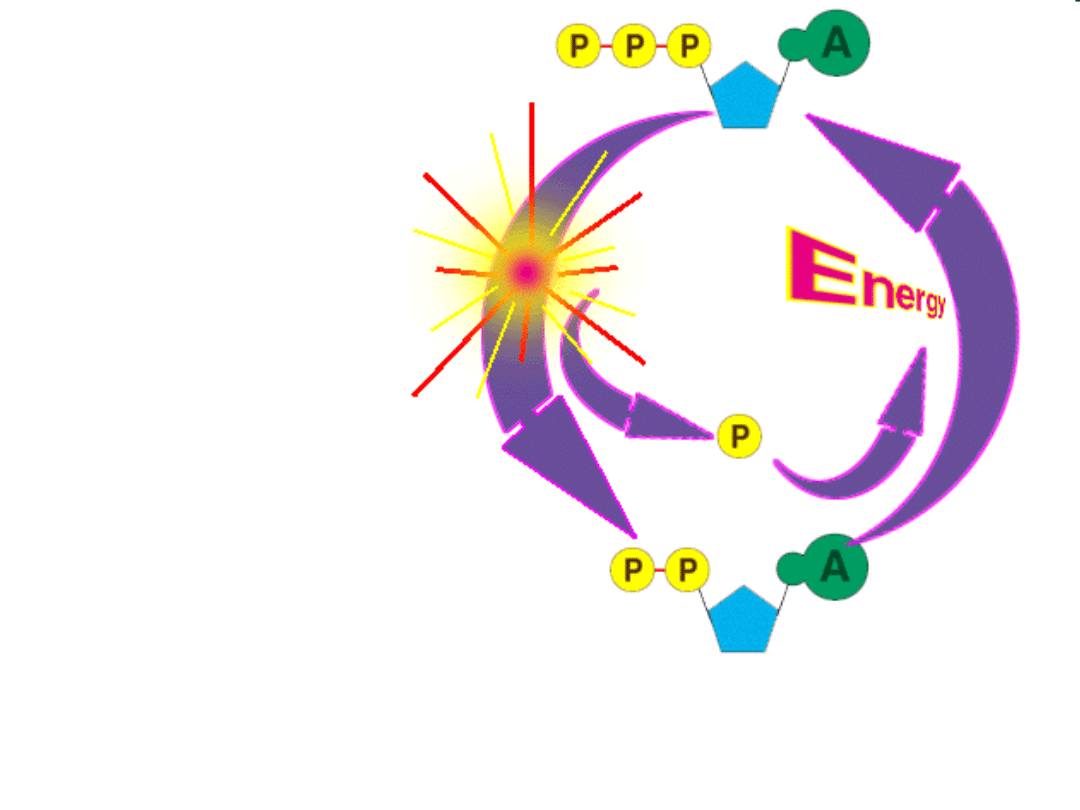
ATP
• ADP
+
P
+
Energia
--->
ATP
• ATP
-->
ADP
+
P
+
Energia
Rozerwaniu wiązania
ostatniego fosforanu
z ATP towarzyszy
wydzielenie energii
7300 cal/mol czyli
30500 J/mol lub
30,5kJ/mol.
Masa cząsteczkowa
ATP wynosi 507.2
czyli około 60 kJ/ kg.
+
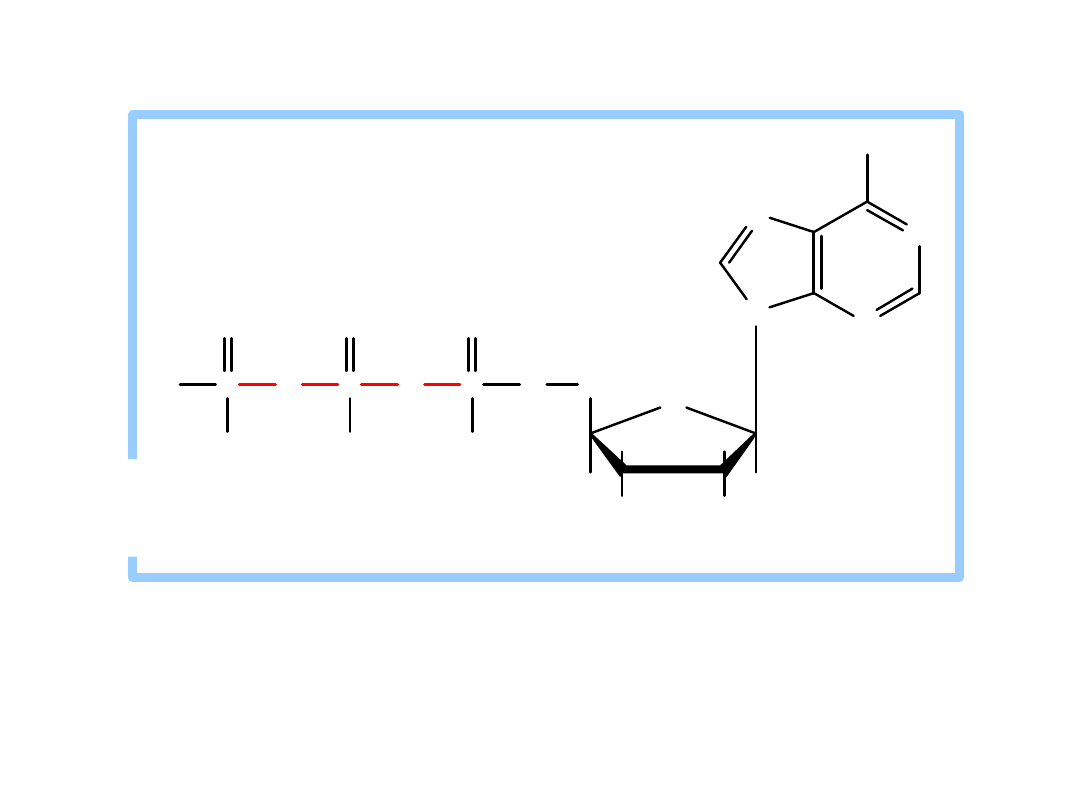
Wysokoenergetyczne wiązania
Bezwodnikowe wiązania
(tworzone między cząsteczkami
kwasu fosforowego lub między cząsteczką kwasu fosforowego
i grupą karbonylową przez odłączenie cząsteczki wody) mają
duży ujemny potencjał termodynamiczny (dG hydrolizy)
N
N
N
N
NH
2
O
OH
OH
H
H
H
CH
2
H
O
P
O
P
O
P
-O
O
O-
O-
O
O
O-
adenine
ribose
ATP
adenosine triphosphate
phosphoanhydride
bonds (
~
)
adenozynotrójfosforan
Wiązania bezwodnikowe
wysokoenergetyczne (
-
)
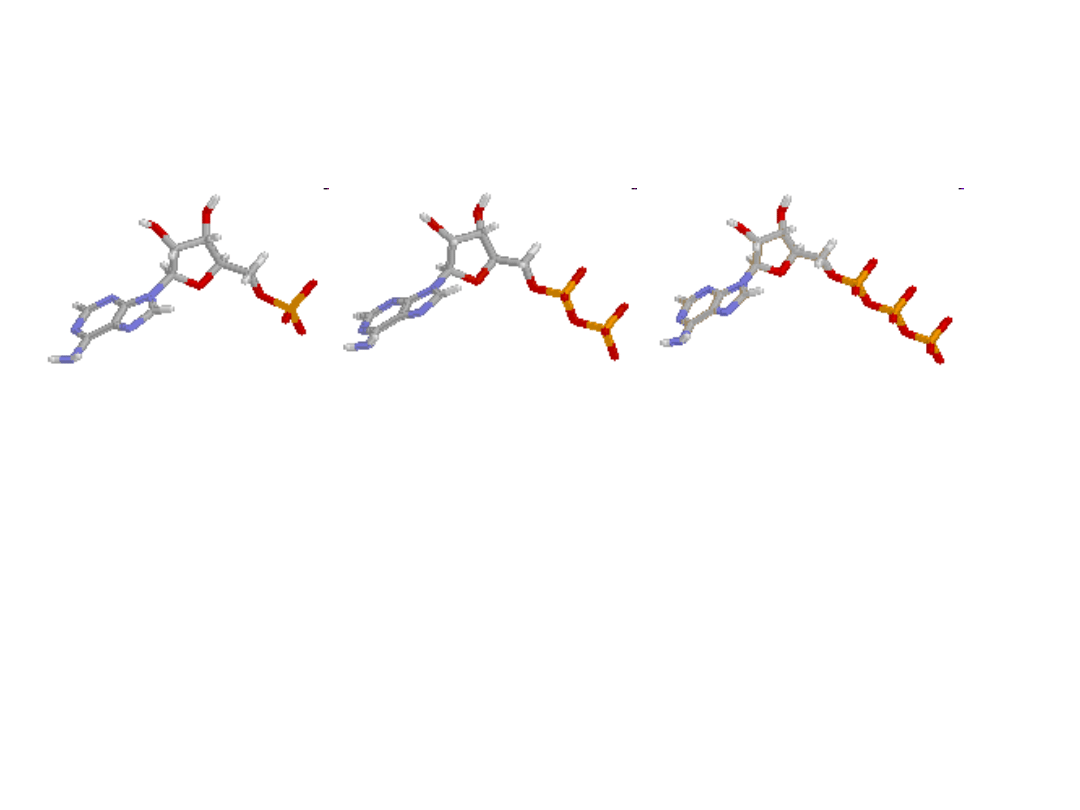
AMP, ADP, ATP
• AMP ADP ATP
• Cząsteczka ATP jest nukleotydem składającym się z
zasady azotowej - adeniny połączonej wiązaniem N-
glikozydowym z cząsteczką cukru - rybozy i trzech reszt
fosforanowych połączonych ze sobą dwoma wiązaniami
bezwodnikowymi. Reszty fosforanowe są oznaczane w
ogólnie przyjętej notacji greckimi literami α, β i γ.

Zużycie ATP
• Najbardziej wszechstronnym nośnikiem energii
w komórce jest ATP.
• Jest to donor energii a nie forma
magazynowania
• W ciągu doby człowiek zużywa około 40 kg
• W czasie wysiłku może zużywać 0,5 kg na
minutę
• ATP jest cały czas zużywane i tworzone
Stężenia związków zawierających
fosforan w mięśniach w stanie
spoczynku
ATP 4 mM
ADP 0,013 mM
Fosforan kreatyny 25 mM
Kreatyna 13 mM

Metabolizm to całokształt reakcji biochemicznych
zachodzących w komórkach organizmu. Związany jest z
przepływem materii i energii.
Anabolizm to reakcje syntez związków złożonych z
prostszych, wymagające dostarczenia energii. Energia ta
podnosi poziom energetyczny związków w czasie procesu
chemicznego. Powstający produkt rekcji zawiera większą
ilość energii, niż poszczególne substraty. Dostarczona
energia zostaje zmagazynowana w postaci wiązań
chemicznych.
Katabolizm to reakcje chemiczne podczas których
następuje obniżanie poziomu energetycznego substratów
w wyniku ich rozkładu na związki proste, oraz wydzielana
jest energia wiązań wysokoenergetycznych substratów.
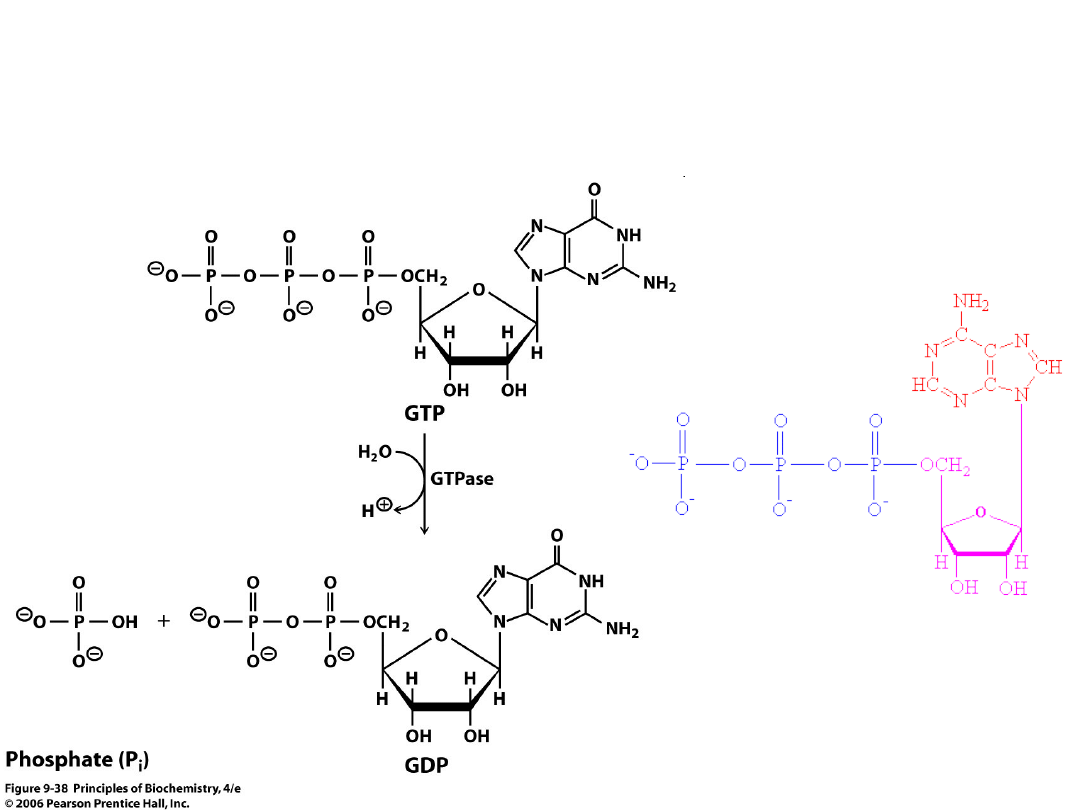
GTP, GDP ATP

• energia z GTP jest łatwo zamieniana na
energię ATP po przez przeniesienie grupy
fosforanowej z GTP na ADP.
• NADH i FADH
2
biorą udział w
mitochondrialnym łańcuchu oddechowym -
przemianie, która zamienia energię tych
zredukowanych związków na energię wiązań
ATP. Niezbędnym uczestnikiem łańcucha
oddechowego jest tlen.
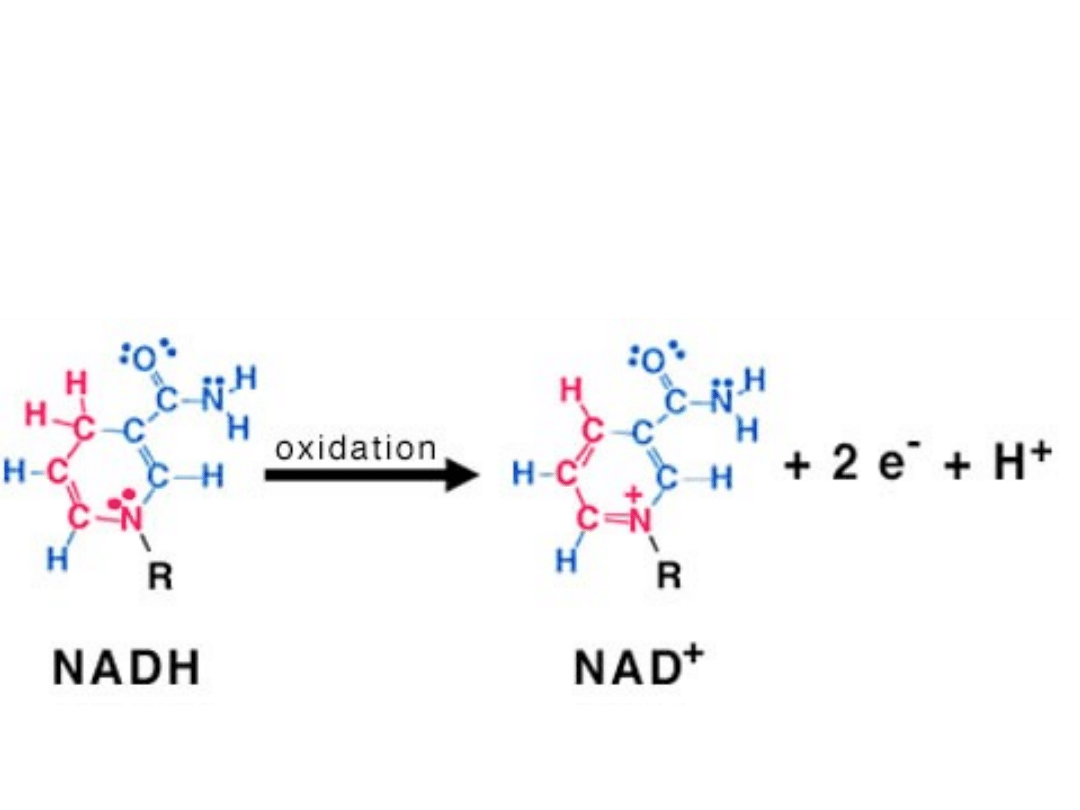
utlenianie NADH
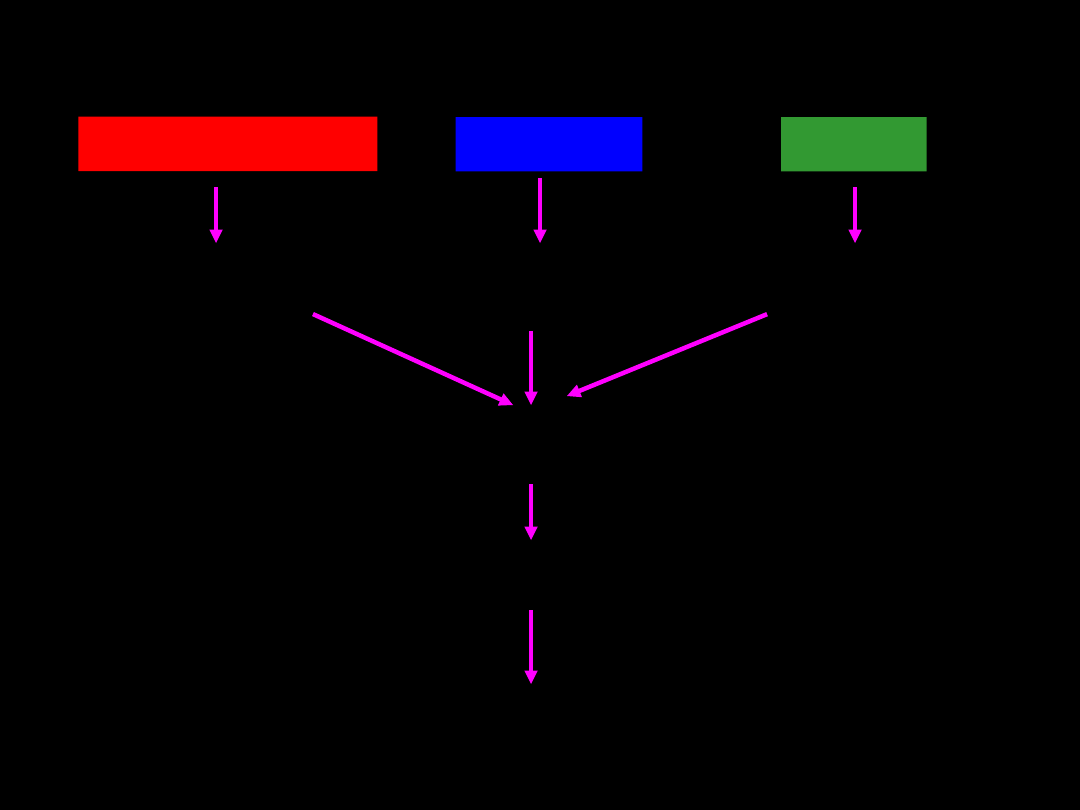
Przemiany energetyczne
organizmu
węglowodany
białka
tłuszcze
Cukry proste
kwasy tłuszczowe
aminokwasy
Acetylo-
koenzym
Cykl Krebsa
Energia w postaci powstającego ATP, NADH oraz FADH2
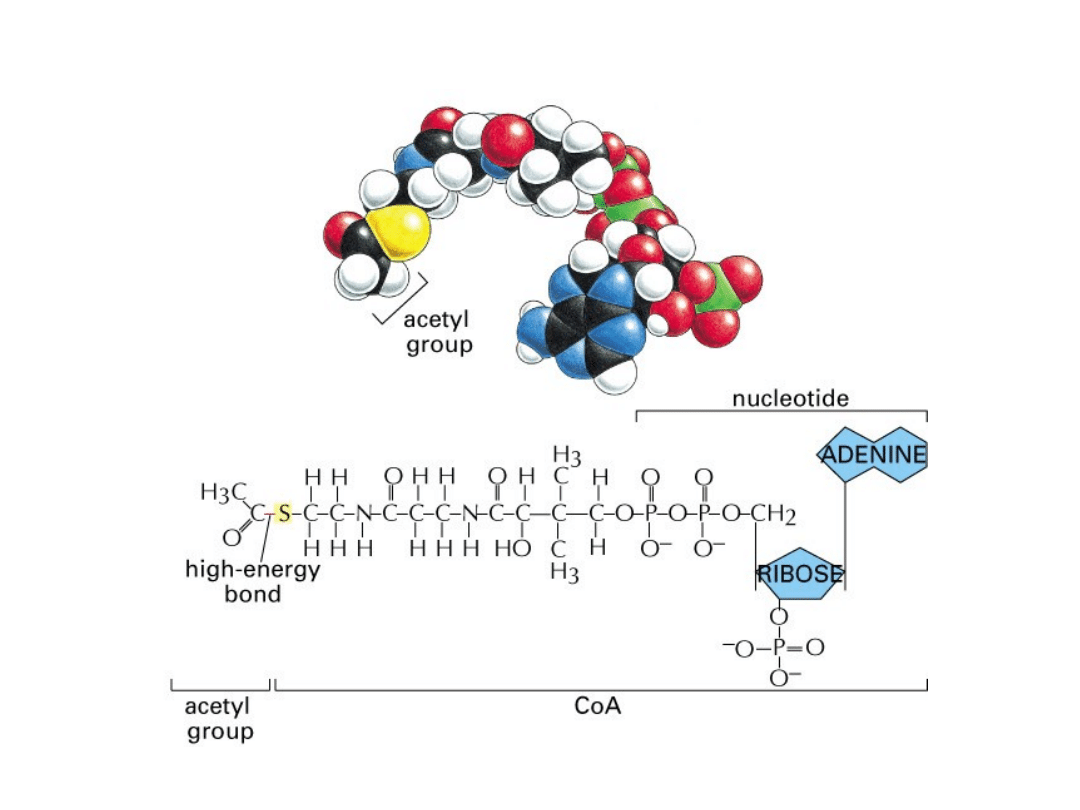
acetyloCoA
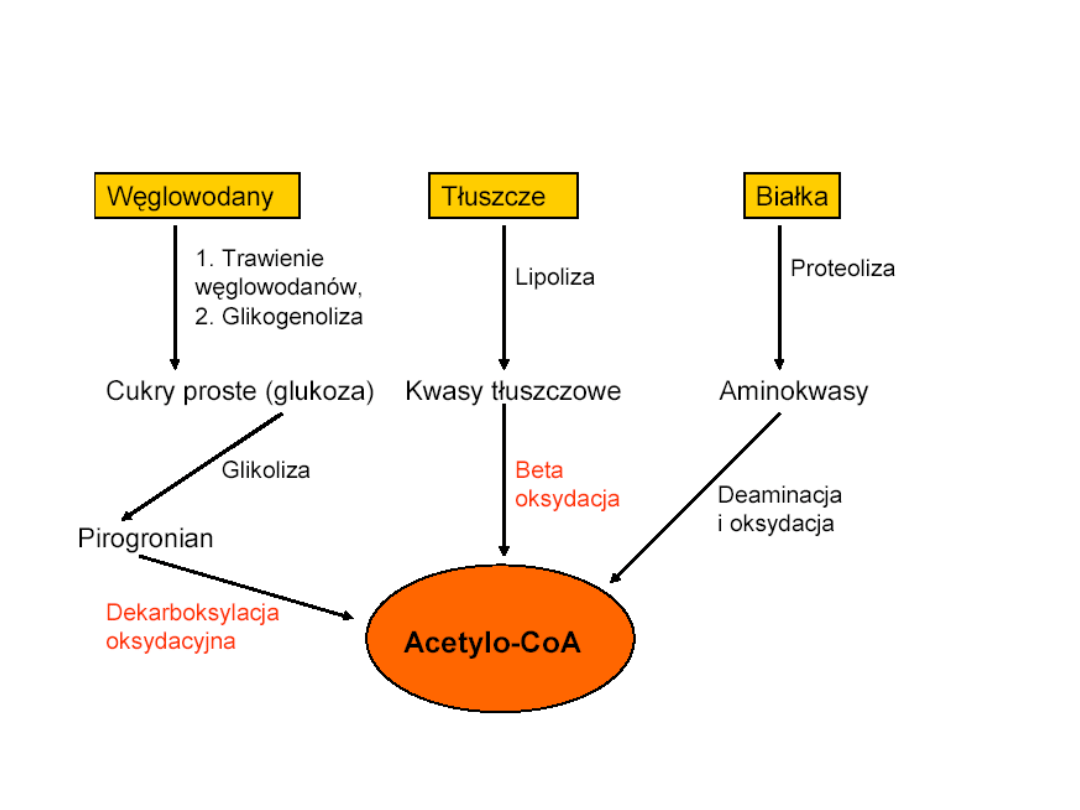
Powstawanie acetylo-CoA

• Cykl Krebsa
, czyli
cykl kwasu cytrynowego
to cykl przemian metabolicznych, który
przebiega w komórkach wszystkich organizmów
oddychających tlenem. Został odkryty w 1937
roku przez Hansa Krebsa.
• Cykl Krebsa u eukariotów zlokalizowany jest
wewnątrz mitochondriów - ważnych organelli
komórkowych (u prokariotów przebiega w
cytoplazmie).
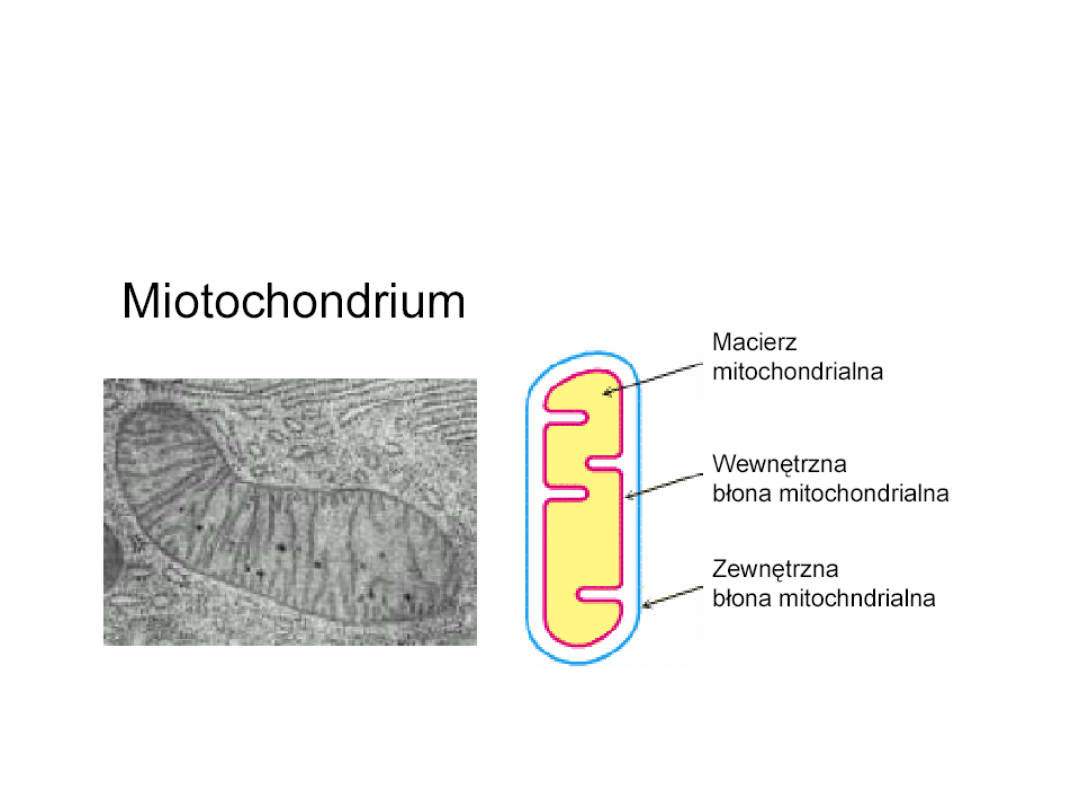
Lokalizacja

• Cykl Krebsa jest końcowym, wspólnym szlakiem
utleniania cząsteczek będących źródłem energii
dla organizmu, takich jak białka (aminokwasy),
kwasy tłuszczowe, węglowodany.
• Jest ciągiem reakcji zachodzących w
mitochondriach,w wyniku których reszty
acylowe ulegają katabolizmowi z uwolnieniem
równoważników wodorowych.

Funkcje cyklu Krebsa
• Dostarcza równoważników redukujących
zamienianych na energię magazynowaną
w ATP w łańcuchu oddechowym
• Dostarcza energii w postaci GTP
• Dostarcza ważnych prekursorów do
syntezy innych cząsteczek

• Zadaniem cyklu Krebsa jest utlenić związek o
nazwie: acetylokoenzym A (acetylo-CoA) do
2 cząsteczek dwutlenku węgla (CO
2
), pozyskaną
w tym procesie energię ulokować w chemicznych
nośnikach energii: GTP, NADH i FADH
2
.
Sumaryczny wzór cyklu Krebsa to:
acetylo
-CoA + GDP + P
i
+ 3NAD
+
+ FAD +
2H
2
0 →
CoA + GTP + 3NADH + 3H
+
+ FADH
2
+ 2
CO
2
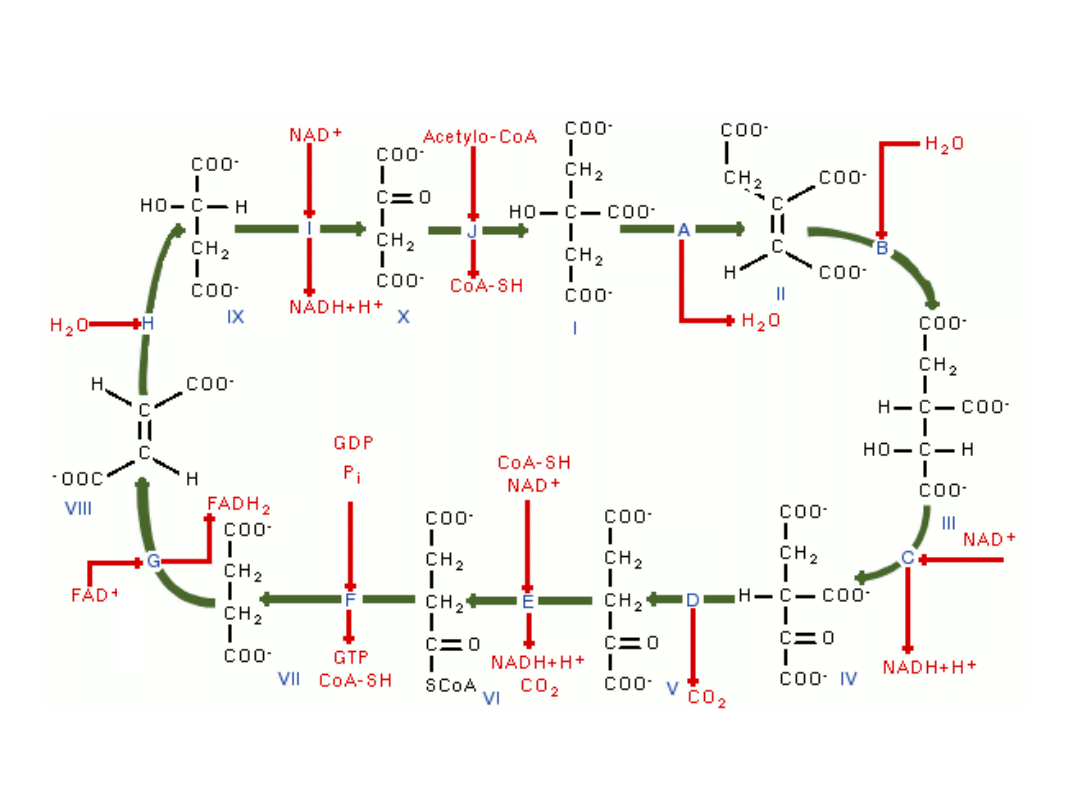
Cykl kwasu cytrynowego (Krebsa)
bursztynian bursztynyloCoA -ketoglutaran
szczawiobursztynian
Jabłczan szczawiooctan
cytrynian
iz
o
cy
tr
y
n
ia
n
Fu
m
a
ra
n
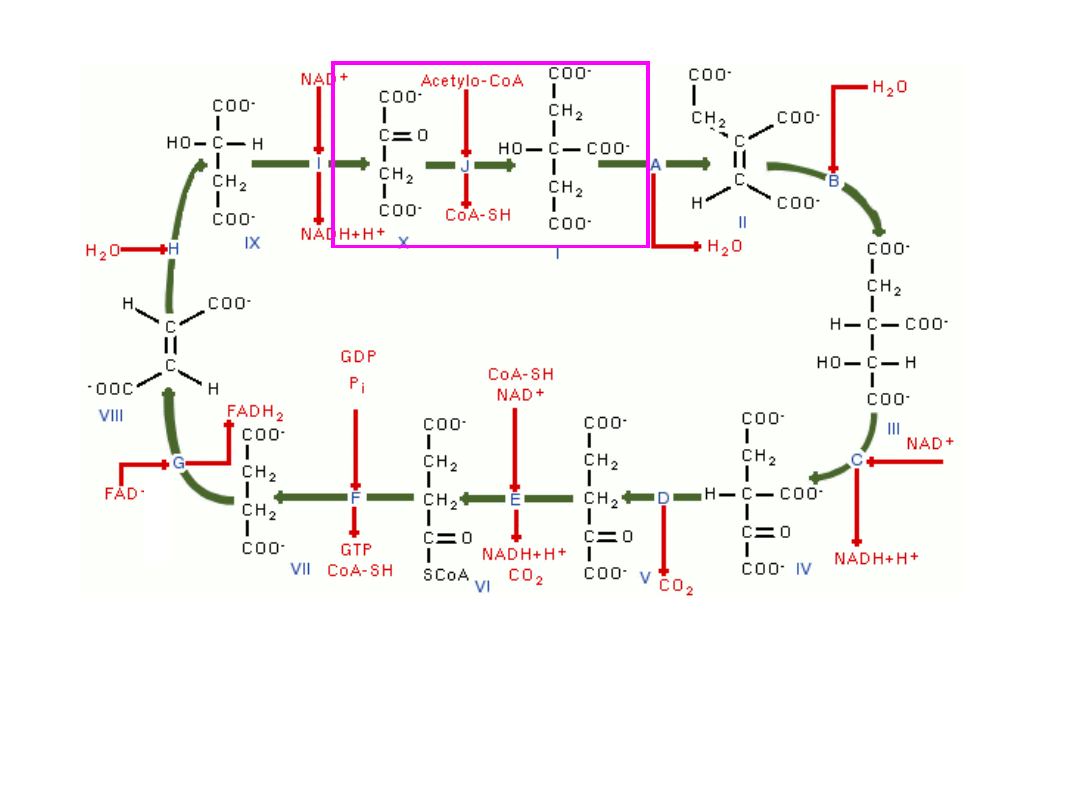
4-węglowy i 2-karboksylowy (4C, 2COOH) szczawiooctan łączy się z
grupą acetylową pochodzącą z acetylo-CoA i tworzy 6-węglowy i 3-
karboksylowy cytrynian –(
enzym-syntaza cytrynianowa
)
bursztynian bursztynyloCoA -ketoglutaran
szczawiobursztynian
Jabłczan szczawiooctan
cytrynian
iz
o
cy
tr
y
n
ia
n
Fu
m
a
ra
n
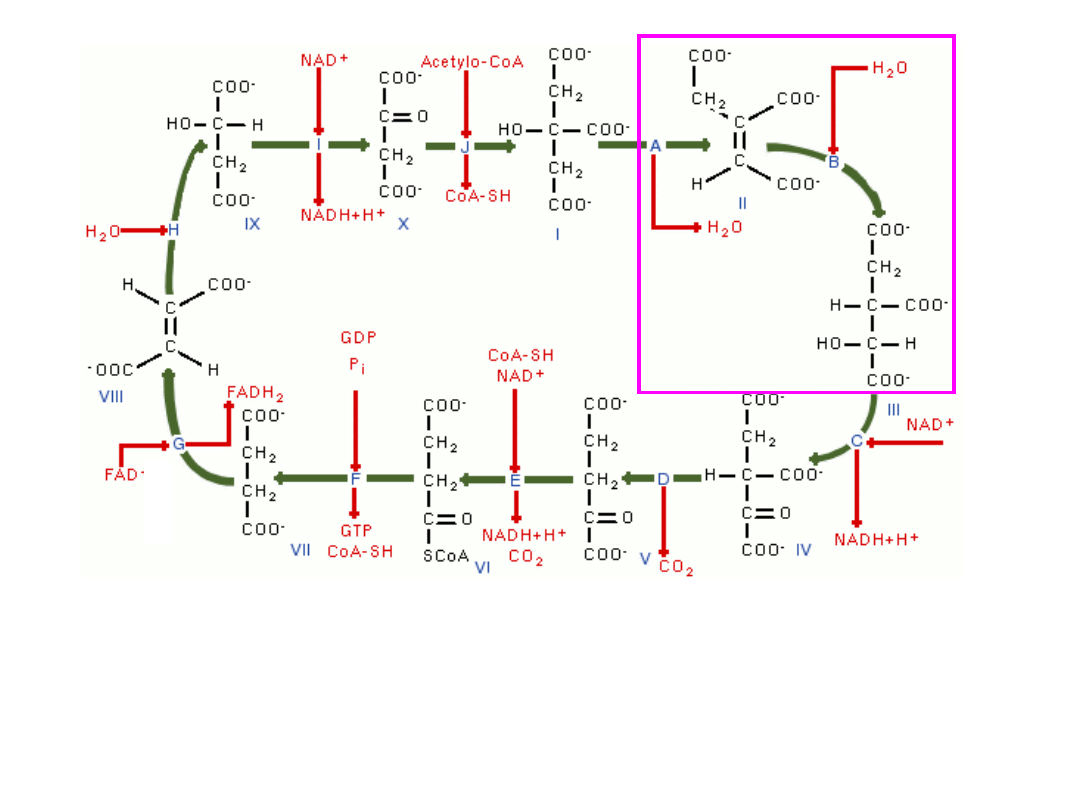
• Następuje izomeryzacja cytrynianu do izocytrynianu czemu
towarzyszy odłączenie i przyłączenie cząsteczki wody –
(
enzym-hydrataza akonitanowa
)
bursztynian bursztynyloCoA -ketoglutaran
szczawiobursztynian
Jabłczan szczawiooctan
cytrynian
iz
o
cy
tr
y
n
ia
n
Fu
m
a
ra
n
6-węglowy i 3-karboksylowy izocytrynian (6C, 3COOH) ulega
utlenieniu (odłączane są dwa atomy wodoru) i przechodzi w
szczawiobursztynian jednocześnie następuje redukcja NAD+ do NADH
–(
enzym-dehydrogenaza izocytrynianowa
)
bursztynian bursztynyloCoA -ketoglutaran
szczawiobursztynian
Jabłczan szczawiooctan
cytrynian
iz
o
cy
tr
y
n
ia
n
Fu
m
a
ra
n
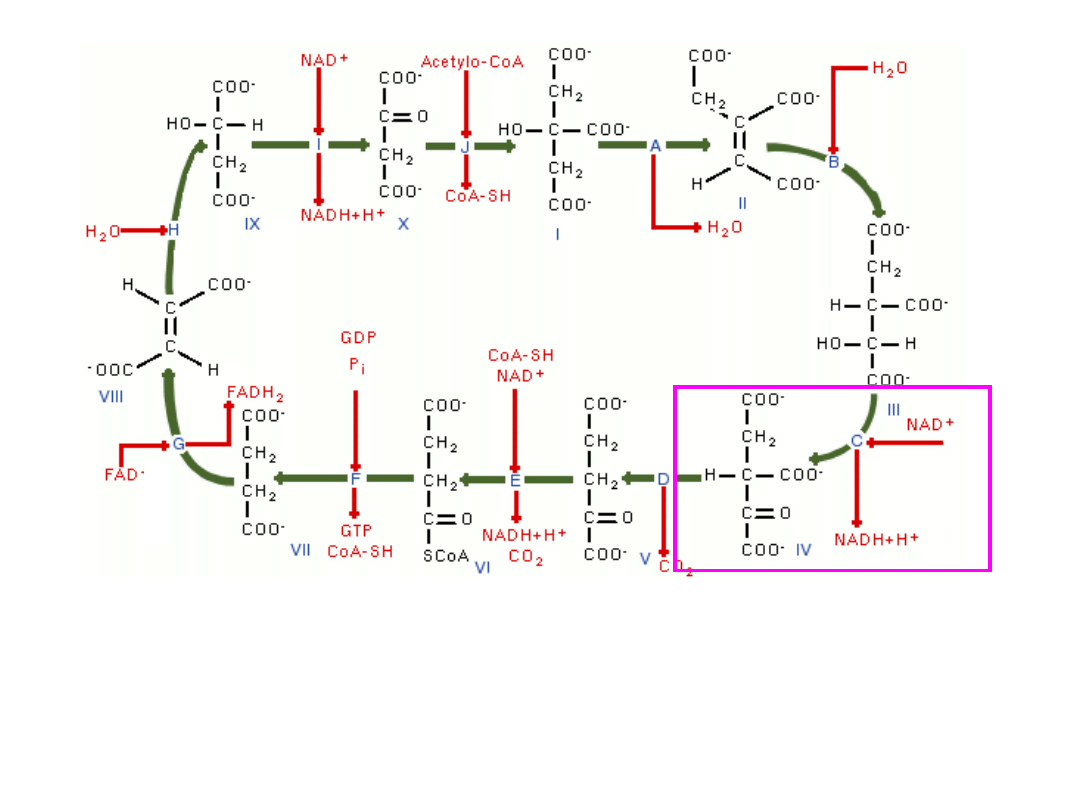
Następuje odłączenie cząsteczki CO
2
, ze szczawiobursztynianu (6C, 3COOH)
powstaje α-ketoglutaran(5C,2COOH) –(
enzym-dehydrogenaza izocytrynianowa
)
Bursztynian bursztynyloCoA -ketoglutaran
szczawiobursztynian
Jabłczan szczawiooctan
cytrynian
iz
o
cy
tr
y
n
ia
n
Fu
m
a
ra
n
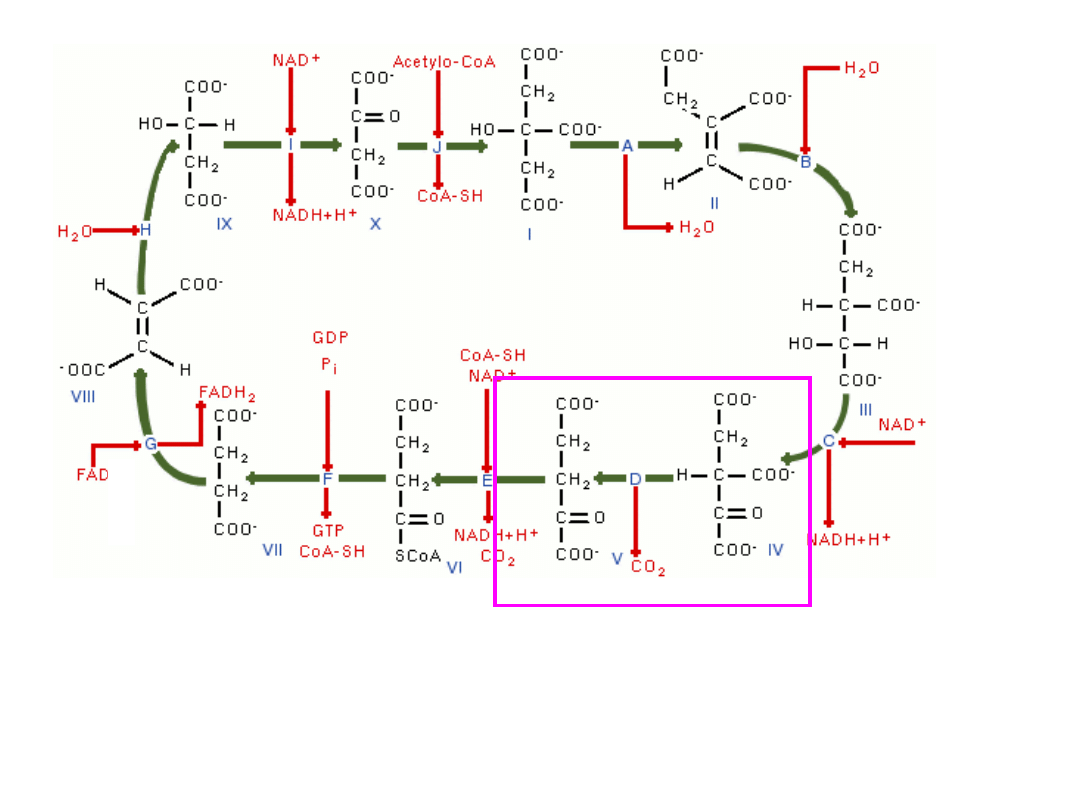
5-węglowy i 2-karboksylowy α-ketoglutaran (5C, 2COOH)
analogicznie do poprzedniego etapu ulega utlenieniu i
dekarboksylacji tworząc
4-węglowy i 1-karboksylowy związek, który łącząc się z koenzymem
A daje bursztynylo-CoA (4C, 1COOH) –(
enzym-dehydrogenaza -
ketoglutaranowa
)
bursztynian bursztynyloCoA -ketoglutaran
szczawiobursztynian
Jabłczan szczawiooctan
cytrynian
iz
o
cy
tr
y
n
ia
n
Fu
m
a
ra
n
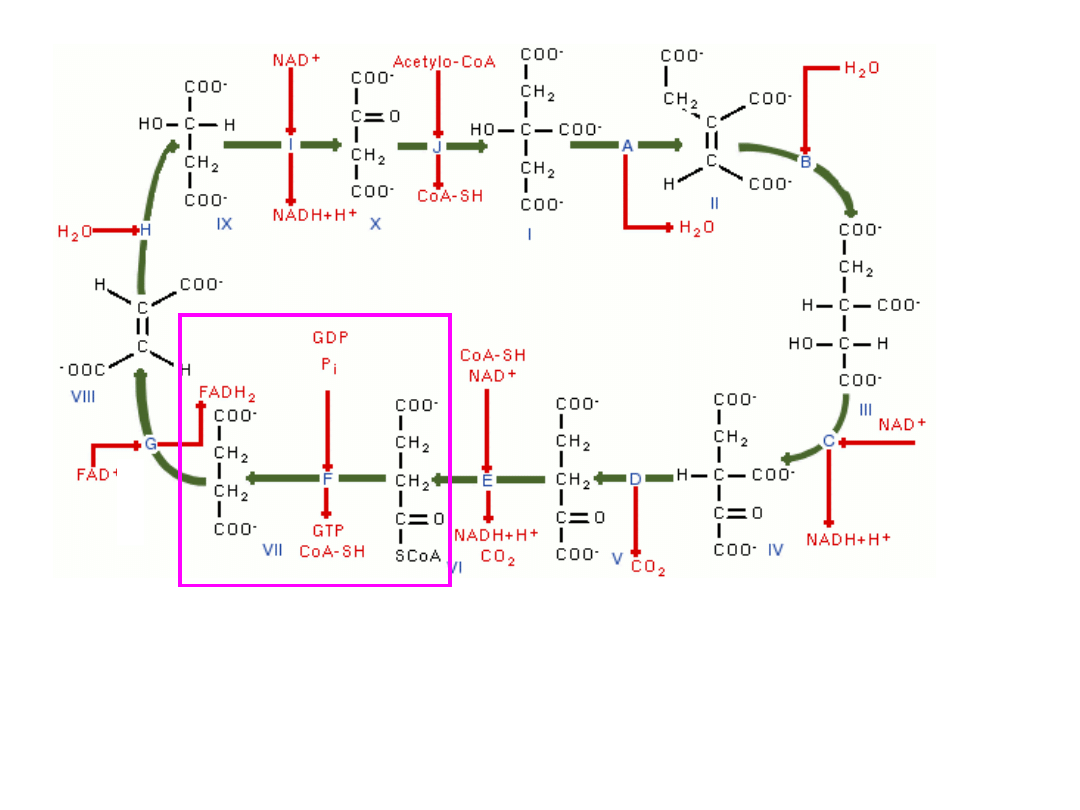
bursztynylo-CoA (4C, 1COOH) odłączając koenzym A tworzy
bursztynian (4C, 2COOH) uwalniając jednocześnie dużą porcję
energii, która za pośrednictwem nukleotydu GTP przekazywana jest
na ADP fosforylując go do ATP – (fosforylacja substratowa} –
(
enzym-syntetaza bursztynylo-CoA
)
Bursztynian bursztynyloCoA -ketoglutaran
szczawiobursztynian
Jabłczan szczawiooctan
cytrynian
iz
o
cy
tr
y
n
ia
n
Fu
m
a
ra
n
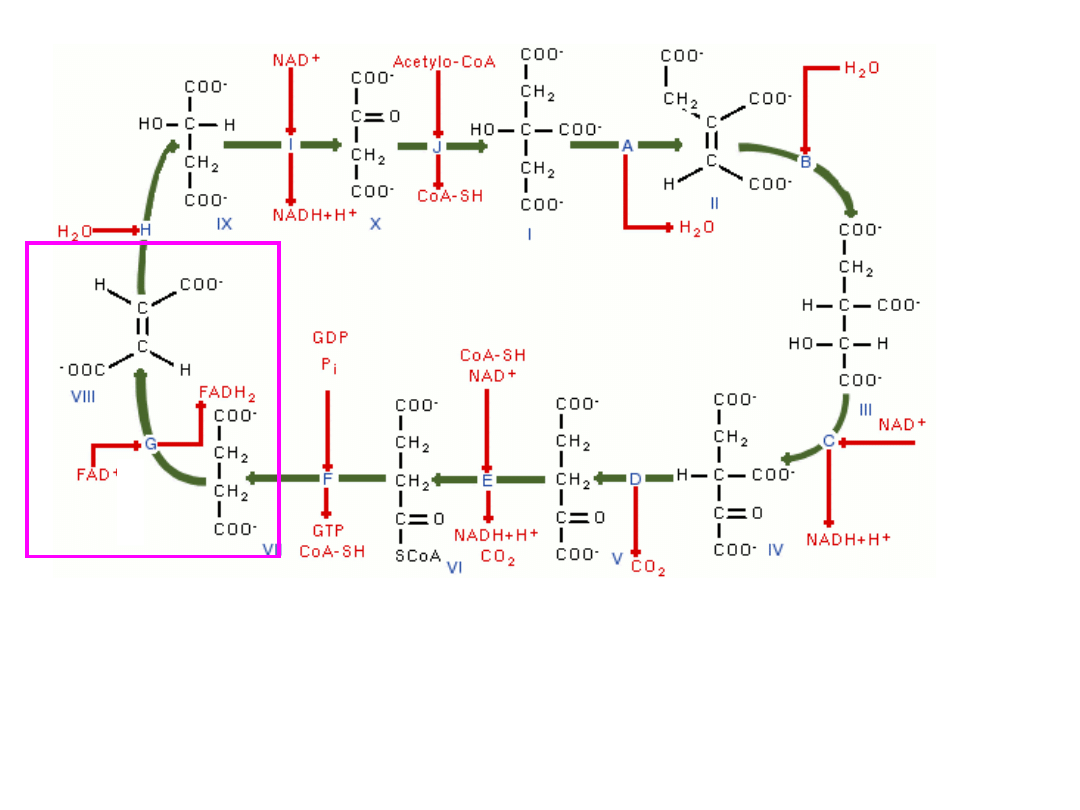
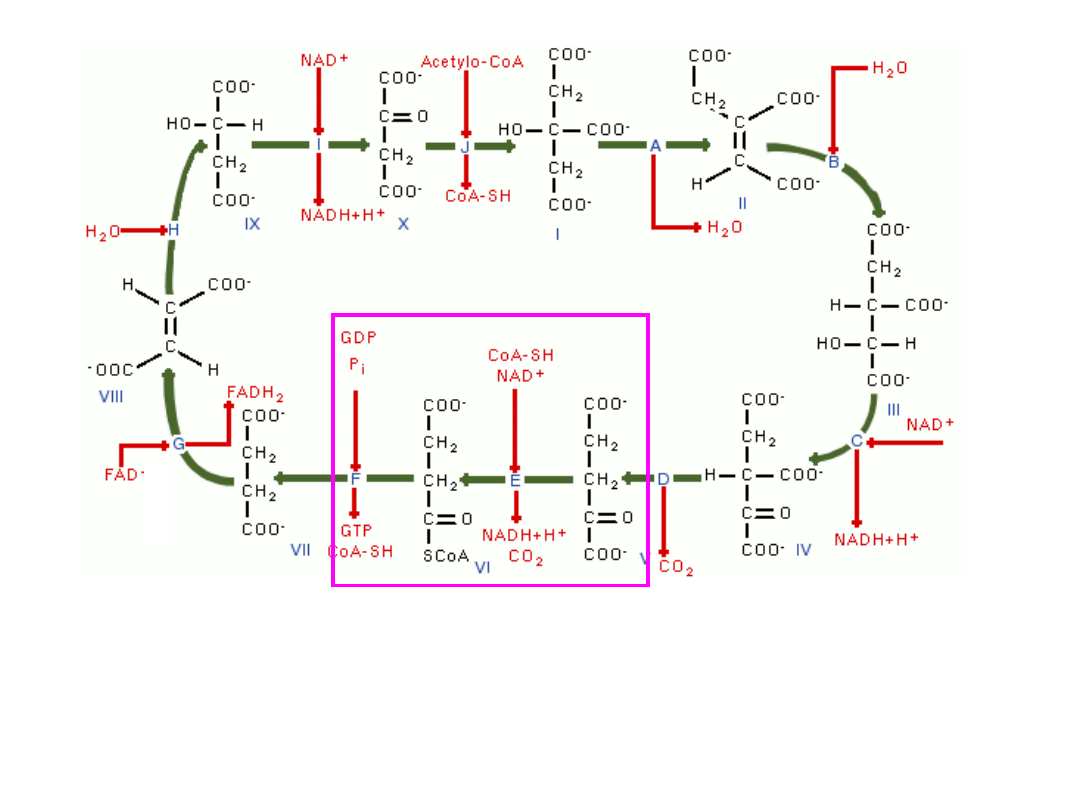
4-węglowy i 2-karboksylowy bursztynian (4C, 2COOH) ulega
dehydrogenacji i utlenieniu do 4-węglowego i 2-karboksylowego
fumaranu (4C, 2COOH) towarzyszy temu redukcja FAD do FADH
2
–
(
enzym-dehydrogenaza bursztynianowa
)
Bursztynian bursztynyloCoA -ketoglutaran
szczawiobursztynian
Jabłczan szczawiooctan
cytrynian
iz
o
cy
tr
y
n
ia
n
Fu
m
a
ra
n
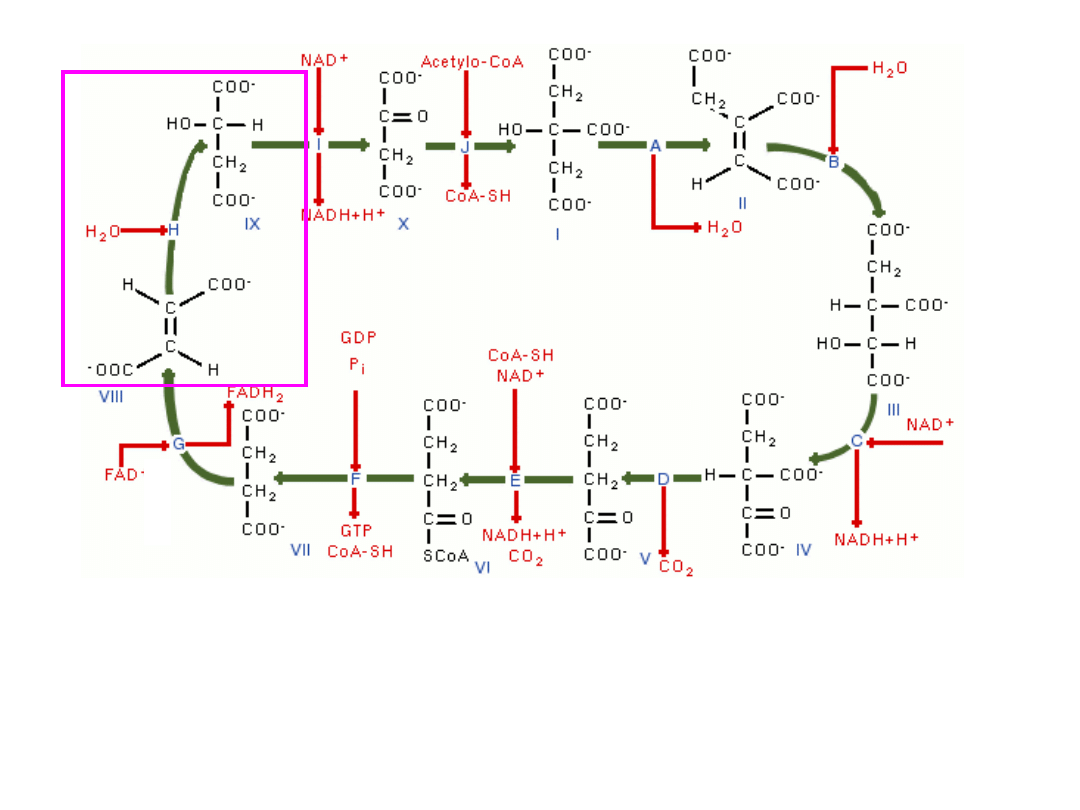
fumaran (4C, 2COOH) ulega hydratacji do jabłczanu (4C, 2COOH);
jabłczan jest bardzo podobny pod względem budowy chemicznej do
bursztynianu – zamiast jednego wodoru przy C2 (bursztynian)
występuje grupa hydroksylowa (jabłczan) –(
enzym-fumaraza
)
bursztynian bursztynyloCoA -ketoglutaran
szczawiobursztynian
Jabłczan szczawiooctan
cytrynian
iz
o
cy
tr
y
n
ia
n
Fu
m
a
ra
n
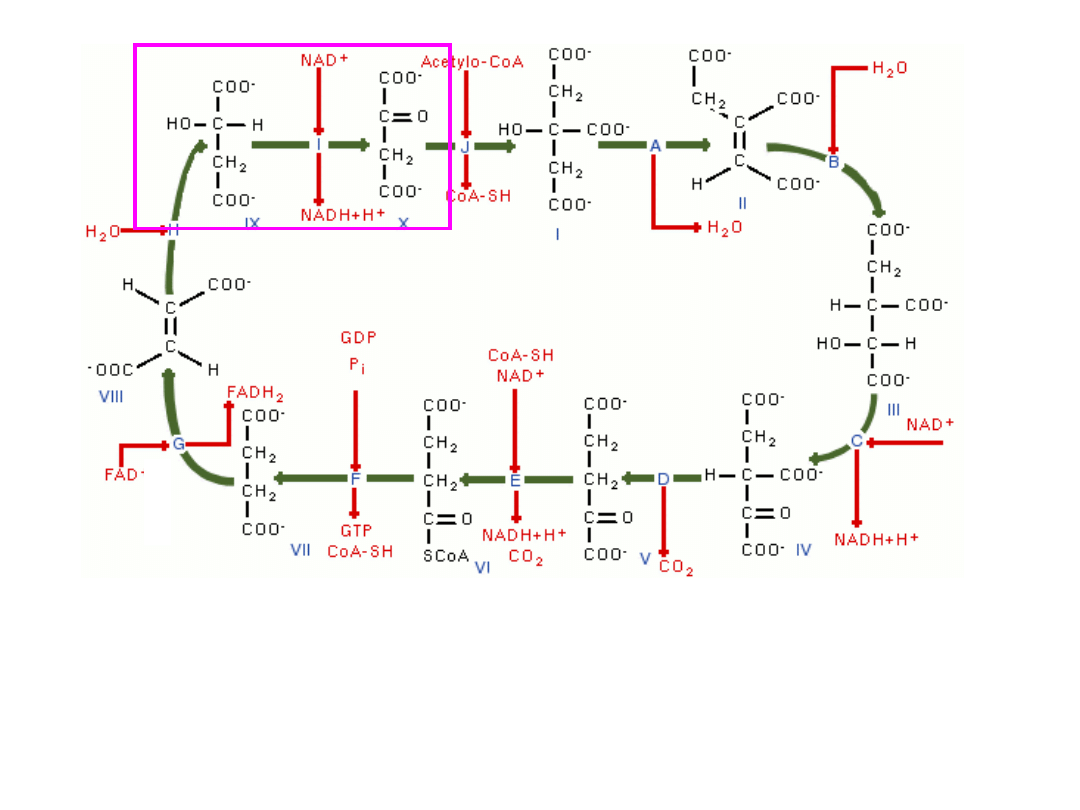
4-węglowy i 2-karboksylowy jabłczan (4C, 2COOH) ulega
dehydrogenacji do 4-węglowego i 2-karboksylowego szczawiooctanu
(4C, 2COOH), który w kolejnym etapie łącząc się z grupą acetylową
acetylo-CoA rozpoczyna kolejny cykl od nowa–(
enzym-
dehydrogenaza jabłczanowa
)
bursztynian bursztynyloCoA -ketoglutaran
szczawiobursztynian
Jabłczan szczawiooctan
cytrynian
iz
o
cy
tr
y
n
ia
n
Fu
m
a
ra
n

• Jedna cząsteczka NADH pozwala
wyprodukować 2,5 cząsteczki ATP
• jedna cząsteczka FADH
2
– 1,5 cząsteczki
ATP.

Bilans cyklu Krebsa
• W każdym cyklu powstają 3 cząsteczki NADH,
1cz. FADH
2
, 2 cz. CO
2
i jedna cząsteczka
ATP (pośrednio przez GTP).
• W glikolizie – procesie metabolizmu glukozy,
podstawowego surowca energetycznego
organizmu, z jednej cząsteczki glukozy powstają
dwa acetylo-CoA, czyli całkowite spalenie 1cz.
glukozy wymaga obiegu dwóch cykli Krebsa,
czyli bilans należy liczyć podwójnie

Bilans energetyczny
cyklu Krebsa II
Sposób wytwarzania
energii
Liczba utworzonych
cząsteczek ATP
Utlenianie 3 cząsteczek
NADH w łańcuchu
oddechowym
7,5 (3 x 2,5)
Utlenianie FADH2 w
łańcuchu oddechowym
1,5
1 Fosforylacja
substratowa
1
razem
10

Regulacja cyklu kwasu cytrynowego
Na szybkość cyklu ma wpływ:
dostępność substratów,
hamujące działanie nagromadzonych produktów i oparte na
mechanizmach sprzężenia zwrotnego allosteryczne hamowanie przez
następne intermediaty cyklu
. Najbardziej prawdopodobnymi miejscami regulacji są reakcje
nieodwracalne katalizowane przez następujące enzymy:
- syntazę cytrynianową (
hamowana przez cytrynian, a także przez ATP
)
- dehydrogenazę izocytrynianową (
hamowana przez NADH i ATP
, a
aktywowana przez ADP
)
- dehydrogenazę α-ketoglutaranową (
hamowana przez NADH i
bursztynylo-CoA
)
- dehydrogenazę pirogronianową (
hamowana przez NADH i acetylo-CoA
).

Wniosek 1
Cykl Krebsa przebiega szybciej, gdy poziom energii w
komórce jest niski
(duże stężenie ADP, a małe stężenie ATP i NADH),
zwalnia swój przebieg, gdy dochodzi do akumulacji ATP
(jak i również NADH, byrsztynylo-CoA oraz cytrynianu).

Wniosek 2
Cykl Krebsa zachodzi wyłącznie w
warunkach tlenowych. Tlen choć nie
bierze bezpośredniego udziału w cyklu
Krebsa to jest niezbędny do utleniania
FADH
2
i NADH
Document Outline
- Slide 1
- Slide 2
- Slide 3
- Slide 4
- Slide 5
- Slide 6
- Slide 7
- Slide 8
- Slide 9
- Slide 10
- Slide 11
- Slide 12
- Slide 13
- Slide 14
- Slide 15
- Slide 16
- Slide 17
- Slide 18
- Slide 19
- Slide 20
- Slide 21
- Slide 22
- Slide 23
- Slide 24
- Slide 25
- Slide 26
- Slide 27
- Slide 28
- Slide 29
- Slide 30
- Slide 31
- Slide 32
- Slide 33
Wyszukiwarka
Podobne podstrony:
ZW3 Radiata
więcej podobnych podstron