AKTYWNOŚĆ
BIOLOGICZNA
NEUROMEDIATORÓW W
OUN
Część 1
UKŁAD DOPAMINERGICZNY
I JEGO STRUKTURY

UKŁADY FUNKCJONALNE
MÓZGU
Zachowanie człowieka i zwierząt (ssaków)
jest wypadkową działania trzech wielkich
układów funkcjonalnych mózgu:
UKŁADU POBUDZENIA (arousal)
UKŁADU NAGRODY (reward)
UKŁADU POZNAWCZEGO (cognition)
Powyższe układy stanowią system sterujący
zachowaniem człowieka i zwierząt.

UKŁAD POBUDZENIA
Odpowiada za stany czuwania
i snu.
Składa się z trzech podsystemów:
Pobudzenia ogólnego
regulacja pobudliwości OUN
Pobudzenia ukierunkowanego
wykazującego związek z motywacją
i emocjami
Regulacji pobudzenia
obwodowego
oddziaływanie
CNS na ciało

UKŁAD POBUDZENIA
Podsystem pobudzenia ukierunkowanego
Dostarcza jakości emocjonalnych
zróżnicowanych, często
przeciwwstawnych doznań
lęk, ciekawość, gniew,
przyjemność, wstręt, złość, zadowolenie
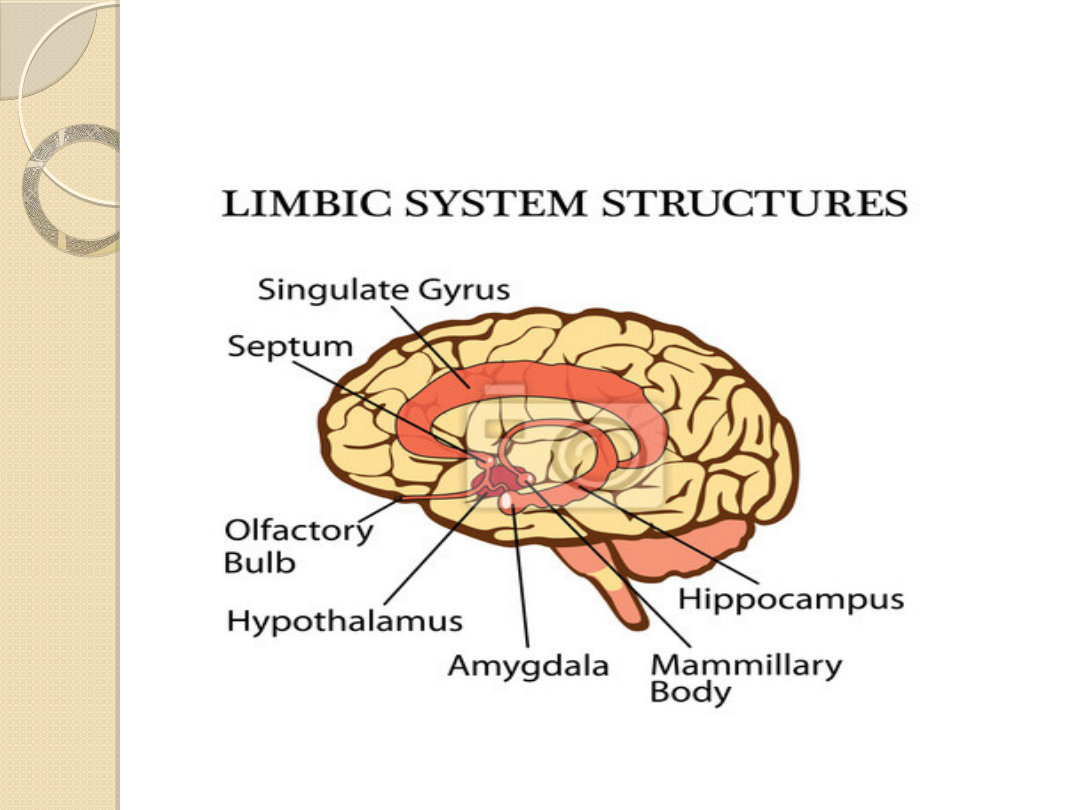
Układ limbiczny stanowi podłoże anatomiczne
podsystemu pobudzenia ukierunkowanego.
W układzie limbicznym jest zlokalizowany układ
nagrody
.
Podsystem pobudzenia ukierunkowanego odpowiada za
mechanizm wybierania odpowiednich celów, inicjuje
zachowania potrzebne do ich osiągnięcia i sygnalizuje, że
zostały osiągnięte.
Cele korzystne:
przeżycie osobnika czy sukces
reprodukcyjny
zachowania wzmacniane nagrodą
Cele niekorzystne
hamowanie przez karę.

UKŁAD NAGRODY
Układ nagrody jest systemem oceniającym i
pozwalającym na odczuwanie przyjemności i
kary za odpowiednie rodzaje zachowań.
Został odkryty na początku lat 50-tych XX wieku
Po zmodyfikowaniu metod eksperymentalnych
przez Oldsa i Milnera (J.Comp. Physiol. Psychol,
1954, 47, 419-427) po drażnieniu odpowiednich
obszarów mózgu szczura opisano i opracowano
„mapę obszarów przyjemności w mózgu”.
Drażnienie miejsc powodujących przyjemność
ogólne dobre samopoczucie
Drażnienie miejsc awersyjnych
lęk, poczucie
niebezpieczeństwa, opuszczenie

UKŁAD NAGRODY
UKŁAD NAGRODY

UKŁAD NAGRODY
Układ nagrody jest zaangażowany we
wszystkie zasadnicze zachowania człowieka
czy zwierząt:
Pobieranie pokarmu
Pobieranie wody
Aktywność seksualna
Agresja
Odczuwanie przyjemności
Zaburzenia układu nagrody:
Anomalie behawioralne
Bulimia, anoreksja
Zaburzenia psychoseksualne
Nadmierna agresja

UKŁAD NAGRODY
Układ nagrody określany jest także jako
ośrodek przyjemności.
Jest to zbiór struktur w mózgu, które
regulują kwestie związane z motywacją i
kontrolą zachowania.
Mechanizm ten powstał w wyniku ewolucji i
ma na celu wyrobienie w organiźmie
nawyków, które zwiększają
prawdopodobieństwo występowania
korzystnych zachowań.
Zasada:
coś sprawia przyjemność to staje
się to doskonałym motywatorem !!!!

UKŁAD NAGRODY
W przypadku wystąpienia korzystnego dla
jednostki zachowania uwalnia się dopaminia
(hormon szczęścia)
, która pobudzając
receptory w strukturach układu nagrody i
wywołuje u osobnika poczucie przyjemności.
Wystąpienie poczucia przyjemności
powoduje odczucie spełnienia i zadowolenia.
Układ nagrody jest aktywowany w sytuacjach
zaspokajania podstawowych popędów
(jedzenie, zachowania seksualne)
oraz w
trakcie wykonywania czynności nie
popędowych odczuwanych jako przyjemne.

UKŁAD NAGRODY
W warunkach fizjologicznych na
struktury obejmujące układ nagrody
OUN działają neuroprzekaźniki.
W pobudzenie układu nagrody
zaangażowane są:
1.
DOPAMINA
2.
SEROTONINA
3.
ENDOGENNE PEPTYDY OPIOIDOWE
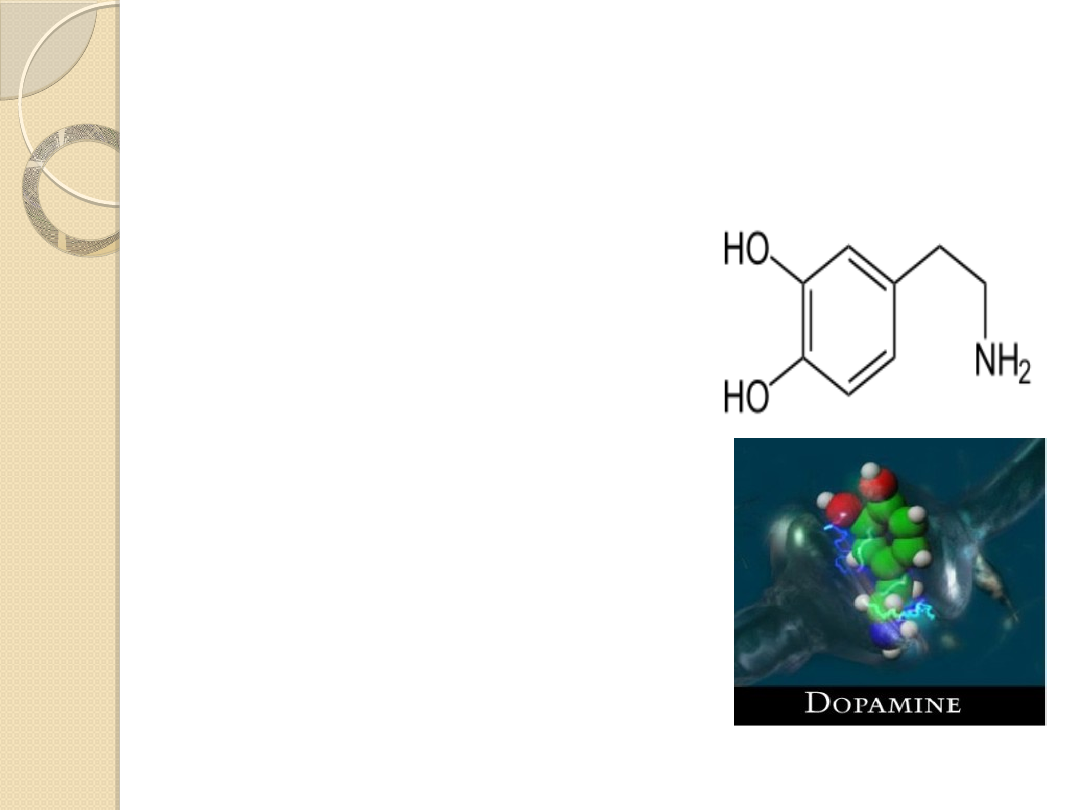
UKŁAD DOPAMINERGICZNY
Dopamina (C
8
H
11
NO
2
) pełni różne
funkcje w zależności od miejsca
działania w OUN:
Układ pozapiramidowy
napęd
ruchowy, koordynacja, napięcie
mięśni szkieletowych
Układ limbiczny
procesy
emocjonalne, wyższe czynności
psychiczne.
Podwzgórze
regulacja uwalniania
hormonów
(prolaktyna, ACTH,
gonadotropiny)

UKŁAD DOPAMINERGICZY
Dopamina (DA) jest aminą katecholową.
Jest prekursorem w biosyntezie
noradrenaliny i adrenaliny.
Jest samodzielnym neuroprzekaźnikiem w
OUN, ale też pełni rolę w czynności
obwodowego układu nerwowego.
Dopamina wywiera działanie poprzez
receptory dopaminergiczne:
grupa D1-like
(D1 i D5) oraz
grupa D2-like
(D2, D3 i D4).
Receptory dla DA należą do receptorów
metabotropowych, związanych funkcjonalnie
z białkami regulacyjnymi G.
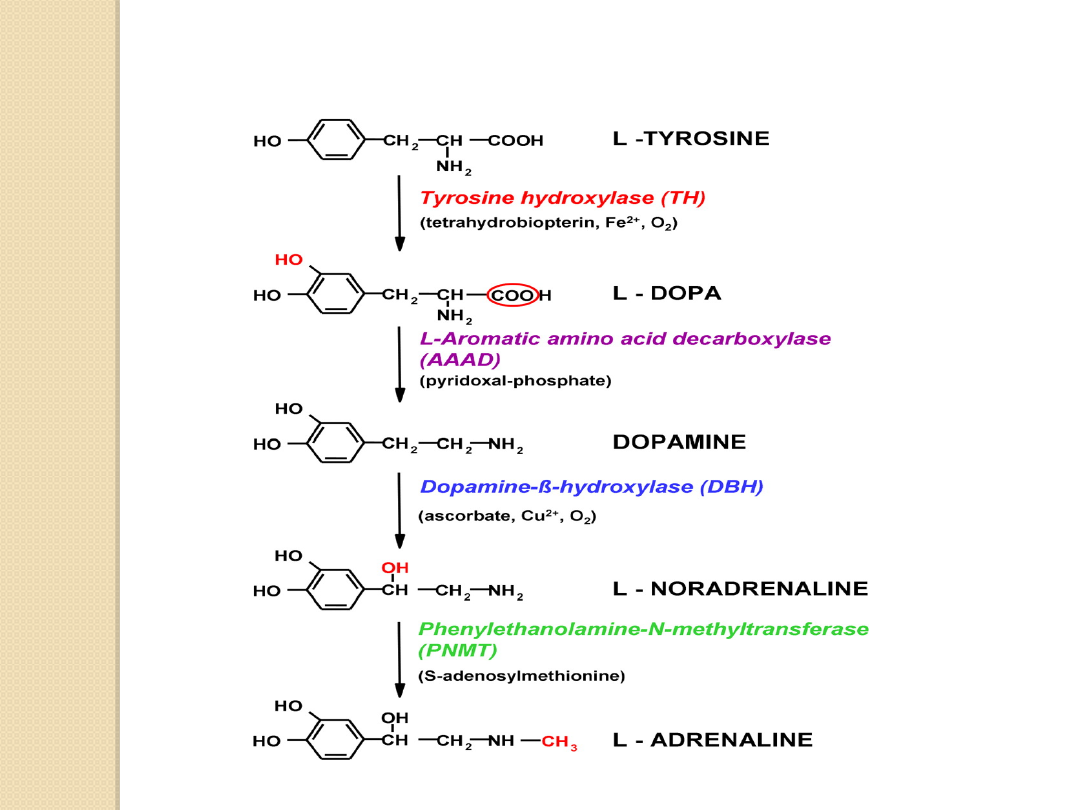
BIOSYNTEZA AMIN KATECHOLOWYCH
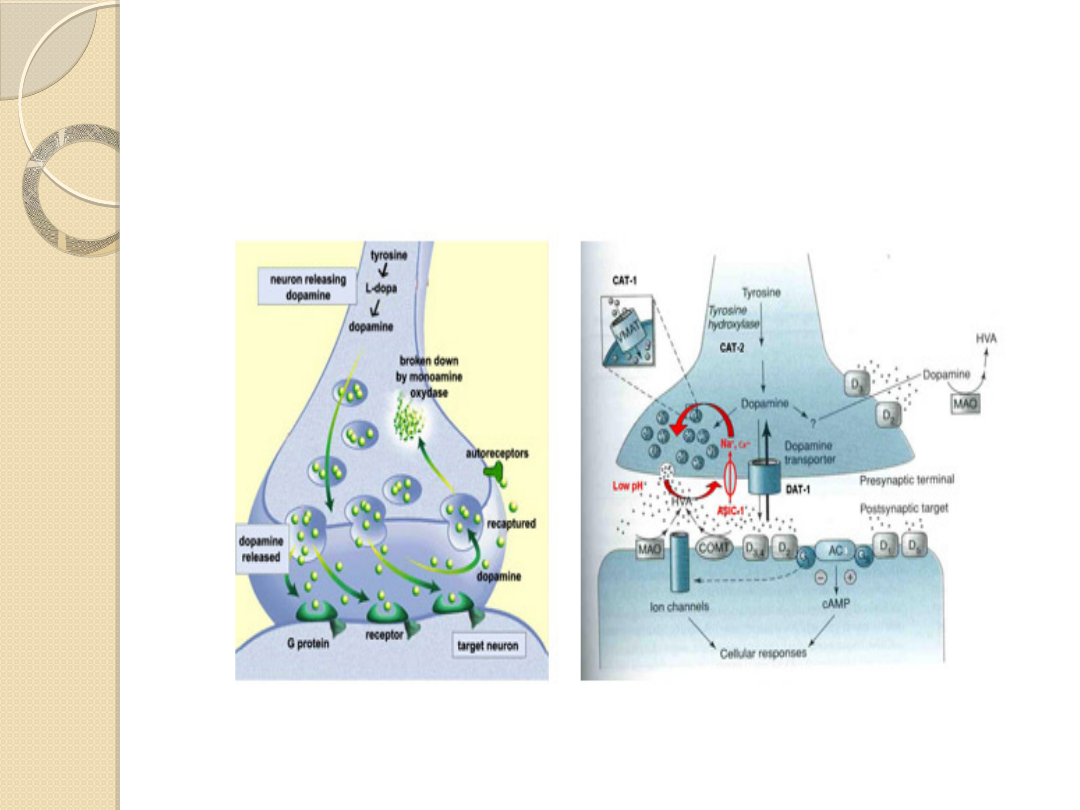
SYNAPSA DOPAMINERGICZNA
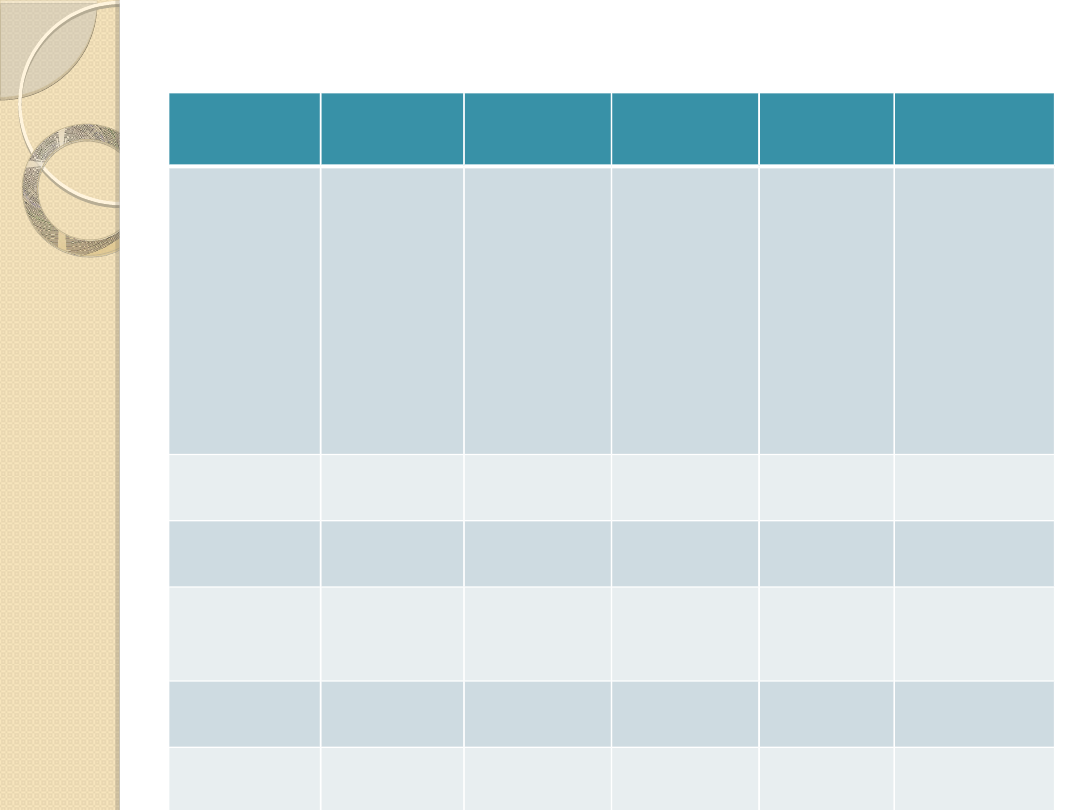
Układy efektorowe receptorów dopaminergicznych.
Recepto
r
D1
D5
D2
D3
D4
Lokalizacj
a
Prążkowie
, nowa
kora w
tym cześć
przed-
czołowa
Hipokamp,
kora
węchowa,
wzgórze
Prążkowie
,
hipokamp
, kora
węchowa,
okolica
migdałow
ata
Prążkowie
zwłaszcza
cześć
brzuszna,
móżdżek,
Kora
przedczoło
wa,
hipokamp,
okolica
migdałowa
ta, rdzeń
przedłużon
y, brak w
prązkowiu
Auto-
receptor
---
----
tak
tak
----
Cyklaza
adenylowa
+
+
-
-
-
Fosfolipaz
a C (IP3,
DAG)
+
-
-
-
-
Kanały
wapniowe
+
-
-
-
-
Kanały
potasowe
?
?
+
?
?

Receptory DA
Receptory grupy D1 aktywują cyklazę adenylową i
zwiększają aktywność neuronów OUN
Receptory grupy D2 wywierają efekt przeciwwstawny.
Pobudzenie motoryczne i emocjonalne (agresja)
oraz zachowania steoretypowe związane są z
pobudzeniem receptorów D2<D1.
Receptor D2 odgrywa istotna rolę w pobudzeniu
układu nagrody
(oczekiwanie na przyjemność)
Receptory D3 związane są tak jak receptory D2 z
regulacją procesów emocjonalnych.
Receptory D2 i D3 mogą być autoreceptorami
hamującymi uwalnianie dopaminy i aktywność
neuronów dopaminergicznych.
Receptor D2 w przysadce związany jest z
hamowaniem uwalniania prolaktyny.

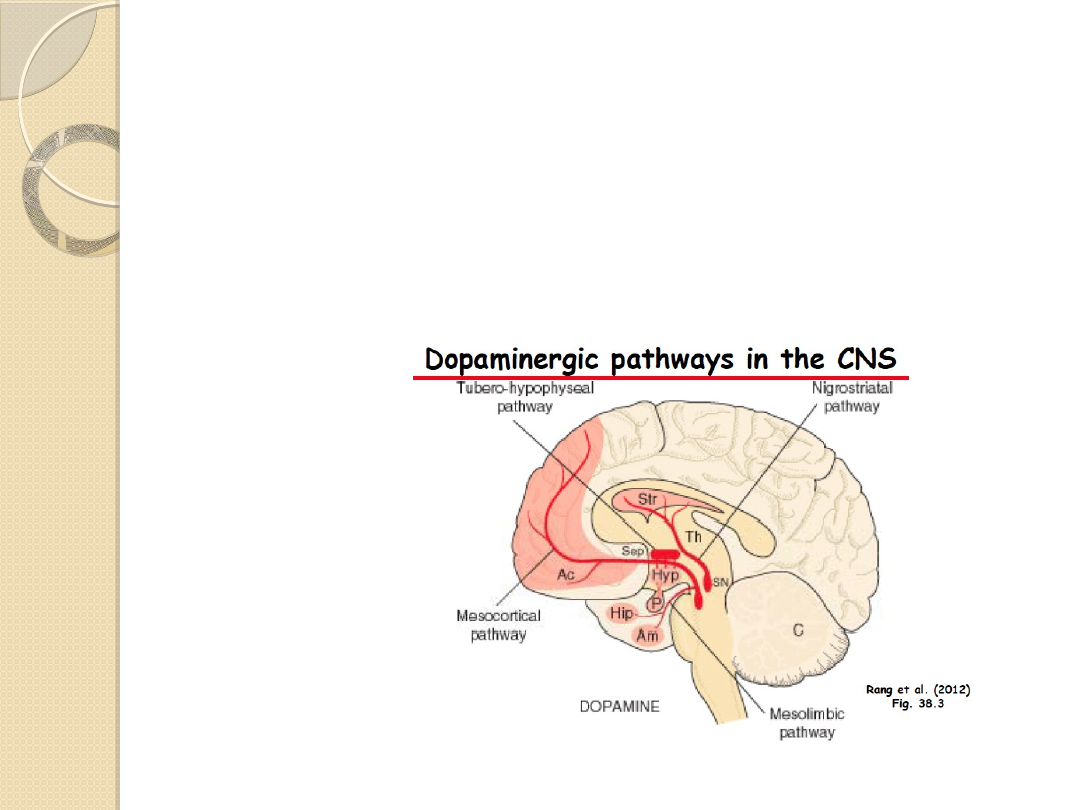
SZLAKI DOPAMINERGICZNE W
MÓZGU.
Szlak nigro-striatalny (czarno-
prążkowiowy)
tworzy włókna osiowe komórek
dopaminergicznych znajdujących się w istocie
czarnej (substantia nigra), łączące ten obszar
z prążkowiem (striatum), szczególnie z
jądrem ogoniastym (nucleus caudatus).
Szlak ten jest związany z regulacją
procesów ruchowych
napędu ruchowego,
koordynacji i napięcia mięśniowego
układ czynnościowy regulujący funkcje
ruchowe tzw. układ pozapiramidowy

Zaburzenia czynności tego układu prowadzą do
poważnych zmian w napięciu mięśni
szkieletowych, aktywności ruchowej i koordynacji
ruchów.
W chorobie Parkinsona (
drżenia mięśniowe,
upośledzenie napędu ruchowego i wzmożone
napięcie mięśni)
występują zmiany
zwyrodnieniowe szlaku nigro-striatalnego i
zmniejszenie stężenia dopaminy w prążkowiu.
Leki, których mechanizm działania związany jest z
antagonizmem w stosunku do receptorów D2
(neuroleptyki, metoklopramid) mogą wywierać
działania niepożądane w postaci objawów
Parkinsona!!!!!

Szlak mezolimbiczny i szlak mezokortykalny
układ mezokortykolimbiczny
łączy
dopaminergiczne komórki śródmózgowia ze
strukturami limbicznymi, głównie jądrami
przegrody oraz polami korowymi.
Szlak ten jest odpowiedzialny za procesy
emocjonalne ( agresji), za procesy związane z
układem nagrody i czynności motywacyjne ,
ale też odgrywa istotną rolę w regulacji procesów
ruchowych.
Szlak mezokortykalny uczestniczy również w
reakcjach organizmu na bodźce stresowe.

Szlak guzowo-lejkowaty
tworzy grupa krótkich
neuronów biegnących z jądra łukowatego (nucleus
arcuatus) podwzgórza do wyniosłości pośrodkowej i
przysadki.
Szlak ten jest odpowiedzialny za regulację czynności
wydzielniczej przysadki.
Dopamina hamuje uwalnianie prolaktyny i ACTH a
pobudza wydzielanie hormonu wzrostu oraz wpływa
regulująco na wydzielanie gonadotropin
przysadkowych.
Receptory dopaminergiczne w obrębie puszki są związane
z odruchem wymiotnym
leki blokujące receptory DA
wykazują działanie przeciwwymiotne.
Neurony dopaminergiczne wstawkowe występują w części
wzrokowej kory, siatkówce oka i rdzeniu przedłużonym
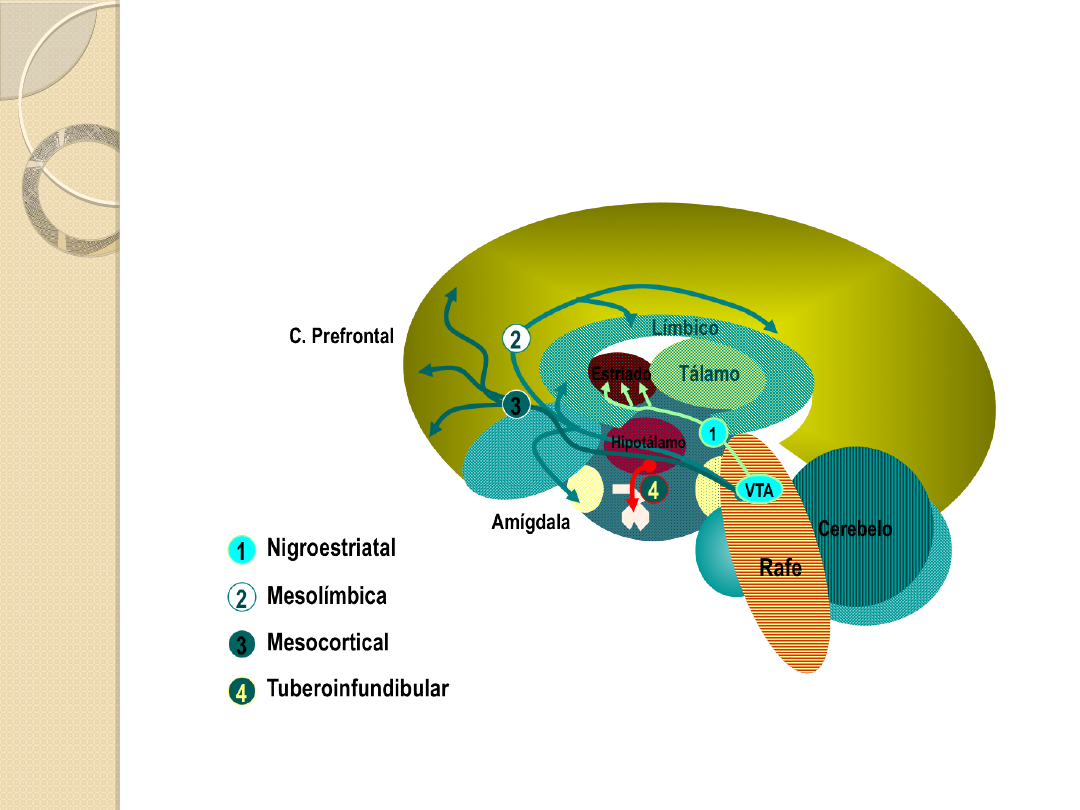
SZLAKI DOPAMINERGICZNE
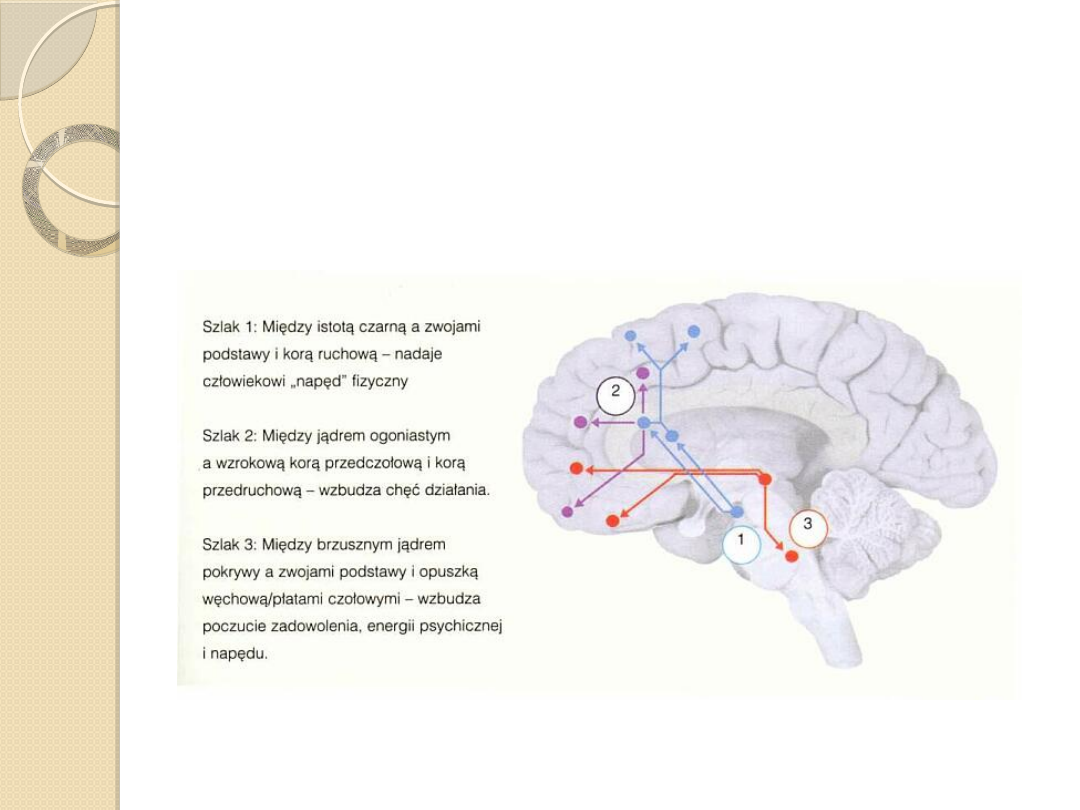
Szlak 1: nigrostriatalny
Szlak 2 mezolimbiczny
Szlak 3. mezokortykalny

Interakcje neuronów dopaminergicznych
z innymi typami neuronów.
Neurony dopaminergiczne szlaku
nigrostriatalnego znajdują się pod hamującym
wpływem neuronów GABA-ergicznych,
enkefalinergicznych i cholinergicznych.
Silnie hamująco na komórki dopaminergiczne
istoty czarnej działają zstępujące neurony
GABA-ergiczne.
Aktywująco działają na przekaźnictwo
dopaminergiczne neurony glutaminergiczne.
Zaburzenia w równowadze poszczególnych
typów neuronów w prążkowiu i istocie czarnej
stanowią podstawowy mechanizm choroby
Parkinsona.

Farmakologiczna modulacja aktywności
receptorów dopaminergicznych
Blokowanie receptorów dopaminergicznych w prążkowiu
manifestuje się:
Osłabieniem lub całkowitym zniesieniem napędu
ruchowego
Zmianami napięcia mięśniowego
Katalepsją i katatonią
efekt wykorzystywany w terapii
Blokowanie tych receptorów w strukturach limbicznych
prowadzi do uspokojenia i osłabienia reakcji
emocjonalnych (agresji)
efekt wykorzystywany w terapii
.
Blokowanie receptorów w opuszce hamuje odruch
wymiotny
efekt wykorzystywany w terapii
.
Pobudzenie receptorów dopaminergicznych w podwzgórzu
hamuje wydzielanie prolaktyny i ACTH
efekt
wykorzystywany w terapii
, natomiast blokowanie tych
receptorów nasila wydzielanie prolaktyny

UKŁAD NORADRENERGICZNY
OŚRODKOWEGO UKŁADU NERWOWEGO
Noradrenalina nie jest równomiernie
rozmieszczona w OUN.
Tworzy kilka dobrze wyodrębnionych szlaków
rozpoczynających się w obrębie pnia mózgu i
tworzących projekcje wstępującą i w mniejszym
stopniu zstępującą.
Szlak grzbietowy
powstaje w ciałach komórek
skupionych w obrębie tzw. jądra miejsca
sinawego (nucleus locus coeruleus), w
grzbietowej części mostu.
Szlak ten dociera do struktur korowych
kory mózgu, kory hipokampa i kory móżdżku
oraz
struktur podkorowych
jądra migdałowatego i
podwzgórza
.

Funkcja biologiczna szlaku grzbietowego:
regulacja czynności bioelektrycznej kory
regulacja procesów snu i czuwania
regulacja koncentracji i uwagi
procesy uczenia się i pamięci.
Nadmierna czynność szlaku grzbietowego
prowadzi do:
pobudzenia psychoruchowego
zaburzeń emocjonalnych (nasilenie lęku)
zahamowania aktywności poznawczej
utrudnienia uczenia się

Szlak brzuszny
powstaje z licznych skupisk komórek
umiejscowionych w brzusznej części nakrywki mostu i
dociera do wielu struktur podkorowych m.in.
podwzgórza
Funkcja biologiczna szlaku brzusznego :
regulacja czynności hormonalnych podwzgórza
działanie hamujące na czynnik podwzgórzowy CRF
kierujący syntezą i uwalnianiem ACTH, pobudzenie
uwalniania hormonu wzrostu
regulacja czynności ruchowych
regulacja procesów łaknienia
uszkodzenie szlaku
brzusznego powoduje zwiększone pobieranie pokarmu
(hyperphagia)
procesy uczenia się
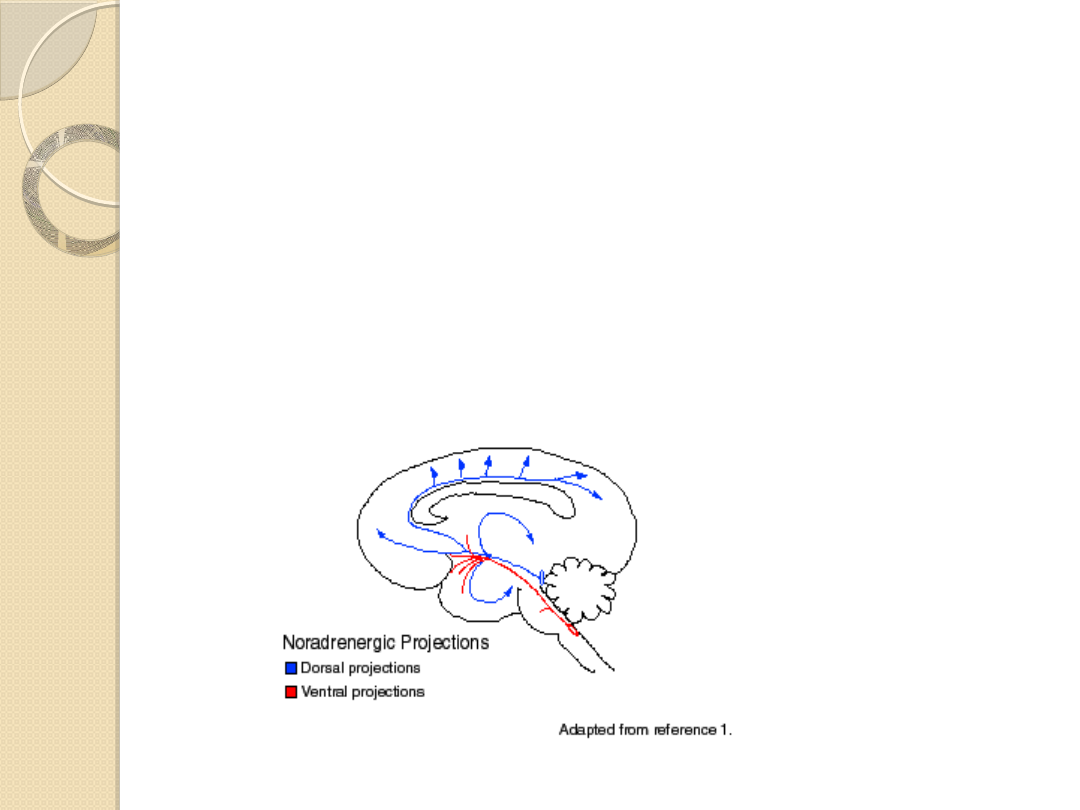
Pęczek okołokomorowy
(fasciculus
periventricularis) rozpoczyna się w
komórkach istoty szarej okołokomorowej i
okołowodociągowej mózgu
unerwia
podwzgórze i istotę szarą śródmózgowia.

W OUN występują wszystkie typy
receptorów adrenergicznych.
Postsynaptyczne:
α1, β
adrenergiczne
Presynaptyczne :
α2
adrenergiczne
(
autoreceptory
)
pobudzenie tych receptorów
prowadzi do zahamowania
uwalniania noradrenaliny do
szczeliny synaptycznej.

Pobudzenie receptorów α2 doprowadza do osłabienia
funkcji neuronów noradrenergicznych i do
zmniejszenia uwalniania noradrenaliny.
Związki pobudzające te receptory wywołują
wyraźny wpływ uspokajający, hamują aktywność
ruchową, wywołują miorelaksację i wywołują
analgezję.
Mechanizm wielu chorób psychicznych może być
związany ze zmianami wrażliwości tych receptorów.
W depresji endogennej może występować zwiększona
wrażliwość receptorów α2 i postsynaptycznych
receptorów β.
Leki będące antagonistami receptorów β
adrenergicznych hamują aktywność komórek
miejsca sinawego, hipokampa i kory móżdżku co
manifestuje się wystąpieniem uspokojenia.

INTERAKCJE NEURONÓW
NORADRENERGICZNYCH Z INNYMI TYPAMI
NEURONÓW
Antagonizm między neuronami
noradrenergicznymi a serotoninergicznymi.
Odgrywa to istotną rolę regulacyjną dla wielu
czynności OUN
snu i czuwania, napędu
ruchowego, emocji, nabywania czynności
warunkowych.
Neurony noradrenergiczne są również
hamowane przez neurony wytwarzające
enkefaliny i GABA
jest to hamowanie typu
presynaptycznego poprzez synapsy aksono-
aksonalne.

5-HYDROKSYTRYPTAMINA I
NEURONY SEROTONINERGICZNE
Serotonina w OUN stanowi ok. 1% całkowitej puli
występującej w organiźmie.
Jest ona bardzo ważnym neurotransmiterem w
OUN.
W szyszynce serotonina jest przekształcana
do N-acetyloserotoniny, a następnie do
melatoniny
hormonu odpowiedzialnego za
regulację rytmu dobowego organizmu i snu.
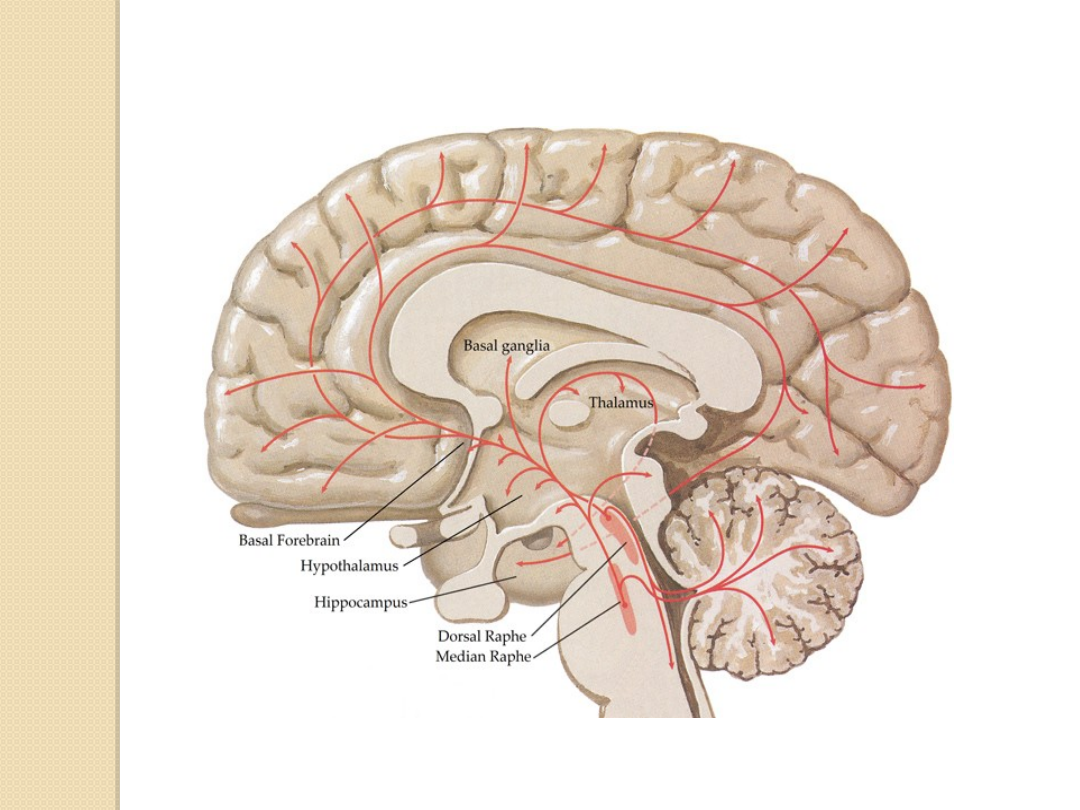
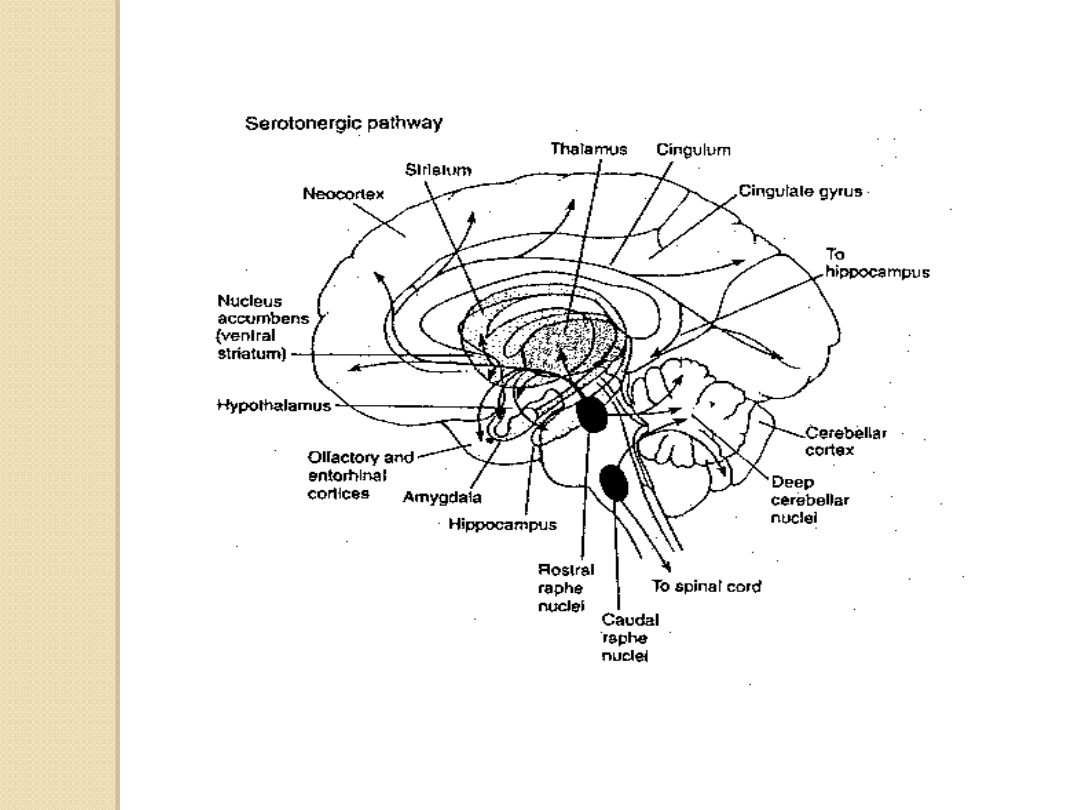
Neurony wytwarzające serotoninę są zgrupowane
w środkowych częściach pnia mózgu w tzw.
jądrach szwu.
Włókna osiowe wytwarzają liczne szlaki
wstępujące i zstępujące.

Do najlepiej poznanych skupisk komórek
wytwarzających serotoninę należą:
Jądro środkowe szwu
(nucleus raphe
medianus) unerwiające struktury korowe i
limbiczne (hipokamp)
Jądro grzbietowe szwu
(nucleus raphe
dorsalis) unerwiające struktury prążkowia,
korę i ciało migdałowate
Jądro wielkie
(nucleus raphe magnum)
unerwiające móżdżek i rdzeń kręgowy.
SKUPISKA KOMÓREK W OUN WYTWARZAJĄCYCH
SEROTONINĘ

SEROTONINA
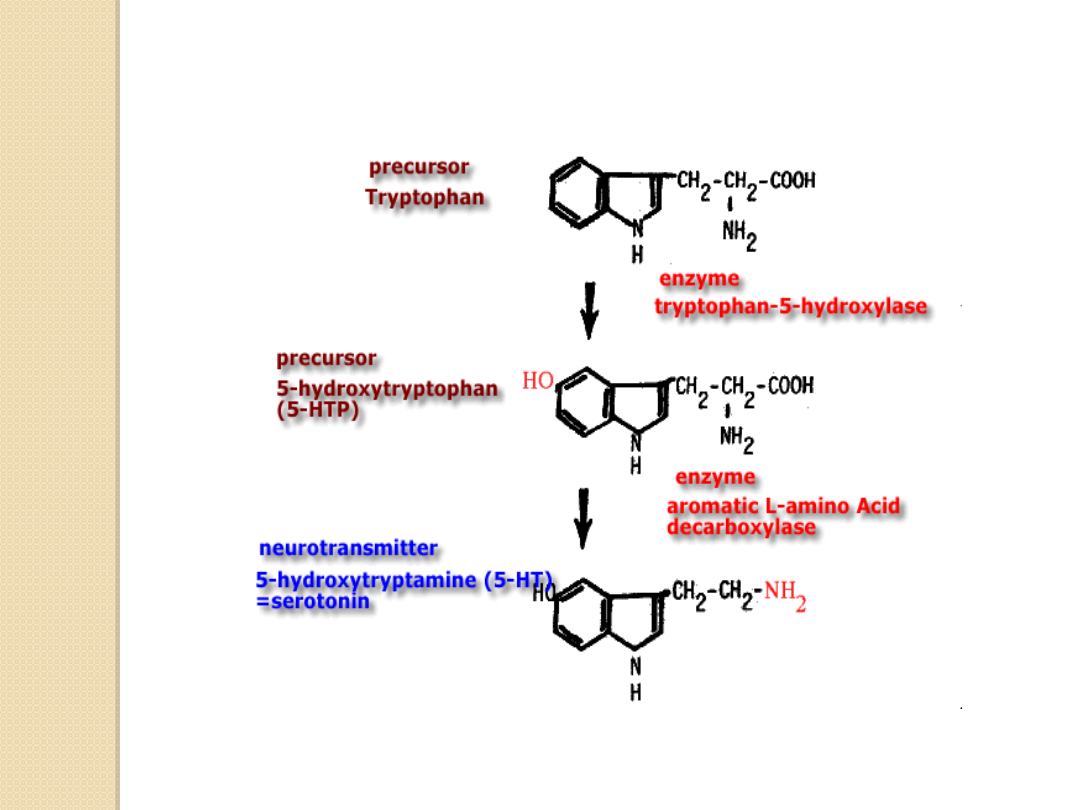
Serotonina jest indolaminą syntetyzowaną
blisko zakończenia nerwowego z aminokwasu
tryptofanu.
Tryptofan pod wpływem hydroksylazy
tryptofanu zostaje przekształcany w 5-
hydroksytryptofan podlegający działaniu
dekarboksylazy L-aminokwasów aromatycznych
następstwem czego jest powstanie cząsteczki
aktywnego biologicznie neuroprzekaźnika jakim
jest serotonina.
Stężenie serotoniny w strukturach ośrodkowego
układu nerwowego jest zależne od ilości
tryptofanu w pożywieniu.

SEROTONINA
Prędkość syntezy serotoniny jest dostosowana
do częstotliwości wyładowań neuronu.
Serotonina nie ma zdolności przenikania przez
barierę krew mózg, jej synteza i uwalnianie w
strukturach ośrodkowego systemu nerwowego
zależy od zdolności przenikania tryptofanu
przez barierę krew- mózg.
W medycynie weterynaryjnej stosuje się
tryptofan jako suplement diety zarówno u
zwierząt hodowlanych
(świnie, drób)
oraz
towarzyszących
(psy)
w celu zwiększenia
przyjmowania pokarmu oraz eliminowania
nieprawidłowych zachowań.

SEROTONINA
Serotonina odgrywa rolę hamującą w procesach
regulacyjnych w OUN.
Reguluje procesy snu (sen wolnofalowy i paradoksalny)
Wpływa na łaknienie
Wpływa na popęd płciowy
Wpływa na procesy emocjonalne
Odpowiedzialna jest za pobudzenie ruchowe.
Wpływa modulująco na przekaźnictwo bodźców
czuciowych i bólowych (drażnienie jąder szwu
wywołuje analgezję)
Serotonina bierze udział w regulacji czynności
hormonalnych podwzgórza i przysadki
wydzielanie
gonadotropin, ACTH
.
Bierze udział w indukcji odruchu wymiotnego

U zwierząt uszkodzenie jąder szwu powoduje:
Silne pobudzenie ruchowe
Impulsywność i zwiększoną drażliwość
Agresywność
Zaburzenia popędu płciowego
Antagonizm czynnościowy pomiędzy neuronami
serotoninergicznymi a noradrenergicznymi
Równowaga pomiędzy układami neuronów
noradrenergicznych a serotoninergicznych
odgrywa istotną rolę w procesach snu i czuwania

RECEPTORY DLA SEROTONINY
Wpływa na komórki efektorowe poprzez
kilka typów receptorów serotoninowych: 5-
HT1A, 5-HT1B, 5-HT1C, 5-HT1D, 5-HT2, 5-
HT3, 5-HT4
Receptory grupy 5-HT1A związane są z
hamowaniem aktywności neuronów (mogą
być także autoreceptorami)
Agoniści tych receptorów
(buspiron)
działają
przeciwlękowo i mogą wywierać działanie
przeciwdepresyjne
(flesinoksan)
Receptory 5HT1A regulują uwalnianie ACTH
i wpływają na ośrodkowe działanie
glikokortykosterydów.

Receptory 5-HT2 związane są z interakcją z
układem dopaminergicznym
antagoniści
receptora 5-HT2 nasilają uwalnianie
dopaminy.
Leki będące antagonistami tego receptora
stosuje się w leczeniu migreny u ludzi.
Antagoniści receptorów 5HT2 a także
agoniści postsynaptycznych receptorów
5HT1A osłabiają niekorzystny wpływ stresu i
niedotlenienia na komórki hipokampa.

Leki będące antagonistami receptora 5HT3
wywierają silne działanie przeciwwymiotne
Ondansetron jest silnym wybiórczym antagonistą
receptorów dla serotoniny (5-HT3) zlokalizowanych
centralnie w obszarze chemowrażliwej strefy
wyzwalającej (chemoreceptor trigger zone) na dnie
komory IV oraz w zakończeniach nerwu błędnego
ośrodkowo w jądrze pasma samotnego i obwodowo
w splocie śródściennym przewodu pokarmowego.
Ondansetron hamuje nudności i wymioty związane
z działaniem serotoniny uwalnianej w jelicie
cienkim przez leki cytostatyczne i radioterapię, a
także blokując ośrodkowe receptory 5-HT3.

ACETYLOCHOLINA I NEURONY
CHOLINERGICZNE
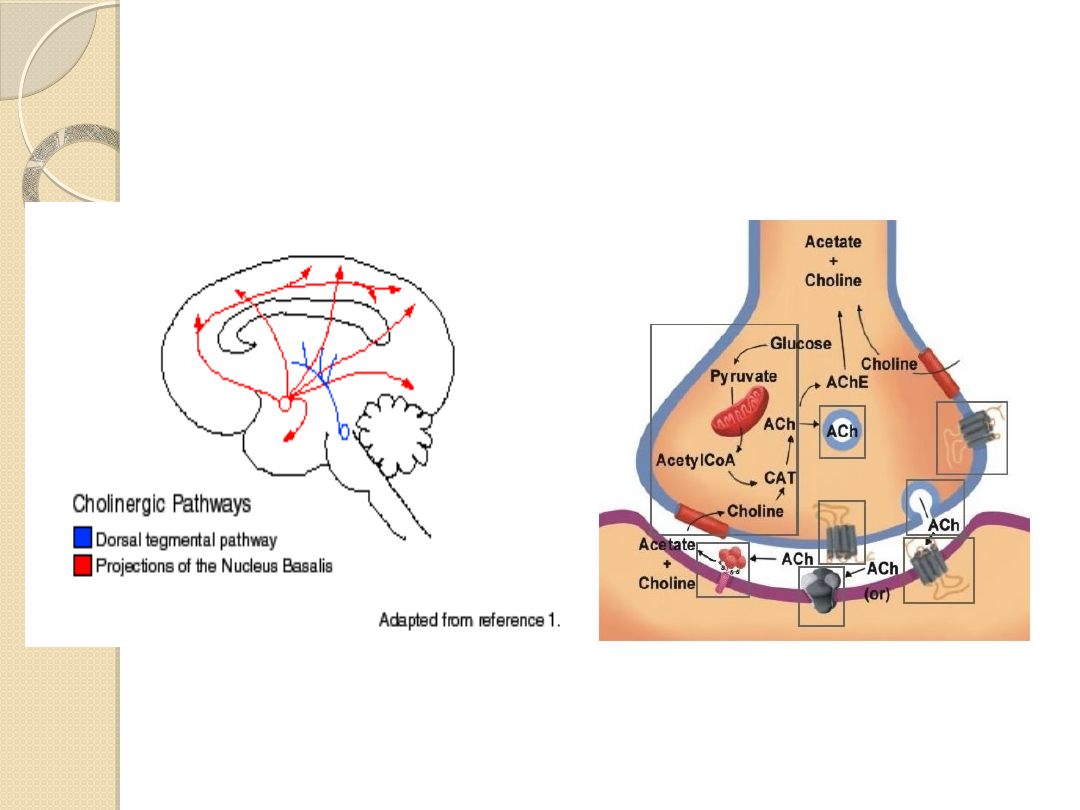
Ciała komórkowe neuronów
cholinergicznych są rozsiane w wielu
strukturach OUN:
w szlaku limbicznym
połączenie
cholinergiczne między przegrodą i
hipokampem
prążkowiu
podwzgórzu
korze mózgu
pniu mózgu
UKŁAD CHOLINERGICZNY W OUN

Funkcja biologiczna neuronów
cholinergicznych:
aktywność ruchowa
emocje
regulacja procesów snu i czuwania
kontrola budzenia się
procesy pamięci
procesy uczenia się
Wstrzyknięcie związków pobudzających
neurony cholinergiczne do podwzgórza
wywołuje silną agresję u zwierząt
doświadczalnych.

Podanie związków cholinergicznych do
hipokampu powoduje powstanie wyładowań
drgawkowych rozprzestrzeniających się na
inne struktury mózgu.
Podanie systemowe fizostygminy
zwierzętom doświadczalnym wywołuje
desynchronizację w obrazie korowym EEG i
pobudzenie rytmu w hipokampie.
Leki blokujące receptory cholinergiczne
(atropina) wywołuje synchronizację
czynności korowej w EEG.

Wpływ ACh i DA na czynność
struktur układu pozapiramidowego
ACh DA
stan dynamicznej
równowagi
o
słabienie czynności DA-ergicznej
(stan parkinsonoidalny, katalepsja)
przewaga czynnościowa układu DA-
ergicznego
(pobudzenie aktywności ruchowej,
stereotypia)

W OUN tak jak na obwodzie ACh wywiera
działanie receptorowe
Receptory cholinergiczne typu M1
(muskarynowe neuronalne)
i N
(nikotynowe).
Leki blokujące nieselektywnie receptory M
stosowane są w leczeniu choroby Parkinsona.
Leki zwiększające stężenie Ach w OUN
(
inhibitory acetylocholinoesterazy o działaniu
ośrodkowym
donepezil, fizostygmina,
galantamina
) usprawniają funkcje poznawcze i
pamięć i stosowane są w leczeniu choroby
Alzheimera u ludzi.

HISTAMINA
Rola histaminy w OUN jest słabo poznana.
Duże stężenia histaminy występują w
podwzgórzu wpływ na wydzielanie
wazopresyny i prolaktyny oraz pełni rolę
regulacyjną w procesie termoregulacji.
Histamina wpływa hamująco na dużą część
neuronów w korze mózgowej i osłabia
przekazywanie bodźców przez spoidło wielkie.
W OUN występują receptory H1, H2 i H3
Receptor H3 jest autoreceptorem hamującym
osłabia uwalnianie histaminy.

Receptory H2 mogą znajdować się presynaptycznie
na neuronach noradrenergicznych, działając
hamująco na wydzielanie noradrenaliny.
Histamina bierze udział w regulacji odruchu
wymiotnego (antagoniści H1 – efekt
przeciwwymiotny
Amina ta reguluje również wydzielanie
prolaktyny
pobudzenie receptorów H1 wzrost
wydzielania
,
pobudzenie receptorów H2
hamowanie wydzielania
Blokowanie receptorów H1 sedacja i efekt
przeciwwymiotny.
Document Outline
- Slide 1
- Slide 2
- UKŁADY FUNKCJONALNE MÓZGU
- UKŁAD POBUDZENIA
- UKŁAD POBUDZENIA
- UKŁAD NAGRODY
- UKŁAD NAGRODY
- UKŁAD NAGRODY
- UKŁAD NAGRODY
- UKŁAD NAGRODY
- UKŁAD NAGRODY
- UKŁAD NAGRODY
- UKŁAD DOPAMINERGICZNY
- UKŁAD DOPAMINERGICZY
- Slide 15
- SYNAPSA DOPAMINERGICZNA
- Układy efektorowe receptorów dopaminergicznych.
- Receptory DA
- SZLAKI DOPAMINERGICZNE W MÓZGU.
- Slide 20
- Slide 21
- Slide 22
- SZLAKI DOPAMINERGICZNE
- Slide 24
- Interakcje neuronów dopaminergicznych z innymi typami neuronów.
- Slide 26
- UKŁAD NORADRENERGICZNY OŚRODKOWEGO UKŁADU NERWOWEGO
- Slide 28
- Slide 29
- Slide 30
- Slide 31
- Slide 32
- INTERAKCJE NEURONÓW NORADRENERGICZNYCH Z INNYMI TYPAMI NEURONÓW
- 5-hydroksytryptamina i neurony serotoninergiczne
- Slide 35
- Slide 36
- Slide 37
- SEROTONINA
- SEROTONINA
- Slide 40
- SEROTONINA
- Slide 42
- RECEPTORY DLA SEROTONINY
- Slide 44
- Slide 45
- ACETYLOCHOLINA I NEURONY CHOLINERGICZNE
- UKŁAD CHOLINERGICZNY W OUN
- Slide 48
- Slide 49
- Wpływ ACh i DA na czynność struktur układu pozapiramidowego
- Slide 51
- HISTAMINA
- Slide 53
Wyszukiwarka
Podobne podstrony:
3 AKTYWNOŚĆ BIOLOGICZNA NEUROMEDIATORÓW W OUN część II
Flokulacja kłaczków osadu czynnego przez stymulacją tlenowej aktywności biologicznej
Aktywność biologiczna izoflawonoidów i ich znaczenie terapeutyczne i kosmetyczne
Wykład 9 2 Podstawy biologicznego oczyszczania ścieków część II
Aktywność biologiczna białek w surowcach i żywności
Aktywność biologiczna flawonoidów i ich zastosowanie w kosmetykach
[21]aktywność biologiczna flawonoidów, Bibliografia
Wykład 9 1 Podstawy biologicznego oczyszczania ścieków część I ppt
Podstawy biologicznego oczyszczania ścieków część II
Enzymy i ich aktywność biologiczna, Kosmetologia, Notatki i wyłady, Biochemia
Profile potencjalnych aktywności biologicznej białek
więcej podobnych podstron