 2 ATP
6. pirogronian może być substratem szeregu rodzajów fermentacji. W trakcie.
Których dochodzi do regeneracji NADH w wyniku przeniesienia elektronów
na pirogronian. W warunkach tlenowych pirogronian przekształcany do
acetylo-CoA, który wchodzi w cykl Krebsa (gdzie następuje generowanie
ATP, tworzenie NADH, który przeniesiony na łańcuch oddechowy daje zysk
energetyczny)
W glikolizie powstaje szereg produktów wykorzystywanych do syntez
komórkowych
CHO
OH
H
H
HO
OH
H
OH
H
CH
2
OH
O
H
HO
H
HO
H
OH
OH
H
H
OH
+ ATP
+ ATP
CHO
OH
H
H
HO
OH
H
OH
H
CH
2
OPO
3
2-
O
H
HO
H
HO
H
OH
OH
H
H
OPO
3
2-
+ ADP + H
+
+ ADP + H
+
Glukoza
Glukozo-6-fosforan
enzym:
heksokinaza
Etap 1:
fosforylacja
glukozy
Etap 2:
izomeryzacj
a
glukozy do
fruktozy
CH
2
OH
-2
O
3
POH
2
C
HO
HO
O
CH
2
OH
O
H
HO
OH
H
OH
H
CH
2
OPO
3
2-
Fruktozo-6-fosforan
CHO
OH
H
H
HO
OH
H
OH
H
CH
2
OPO
3
2-
O
H
HO
H
HO
H
OH
OH
H
H
OPO
3
2-
Glukozo-6-fosforan
Enzym:
izomeraza
glukozofosforanowa
Etap 3:
Druga
fosforylacja
fruktozy
CH
2
OH
HO
HO
-2
O
3
POH
2
C
CH
2
OH
O
H
HO
OH
H
OH
H
CH
2
OPO
3
2-
Fruktozo-6-fosforan
Enzym:
fosfofruktokinaza
CH
2
OPO
3
2-
-2
O
3
POH
2
C
HO
HO
O
CH
2
OPO
3
2-
O
H
HO
OH
H
OH
H
CH
2
OPO
3
2-
Fruktozo-1,6-difosforan
CH
2
OPO
3
2-
-2
O
3
POH
2
C
HO
HO
O
CH
2
OPO
3
2-
O
H
HO
OH
H
OH
H
CH
2
OPO
3
2-
Fruktozo-1,6-difosforan
Rozszczepienie
wi¹zania miêdzy 3 - 4 atomem wêgla
CH
2
OH
C
H
O
C O
CH
2
OPO
3
2-
C
H
OH
CH
2
OPO
3
2-
fosfodihydroksyaceton
aldehyd 3-fosfoglicerynowy
Etap 4:
Rozpad na
2
fragmenty
trójwęglow
e;

Etap 5:
Fragmenty
trójwęglow
e
izomeryzuj
ą
CH
2
OH
C
H
O
C
O
CH
2
OPO
3
2-
C
H
OH
CH
2
OPO
3
2-
fosfodihydroksyaceton
aldehyd 3-fosfoglicerynowy
Izomeraza
triozofosforanowa
Etap 6:
Odwodornieni
e
i fosforylacja
trójwęglowego
fragmentu
C
H
O
C
H
OH
CH
2
OPO
3
2-
aldehyd 3-fosfoglicerynowy
dehydrogenaza
aldehydu
3-fosfoglicerynowego
NAD
+
+ P
i
NADH + H
+
C
O
OPO
3
2-
C
H
OH
CH
2
OPO
3
2-
1,3-difosfoglicerynian
Etap 7:
Utworzenie
ATP
Z ADP –
odzysk
energii.
kinaza
fosfoglicerynianowa
C
O
OPO
3
2-
C
H
OH
CH
2
OPO
3
2-
1,3-difosfoglicerynian
ADP
ATP
C
O
O
-
C
H
OH
CH
2
OPO
3
2-
3-fosfoglicerynian
Etap 8:
Izomeryzacja
fosfogliceromutaza
C
O
O
-
C
H
OPO
3
2-
CH
2
OH
2-fosfoglicerynian
C
O
O
-
C
H
OH
CH
2
OPO
3
2-
3-fosfoglicerynian
Etap 9:
Odszczepienie
cząsteczki
wody
enolaza
C
O
O
-
C
H
OPO
3
2-
CH
2
OH
2-fosfoglicerynian
C
O
O
-
C
H
OPO
3
2-
C
H
H
fosfoenolopirogronian
Etap 10:
Znów odzysk
energii – powstaje
ATP z ADP
kinaza
pirogronianowa
C
O
O
-
C
H
OPO
3
2-
C
H
H
fosfoenolopirogronian
ADP
ATP
C
O
O
-
C O
CH
3
pirogronian

Bilans energetyczny glikolizy
Reakcja
Przemiana ADP ATP
Glukoza glukozo-6-fosforan
-1
Fruktozo-6-fosforan fruktozo-1,6-difosforan
-1
2 cząsteczki 1,3-difosfoglicerynianu
2
cząsteczki 3-fosfogliceraynianu
+2
2 cząsteczki fosfoenolopirogronianu
2 cząsteczki pirogronianu
+2
NETTO
+2

Podział wg zapotrzebowania na
O2:
- ścisłe tlenowce: Streptomyces (antybiotyki jako
wtórny metabolit)
- ścisłe beztlenowce: en. zdobywana na drodze
fermentacji (heterofermentacja) Clostridium.
acetobutylicum (produkcja acetonu) C. botulinum
(wytwarzanie toksyny)
- fakultatywne: zdobywają en. dwoma sposobami
(fermentacja lub/i fosforylacja oksydatywna)

Oddychanie beztlenowe -
fermentacje
Zdobywanie en. na drodze fermentacji jest znacznie
niższe (2ATP)
Fermentacja alkoholowa: glu ->EtOH + CO2 (2ATP)
Warunki tlenowe (36 ATP)
Potencjalne substraty są wydzielane z komórki.
Drożdże - EtOH i kw. mlekowy nie mogą być dalej
metabolizowane i są wydzielane do podłoża.

Typy fermentacji:
homofermentacja (1 produkt) np. mlekowa (glu ->kw.
mlekowy) - Lactococcus lactis
heterofermentacje (2 lub więcej produktów) np.
mlekowa (glu -> kw. mlekowy + CO2 + alkohol)
Lactobacillus; 4 produkty: etanol, metanol, propanol,
butanol C. acetobutylicum
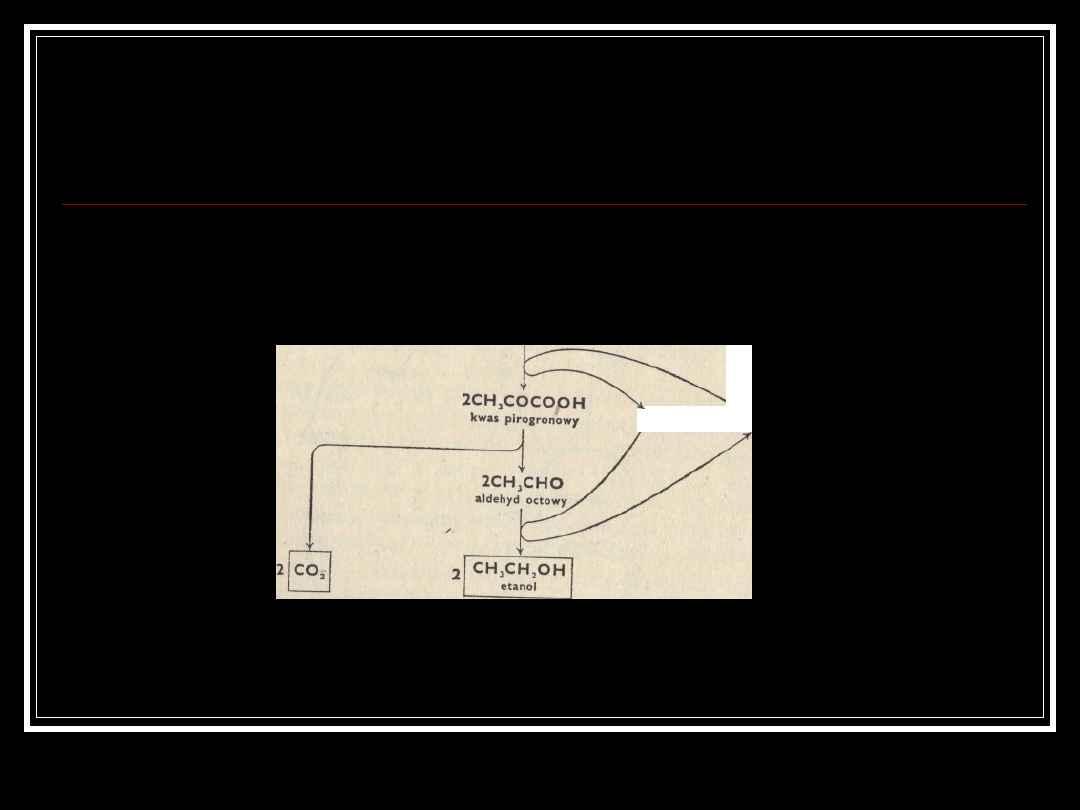
Saccharomyces cerevisiae
-fermentacja alkoholowa
glukoza

Homofermentacje:
Alkoholowa
Mlekowa – (kwas mlekowy) Streptococcus
lactis, Lactobacillus casei

Heterofermentacje:
Rzekomo-mlekowa (kwas mlekowy, kwas octowy,
kwas mrówkowy, etanol, CO2,H2) - Streptococcus
cremonis, Lactobacillus brevis
Masłowa (kwas masłowy, kwas octowy, etanol, CO2,
H2) – Clostridium butyricum
Propionowa (kwas propionowy, kwas octowy, kwas
bursztynowy, CO2) – Propionibacterium spp
Acetonowo-butanolowa (butanol, aceton, CO2, H2)
Clostridium acetobutyricum
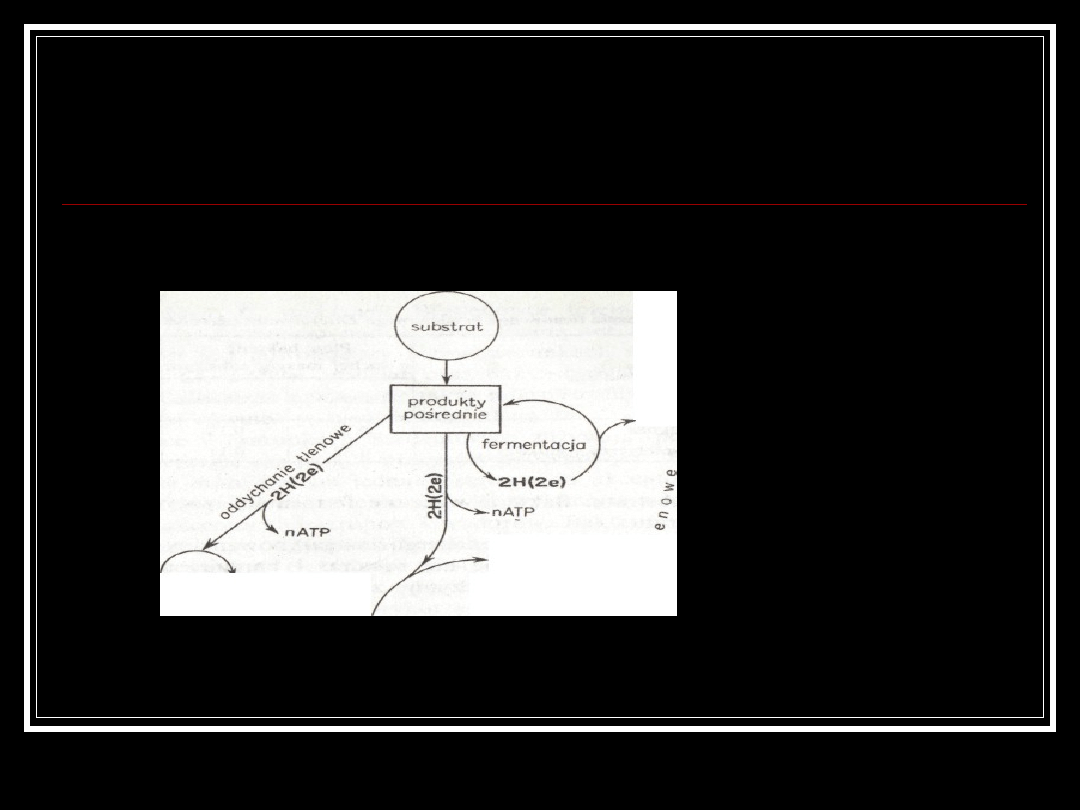
Ogólny schemat oddychania
tlenowego i beztlenowego
utleniony związek
nieorganiczny

Oddychanie z redukcja
związków mineralnych
Denitryfikacja
Desulfurikacja
Redukcja węglanów

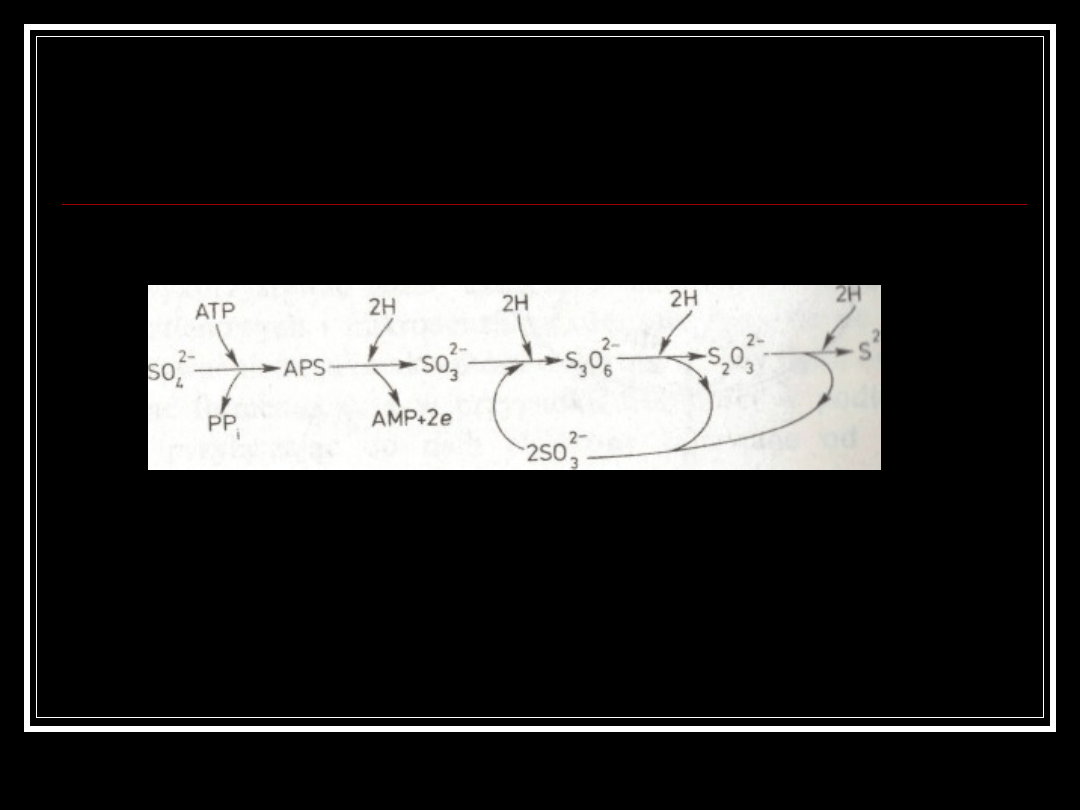
Kataboliczna redukcja
siarczanów.
APS - siarczan adenilowy, PPi — fosforan nieorganiczny

Oddychanie tlenowe
Cykl kwasu cytrynowego

Energia, jaką komórka uzyskuje przy rozpadzie
glukozy na dwa fragmenty (pęka jedno wiązanie)
jest zmagazynowana w dwóch utworzonych
cząsteczkach ATP.
Tymczasem w warunkach dostępu tlenu zachodzi
dalsza przemiana kwasu pirogronowego.
Traci on jeden węgiel i przemienia się w dwu
węglowy fragment C
2
. Ten fragment w postaci
ugrupowania acetylowego (CH
3
CO) przyłącza się
do specjalnego nośnika, jakim jest koenzym A.
Powstaje w ten sposób acetylo-koenzym A
(acetylo-CoA), który dostarcza grupę acetylową
do innego układu, w którym to ostanie pozostałe
z glukozy wiązanie C-C ulegnie rozerwaniu.
W istocie znaczenie acetylo-CoA jest
bardziej ogólne, bo te grupy acetylowe pochodzą
także z rozkładu kwasów tłuszczowych – innego
rodzaju „pożywienia” dla komórki.
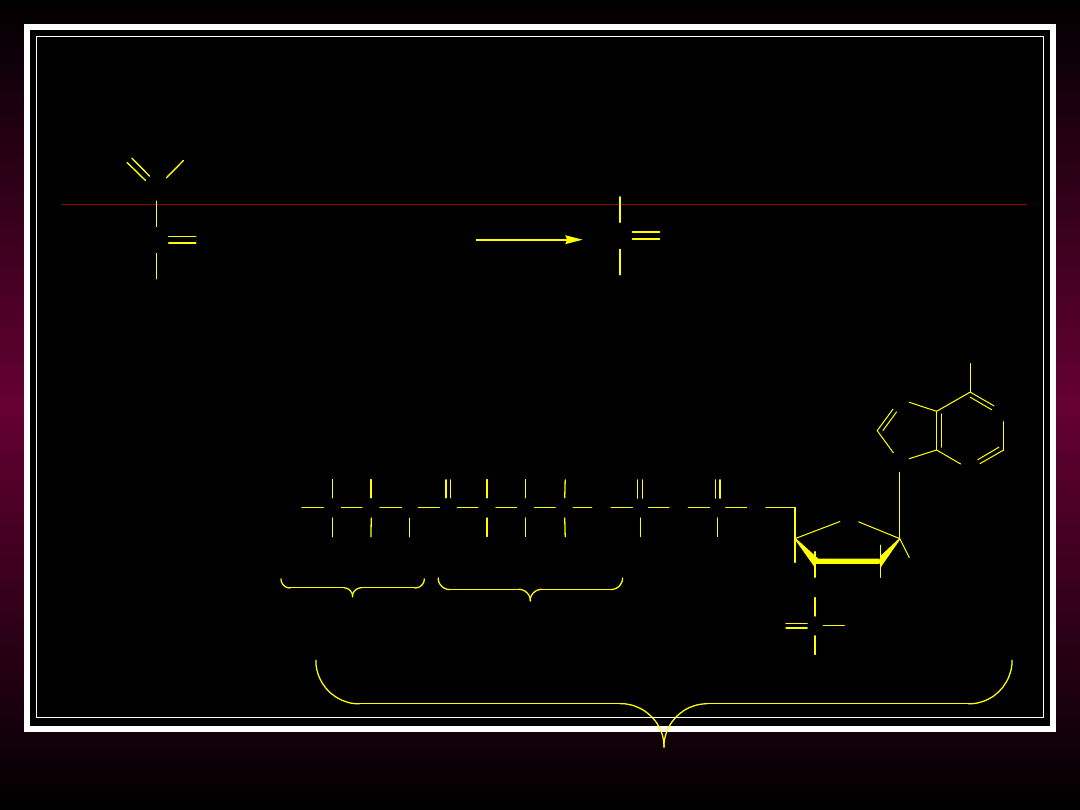
Etap wstępny:
Dekaboksylacja kwasu pirogronowego
C
O
O
-
C O
CH
3
pirogronian
+ CoA + NAD
+
+ CO
2
+ NADH
CoA
C O
CH
3
acetylo-CoA
N
N
N
N
NH
2
O
OH
O
H
H
H
H
O
P
O
O-
O
P O-
O-
O
P
O
O-
C
C
CH
3
CH
3
C
C
N
H
OH
O
H
C
C
HS
H
H
H
H
H
H
O
-merkaptoamina
reszta kwasu
pantotenowego
CoA

Etap 1:
Przyłączenie grupy acetylowej do kwasu szczawiooctowego
(szczawiooctanu)
COO-
C O
CH
2
COO-
CH
2
C COO-
CH
2
COO-
COO-
HO
cytrynian
szczawiooctan
H
3
C C
O
+ H
2
O
enzym: syntaza cytrynianiowa
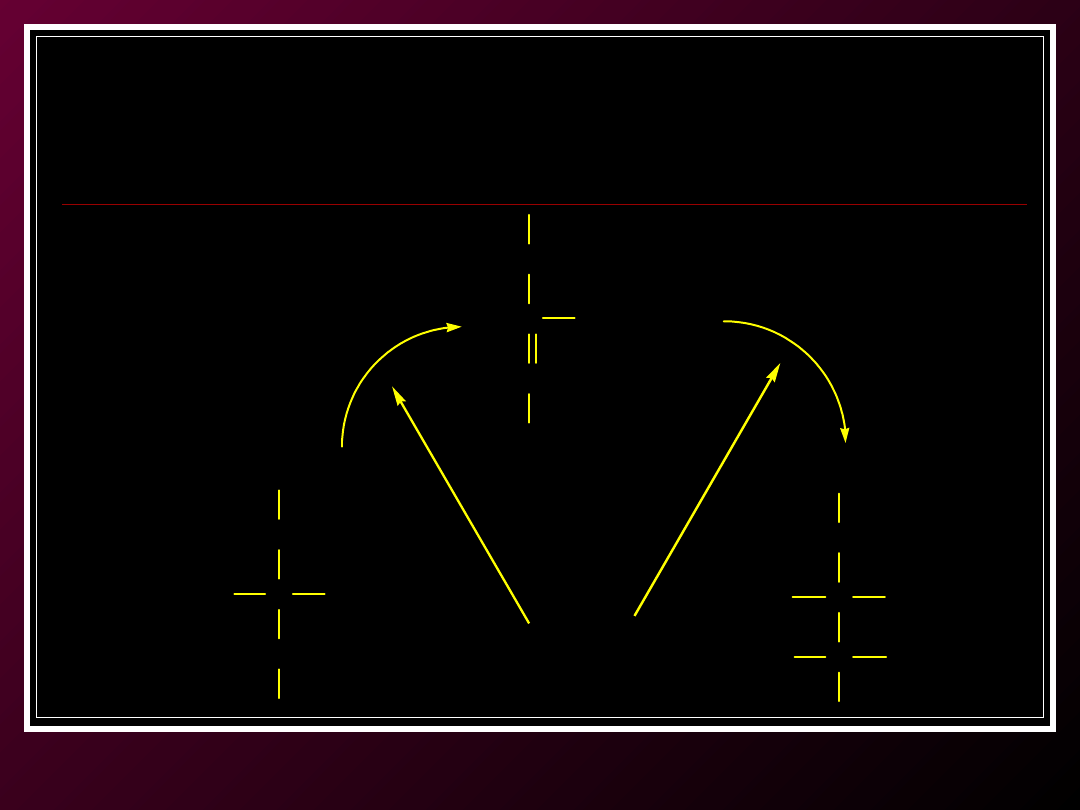
Etap 2:
Przyłączenie grupy acetylowej do kwasu szczawiooctowego
(szczawiooctanu)
CH
2
C COO-
CH
2
COO-
COO-
HO
cytrynian
CH
2
C COO-
CH
COO-
COO-
cis-akonitan
CH
2
C COO-
C
COO-
COO-
H
izocytrynian
H
HO
enzym: akonitaza
-H
2
O
+H
2
O
CH
2
C COO-
C
COO-
COO-
H
izocytrynian
H
HO
enzym: dehydrogenaza izocytrynianowa
CH
2
C COO-
C
COO-
COO-
H
szczawiobursztynian
O
CH
2
C H
C
COO-
COO-
H
-ketoglutaran
O
NAD
+
NADH +
H
+
CH
2
C COO-
C
COO-
COO-
H
szczawiobursztynian
O
H
+
CO
2
Etap 3:
Odszczepienie jednej cząsteczki CO
2
,
energia zmagazywnana w NADH

Etap 4:
Odszczepienie drugiej cząsteczki CO
2
,
energia zmagazywnana w NADH
enzym:
dehydrogenaza -ketoglutarowa
CoA + NAD
+
CO
2
+ NADH
CH
2
C
H
C
COO-
COO-
H
-ketoglutaran
O
C
CH
2
CH
2
COO-
O
bursztynylo-CoA
S-CoA

Etap 5:
Energia przemiany zmagazynowana w GTP
enzym:
syntetaza bursztynylo-CoA
GTP + CoA
C
CH
2
CH
2
COO-
O
bursztynylo-CoA
S-CoA GDP + P
i
C
CH
2
CH
2
COO-
O
bursztynian
O-

Etap 6 - 8:
Przemiany fragmentów czterowęglowych
C
CH
2
CH
2
COO-
O
bursztynian
O-
C
CH
CH
COO-
O
fumaran
O-
FADH
2
H
2
O
C
C
CH
2
COO-
O
jab³czan
O-
HO
H
C
C
CH
2
COO-
O
szczawiooctan
O-
O
NADH
Szczawiooctan odtworzył się. W efekcie całego cyklu
rozerwaniu uległy dwa wiązania węgiel-węgiel. Energia
rozpadu wiązań została zmagazynowana w NADH i
FADH
2
. Proces przebiega wyłącznie w warunkach
tlenowych.

Bioenergetyka – łańcuch oddechowy
Fosforylacja oksydacyjna

Łańcuch oddechowy
Dehydrogenazy
NAD, NADP
FMN, FAD
Cytochrom b
Ubichinon
Cytochrom c
Cytochrom a
Oksydaza cytochromowa
NADH I FADH
2
są głównymi przenośnikami elektronów w
procesie utleniania „paliwa molekularnego”
N
N
N
N
H
3
C
H
3
C
H
H
O
H
R
O
+ 2H
+
+ 2e
-
N
N
N
N
H
3
C
H
3
C
H
H
O
H
R
O
H
H
FAD (forma utleniona)
FADH
2
(forma utleniona)

W rezultacie elektrony te są przekazywane
w kaskadowych reakcjach tlenowi
cząsteczkowemu – O
2
(stopień utlenienia –
zero) w reakcji:
O
2
+ 4H
+
+ 4e
-
2H
2
O

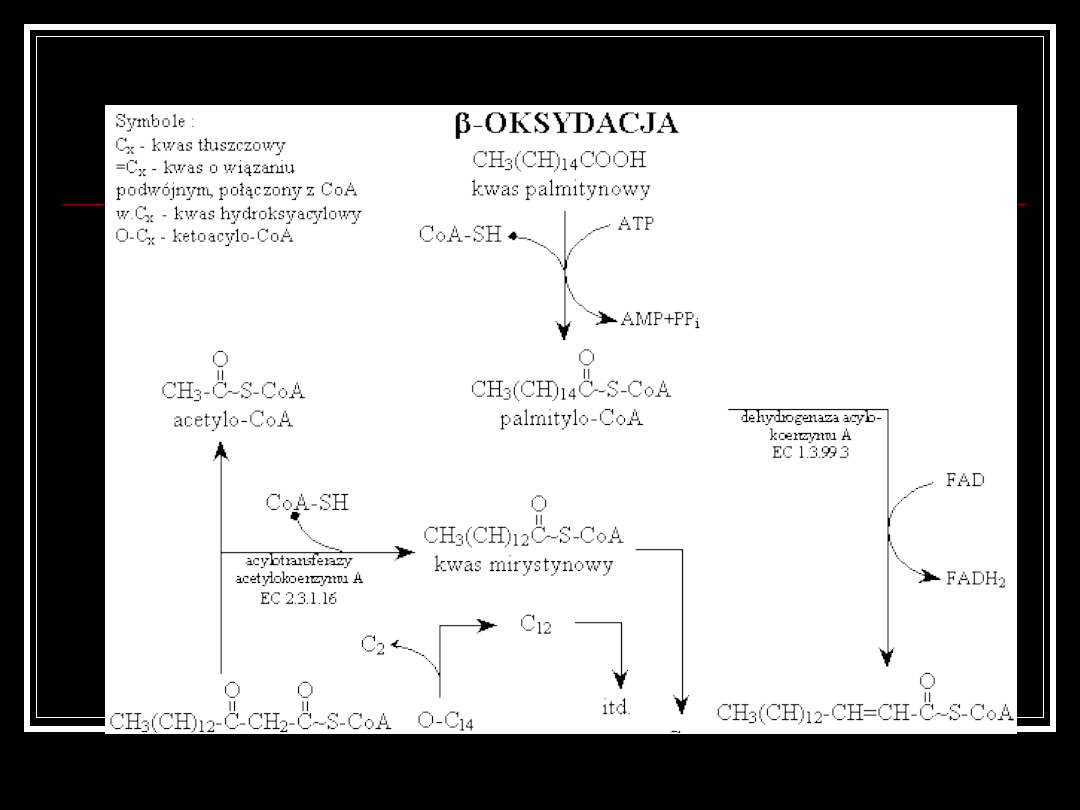
BETA-OKSYDACJA
TŁUSZCZÓW
Kwasy tłuszczowe ulegają spaleniu w procesie
beta-oksydacji.
Polega on na cyklicznym odłączeniu reszty kwasu
octowego połączonej z koenzymem A, która
zostaje rozłożona w cyklu Krebsa.
Łańcuch jest w ten sposób krótszy o dwa atomy
węgla za każdym razem.

W pierwszym etapie kwas tłuszczowy łączy się z CoA i w obecności
dehydrogenazy acylo-koenzymu A (EC 1.3.99.3) ulega odwodornieniu
przez FAD. Powstaje kwas o wiązaniu podwójnym, połączony z CoA.
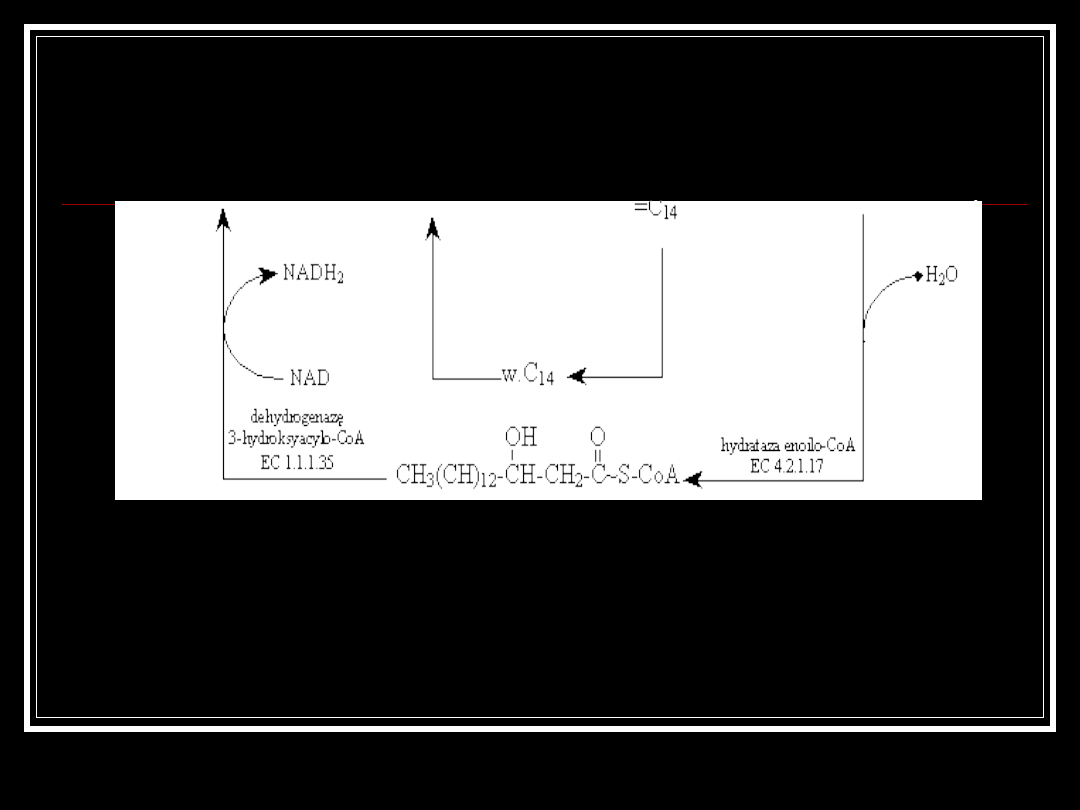
W drugim etapie podwójne wiązanie ulega rozerwaniu w wyniku
przyłączenia wody w obecności enzymu hydratazy enoilo-CoA - EC
4.2.1.17. Powstaje kwas hydroksyacylowy połączony z CoA.
Ten z kolei ulega dehydrogenacji w reakcji katalizowanej przez
dehydrogenazę 3-hydro-ksyacylo-CoA (EC 1.1.1.35), w której
koenzymem jest NAD i powstaje ketoacylo-CoA
W wyniku działania enzymu acylotransferazy acetylokoenzymu A (EC
2.3.1.16) następuje odłączenie acetylo-CoA od dotychczasowego
kwasu, w wyniku czego łańcuch kwasu tłuszczowego jest o dwa
atomy węgla krótszy i do którego ponownie przyłącza się CoA. Proces
się powtarza. Końcowym produktem b-oksydacji jest butyrylo-CoA,
który rozkładany jest na dwie cząsteczki acetylo-CoA

fotosynteza
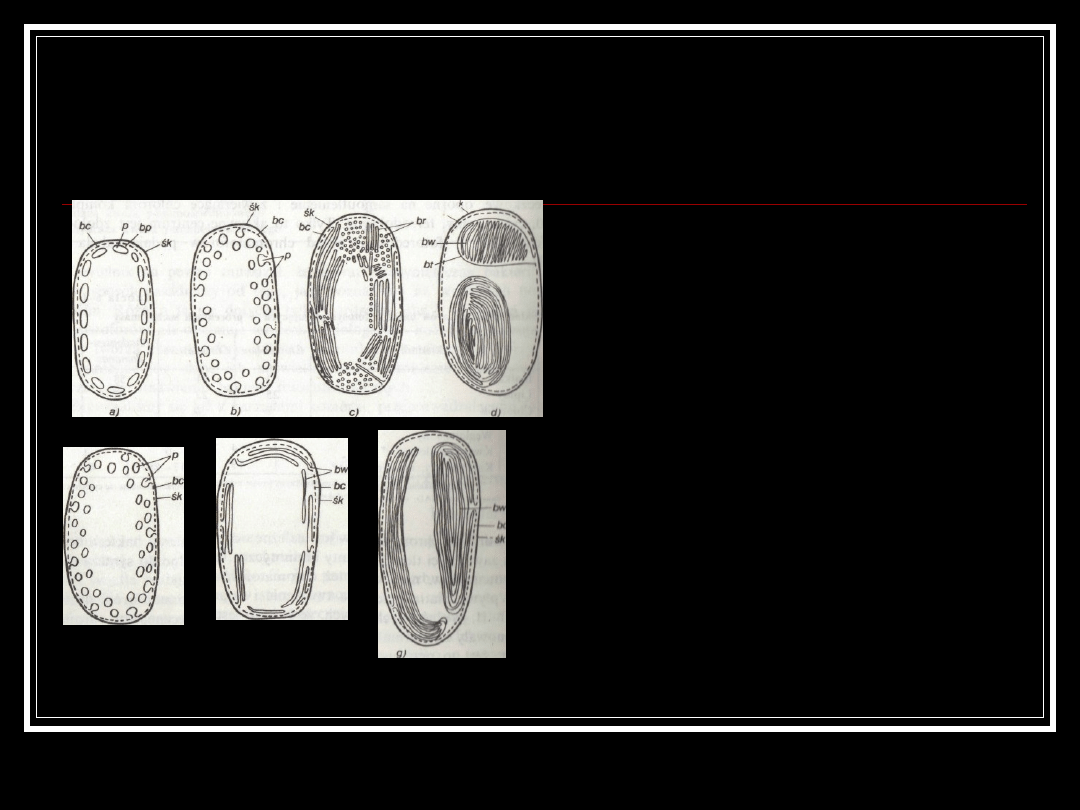
Chromatofory.
U Prokaryota chloroplastów nie ma. U
sinic i u syntetyzujących bakterii ich
rolę spełniają stosunkowo proste,
błoniu twory, tylakoidy, mające formę
pęcherzyków lub warstwowo ułożonych
błon. Bakteryjne organelle nazywamy
chromatoforami. Chromatofory mogą
mieć kształt pęcherzyków niemal
kulistych, 40-100 nm średni
rozproszonych w cytoplazmie, jak u
purpurowych bakterii bezsiarkowych
(Rhodospirillum rubrum,
Rhodopseudomonas spheroides), ciałek
owoidalnj) 30-40x100-150 nm,
położonych w zewnętrznej warstwie
cytoplazmy, u bakterii zielonych
(Chlorobium), lub rozproszonych w
cytoplazmie n purpurowych bakterii
siarkowych (Chromatium,
Thiospirillum).
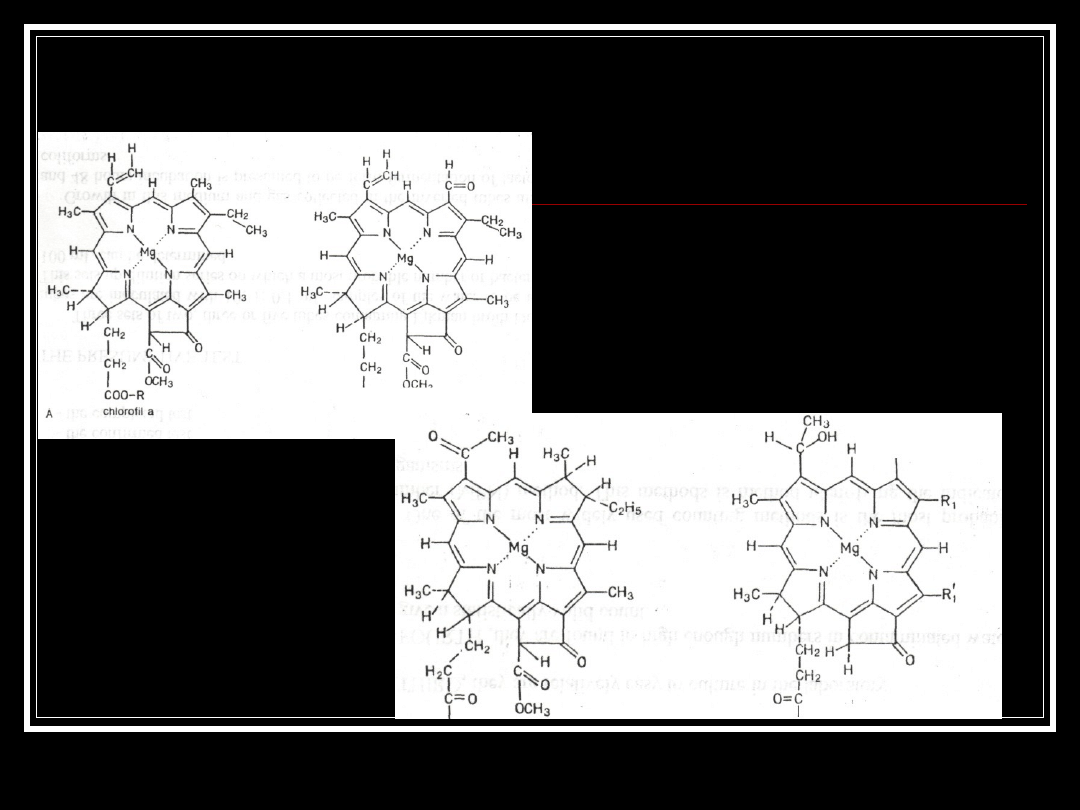
Chlorofile Eucaryota (A) i
Procaryota (B)
0~R
bakteriochlorofil a
0-R;,
chlorofil typu Chlorobium
chlorofil b
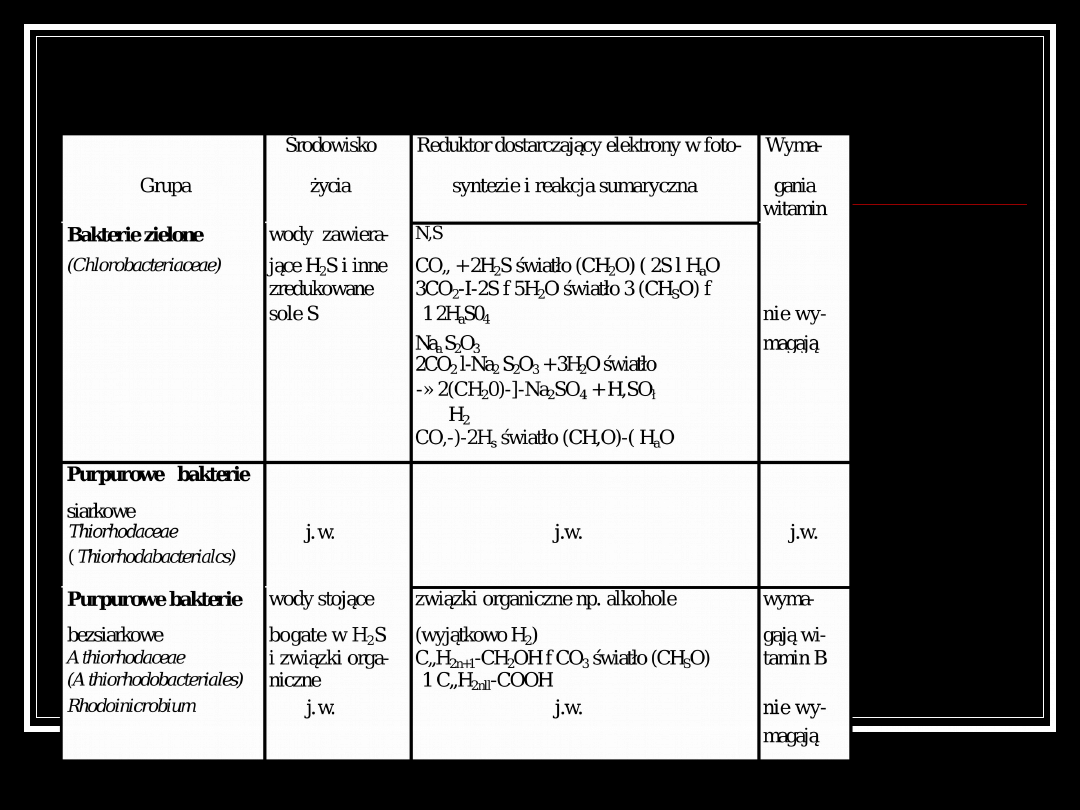
Podział bakterii
fotosyntetyzujących
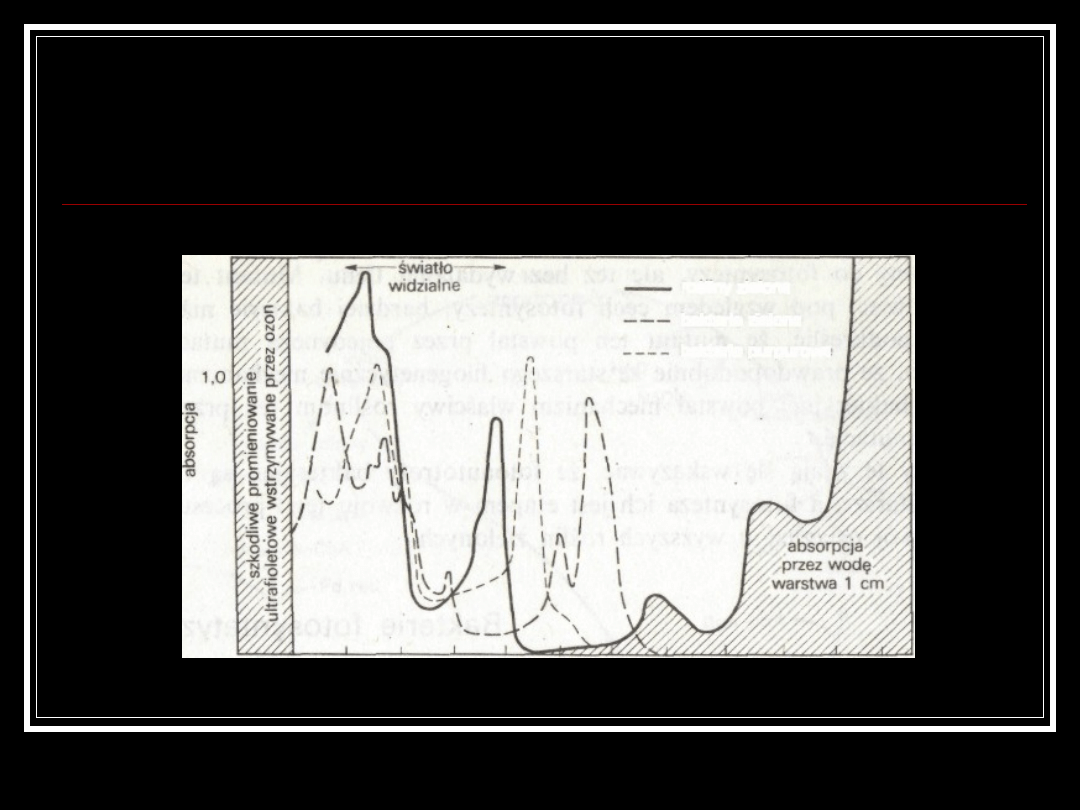
Widma absorpcyjne organizmów
fotosyntetyzujących pokazujące odcinki widma,
jakie mogą być wykorzystane w fotosyntezie.
długość fali, nanometry

Podział bakterii
chemosyntetyzujących
Document Outline
- Metabolizm
- Podział drobnoustrojów wg rodzaju:
- Slide 3
- Typy procesów oddechowych
- Przebieg procesów związanych z pozyskiwaniem energii
- SZLAKI DEGRADACJI CUKRÓW do stadium pirogronianu
- Slide 7
- Szlak pentozofosforanowy
- cykl Endnera Doudoroffa
- Slide 10
- Slide 11
- Slide 12
- W trakcie glikolizy zachodzi:
- Slide 14
- Slide 15
- Slide 16
- Slide 17
- Slide 18
- Slide 19
- Slide 20
- Slide 21
- Slide 22
- Slide 23
- Slide 24
- Slide 25
- Podział wg zapotrzebowania na O2:
- Oddychanie beztlenowe - fermentacje
- Typy fermentacji:
- Saccharomyces cerevisiae -fermentacja alkoholowa
- Homofermentacje:
- Heterofermentacje:
- Ogólny schemat oddychania tlenowego i beztlenowego
- Oddychanie z redukcja związków mineralnych
- Slide 34
- Kataboliczna redukcja siarczanów. APS - siarczan adenilowy, PPi — fosforan nieorganiczny
- Slide 36
- Slide 37
- Slide 38
- Slide 39
- Slide 40
- Slide 41
- Slide 42
- Slide 43
- Slide 44
- Slide 45
- Łańcuch oddechowy
- Slide 47
- Slide 48
- BETA-OKSYDACJA TŁUSZCZÓW
- Slide 50
- Slide 51
- Slide 52
- fotosynteza
- Chromatofory.
- Chlorofile Eucaryota (A) i Procaryota (B)
- Podział bakterii fotosyntetyzujących
- Widma absorpcyjne organizmów fotosyntetyzujących pokazujące odcinki widma, jakie mogą być wykorzystane w fotosyntezie.
- Podział bakterii chemosyntetyzujących
Wyszukiwarka
Podobne podstrony:
Glikoliza prezentacja (ang)
Hydradermabrazja - ablacja połączona z peelingiem glikolowo-salicylowym, Studium kosmetyczne, Pracow
glikoliza, SGGW, biochemia
glikoliza, położnictwo, biochemia, biochemia
Biochemia etapy glikolizy szczegolowo
glikoliza - opis, biologia- studia, Biologia
O glikolach i glicerynie
Glikol dietylenowy
Glikol etylenowy techn
Glikol dietylenowy
Oleksyszyn, Biochemia II, zagadnienia do glikoliza i glukoneogeneza
glikoliza
Krebs 1P 60
Przebieg glikolizy
bioch 8 glikoliza fermentacje szlak fosforanow pentoz
Glikol polietylenowy@0
10 Biochemia glikoliza
glikoliza
więcej podobnych podstron