
ANATOMICZNE PODSTAWY
PAMIĘCI
Danuta M. Kowalska, Paweł Kuśmierek
Joanna Burkot
Uniwersytet Warszawski
Wydział Polonistyki
Warszawa 2008

III. ZABURZENIA PAMIĘCI
OBSERWOWANE PO
USZKODZENIACH PŁATÓW
SKRONIOWYCH
1.Amnezja następcza-
polega na braku zdolności uczenia
się i zapamiętywania nowych zdarzeń występujących po
zadziałaniu czynnika wywołującego zaburzenia pamięci
2. Amnezja wsteczna-
dotyczy pamięci zdarzeń z
przeszłości, przed pojawianiem się czynnika wywołującego
zaburzenia pamięci. Najbardziej zaburzona jest pamięć
wydarzeń z niedawnej przeszłości, najbardziej z przeszłości
odległej. Amnezja wsteczna występuje po dużych
uszkodzeniach płatów skroniowych, obejmujących ich
części boczne, jakie obserwuje się np. w wyniku zakażenia
wirusem opryszczki.

OBSERWACJE KLINICZNE PO
USZKODZENIU PŁATÓW
SKRONIOWYCH
• Pacjent H.M. cierpiał na lekooporna epilepsję- usunięto mu przyśrodkowe
części płatów skroniowych- kompleks ciała migdałowatego, znaczną część
formacji hipokampa, obszary kory znajdujące się w pobliżu tych struktur
• Wystąpiły:
• -zaburzenia pamięci deklaratywnej-pacjent utracił zdolność
zapamiętywania nowych informacji dotyczących bodźców i zdarzeń.
Zaburzenia nie były aż tak drastyczne bowiem ostatecznie H.M mógł nabywać
umiejętności ruchowe, uczyć się przy wielokrotnym powtarzaniu zdań.
Prawidłowo przebiegał też proces prymowania( zaburzone było prymowanie
słów które weszły do użycia po operacji; w pamięci semantycznej H.M nie
istnieją bowiem reprezentacje tych słów, które mogłyby zostać aktywowane
podczas reekspozycji) Świadczy to o zaburzeniu pamięci deklaratywnej przy
zachowaniu pamięci proceduralnej.
• Podsumowując wystąpiła więc amnezja następcza i wsteczna- (pacjent
nie pamiętał zdarzeń poprzedzających na 3 lata zabieg chirurgiczny).
Pamięć starszych zdarzeń jest utrzymana na dobrym poziomie.
• Naukowcy doszli do wniosku, iż zaburzenia pamięci nasilają się gdy
uszkodzone są szczególnie tylne części płatów skroniowych i przede
wszystkim hipokamp. Stwierdzili, że to on jest odpowiedzialny za tworzenie
się nowych śladów pamięciowych. Czy rzeczywiście tak jest?

2. Badania przeprowadzone na zwierzętach
Doniesienia o amnezji następczej, spowodowanej
interwencją chirurgiczną u ludzi zainspirowały serię
eksperymentów na zwierzętach, zwłaszcza na małpach.
Po pewnych doświadczeniach naukowcy sformowali
hipotezę, iż amnezja nie jest spowodowana uszkodzeniem
jedynie hipokampa (tylna część przyśrodkowych płatów
skroniowych). Wiąże się ona też z uszkodzeniem przednich
części płatów skroniowych a więc- ciała migdałowatego,
kory gruszkowatej, kory okołomigdałowatej, kory
węchowej, w skład której wchodzą obszary kory
okołowęchowej, rozciągającej się wzdłuż bruzdy węchowej i
położone przyśrodkowo od tej bruzdy obszary kory
śródwęchowej.
Dla poparcia niniejszej tezy przeprowadzono liczne
eksperymenty w USA, głownie na małpach.

a) Rozpoznawanie wzrokowe
Stosowany jest test: „dobierania nie według wzoru”(DNMS)-
polega na jednopróbowym uczeniu się i następnie rozpoznawaniu
obiektów wzrokowych. Dzięki temu testowi możemy stwierdzić
czy zwierze odróżnia znajome zjawiska sensoryczne od nowych.
Bada się pamięć rozpoznawczą.
Po opanowaniu testu DNMS małpy były poddawane zabiegom
chirurgicznym.
Grupa1-uszkodzone przednie części przyśrodkowych płatów
skroniowych(ciało migdałowate wraz z kora około migdałowatą i
przednimi częściami kory węchowej)
Grupa2- uszkodzono tylne części (hipokamp wraz z tylnymi
obszarami kory węchowej i korą przyhipokampalną)
Grupa3- łącznie przednie i tylne części przyśrodkowych płatów
skroniowych
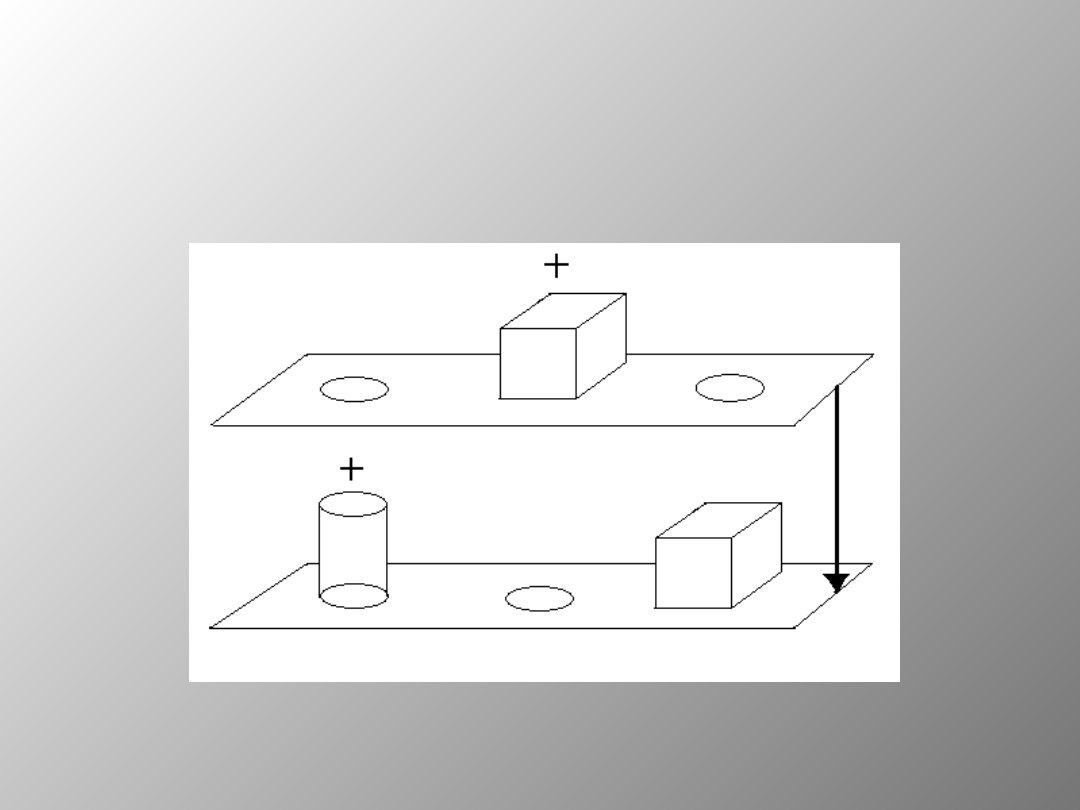
Etapy pojedynczej próby w
teście DNMS
Odroczenie
10-120s.
NABYWANIE
TESTOWANIE

Wzrokowa pamięć rozpoznawcza najsilniej zaburzona była w
grupie 3. Potwierdziło to tezę sformułowaną na podstawie badań
ludzi, iż rozległe uszkodzenia przyśrodkowych płatów skroniowych
silnie zaburzają pamięć rozpoznawczą. Wyniki tego eksperymentu
nie dały jednak ostatecznego dowodu określającego funkcje
poszczególnych struktur limbicznych płata skroniowego w pamięci
rozpoznawczej. Sformułowano następujące pytania:
1.Czy obserwowane zaburzenia pamięci są wynikiem uszkodzenia
ciała migdałowatego i hipokampa, czy też obszarów kory węchowej i
przyhipokampalnej?
2. Czy uszkodzenia ograniczone jedynie do ciała migdałowatego
bądź też hipokampa(bez naruszania obszarów korowych) w równym
stopniu zaburzają pamięć rozpoznawczą?
Dokonano więc eksperymentów w których uszkodzono niniejsze
okolice mózgu, bardziej wybiórczo.
Okazało się że uszkodzenie
ciała migdałowatego wraz z hipokampem nie narusza pamięci
rozpoznawczej. Zaburzenia pojawiają się dopiero gdy uszkodzi
się korę węchową. Okazało się więc, że pierwotne uszkodzenie
u małp ciała migdałowatego i hipokampa w rzeczywistości nie
wpłynęło na pogorszenie pamięci wzrokowej. Za utratę
pamięci wzrokowej były odpowiedzialne obszary kory
węchowej i połączenia nerwowe, uszkodzone przy okazji
uszkadzania hipokampa i ciała migdałowatego.
WNIOSKUJĄC: To kora węchowa i jej połączenia nerwowe
odpowiadają za pamięć wzrokową.

b) Rozpoznanie dotykowe
Stosowany jest test DMS różni się od DNMS tym, że w etapie
testowania nagradzany jest wybór znajomego
obiektu(pokazywanego na etapie nabywania) nie zaś obiektu
nowego. Zwierzęta testowano w ciemności podając do
rozpoznania obiekty różniące się kształtem, wielkością, fakturą
powierzchni. Dotykowa pamięć była dramatycznie zaburzona w
grupie 3, tak jak poprzednio. Dla normalnego funkcjonowania
pamięci dotykowej konieczna jest nieuszkodzona kora węchowa.
WNIOSKUJĄC: Za pamięć dotykową podobnie jak za
pamięć wzrokowa jest odpowiedzialna kora węchowa.

c) Rozpoznawanie słuchowe
Stosowany jest test DMS. Testowane są psy. W fazie nabywania z
głośnika umieszczonego w pozycji centralnej podawano pewien
dźwięk, następnie po 1,5-sekundowym odroczeniu, w fazie
testowania ten sam dźwięk podawano naprzemiennie z nowym,
nieznanym dźwiękiem przez dwa głośniki umieszczone po lewej i
prawej stronie zwierzęcia. Poprawna reakcja- skierowanie do
głośnika emitującego dźwięk znajomy- była nagradzana
pokarmem. W każdej próbie stosowano nową parę dźwięków. Psy
łatwo opanowały test.
Po uszkodzeniu tylko hipokampa pamięć słuchowa nie została
zaburzona-potwierdza to wcześniejszą tezę, że dla pamięci
słuchowej nie jest istotny hipokamp. Zaskakujący wynik dało
uszkodzenie kory węchowej. U małp takie uszkodzenie
powodowało zaburzenie pamięci rozpoznawczej wzrokowej, nie
zaburzało natomiast pamięci rozpoznawczej słuchowej u psów.
Brak udziału kory okołowęchowej w rozpoznawczej pamięci
słuchowej pozostaje zagadką.

d) Zapamiętywanie asocjacji między bodźcami
Używa się zestawu dwuwymiarowych bodźców wzrokowych,
zgodnie z zasadą:” Jeśli na etapie nabywania pojawi się bodziec A,
to po okresie odroczenia, na etapie testowania należy wybrać
bodziec X, a nie Y; natomiast jeśli pojawi się bodziec B, to należy
wybrać bodziec Y, a nie X. Zwierzęta uczyły się 10 takich zadań
pamięciowych opartych na asocjacji dwóch bodźców. Zwierzęta z
grupy 3 miały dramatycznie zaburzoną pamięć asocjacyjną. U tych,
u których usunięto tylko hipokamp i obszary korowe, lub tylko ciało
migdałowate i obszary korowe nie wykryto zmian. Potrafiły wykonać
zadanie.
WNIOSKUJĄC: To kora węchowa a nie ciało
migdałowate lub hipokamp jest istotna dla uczenia się i
zapamiętywania nowych asocjacji między bodźcami.

e) Pamięć przestrzenna
Zdrowe zwierzęta testowane są przy pomocy aparatu Nenckiego
.Po wykonaniu zadania- obustronne wybiórcze uszkodzenie
hipokampa. Po operacji testowano pamięć przestrzenną z
odroczeniem 10s. stopniowo wydłużanym do 120s. Dodawano też
zakłócenia- podczas odroczenia stosowano dodatkowy bodziec
odwracający uwagę zwierzęcia i zmuszający go do zmiany pozycji
ciała. W ten sposób zapobiegano rozwiązaniu testu przez
utrzymanie podczas odroczenia pozycji skierowanej do uprzednio
sygnalizowanego karmnika. Grupa z uszkodzonym hipokampem
wykazywała znaczne zaburzenia, grupa z uszkodzeniami przednich
płatów skroniowych nie różniła się istotnie od zwierząt
normalnych. Wyniki potwierdziły tezę.
WNIOSKUJĄC: To hipokamp jest odpowiedzialny za
pamięć przestrzenną. Naruszenie kory węchowej nie
powodowało zmian. Takie same wyniki uzyskali
naukowcy już wcześniej, podczas badań nad
szczurami.
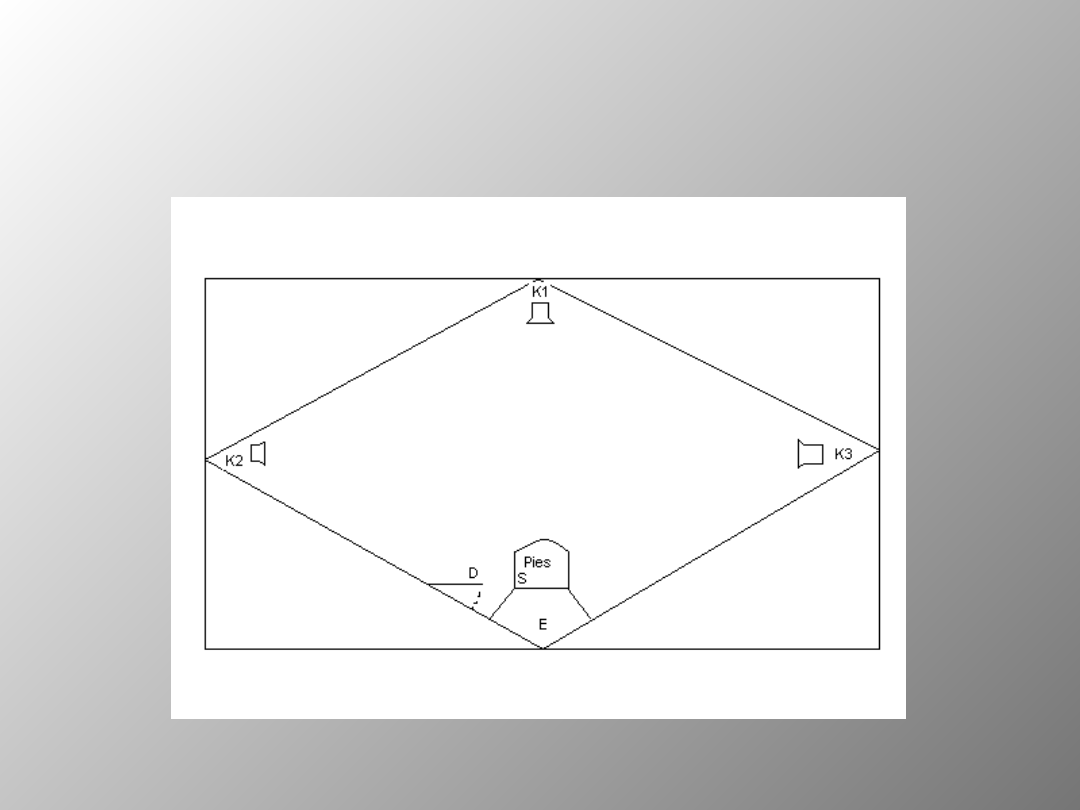
Schemat przedstawiający tzw.
Aparat Nenckiego

IV. ZABURZENIA PAMIĘCI PO
USZKODZENIACH
MIĘDZYMÓZGOWIA
Zaburzenia pamięci występują nie tylko w wyniku uszkodzenia
płatów skroniowych ale też w wyniku uszkodzenia
międzymózgowia- szczególnie ciał suteczkowatych i
przyśrodkowej części wzgórza. Uszkodzenia przyśrodkowej
części wzgórza były powodowane urazami mechanicznymi np.
guzem mózgu czy zespołem Korsakowa, będącym następstwem
alkoholizmu. Silne połączenia między przyśrodkowymi częściami
płatów skroniowych i przyśrodkowymi częściami wzgórza
sugerowały, że oba te rejony wchodzą w skład neuronalnych
substratów pamięci. Potwierdziły to badania na małpach. Przy
uszkodzeniu przedniej i tylnej części wzgórza małpy miały dużo
większe trudności z wykonywaniem zadań niż w przypadku
uszkodzenia tylko jednego elementu.
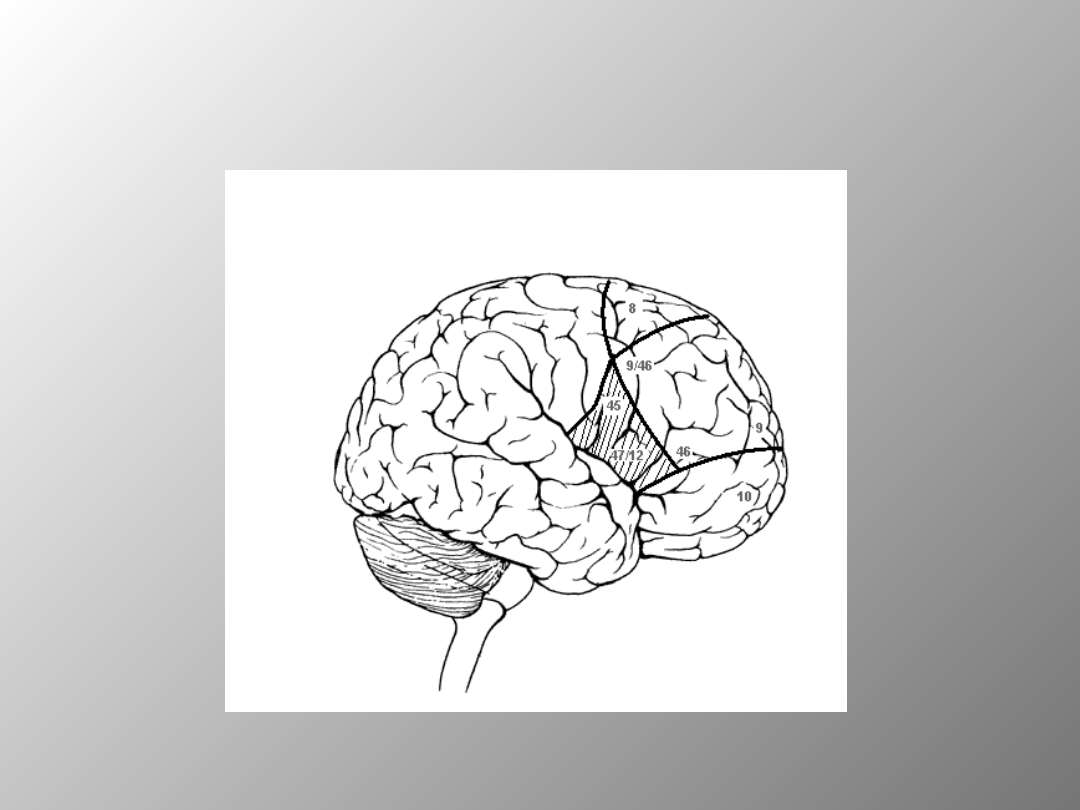
V. ROLA KORY PRZEDCZOŁOWEJ
W PAMIĘCI
Schemat bocznej powierzchni mózgu człowieka z zaznaczeniem obszarów kory
przedczołowej
10-biegun czołowy;8-tylna kora przedczołowa; 9- środkowa część grzbietowo-bocznej kory
przedczołowej; 45 oraz 47/12- środkowa część brzuszno-bocznej kory przedczołowej

a)
Obszary bocznej kory przedczołowej a rodzaj
przetwarzania informacji
Wcześniejsze badania wykazały, że obszary
bocznej kory przedczołowej a zwłaszcza obszary
grzbietowo- boczne są odpowiedzialne za pamięć
krótkotrwałą.
Udowodniono to poprzez badania na małpach. W
obecności małpy umieszczono nagrodę w jednym z
dwóch bocznych karmników znajdujących się na
tacce testowej. Następnie oba karmniki zakryto
identycznymi płytkami oraz zasłonięto całą tackę
testową. Po okresie odroczenia odsłonięto tackę
testową, aby zwierze mogło dokonać wyboru.
Poprawnym wyborem było odsłonięcie płytki pod
którą znajdowała się nagroda. Z kolei w teście
odroczonej alternacji nagroda była umieszczana
naprzemiennie, raz w lewym, raz w prawym
karmniku. Aby odnaleźć nagrodę zwierze musiało
pamiętać po której stronie znajdowała się ona w
poprzedniej próbie. W założeniu oba doświadczenia
były oparte na pamięci przestrzennej. Zaburzenia w
wykonaniu tych zadań pojawiły się w grupie małp z
uszkodzoną grzbietowo- boczna korą przedczołową.
Badania wykazały że punktem krytycznym jest bruzda
główna.
WNIOSKUJĄC: grzbietowo- boczna korą
przedczołowa i bruzda główna są
odpowiedzialne za pamięć krótkotrwałą, a
szczególnie krótkotrwałą pamięć
przestrzenną.

2.Rola brzusznych i przyśrodkowych obszarów kory
przedczołowej w pamięci
Są to obszary mózgu istotne dla formowania pamięci
deklaratywnej. Potwierdziły to nie tylko badania na małpach ale
tez na ludziach. W warszawskim laboratorium A. Grabowskiej
wykazano krótkotrwałe zaburzenie pamięci obiektów (nie ich
lokalizacji) po niewielkich uszkodzeniach tylnej części zakrętu
prostego. Z kolei w innym, amerykańskim laboratorium
obserwowano zaburzenie pamięci obiektów oraz pamięci
przestrzennej po uszkodzeniu tylnej części brzuszno-
przyśrodkowej kory przedczołowej. Potwierdzeniem tych
wyników są badania na ludziach, które wskazują na aktywację
obszarów dolnej kory przedczołowej podczas przypominania
faktów i zdarzeń. Zaproponowano również rolę brzuszno-
przyśrodkowej kory przedczołowej w asocjacji faktów i sytuacji za
stanami emocjonalnymi wywołanymi uprzednio przez podobne
fakty i sytuacje. Konsekwencją tego jest zaburzenie procesu
podejmowania decyzji u pacjentów z uszkodzeniami tych części
kory z powodu niezdolności do odczucia emocjonalnego
zabarwienia konsekwencji podejmowanych decyzji.
WNIOSKUJĄC: Kora przedczołowa odpowiedzialna
jest nie tylko za pamięć krótkotrwałą ale również
pamięć długotrwałą.

VI. ROLA ZWOJÓW PODSTAWY W
PAMIĘCI PROCEDURALNEJ
Zwoje podstawy- jądra leżące pod płaszczem korowym, w
skład których wchodzą: jądro ogniste, skorupa, gałka blada,
jądra niskowzgórzowate i istota czarna. Wśród badaczy
istnieje wiele poglądów na to za co odpowiedzialne są zwoje.
Pacjenci chorzy na amnezję wykazują silne zaburzenia w
zdolności uczenia się nowych faktów i zdarzeń opartych na
pamięci deklaratywnej, lecz formowanie trwałych odruchów i
umiejętności związanych z pamięcią proceduralną przebiega
u nich bez trudności. Natomiast pacjenci cierpiący na
chorobą Parkinsona i Huntingtona, u których występują
degeneracje w zwojach podstawy, przejawiają odwrotny
wzorzec zaburzeń. Mogą oni rozwiązywać zadania oparte na
pamięci deklaratywnej, przy jednoczesnym silnym zaburzeniu
uczucia opartego na pamięci proceduralnej.

Przykładem może być test uczenia probabilistycznego. –
Stosowano 4 abstrakcyjne rysunki, z których każdy był z pewnym
(nieznanym osobom badanym) prawdopodobieństwem związany z
wynikiem „słońce” lub „deszcz”. W kolejnych próbach badanym
osobom prezentowano jeden, albo jednocześnie 2 lub 3 z czterech
rysunków, na podstawie których starały się „przewidzieć pogodę”.
Ponieważ związek każdego z rysunków z wynikiem był tylko
statystyczny i dodatkowo skomplikowany przez prezentacje kilku
rysunków w jednej próbie, wykrycie go w sposób świadomy
(deklaratywny) było bardzo trudne. Osoby zdrowe oraz pacjenci z
amnezją będącą wynikiem uszkodzenia przyśrodkowych płatów
skroniowych uczyli się „przewidywać pogodę” metodą prób i błędów:
ich poziom wykonania wzrastał od 50% na początku treningu do 65%
po 50 próbach. Nie mieli jednak świadomości, że uczyli się związku
rysunków z pogodą. Zarówno pacjenci z choroba Huntingtona, jak i ci
z chorobą Parkinsona nie byli w stanie osiągnąć poziomu wyższego
niż 53%.Wiele innych badań na małpach i szczurach potwierdziły
znaczenie zwojów podstawy w pamięci proceduralnej. Badania u
ludzi z wykorzystaniem technik obrazowania (fMRI) wykazują
aktywację jądra ogoniastego w zadaniach takich jak np. uczenie się
czytania tekstu w lustrze. Chociaż nabycie odruchów lub też
umiejętności może się wydawać bardziej prymitywne od świadomego
zapamiętywania, to jednak neuronowe podłoże tych procesów nie jest
zorganizowane w sposób prosty
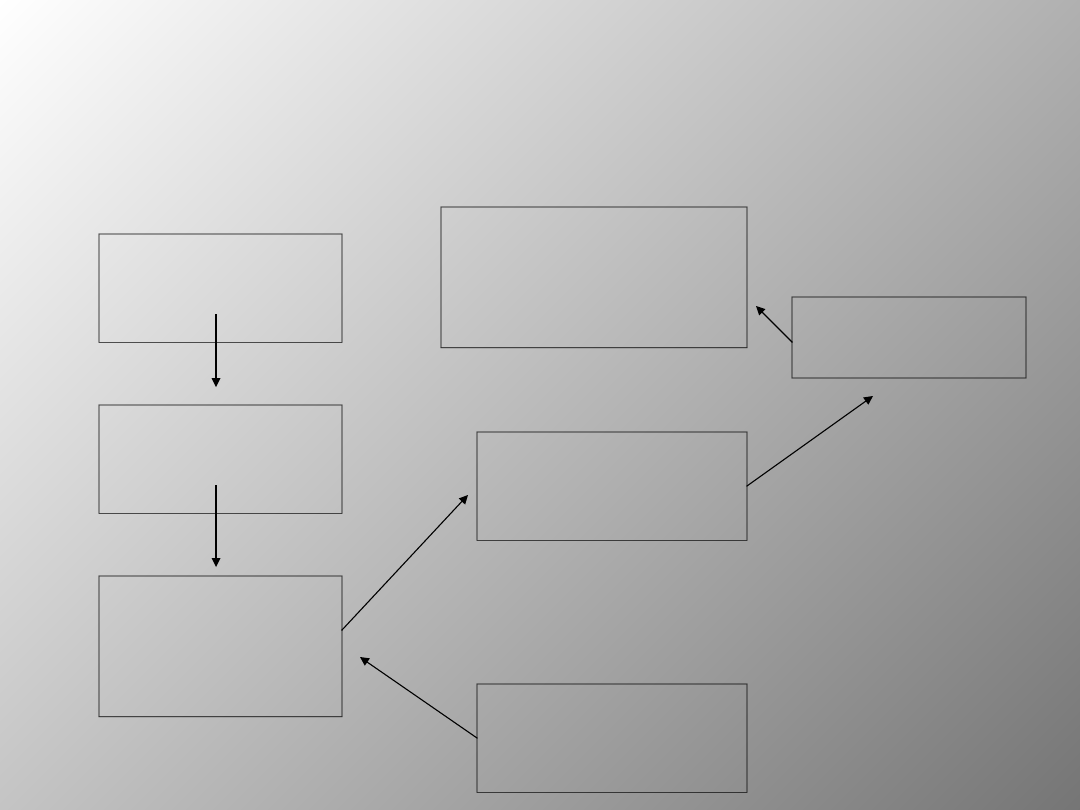
Schemat przedstawiający
neuronowe podłoże pamięci
proceduralnej
Pierwszorzędne
obszary
czuciowe
Obszary
czuciowe
wyższego rzędu
Tylna część
jądra
ogoniastego i
skorupa
Gałka blada i
część siatkowata
istoty czarnej
Część zbita istoty
czarnej, system
dopaminergiczny
Brzuszne jądra
wzgórza
Kora przedruchowa i
dodatkowe pole
ruchowe kory
czołowej

Badania sugerują, iż różne formy warunkowania mogą mieć
odmienne podłoże neuronowe, ponadto informacje o
nabytych umiejętnościach mogą być przechowywane w
różnych częściach mózgu. Wiele danych wskazuje na istotna
rolę zwojów podstawy oraz ich połączeń z jądrami brzusznymi
wzgórza i następnie z czołowymi obszarami korowymi w
warunkowaniu reakcji instrumentalnych. Wykazano również
że u podstaw klasycznej warunkowej reakcji mrugania leżą
połączenia zwojów podstawy z móżdżkiem, natomiast
warunkowanie klasycznych reakcji autonomicznych takich jak
częstotliwość czynności serca, zależy od połączeń z ciałem
migdałowatym.
Wielu badaczy uważa, że zwoje podstawy zawierają całą
maszynerię odpowiedzialną za pamięć proceduralną.
Najnowsze badania sugerują, że zwoje podstawy mogą
uczestniczyć zarówno w formowaniu pamięci
proceduralnej i deklaratywnej. Inne badania wskazują na
pośrednicząca rolę kory czołowej, łącznie z wysoko
zorganizowanymi obszarami kory przedczołowej, w
wytwarzaniu trwałych odruchów.
WNIOSKUJĄC: systemy neuronowe stanowiące
substraty dla pamięci deklaratywnej i
proceduralnej ściśle współpracują ze sobą w
kształtowaniu zachowań u ludzi i u zwierząt.

VII. PODSUMOWANIE
W pamięci deklaratywnej uczestniczą struktury
przyśrodkowej części płata skroniowego, przyśrodkowe
części wzgórza oraz kora przedczołowa.
Kora węchowa jest związana z rozpoznawaniem
wzrokowym i dotykowym oraz z pamięcią asocjacyjną.
Hipokamp jest istotny dla wykonania zadań opartych na
pamięci przestrzennej, na pamięci epizodycznej.
Ciało migdałowate istotne jest w pamięci emocjonalnej i
kojarzeniu bodźców ze wzmocnieniem.
Grzbietowo-boczna kora przedczołowa odgrywa dużą rolę
w wykonaniu zadań opartych na pamięci operacyjnej,
natomiast brzuszna część kory przedczołowej jest istotna
dla pamięci rozpoznawczej.
Pamięć proceduralna jest związana z funkcją zwojów
podstawy oraz ich interakcją ze wzgórzem i korą
przedczołową. W procesach warunkowania zwoje podstawy
współpracują również z wyspecjalizowanymi obszarami
korowymi istotnymi dla pamięci deklaratywnej.

Dziękuję
Document Outline
- Slide 1
- Slide 2
- Slide 3
- Slide 4
- Slide 5
- Slide 6
- Slide 7
- Slide 8
- Slide 9
- Slide 10
- Slide 11
- Slide 12
- Slide 13
- Slide 14
- Slide 15
- Slide 16
- Slide 17
- Slide 18
- Slide 19
- Slide 20
- Slide 21
- Slide 22
Wyszukiwarka
Podobne podstrony:
mozg www prezentacje org
MÓZG MĘŻCZYZNY PREZENTACJA
prezentacja finanse ludnosci
prezentacja mikro Kubska 2
Religia Mezopotamii prezentacja
Prezentacja konsument ostateczna
Strategie marketingowe prezentacje wykład
motumbo www prezentacje org
lab5 prezentacja
Prezentacja 18
Materialy pomocnicze prezentacja maturalna
Prezentacja na seminarium
Lato prezentacja 3
Prezentacja1
więcej podobnych podstron