
Biomaterials 21 (2000) 667}681
Review
Osteoblast adhesion on biomaterials
K. Anselme*
Institut de Recherche sur les Maladies du Squelette, Institut Calot, Rue du Dr Calot, 62600 Berck sur mer cedex, France
Received 11 January 1999; accepted 21 October 1999
Abstract
The development of tissue engineering in the "eld of orthopaedic surgery is now booming. Two "elds of research in particular are
emerging: the association of osteo-inductive factors with implantable materials; and the association of osteogenic stem cells with these
materials (hybrid materials). In both cases, an understanding of the phenomena of cell adhesion and, in particular, understanding of
the proteins involved in osteoblast adhesion on contact with the materials is of crucial importance. The proteins involved in osteoblast
adhesion are described in this review (extracellular matrix proteins, cytoskeletal proteins, integrins, cadherins, etc.). During
osteoblast/material interactions, their expression is modi"ed according to the surface characteristics of materials. Their involvement
in osteoblastic response to mechanical stimulation highlights the signi"cance of taking them into consideration during development
of future biomaterials. Finally, an understanding of the proteins involved in osteoblast adhesion opens up new possibilities for the
grafting of these proteins (or synthesized peptide) onto vector materials, to increase their in vivo bioactivity or to promote cell
integration within the vector material during the development of hybrid materials.
( 2000 Elsevier Science Ltd. All rights reserved.
Keywords: Osteoblast; Adhesion; Biomaterial; Tissue engineering
1. Introduction
Cell adhesion is involved in various natural phe-
nomena such as embryogenesis, maintenance of tissue
structure, wound healing, immune response, metastasis
as well as tissue integration of biomaterial. The biocom-
patibility of biomaterials is very closely related to cell
behaviour on contact with them and particularly to cell
adhesion to their surface. Surface characteristics of ma-
terials, whether their topography, chemistry or surface
energy, play an essential part in osteoblast adhesion on
biomaterials. Thus attachment, adhesion and spreading
belong to the "rst phase of cell/material interactions and
the quality of this "rst phase will in#uence the cell's
capacity to proliferate and to di!erentiate itself on con-
tact with the implant.
It is essential for the e$cacy of orthopaedic or dental
implants to establish a mechanically solid interface with
complete fusion between the material's surface and the
bone tissue with no "brous tissue interface. Moreover,
the recent development of tissue engineering in the "eld
* Tel.: #33-0321-892029; fax: #33-0321-892070.
E-mail address: kanselme@hopale.com (K. Anselme)
of orthopaedic research makes it possible to envisage the
association of autologous cells and/or proteins that pro-
mote cell adhesion with osteoconductive material to cre-
ate osteoinductive materials or &hybrid materials'. Thus,
a complete understanding of cell adhesion and parti-
cularly osteoblast adhesion on materials is now essential
to optimize the bone/biomaterial interface at the heart of
these hybrid materials. This includes an understanding
of the molecules involved in bone cell adhesion, parti-
cularly regarding interaction with the materials but also
the need to take into account osteoblastic reaction to the
mechanical constraints which will be applied to im-
planted materials in vivo. The application of non-de-
structive in vitro mechanical constraints to the cell/
biomaterial interface permits understanding of the
e!ects of mechanical stimulation on the synthesis of
adhesion proteins, cell growth and cell di!erentiation
and provides essential information for the development
of hybrid materials.
This paper o!ers a review of present knowledge of
osteoblast adhesion, focused on in vitro adhesion on
orthopaedic biomaterials. The aim of the author is to
highlight useful information for the improvement of pres-
ent biomaterials and the future development of new
biomaterials.
0142-9612/00/$ - see front matter
( 2000 Elsevier Science Ltd. All rights reserved.
PII: S 0 1 4 2 - 9 6 1 2 ( 9 9 ) 0 0 2 4 2 - 2

2. Description of bone cell adhesion
The term &adhesion' in the biomaterial domain covers
di!erent phenomena: the attachment phase which occurs
rapidly and involves short-term events like physico-
chemical linkages between cells and materials involving
ionic forces, van der Walls forces, etc. and the adhesion
phase occurring in the longer term and involving various
biological molecules: extracellular matrix proteins, cell
membrane proteins, and cytoskeleton proteins which in-
teract together to induce signal transduction, promoting
the action of transcription factors and consequently regu-
lating gene expression. Those various proteins involved
in cell adhesion will be described in this "rst part.
2.1. Proteins involved in osteoblast cell adhesion
2.1.1. Extracellular matrix proteins
The extracellular matrix of bone is composed of 90%
collagenic proteins (type I collagen 97% and type V col-
lagen 3%) and of 10% non-collagenic proteins (NCP)
(osteocalcin 20%, osteonectin 20%, bone sialoproteins
12%, proteoglycans 10%, osteopontin, "bronectin,
growth factors, bone morphogenetic proteins, etc.). All
these proteins are synthesized by osteoblasts and most
are involved in adhesion. In vitro, other proteins such as
"bronectin
or vitronectin have been shown to be in-
volved in in vitro osteoblast adhesion. Some other pro-
teins with a plasmatic origin are also associated with
mineralized bone matrix but their role in osteoblast
adhesion is not fully de"ned (
a2HS glycoprotein, al-
bumin, immunoglobulin, transferrin, etc.) [1].
Some of the bone proteins have chemotactic or adhes-
ive properties, notably because they contain an Arg}
Gly}Asp (RGD) sequence which is speci"c to the "xation
of cell membrane receptors like integrin ("bronectin, os-
teopontin, bone sialoprotein, thrombospondin, type I
collagen, vitronectin) [1,2].
In order to determine the proteins involved in bone cell
adhesion, experiments consisting of bone cell cultures on
dishes coated with extracellular matrix proteins are cur-
rently used. Human osteoblasts adhere preferentially to
"bronectin as compared with type I, type IV collagen,
and vitronectin but weakly to laminin and type V col-
lagen. In contrast, they do not adhere to type III collagen
[3].
Other adhesion experiments use arti"cially synthesized
peptides like GRGDS (Gly}Arg}Gly}Asp}Ser) [2] or
RGDS (Arg}Gly}Asp}Ser) [4]. After mixing the peptide
with culture medium, it was demonstrated that RGDS
peptide partially inhibited rat calvarial bone cell attach-
ment to "bronectin-coated plates in a competitive dose-
dependent manner although RGES control peptide had
a minimal e!ect on cell attachment [4]. A GRGDS
peptide almost completely blocked the attachment to
bone sialoprotein and vitronectin and slightly decreased
the attachment to "bronectin, type I collagen and throm-
bospondin [2]. The addition of the peptide GRGDSP
(Gly}Arg}Gly}Asp}Ser}Pro) to human osteoblast-like
osteosarcoma cells SaOs-2 cells in a serum free medium,
inhibited cell adhesion by 28% on titanium-based alloys
(Ti6Al4V) and by 40% on CoCrMo alloys although
adhesion on glass and plastic was not a!ected. The con-
trol peptide GRADSP (Gly}Arg}Ala}Asp}Ser}Pro) had
no e!ect on cell adhesion [5]. The strength of rat cal-
varial cell adhesion, measured with a radial #ow appar-
atus, was signi"cantly higher on a RGD-peptide-coated
surface compared to a RGE-peptide-coated surface [6].
The role of RGD-peptides conformation on osteo-
progenitor cell adhesion has been highlighted [7]. These
experiments have demonstrated the importance of iso-
lated RGD-sequence containing peptides in promoting
adhesion of bone cells. Biomaterials may be improved by
a preliminary adsorption of these peptides.
2.1.2. Cytoskeleton proteins
The sites of adhesion between tissue cultured cells and
substrate surfaces are called focal contacts or adhesion
plaques. Focal contacts are closed junctions where the
distance between the substrate surface and the cell mem-
brane is between 10}15 nm. This type of junction is rare
in vivo except for endothelial cells in vessels with high
hydrodynamic stress [8]. They also appear to be analo-
gous to sarcolemnal dense plaques of smooth muscle cells
in vivo.
The external faces of focal contacts present speci"c
receptor proteins such as integrins. On the internal face,
some proteins like talin, paxillin, vinculin, tensin are
known mediating interactions between actin "laments
and membrane receptor proteins (integrins) (Fig. 1).
Many proteins colocalize with vinculin and talin in the
adhesion plaque: integrin, cytoskeletal proteins, pro-
teases, protein kinases and phosphatases, signalling
molecules, etc. These proteins are involved in signal
transduction.
The formation of focal contacts occurs essentially in
cells with low motility and is promoted in vitro by ex-
tracellular matrix proteins like "bronectin or vitronectin.
The architecture of the actin cytoskeleton is essential to
the maintenance of cell shape and cell adhesion. If assem-
bled in long bundles, F-actin supports "nger-like protru-
sions of the plasma membrane known as "lopodia; if
assembled in the form of a mesh, it supports sheet-like
protrusions known as lamellipodia. If present in bundles
coupled with adhesion plaques, actin &stress "bers' may
transmit forces to the substrate [9].
2.1.3. Adhesion molecules
Adhesion molecules are characterized by their capacity
to interact with a speci"c ligand. These ligands may be
situated on the membrane of neighbouring cells or may
be extracellular matrix proteins. Adhesion molecules
668
K. Anselme / Biomaterials 21 (2000) 667}681

Table 1
The integrin family of adhesion receptors
!
Sub-units
Other names
Ligands
Cells
b1
a1
VLA-1
Collagen-I, collagen-IV and laminin
Bone cells, cultured osteoblastic cells, activated
T-cells, monocytes, melanoma cells and smooth
muscle cells
a2
VLA-2, GPIa-IIa,
ECMRII
Collagens-I to IV, laminin and possibly
Fibronectin
Bone cells, cultured osteoblastic cells, B and
T lymphocytes, platelets, "broblasts, endothelial cells
and melanoma cells
a3
VLA-3, VCA-2,
ECMRI, Gapb-3
Fibronectin, Collagen-I, Epiligrin,
Laminin and Nidogen/Entactin
Cultured osteoblastic cells, B-lymphocytes, kidney
glomerulus and most cultured cell lines
a4
VLA-4, LPAM-2
Fibronectin, VCAM-1
Cultured osteoblastic cells, lymphocytes, monocytes,
eosinophils, NK-cells and thymocytes
a5
VLA-5, FNR,
GPIc-IIa, ECMRVI
Fibronectin
Bone cells, cultured osteoblastic cells, memory-
T-cells, monocytes, platelets and "broblasts
a6
VLA-6, GPIc-IIa
Laminin-1, -2, -4 and -5 (only in vitro)
Platelets, lymphocytes, monocytes, thymocytes and
epithelial cells
a7
VLA-7
Laminin-1
Skeletal and cardiac muscle at speci"c stages during
muscle development and melanoma cells
a8
Fibronectin
?
a9
Tenascin-C, osteopontin
Adult skeletal muscle, visceral smooth muscle, skin
and corneal epithelial cells, hepatocytes
aV
Fibronectin, vitronectin
Cultured osteoblastic cells
b2
aD
?
?
aL
LFA-1, CD11a-CD18
ICAM-1 to -3
Leucocytes
aM
Mac-1, CR-3,
CD11b-CD18
C3bi, coagulation factor X,
"brinogen and ICAM-1
Monocytes, macrophages, NK cells and granulocytes
aX
p150, CR-4,
CD11c-CD18
Fibrinogen
Monocytes, macrophages, granulocytes, NK cells
and activated lymphocytes
b3
aV
VNR, CD51
Fibrinogen, "bronectin, Von Willebrand's
factor, vitronectin, thrombospondin,
osteopontin and bone sialoprotein 1
Bone cells, cultured osteoblastic cells, endothelial
cells, some B-cells, platelets and monocytes
aIIb
GPIIb-IIIa, CD41
Fibrinogen, "bronectin, Von Willebrand's
factor and vitronectin
Platelets
b4
a6
Laminin-1
Immature thymocytes, squamous epithelia, subsets
of endothelial cells, Schwann cells and "broblasts
in the peripheral nervous system
b5
aV
aVbS, aVb3b
Vitronectin
Cultured osteoblastic cells, hepatoma cells,
"broblasts and carcinoma cells
b6
aV
Fibronectin
Carcinoma cells
b7
a4
LPAM-1
MadCAM, "bronectin and VCAM-1
Leukocytes (directed to the Peyer's patches of the gut)
aIELb M290 IEL, aH
?
?
b8
aV
?
Mature synapses of central nervous system
!VLA: very late activation antigen, VCA: very common antigen, ECMR: extra cellular matrix receptor, GP: glycoprotein, Gap: galactoprotein, LFA:
leukocyte function associated antigen, M290 IEL: mouse intraepithelial lymphocyte antigen recognized by monoclonal antibody M290, FNR:
"bronectin receptor, ICAM: inter cellular adhesion molecule, LPAM: lymphocyte Peyer's patch HEV adhesion molecule (mouse), VCAM: vascular
cellular adhesion molecule, CR: C3bi receptor, VNR: vitronectin receptor, MAC: macrophage receptor.
belong to di!erent families. The four main classes are
selectins, immunoglobulin superfamily, cadherins and in-
tegrins. Amongst them, only cadherins and integrins have
been described at this time in osteoblastic cells.
Integrins: cell}substrate adhesion. The integrin family is
composed of 22 heterodimers of two types of sub-units
a and b. 16 a sub-units and 8 b sub-units have been
discovered. This diversity of structures is associated with
various ligand-binding possibilities (Table 1). Integrins
are transmembrane heterodimers consisting of non-
covalently associated
a and b sub-units. Each sub-unit is
made up of a large extracellular domain, a transmem-
brane domain and a short cytoplasmic domain. The
integrin spanning the cell membrane acts as an interfacer
between the intra- and extra-cellular compartments and
can translate the attachment of external ligands to inter-
nal information which induces adhesion, spreading, or
cell migration and consequently regulates cell growth
and di!erentiation (Fig. 1).
Recently, the expression of integrin in bone and in
cultured bone cells has been demonstrated. On human
bone sections, all bone cell types expressed
a1 and
K. Anselme / Biomaterials 21 (2000) 667}681
669
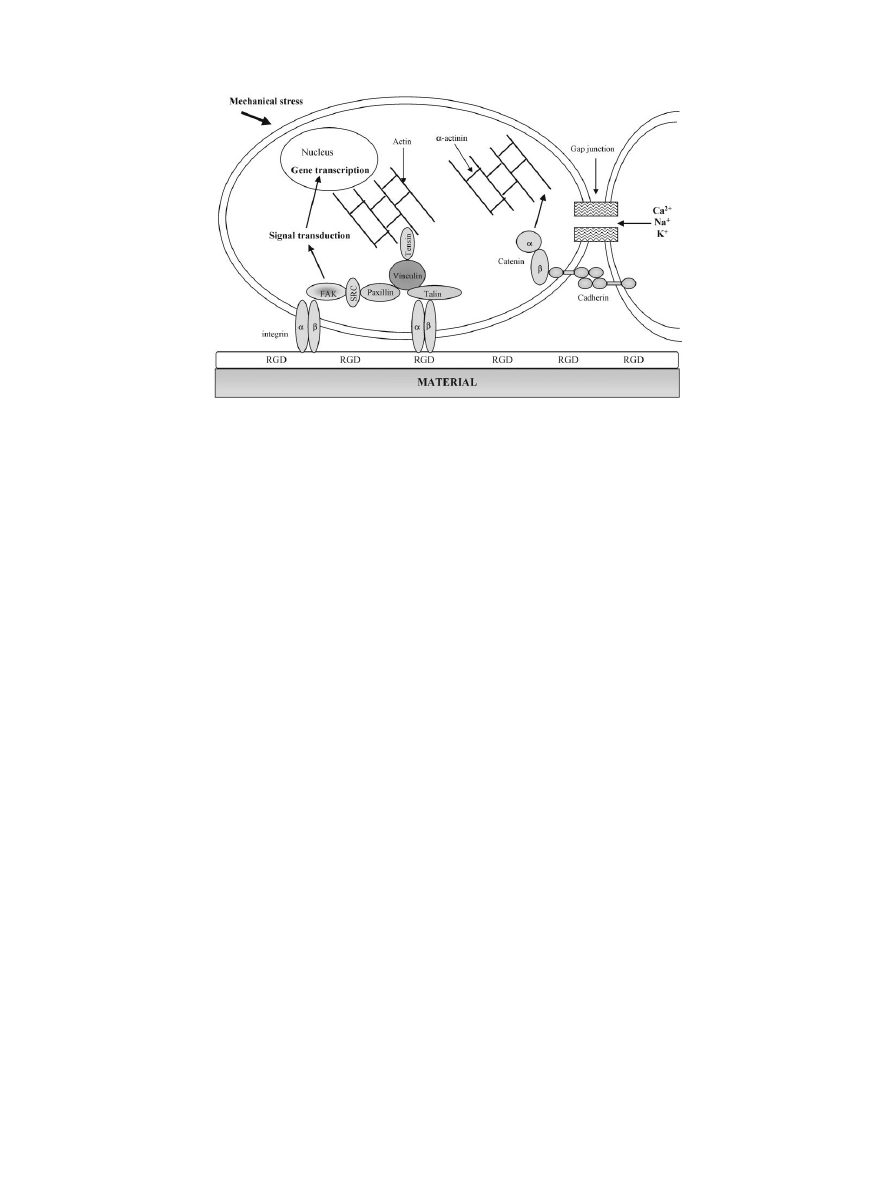
Fig. 1. Representation of the cell proteins involved in cell adhesion on biomaterial.
a5 sub-units although a subpopulation of osteoblastic
cells expressed
a2, a7 and a7b3 [10]. Cultured human
osteoblasts expressed high levels of
a1b1, a3b1, a5b1 and
a7b5 and much lower levels of a2b1, a4b1, a7b1 and a7b3
integrins [11].
b1 sub-unit appears to be the predomi-
nant receptor involved in osteoblast adhesion to collagen
and laminin and is partially involved in adhesion to
"bronectin [3]. In cells adhering in the presence of serum,
b3 and a7 integrin were recovered in focal contacts.
a4 and b1 subunits are expressed by primary human
osteoblasts on "bronectin, type I collagen and laminin-
coated control polystyrene dishes.
a2, a6 and a7 are
expressed on control dishes,
a2, a3 and a7 on collagen-
coated dishes and
a6 and a7 on laminin-coated dishes.
b3 subunit is expressed on all surfaces except on
laminin-coated dishes [12]. Adhesion to "bronectin, col-
lagen- and laminin-coated dishes is inhibited by anti-
b1
integrin subunit antibody although anti-
a5 integrin
subunit antibody a!ects adhesion only to "bronectin
[13]. An understanding of the integrin subunits involved
in osteoblast adhesion provides essential information for
biomaterial improvement and notably for de"nition of
the proteins which may be useful to adsorb on materials
before implantation.
Cadherin: cell}cell adhesion. As cell}substrate ad-
hesions are based on integrin-type receptors, adherens
junctions containing cadherins mediate cell}cell ad-
hesion (CAM). Cadherins are transmembrane glycopro-
teins acting with intracellular partners: catenins which
interact with intracellular proteins [14] (Fig. 1). Associ-
ation with
a, b or c-catenin is a prerequisite for the
adhesive function of cadherins. The cadherin family is
composed of numerous types of calcium-dependent mol-
ecules (E, P, N, L, R, 6B, 7, 11, 4). They associate in
a zipper homophilic model of interactions between cad-
herin molecules exposed on the plasma membrane of
adjacent cells [15]. Firstly, osteoblasts have been shown
to express E-cadherins [16] and cadherin-11 (or OB-
cadherin) [17]. More recently, human osteoblasts have
been shown to express mRNA for cadherin-11, N-cad-
herin and low levels of cadherin-4. The expression of
cadherin-4 is modulated by bone morphogenetic pro-
tein-2 (BMP-2) contrary to cadherin-11 and N-cadherin
expression [18]. Cadherin-11 gene is expressed in the
bone marrow and bone cells obtained from rabbits of
various age groups. The relative level of cadherin-11 gene
is greater in mature rabbit marrow than in young or aged
animals [19].
Gap junctions: cell}cell communications. Cell rec-
ognition and adhesion precede and control cell}cell
communication via gap junctions [20]. Intercellular
communications occur through direct exchange of ions
via gap junctions or through signals produced by the
action of CAM.
Gap junctions are constituted by homohexamus de-
rived from a family of proteins called connexins. When
the connexin of one cell (composed of six connexin mol-
ecules) is in register with a similar structure on a neigh-
bouring cell, a transcellular channel is formed [20]. Tight
and adherens junctions or desmosomes provide anchor-
age to surrounding cells and allow direct exchange of
ions or small molecules between cells (Fig. 1).
Osteoblasts express in vitro two connexins, connexin
43 and connexin 45 [21,22]. Connexin 43 expression by
human osteoblastic cells is regulated by retinoic acid and
transforming growth factor-
b1 (TGF-b1) [23] and by
parathyroid hormone (PTH) and prostaglandin E2
(PGE2) [20,24].
670
K. Anselme / Biomaterials 21 (2000) 667}681

2.2. Signal transduction
As previously mentioned, integrins, CAM molecules
and gap junctions regulate gene expression by a signal
transduction induced by cell}substrate or cell}cell ad-
hesion. Integrins and cadherins have direct interactions
with the cytoskeleton (actin) via
a-actinin, talin and
catenins. Integrin clustering and occupancy provoke the
recruitment of tensin and focal adhesion kinase (FAK),
their phosphorylation and subsequently the recruitment
of talin, vinculin, and
a-actinin [25] (Fig. 1). Various
other intracellular signalling pathways are activated by
cell adhesion: protein tyrosine phosphorylations, mito-
gen-activated protein kinase (MAP) activation, Ca
2` in-
#ux, pH alterations and inositol lipid turnover.
b-catenin
molecules can interact with the cadherin system but also
with fascin (a molecule involved in the formation of
F-actin bundles) and with transcription factors [14].
Some other molecules like FAK may play a mechanical
role in binding a number of signalling and cytoskeletal
molecules in parallel with its kinase activity (Fig. 1).
Talin, vinculin and
a-actinin link the F-actin
"bers to
the plasma membrane. The rearrangement of F-actin
"bre bundles induces the cell shape changes producing
the "nger-like or sheet-like protrusions ("lopodia or
lamellipodia, respectively). The signals mediated by cell
shape may be processed by the nuclear matrix which is
physically linked to the cytoskeleton via the nuclear
lamins. Subsequently, adhesion plaque activation is es-
sential for signal transduction and then regulation of
gene expression. ECM-mediated changes in cell shape
can modify the nuclear matrix and therefore modify
gene expression. ECM-mediated changes in cell shape
initiate a switch between the proliferative and di!erenti-
ative state. For example, as osteoblasts shift from a
proliferative to a rounded di!erentiated state, a novel
nuclear matrix protein NMP-2 appears. NMP-2 binds to
the osteocalcin gene promoter and induces its expression
[26].
2.3. Adhesion and cell migration
Cell migration requires a dynamic interaction between
the cell, its substrate and its cytoskeleton. Firstly, cells
develop a protrusion of their leading edge to form
a lamellipodium. Secondly, after formation and "xation
of the lamellipodium, cells use adhesive interactions to
generate the traction and energies required for cell move-
ment. The last step of the migratory cycle is the release of
adhesions at the rear of the cell followed by its detach-
ment and retraction. Integrin have been shown to be
involved in cell migration [27]. In general, cells with
a low motility form strong focal adhesions while motile
cells form less adhesive structures. An intermediate level
of attachment force induces a maximal migration rate
[28]. Some antiadhesive extracellular matrix proteins
play a role in cell migration: tenascin, thrombospondin,
laminin, muscin, proteoglycans [29]. Migration tests on
biomaterials with surface grooves demonstrated that cell
migration was faster on materials with deeper grooves
[30].
3. Osteoblast/material interactions
Osteoblast/material interaction depends on the surface
aspects of materials which may be described according to
their topography, chemistry or surface energy. These
surface characteristics determine how biological molecu-
les will adsorb to the surface and more particularly deter-
mine the orientation of adsorbed molecules [31]. They
also determine the cell behaviour on contact. As pre-
viously shown, cells in contact with a surface will "rstly
attach, adhere and spread. This "rst phase depends on
previously described adhesion proteins. Thereafter, the
quality of this adhesion will in#uence their morphology,
and their capacity for proliferation and di!erentiation.
Early in vitro cytocompatibility studies focused on the
morphological aspect, growth capacity and the state of
di!erentiation of cells on materials with various chemical
compositions [32}38]. The diversity of cell responses to
the di!erent materials tested highlighted the capacity of
cells to discriminate between di!erent chemistries. How-
ever, the sensitivity of in vitro biological tests was some-
times too low to distinguish the e!ects of subtle changes
in substratum surface chemistry. No di!erences were
observed in cell colonization of polystyrene dishes
treated by sulphuric acid and gamma-irradiation al-
though X-ray photoelectron spectroscopy (XPS) analysis
demonstrated that the two treatments introduced di!er-
ent chemical groups onto the polymer surface [39].
Surfaces are di!erent from the corresponding bulk of
the material. For thermodynamic reasons they contain
unsaturated bonds which lead to the formation of surface
reactive layers and adsorbed contamination layers. Prep-
aration technique e!ects such as sterilization e!ects have
been studied by several authors. They have demonstrated
the crucial e!ect of the sterilization methods of commer-
cially pure titanium (cpTi) on in vitro subsequent cell
adhesion [40,41]. These e!ects may be related to steriliz-
ation-induced surface chemical modi"cations [42]. Some
clinical surface preparation on titanium-based alloys are
known to considerably modify the surface chemical char-
acteristics. Improper glow discharge plasma treatment (a
frequently used method for cleaning, preparation and
modi"cation of biomaterial) can produce unwanted and
irreproducible results [43]. The surface oxide on tita-
nium implant materials is mainly TiO2 and is about 2 nm
thick. One signi"cant di!erence, as compared to the
machined unalloyed Ti, was that AlOx and no V was
detected on the machined Ti6Al4V surfaces. The concen-
tration of Al on the outermost surface may constitute
K. Anselme / Biomaterials 21 (2000) 667}681
671

a potential risk for Al dissolution in the surrounding
tissues [44]. Consequently, caution must be taken with
the surface preparation of these alloys. The surface oxide
formed on the alloy might play a role in the extent to
which the materials will be accepted. So, the e!ects of
surface preparation on cell cytocompatibility must be
previously veri"ed before surgical use because the biolo-
gical properties of bulk material could be largely modi-
"ed by the process of surface preparation [45].
3.1. Morphological aspects of osteoblast on materials
The comparison of the behaviour of di!erent cell types
on materials shows that they react di!erently according
to surface roughness [30,46}48]. Scanning electron
microscopic examination of bone cells on materials with
various surface roughness generally demonstrated that
cell spreading and continuous cell layer formation was
better on smooth surfaces compared to rough ones
[35,49,50].
The organization of surface roughness is an important
parameter to consider. In vitro, many authors have dem-
onstrated the contact guidance phenomenon using
"broblasts or epithelial cells. Epithelial cells were mark-
edly oriented along the long axis of 10
lm deep grooves
on a titanium-coated implant [51]. The modi"cations of
groove and ridge dimensions of a silicone rubber substra-
ta suggested that a ridge width )4
lm was necessary to
observe rat dermal "broblast orientation along the sur-
face grooves. The modi"cations of groove width or
groove depth did not a!ect the cellular orientation [52].
The contact guidance phenomenon has also been de-
scribed on osteoblastic cells. On smooth surfaces, bone
cells were randomly oriented although they were aligned
parallel to the direction of the grooves in an end-to-end
fashion in 5
lm-deep grooves. In contrast, they
&ignored'
the surface topography on an 0.5
lm grooved surface
[30]. In vivo, it appears that the surface characteristics of
an implant, particularly roughness, may control tissue
healing and therefore subsequent implant success
[53,54]. The hydrophilic and hydrophobic character-
istics of a material are also of great importance for cell
adhesion. Cell adhesion is generally better on hydrophilic
surfaces [55]. Spreading of human skin "broblasts in-
creased along chemically characterized gradient surfaces
going from the hydrophobic to the hydrophilic end [56].
The morphological aspect of neonate rat calvarial os-
teoblasts cultured on a positively or negatively charged
polymer substrata was signi"cantly di!erent. Cells #at-
tened out so closely onto the positively charged substrata
that the ventral cell membrane was not distinguishable
through the transmission electron microscope. On nega-
tively charged substrata, the ventral cell membrane was
readily visible with only focal contacts with the substrata
[57]. Bone calvarial cells randomly plated on materials
with patterned surface chemistry rapidly (t(30 min) or-
ganized on positive charged regions in the presence of
serum. After 30 min, cells started to align and spread
exclusively on these regions. By day 2, cells started to
extend from positive charged regions to negative charged
regions due to lack of surface area available and were
con#uent after 4 days [46].
3.2. Osteoblast adhesion on material
Cells do not interact with a naked material either
in vitro or in vivo. At each step, the material is
conditioned by the biological #uid components [31]. The
pH as well as the ionic composition and strength of
solution, temperature and the functional group of pro-
teins and substrates are the factors determining protein
adsorption.
Surface energy may in#uence protein adsorption and
the structural rearrangement of the proteins on the ma-
terial [31]. Protein adsorption was di!erent on positively
and negatively charged polymer substrates [57]. Protein
from serum containing media adsorbed on surfaces form-
ing multiple molecular layers [30]. Many recent experi-
ments using materials with patterned surface chemistry
obtained by photolithography demonstrated the di!er-
entiated adsorption of serum proteins on the negative
(DMS) or positive (EDS) charged regions. The role of
vitronectin for in vitro cell adhesion has been highlighted
by several authors [36,47,58]. Sera depleted of vitronec-
tin, with or without "bronectin, greatly reduced cell at-
tachment and spreading on patterned surfaces. Thus, the
presence of vitronectin is essential for in vitro spatial
distribution, attachment and spreading of bone-derived
cells in an EDS region [47].
The role of plasma "bronectin (pFN) adsorption in cell
adhesion has been also established [55]. Moreover, it is
clear that not only the nature of adsorbed biological
molecules but also their conformation will in#uence con-
sequent cell adhesion. Changes in pFN conformation
a!ected "bronectin cell binding domain conformations
and then a!ect FN a$nity with its cell surface receptor
[59].
A direct relationship exists between roughness and
surface energy of materials. A systematic study evaluated
cell adhesion on polymethylmetacrylate (PMMA) mate-
rials with various degrees of surface roughness. It was
demonstrated that the apolar component of surface en-
ergy increased signi"cantly with roughness although the
basic component decreased and that cell adhesion en-
hancement was related to the degree of roughness and
the hydrophobicity [60].
In a recent work, the adhesion of human osteoblasts
on Ti6Al4V substrates with "ve di!erent degrees of sur-
face roughness was studied. Cell adhesion was correlated
with roughness parameters and in particular with para-
meters describing the organization of the surface rough-
ness. More so than the roughness amplitude evaluated by
672
K. Anselme / Biomaterials 21 (2000) 667}681

Ra or Rt, the developed surface or Delta parameter
describing the surface organization was the more in#uen-
tial parameters on human osteoblastic cell adhesion to
orthopaedic alloys [50].
Osteoblast adhesion on materials may also be con-
sidered in relation to the expression of the various
adhesion proteins. Numerous studies using immuno-his-
tochemical methods have shown the presence of actin
and vinculin in cultured osteoblasts on various materials.
Cell adhesion was higher on Ti6Al4V'polystyrene'
CoCrMo alloy. The rate of actin and vinculin cyto-
skeletal reorganization was enhanced on Ti6Al4V. Focal
contacts remained peripherally located in cells on
Ti6Al4V and CoCrMo although on polystyrene the focal
contacts quickly became dispersed along actin "laments.
The authors suggested that osteoblast attachment was
greater on Ti6Al4V because cell spreading and cytos-
keletal organization were enhanced [61]. In human
osteoblasts cultured on a titanium alloy with various
degrees of roughness, we showed the presence after 1,
7 and 14 days of short, thin and dense vinculin positive
patches. Logically, the distribution of focal contacts illus-
trated the mode of adhesion of osteoblasts on the di!er-
ent roughnesses. On smooth surfaces, focal contacts were
distributed uniformly on all the membrane surface which
was in contact with the substratum. On rough surfaces,
focal contacts were visible only at the extremities of cell
extensions where cell membranes were in contact with
the substrate [50].
Not only topographical but also physico-chemical
characteristics of the surface in#uence the distribution of
focal contacts. Analysis of focal contacts using speci"c
labelling of actin and vinculin showed their presence only
on electropolished and etched titanium surfaces but not
on sandblasted ones [62]. The F-actin cytoskeleton
reorganization on various materials showed a typical
sequence: at 6 h, the F-actin was generally di!usely dis-
tributed with circumferential banding near the edge of
cells. At 12 h, thin micro"lament bundles were visible. At
24 h, cells were spread and numerous well-de"ned stress
"bres were observed [37]. After 24 h, an actin micro"la-
ment system ran parallel to the long axis of cells [50].
Double localization of actin and vinculin showed that
actin formed numerous stress "bres thoughout the
cytosol which terminated at the cell periphery and that
the vinculin-containing adhesion plaques were situated
at the terminal &ends' of the actin stress "bres [38].
More recently, the integrins mediating osteoblast ad-
hesion on various materials were studied. Some experi-
ments consisted of inhibiting cell attachment to various
materials and of determining the integrin subunits in-
volved in cell adhesion using anti-integrin subunit anti-
bodies. An antibody to the "bronectin receptor
a5b1
signi"cantly inhibited adhesion on Ti6Al4V by 63% and
to CoCrMo by 49%. Serum had no e!ect on the number
of cells that attached to Ti6Al4V and CoCrMo alloys but
did increase the number of cells that attached to glass.
Osteoblast-like cell adhesion on glass contrary to
Ti6Al4V and CoCrMo was not mediated by integrins but
did require the adsorption of vitronectin [5]. Integrin
expression of primary human osteoblasts was analysed
after culture on rough and polished CoCrMo and
Ti6Al4V alloys.
a3 and a6 integrin sub-units were ex-
pressed only on polished Ti6Al4V.
a2, a4, a7 and b1 in-
tegrin sub-units were expressed on all surfaces and
b3 sub-unit was expressed on all surfaces except on rough
CoCrMo [12]. We also observed the expression of
a3 and b1 integrin sub-units by primary human osteo-
blasts but no expression of
a2 integrin sub-units [50].
The anti-
b1 sub-unit antibody revealed thin, short
"laments parallel to the cytoskeleton arrangement on all
the membrane surface. The morphology of
b1 "laments
varied slightly on di!erent substratum roughnesses
[12,50]. It seems that
b1 integrin was the major integrin
sub-unit involved in osteoblast adhesion on biomaterials.
3.3. Osteoblast diwerentiation on materials
As previously described, some proteins can be adsorb-
ed in vitro from the serum containing media or in vivo
from biological #uids. Vitronectin [63] or "bronectin
[50], extracellular matrix proteins synthesized by bone
cells in vitro or adsorbed from serum, are essential for
osteoblast in vitro adhesion [58]. After the attachment
phase, extracellular collagenous and non-collagenous
matrix proteins are synthesized by cells [30,64].
Neonatal rat calvarial osteoblasts expressed mRNA en-
coding osteonectin and osteopontin when cultured on
Ti6Al4V, HA or tissue culture polystyrene [65]. Some
qualitative and quantitative di!erences in bone protein
expression has been observed between various substra-
tes: rat bone marrow stromal cells expressed more os-
teopontin and bone sialoprotein on HA compared to
cpTi or glass-ceramic [66]. SaOs-2 human osteoblast-
like cell line synthesized a higher collagen and non-
collagen protein quantity when cultured on a CoCrMo
alloy compared to glass control discs [5].
Using human osteoblast-like cells (MG63), it was
found that on titanium disks with various degrees of
roughness, proliferation and alkaline phosphatase activ-
ity was reduced when roughness increased. In contrast,
collagen synthesis increased with roughness although
non-collagenous protein synthesis was not a!ected by
roughness [64]. PGE2 and TGF
b1 production was high-
er on rough surfaces than on smooth ones [49]. More-
over, the roughness increased the MG63 cells response to
1
a,25(OH)2D3 vitamin and notably promoted the in-
crease of osteocalcin synthesis and alkaline phosphatase
activity [67]. Adult jaw bone cells had a lower prolifer-
ation on rough-surface (plasma-sprayed) hydroxyapatite
(HA) or titanium substrates compared to smooth surface
(polished), a lower alkaline phosphatase activity but
K. Anselme / Biomaterials 21 (2000) 667}681
673

a higher osteocalcin synthesis [34]. Human bone-derived
cells cultured on titanium-based metallic substrates [68]
or ceramic substrates [69] expressed and translated dif-
ferently mRNAs for speci"c osteoblastic proteins like
alkaline phosphatase, thrombospondin, osteopontin, os-
teocalcin, osteonectin, type I collagen and bone sialo-
protein. This di!erential regulation of proteins occurring
between cells from the same patient implies that human
bone-derived cells respond to small di!erences in the
surface chemistry and or/microcrystallinity.
Bone cells cultured on materials with patterned surface
chemistry during 15 days formed multilayered cell layer
on the entire surface but Von Kossa staining demon-
strated the presence of mineralized tissue consistent with
the underlying chemical pattern. Mineralized tissue was
preferentially localized on positive charged regions [46].
4. Mechanical aspects of osteoblasts/biomaterial
interactions
The role of mechanical load in bone remodelling is well
known. In vitro, osteoblastic cells respond to mechanical
stimuli with alterations in proliferation and/or pheno-
typic expression. Adhesion is also a!ected by mechanical
stimulation.
Firstly, the in#uence of mechanical stimulation on
in vitro osteoblast adhesion, proliferation and di!erenti-
ation will be considered. Secondly, the use of mechanical
strain for cell detachment and quantitative cell adhesion
evaluation will be presented.
4.1. Mechanical ewects on osteoblasts
The osteoblastic cell response to mechanical stimula-
tion has been widely studied using various cell strains
and various methods of load application. Consequently,
heterogenous results were obtained. The mechanism of
cellular response to #ow was demonstrated to be shear
stress independent [70]. In some cases, a dose depen-
dence was observed between the number of responsive
cells and shear stress magnitude [71,72]. On the other
hand, other authors suggest that bone and bone-like cells
respond to mechanical signals in a trigger-like rather
than a dose}response fashion [73].
Generally, cell proliferation was stimulated by mech-
anical stress [74}78]. Optimum magnitude of tensional
forces for osteoblastic cell division was determined
[75,76]. Applications of mechanical strains on human
bone cells caused a signi"cant e!ect on proliferation but
only in a proportion of subjects [78].
Osteoblast di!erentiation generally implies alkaline
phosphatase activity (ALP) and speci"c protein expres-
sion like osteocalcin, osteopontin, type I collagen and
in vitro mineralization capacity. In vitro mechanical
stimulation has shown various e!ects on ALP activity of
cells [73,75,79}81]. Low vacuum pressure depressed
ALP activity in the cultures of rat calvaria bone cells and
mouse calvaria cell line (MC3T3-E1) [73] although inter-
mittent hydrostatic compression upregulated ALP
activity in mouse calvarial bone cells [80]. Rat calvarial
osteoblasts subjected to #uid #ow expressed lower ALP
mRNA notably under pulsatile #uid #ow [82]. These
observations illustrate the signi"cance of strain charac-
teristics and methods used to apply mechanical strain on
bone cells: continued or intermittent strains, applied by
compression, stretch or #uid #ow methods.
Speci"c osteoblastic protein expression is also modi-
"ed
by strains. Osteopontin synthesis is generally in-
creased [75,83}85]. On the other hand, osteocalcin
synthesis is di!erently in#uenced [73,86]. Collagen ex-
pression by cells under mechanical strains is also highly
variable [82,87]. Osteoprogenitor cells reacted di!erently
to osteoblastic cells. Collagen expression of osteoblastic
cells after intermittent hydrostatic compression was
decreased although collagen expression by osteoprogeni-
tor cells was increased under the same conditions [82].
This illustrates the in#uence of the cell line used for the
experiment.
The cytoskeleton is also modi"ed by mechanical
stimulation. Intermittent hydrostatic compression up-
regulated actin expression [82]. Mechanical strain in-
creased formation and thickening of actin stress "bers
[82]. A 250% increase of vinculin was observed following
mechanical stimulation and especially at the peripheral
edges of the cells [88]. However, vinculin appears not to
play a role in mechanical transduction because depletion
of vinculin from focal contacts did not prevent the re-
sponse of cells to mechanical stimulation [33]. The
microtubule role in mechanical force transduction has
been also highlighted [88,89].
Prostaglandin E2 (PGE2) or prostacyclin synthesis by
osteoblasts are systematically increased by mechanical
strains [78,90}95]. PGE2 synthesis is one of the early
responses of osteoblasts and induces a secondary produc-
tion of cAMP which is involved in signal transduction
[91]. In mechanically stimulated osteoblastic cells, cAMP
increase was also largely demonstrated [70,86,91,96].
As previously described, cell adhesion may occur on
RGD-sequence-containing proteins like collagen or os-
teopontin via speci"c membrane receptors called integ-
rins. Some authors focused on expression of
b1 integrin
sub-units and
a2 integrin sub-units by osteoblasts sub-
mitted to mechanical stimulation [97}99]. Using para-
magnetic beads coated with anti-integrin antibodies, it
was possible to apply de"ned physical forces on indi-
vidual integrin receptor sub-units [97,99]. These experi-
ments con"rmed previous results demonstrating the role
of integrin in transmission of mechanical signals into cells
[100,101].
Cell}cell adhesion proteins are also involved in osteo-
blast response to mechanical stimulation. As previously
674
K. Anselme / Biomaterials 21 (2000) 667}681

Table 2
Techniques for the detachment of cells and quantitative evaluation of
cell adhesion
Technique
Cells
Micropipet aspiration
Human blood cells [110,111]
Porcine aorta endothelial cells [112]
Normal and SV-40 transformed human
dermal "broblasts [113]
Centrifugation
SV-40 transformed murine peritoneal
macrophages and human erythrocytes
[114]
Vero Green monkey kidney [115]
Fibroblasts and glioma cells [116]
Normal and SV-40 transformed human
dermal "broblasts [113]
Paramagnetics beads
Fibroblasts [117,118]
Osteoblastic cells [99]
Osteoblast-like cells [107]
Fluid #ow
Mouse macrophage cell line [119]
3T3 "broblasts [120]
Bovine aortic endothelial cells [121]
Enzymatic detachment
Chick embryo aortic explants [118]
Human and chick embryo trabecular
bone explants [122]
Primary human osteoblastic cells [50]
Spinning disk
3T3 "broblasts [123]
Osteoblast-like cells [108,109]
Microplate manipulation
Chick "broblasts [124]
Microcantilever
L929 murine "broblasts [125]
shown, osteoblasts express connexin 43 and 45 [21,22].
Cyclic stretch enhances gap junctional communications
between osteoblastic cells [102]. These gap junctions
are involved in intercellular calcium wave propaga-
tion [103] and then induce mechanosensitive signal
transduction pathways. Many experiments have shown
that substratum stretch, hydrostatic pressure and
#uid shear stress increased inositol
triphosphate (IP3)
in bone cells with IP3 causing the release of calcium
from intracellular stores. The IP3 biochemical pathway is
one of the mediators of the response of bone cells to
mechanical stimulation [71,90]. Mechanosensitive ion
channels have also been demonstrated in osteoblasts
[104,105].
The principle mechanical detection system of cells is
the matrix-integrin mechanosensory protein complex-
cytoskeleton machinery which is linked to a kinase cas-
cade system. Mechanosensory protein complex contains
talin, vinculin, tensin, paxillin, Src and focal adhesion
kinase (FAK) (Fig. 1). The kinetics of response events in
mechanically loaded cells may entail, during milliseconds
to seconds after stimulation, a signalling response involv-
ing mechanically active channels Ca
2`, Ma`, K`, H`,
IP3, cAMP, PGE2, kinases, G proteins, etc. After min-
utes to hours after stimulation, signalling with kinases,
transcription and transduction proceeds, and cyto-
skeletal protein polymerization and focal adhesion
rearrangement occur. After some days, cells migrate, ex-
press and degrade extracellular matrix, divide or die.
A new state of equilibrium is established [106].
4.2. Quantitative evaluation of cell adhesion
Quantitative evaluation of cell adhesion generally im-
plies the detachment of cells. To detach cells, various
techniques may be used (Table 2).
Most experiments attempted to apply external forces
to cells cultured on control surfaces and either to quan-
tify and mathematically create a model of the adhesive
cell/substrate connections or the mechanical deforma-
tions of individual cells. Only recently have studies fo-
cused on measurement of osteoblastic cell adhesion:
some on control surfaces [99,107], others on bioactive
biomaterials [108,109].
The optimal tensional force that stimulates osteoblast
activity was determined [75]. An application of over
10 000
l-strain resulted in a dedi
!erentiation of the os-
teoblasts and a change in cell morphology to become
"broblast like [126]. The proliferation of human osteob-
lasts was increased signi"cantly by 1% strain although
higher strain magnitudes had lesser (non-signi"cant) ef-
fects or decreased the mitotic activity of the cells [76].
A spinning disc device permitted quanti"cation of rat
osteosarcoma cells on various substrates. Cell attach-
ment strength approximated 20 dyn/cm
2 (2 N/m2) on
bioactive glass or borosilicate control glass. If "bronectin
was previously adsorbed on these materials, cell attach-
ment strength was comprised between about 50 and
70 dyn/cm
2 [109].
Optical techniques such as total internal re#ection
#uorescence microscopy (TRIFM) or internal re#ection
microscopy (IRM) permit visual examination of cell/sub-
strate contacts in real time. Additionally, TRIFM allows
quanti"cation of the separation distance of cells from
a biomaterial surface and has been used to evaluate the
adhesion strength on a biomaterial surface of bovine
endothelial cells following exposure to #ow [127].
5. Perspectives: improvement of biomaterials
Biomaterials currently available for clinical use are
known for their good biocompatibility. However, if most
of them have the mechanical properties required for
a de"ned implantation site, they do not all possess the
necessary bioactivity properties for good tissue regenera-
tion, i.e. not only osteoconductive properties but also
osteoinductive properties.
Several means are currently being developed for bio-
materials improvement like the surface modi"cation of
materials by protein adsorption procedures; or bone tis-
sue engineering associating autologous bone cells with
biomaterial.
K. Anselme / Biomaterials 21 (2000) 667}681
675

5.1. Adhesion protein adsorption
The proteins currently being considered for chemical
surface modi"cation of materials are growth factors (or
related proteins) and adhesion proteins (or related peptides).
Among the bone-related growth factors, some are at
the present time being tested for their ability to promote
bone regeneration. Notably, members of the Transform-
ing Growth Factor-
b family are being widely studied:
TGF-
b1, BMP-2, BMP-7 (or osteogenic protein-1 [OP-
1]). BMPs were identi"ed following puri"cation of
bovine bone proteins after it was discovered that de-
mineralized bone segments or extracts of demineralized
bone-induced bone formation in ectopic sites [128,129].
BMPs associated with various carriers have experi-
mentally shown their e$cacy [130}133] and are current-
ly under clinical investigation [134].
Amongst adhesion proteins, as previously described,
RGD-peptides have shown their e$cacy in promoting
osteoblast adhesion. The use of RGD-peptide may be
a future way of improving biomaterial surface. However,
the promotion of adhesion by RGD-peptides was not
associated with a subsequent enhancement of cellular
functions [135]. OP-1 promoted mineralization of os-
teoblast cultured on a RGD-peptide. These results sug-
gest that the combination of selected bioactive agents
and proactive biomaterials may synergistically enhance
clinically desirable cellular functions [136]. Other pep-
tides were explored. One method consists of synthesizing
a 15-residue peptide related to a biologically active por-
tion of type I collagen. This peptide was adsorbed on
inorganic bovine bone mineral particles to encourage
attachment of human dermal "broblasts. Cell attach-
ment on the coated particles increased with augmented
peptide content on the surface of bone particles. The
presence of P-15 promoted the formation of tri-dimen-
sional colonies. P-15 coated bovine bone mineral par-
ticles may be a useful matrix for bone repair [137].
5.2. Bone engineering: osteoblast/biomaterial association
Recently, a new "eld of biological/biomedical research
has developed: cellular and tissue engineering. This is
a consequence of advances in cell isolation and culture
procedures, combined with a growing understanding of
cell physiology. In particular, the previously described
knowledge of osteoblast adhesion on biomaterials may
provide essential information on the development of
&hybrid
materials' containing biocompatible osteocon-
ductive structural materials and autologous bone cells
for a self-cell therapy.
Former studies in this "eld concerned porous calcium
phosphate
ceramics
associated
with
non-cultured
autologous bone marrow cells. They were demonstrated
to produce bone after subcutaneous implantation
[138}140] and after experimental implantation in bone
defects [141}144]. In fact, bone marrow is known to
contain osteogenic precursor cells [145,146]. After in
vivo implantation in di!usion chambers, human os-
teogenic precursor cells developed an osteogenic tissue
consisting of a mineralizing "brous component and carti-
lage [147,148].
More recently, in vitro long-term culture of osteoblasts
in porous materials like ceramics [149}152], collagen
[153,154], commercial bone substitutes [155,156] or
polymer sca!olds [157] have been developed. They have
allowed investigation of how bone marrow cells adhere
to the surface of porous calcium phosphate ceramics
[150,152]. Cell di!erentiation during cultures was
monitored [151,153,156,158] and osteoblastic cell pen-
etration inside porous material and across inter-pore
connections was illustrated [151,158]. The sizes of inter-
pore connections were the most important parameters
for a deep colonization of porous materials after im-
plantation [158].
Some of these works were at the root of the use of
previously cultured bone cells for hybrid material prep-
aration [159}165]. The culture period may allow stimu-
lation of the inherent osteogenic ability of marrow
stromal stem cells in pores of porous hydroxyapatite
using for example a dexamethasone stimulation of cells
[165]. Frozen-preservation of cells could be used because
recultured cells after thawing also formed bone in in vivo
porous ceramic assay [159]. Cells may be cultured before
implantation either in tissue culture dishes before being
mixed with the material [161,162,166] or directly cul-
tured in the material [159,163,165,167].
6. Conclusion
This review has highlighted the complexity of the phe-
nomena occurring in cell/material interactions and parti-
cularly the role of cell adhesion, which conditions
subsequent cell behavior at the interface with the mate-
rial. A complete understanding of cell behavior in contact
with the material is becoming more and more essential in
attaining adequate health safety conditions for clinical
use of these hybrid materials. The development of tissular
engineering techniques in the orthopaedic domain is re-
quiring more and more the consideration of osteoblast
adhesion properties whether for the improvement of the
surfaces of materials by adsorption or grafting of speci"c
adhesion factors, or for the development of hybrid mate-
rials associating autologous bone cells and materials.
Acknowledgements
The author thanks B. NoeKl for the tables and "gures
achievement and Dr. P. Hardouin for critical reading of
the manuscript.
676
K. Anselme / Biomaterials 21 (2000) 667}681

References
[1] Malaval L, Chenu C, Delmas PD, ProteHines de l'os. Maladies
meHtaboliques osseuses de l'adulte. Kuntz D, editors. Paris:
Flammarion MeHdecine Sciences, 1996. p. 17}35.
[2] Grzesik WJ, Robey PG. Bone matrix RGD glycoproteins: im-
munolocalization and interaction with human primary osteo-
blastic bone cells in vitro. J Bone Miner Res 1994;9:487}96.
[3] Gronthos S, Stewart K, Graves SE, Hay S, Simmons PJ. Integrin
expression and function on human osteoblast-like cells. J Bone
Miner Res 1997;12:1189}97.
[4] Puleo DA, Bizios R. RGDS tetrapeptide binds to osteoblasts
and
inhibits
"bronectin-mediated
adhesion.
Bone
1991;
12:271}6.
[5] Gronowicz G, McCarthy M-B. Response of human osteoblasts
to implant materials: integrin-mediated adhesion. J Orthopaed
Res 1996;14:878}87.
[6] Rezania A, Thomas CH, Branger AB, Waters CM, Healy KE.
The detachment strength and morphology of bone cells contact-
ing materials modi"ed with a peptide sequence found within
bone sialoprotein. J Biomed Mater Res 1997;37:9}19.
[7] Verrier S, Jonkczyk A, Dard M, Amedee J. E!ect of linear and
cyclic RGD-containing peptides on osteoprogenitor cell ad-
hesion onto extracellular matrix proteins. J Bone Miner Res 1998;
23:S535.
[8] Burridge K, Fath K. Focal contacts: transmembrane links be-
tween the extracellular matrix and the cytoskeleton. BioEssays
1989;10:104}8.
[9] Zigmond SH. Signal transduction and actin "lament organiza-
tion. Curr Opin Cell Biol 1996;8:66}73.
[10] Hughes DE, Salter DM, Dedhar S, Simpson R. Integrin expres-
sion in human bone. J Bone Miner Res 1993;8:527}33.
[11] Saito T, Albelda SM, Brighton CT. Identi"cation of integrin
receptors on cultured human bone cells. J Orthopaed Res
1994;12:384}94.
[12] Sinha RK, Tuan RS. Regulation of human osteoblast integrin
expression by orthopedic implant materials. Bone 1996;18:451}7.
[13] Pistone M, Sanguineti C, Federici A, et al. Integrin synthesis and
utilization in cultured human osteoblasts. Cell Biol Int 1996;
20:471}9.
[14] Yamada KM, Geiger B. Molecular interactions in cell adhesion
complexes. Curr Opin Cell Biol 1997;9:76}85.
[15] Takeichi M. Morphogenetic roles of classic cadherins. Curr
Opin Cell Biol 1995;7:619}27.
[16] Babich M, Foti LRP. E-cadherins identi"ed in osteoblastic cells:
e!ects of parathyroid hormone and extracellular calcium on
localization. Life Sci 1994;54:201}8.
[17] Okasaki M, Takeshita S, Kawai S, et al. Molecular cloning and
characterization of OB-cadherin, a new member of cadherin
family expressed in osteoblasts. J Biol Chem 1994;269:12092}8.
[18] Cheng S-L, Lecanda F, Davidson MK, et al. Human osteoblasts
express a repertoire of cadherins, which are critical for BMP-2-
induced osteogenic di!erentiation. J Bone Miner Res 1998;
13:633}44.
[19] Goomer RS, Maris T, Amiel D. Age-related changes in the
expression of cadherin-11, the mesenchyme speci"c calcium-de-
pendent cell adhesion molecule. Calcif Tissue Int 1998;62:532}7.
[20] Civitelli R. Cell}cell communication in bone. Calcif Tissue Int
1995;56:S29}31.
[21] Civitelli R, Beyer EC, Warlow PM, Robertson AJ, Geist ST,
Steinberg TH. Connexin 43 mediates direct intercellular com-
munication in human osteoblastic cell networks. J Clin Invest
1993;91:1888}96.
[22] Steinberg TH, Civitelli R, Geist ST, et al. Connexin 43 and
connexin 45 form gap junctions with di!erent molecular per-
meabilities in osteoblastic cells. EMBO 1994;13:744}50.
[23] Chiba H, Sawada N, Oyamada M, et al. Hormonal regulation
of connexin 43 expression and gap junctional commu-
nication in human osteoblastic cells. Cell Struct Funct 1994;
19:173}7.
[24] Schiller PC, Mehta PP, Roos BA, Howard GA. Hormonal
regulation of intercellular communication: parathyroid hor-
mone increases connexin 43 gene expression and gap-junctional
communication in osteoblastic cells. Mol Endocrinol 1992;
6:1433}40.
[25] Craig SW, Johnson RP. Assembly of focal adhesions: progress,
paradigms, and portents. Curr Opin Cell Biol 1996;8:74}85.
[26] Roskelley CD, Srebrow A, Bissell MJ. A hierarchy of ECM-
mediated signalling regulates tissue-speci"c gene expression.
Curr Opin Cell Biol 1995;7:736}47.
[27] Wu CY, Fields AJ, Kapteijn BAE, McDonald JA. The role of
a4b1 integrin in cell motility and "bronectin matrix assembly.
J Cell Sci 1995;108:821}9.
[28] Huttenlocher A, Sandborg RR, Horwitz AF. Adhesion in cell
migration. Curr Opin Cell Biol 1995;7:697}706.
[29] Chiquet-Ehrismann R. Inhibition of cell adhesion by anti-adhes-
ive molecules. Curr Opin Cell Biol 1995;7:715}9.
[30] Chesmel KD, Clark CC, Brighton CT, Black J. Cellular re-
sponses to chemical and morphologic aspects of biomaterial
surfaces. II. The biosynthetic and migratory response of bone
cell populations. J Biomed Mater Res 1995;29:1101}10.
[31] Boyan BD, Hummert TW, Dean DD, Schwartz Z. Role of
material surfaces in regulating bone and cartilage cell response.
Biomaterials 1996;17:137}46.
[32] Keller JC, Stanford CM, Wightman JP, Draughn RA, Zaharias
R. Characterizations of titanium implant surfaces. III. J Biomed
Mater Res 1994;28:939}46.
[33] Meyer U, Szulczewski DH, MoKller K, Heide H, Jones DB.
Attachment kinetics and di!erentiation of osteoblasts on di!er-
ent biomaterials. Cells Mater 1993;3:129}40.
[34] De Santis D, Guerriero C, Nocini PF, et al. Adult human bone
cells from jaw bones cultured on plasma-sprayed or polished
surfaces of titanium or hydroxyapatite discs. J Mater Sci-Mater
Med 1996;7:21}8.
[35] Naji A, Harmand M-F. Study of the e!ect of the surface state on
the cytocompatibility of a Co}Cr alloy using human osteoblasts
and "broblasts. J Biomed Mater Res 1990;24:861}71.
[36] Howlett CR, Evans MDM, Walsh WR, Johnson G, Steele JG.
Mechanism of initial attachment of cells derived from human
bone to commonly used prosthetic materials during cell culture.
Biomaterials 1994;15:213}22.
[37] Puleo DA, Bizios R. Formation of focal contacts by osteoblasts
cultured on orthopaedic biomaterials. J Biomed Mater Res
1992;26:291}301.
[38] Hunter A, Archer CW, Walker PS, Blunn GW. Attachment and
proliferation of osteoblasts and "broblasts on biomaterials for
orthopaedic use. Biomaterials 1995;16:287}95.
[39] Callen BW, Sodhi RNS, Shelton RM, Davies JE. Behavior of
primary bone cells on characterized polystyrene surfaces. J Bio-
med Mater Res 1993;27:851}9.
[40] Vezeau PJ, Koorbusch GF, Draughn RA, Keller JC. E!ects of
multiple sterilization on surface characteristics and in vitro
biologic responses to titanium. J Oral Maxillofac Surg 1996;
54:738}46.
[41] Stanford CM, Keller JC, Solursh M. Bone cell expression on
titanium surfaces is altered by sterilization treatments. J Dent
Res 1994;73:1061}71.
[42] Baier RE, Meyer AE, Akers CK, et al. Degradative e!ects of
conventional steam sterilization on biomaterial surfaces. Bio-
materials 1982;3:241}5.
[43] Aronsson BO, Lausmaa J, Kasemo B. Glow discharge plasma
treatment for surface cleaning and modi"cation of metallic bio-
materials. J Biomed Mater Res 1997;35:49}73.
K. Anselme / Biomaterials 21 (2000) 667}681
677

[44] Ask M, Lausmaa J, Kasemo B. Preparation and surface spectro-
scopic characterization of oxide "lms on Ti6Al4V. Appl Surf Sci
1988;35:283}301.
[45] Kasemo B, Lausmaa J. Material}tissue interfaces: the role of
surface properties and processes. Environ Health Perspect
1994;102:41}5.
[46] Healy KE, Thomas CH, Rezania A, et al. Kinetics of bone cell
organization and mineralization on materials with patterned
surface chemistry. Biomaterials 1996;17:195}208.
[47] Thomas CH, McFarland CD, Jenkins ML, Rezania A, Steele JG,
Healy KE. The role of vitronectin in the attachment and spatial
distribution of bone-derived cells on materials with patterned
surface chemistry. J Biomed Mater Res 1997;37:81}93.
[48] Meyle J, GuKltig K, Wolburg H, Von Recum AF. Fibroblast
anchorage to microtextured surfaces. J Biomed Mater Res 1993;
27:1553}7.
[49] Kieswetter K, Schwartz Z, Hummert TW, et al. Surface rough-
ness modulates the local production of growth factors and
cytokines by osteoblast-like MG-63 cells. J Biomed Mater Res
1996;32:55}63.
[50] Anselme K, Bigerelle M, Noel B, et al. Qualitative and quantita-
tive study of human osteoblast adhesion on materials with
various surface roughness. J Biomed Mater Res 2000;49:155}66.
[51] Chehroudi B, Gould TRL, Brunette DM. E!ects of a grooved
titanium-coated implant surface on epithelial cell behavior
in vitro and in vivo. J Biomed Mater Res 1989;23:1067}85.
[52] Den Braber ET, De Ruijter JE, Smits HTJ, Ginsel LA, Von
Recum AF, Jansen JA. E!ect of parallel surface microgrooves
and surface energy on cell growth. J Biomed Mater Res 1995;
29:511}8.
[53] Schwartz Z, Kieswetter K, Dean DD, Boyan BD. Underlying
mechanisms at the bone}surface interface during regeneration.
J Periodontal Res 1997;32:166}71.
[54] Wennerberg A, Ektessabi A, Albrektsson T, Johansson C, An-
dersson B. A 1-year follow-up of implants of di!erent surface
roughness placed in rabbit bone. Int J Oral Maxillofac Implants
1997;12:486}94.
[55] Altankov G, Groth T. Reorganization of substratum-bound
"bronectin on hydrophilic and hydrophobic materials is related
to biocompatibility. J Mater Sci Mater Med 1994;5:732}7.
[56] Ruardy TG, Schakenraad JM, Van der Mei HC, Busscher HJ.
Adhesion and spreading of human skin "broblasts on
physicochemically characterized gradient surface. J Biomed Ma-
ter Res 1995;29:1415}23.
[57] Shelton RM, Rasmussen AC, Davies JE. Protein adsorption at
the interface between charged polymer substrata and migrating
osteoblasts. Biomaterials 1998;9:24}9.
[58] Steele JG, McFarland C, Dalton BA, et al. Attachment of human
bone cells to tissue culture polystyrene and to unmodi"ed poly-
styrene: the e!ect of surface chemistry upon initial cell attach-
ment. J Biomater Sci Polym Ed 1993;5:245}57.
[59] Iuliano DJ, Saavedra SS, Truskey GA. E!ect of the confronta-
tion and orientation of adsorbed "bronectin on endothelial cell
spreading and the strength of adhesion. J Biomed Mater Res
1993;27:1103}13.
[60] Lampin M, Warocquier-CleHrout R, Legris C, Degrange M,
Sigot-Luizard MF. Correlation between substratum roughness
and wettability, cell adhesion, and cell migration. J Biomed
Mater Res 1997;36:99}108.
[61] Sinha RK, Morris F, Shah SA, Tuan RS. Surface composition of
orthopaedic implant metals regulates cells attachment, spread-
ing, and cytoskeletal organization of primary human osteoblasts
in vitro. Clin Orthop Relat Res 1994;305:258}72.
[62] KoKnoKnen M, Hormia M, Kivilahti J, Hautaniemi J, Thesle! I.
E!ect of surface processing on the attachment orientation,
and proliferation of human gingival "broblasts on titanium.
J Biomed Mater Res 1992;26:1325}41.
[63] Hayman EG, Pierschbacher MD, Suzuki S, Ruoslahti E. Vitro-
nectin*a major cell attachment-promoting protein in fetal bo-
vine serum. Exp Cell Res 1985;160:245}58.
[64] Martin JY, Schwartz Z, Hummert TW, et al. E!ect of titanium
surface roughness on proliferation, di!erentiation, and protein
synthesis of human osteoblast-like cells (MG63). J Biomed
Mater Res 1995;29:389}401.
[65] Puleo DA, Preston KE, Sha!er JB, Bizios R. Examination of
osteoblast}orthopaedic biomaterial interactions using molecu-
lar techniques. Biomaterials 1993;14:111}4.
[66] Ozawa S, Kasugai S. Evaluation of implant materials (hy-
droxyapatite, glass-ceramics, titanium) in rat bone marrow
stromal cell culture. Biomaterials 1996;17:23}9.
[67] Boyan BD, Batzer R, Kieswetter K, et al. Titanium surface
roughness alters responsiveness of MG63 osteoblast-like cells to
1
a,25-(OH)2D3. J Biomed Mater Res 1998;39:77}85.
[68] Zreiqat H, Howlett CR. Titanium substrata composition in-
#uences osteoblastic phenotype: in vitro study. J Biomed Mater
Res 1999;47:360}6.
[69] Zreiqat H, Evans P, Howlett CR. E!ect of surface chemical
modi"cation of bioceramic on phenotype of human-bone de-
rived cells. J Biomed Mater Res 1999;44:389}96.
[70] Reich KM, Gay CV, Frangos JA. Fluid shear stress as a medi-
ator of osteoblast cyclic adenosine monophosphate production.
J Cell Physiol 1990;143:100}4.
[71] Hung CT, Pollack SR, Reilly TM, Brighton CT. Real-time
calcium response of cultured bone cells to #uid #ow. Clin Or-
thop Relat Res 1995;313:256}69.
[72] Williams JL, Iannotti JP, Ham A, Bleuit J, Chen JH. E!ects of
#uid shear stress on bone cells. Biorheology 1994;31:163}70.
[73] Stanford CM, Morcuende JA, Brand RA. Proliferative and
phenotypic responses of bone-like cells to mechanical deforma-
tion. J Orthopaed Res 1995;13:664}70.
[74] Buckley MJ, Banes AJ, Levin LG, et al. Osteoblasts increase
their rate of division and align in response to cyclic, mechanical
tension in vitro. Bone and Mineral 1988;4:225}36.
[75] Miyajima K. E!ects of periodic tension on osteoblast-like cells
for cell di!erentiation and alkaline phosphatase activity. Nip-
pon Kyosei Shika Gakkai Zasshi 1990;49:226}36.
[76] Neidlinger-Wilke C, Wilke HJ, Claes L. Cyclic stretching of
human osteoblasts a!ects proliferation and metabolism: a new
experimental method and its application. J Orthopaed Res
1994;12:70}8.
[77] Zhuang H, Wang W, Tahernia AD, Levitz CL, Luchetti WT,
Brighton CT. Mechanical strain-induced proliferation of os-
teoblastic cells parallels increased TGF-beta 1 mRNA. Biochem
Biophys Res Commun 1996;229:449}53.
[78] Fermor B, Gundle R, Evans M, Emerton M, Pocock A, Murray
D. Primary human osteoblast proliferation and prostaglandin
E2 release in response to mechanical strain in vitro. Bone 1998;
22:637}43.
[79] Keila S, Pitaru S, Grosskopf A, Weinreb M. Bone marrow from
mechanically unloaded rat bone expresses reduced osteogenic
capacity in vitro. J Bone Miner Res 1994;9:321}7.
[80] Davies JE, Matsuda T. Extracellular matrix production by os-
teoblasts on bioactive substrata in vitro. Scanning Microsc
1988;2:1445}52.
[81] Hillsley MV, Frangos JA. Alkaline phosphatase in osteoblasts is
down-regulated by pulsatile #uid #ow. Calcif Tissue Int 1997;
60:48}53.
[82] Roelofsen J, Klein-Nulend J, Burger EH. Mechanical stimula-
tion
by
intermittent
hydrostatic
compression
promotes
bone-speci"c gene expression in vitro. J Biomech 1995;28:
1493}503.
[83] Toma CD, Ashkar S, Gray ML, Scha!er JL, Gerstenfeld LC.
Signal transduction of mechanical stimuli is dependent on
micro"lament integrity: identi"cation of osteopontin as a
678
K. Anselme / Biomaterials 21 (2000) 667}681

mechanically induced gene in osteoblasts. J Bone Miner Res
1997;12:1626}36.
[84] Owan I, Burr DB, Turner CH, et al. Mechanotransduction in
bone: osteoblasts are more responsive to #uid forces than mech-
anical strain. Am J Physiol 1997;273:C810}5.
[85] Klein-Nulend J, Roelofsen J, Semeins CM, Bronckers AL, Bur-
ger EH. Mechanical stimulation of osteopontin mRNA expres-
sion and synthesis in bone cell cultures. J Cell Physiol 1997;
170:174}81.
[86] Mikuni-Takagaki Y, Suzuki Y, Kawase T, Saito S. Distinct
responses of di!erent populations of bone cells to mechanical
stress. Endocrinology 1996;137:2028}35.
[87] Harter LV, Hruska KA, Duncan RL. Human osteoblast-like
cells repond to mechanical strain with increased bone matrix
production independent of hormonal regulation. Endocrinology
1995;136:528}35.
[88] Maezzini MC, Toma CD, Scha!er JL, Gray ML, Gerstenfeld
LC. Osteoblast cytoskeletal modulation in response to mechan-
ical strain in vitro. J Orthopaed Res 1998;16:170}80.
[89] Guignandon A, Usson Y, Laroche N, et al. E!ects of intermittent
or continuous gravitational stresses on cell-matrix adhesion:
quantitative analysis of focal contacts in osteoblastic ROS
17/2.8 cells. Exp Cell Res 1997;236:66}75.
[90] Brighton CT, Fisher J, Rush S, et al. The biochemical pathway
mediating the proliferative response of bone cells to a mechan-
ical stimulus. J Bone Jt Surg 1996;78-A:1337}47.
[91] Ngan P, Saito S, Saito M, Lanese R, Shanfeld J, Davidovitch Z.
The interactive e!ects of mechanical stress and interleukin-1
beta on prostaglandin E2 and cyclic AMP production in human
periodontal ligament "broblasts in vitro: comparison with
cloned osteoblastic cells of mouse (MC3T3-E1). Arch Oral Biol
1990;35:717}25.
[92] Ozawa H, Imamura K, Abe E, et al. E!ect of a continuously
applied compressive pressure on mouse osteoblast-like cells
(MC3T3-E1) in vitro. J Cell Physiol 1990;142:177}85.
[93] Guignandon A, Vico L, Alexandre C, Lafage-Proust M-H.
Shape changes of osteoblastic cells under gravitational vari-
ations during parabolic #ight-relationship with PGE2 synthesis.
Cell Struct Funct 1995;20:369}75.
[94] Ajubi NE, Klein-Nulend J, Nijweide PJ, Vrijheid-Lammers T,
Alblas MJ, Burger EH. Pulsating #uid #ow increases prosta-
glandin production by cultured chicken osteocytes-a cyto-
skeleton-dependent process. Biochem Biophys Res Commun
1996;225:62}8.
[95] Smalt R, Mitchell FT, Howard RL, Chambers TJ. Induction of
NO and prostaglandin E2 in osteoblasts by wall-shear stress but
not mechanical strain. Am J Physiol 1997;273:E751}8.
[96] Carvalho RS, Scott JE, Suga DM, Yen EHK. Stimulation of
signal transduction pathways in osteoblasts by mechanical
strain potentiated by parathyroid hormone. J Bone Miner Res
1994;9:999}1011.
[97] Pommerenke H, Schreiber E, Durr F, et al. Stimulation of
integrin receptors using a magnetic drag force device induces
intracellular free calcium response. Eur J Cell Biol 1996;
70:157}64.
[98] Carvalho RS, Scott JE, Yen EH. The e!ects of mechanical
stimulation on the distribution of
b1-integrin and expression of
b1-integrin mRNA in TE-85 human osteosarcoma cells. Arch
Oral Biol 1995;40:257}64.
[99] Schmidt C, Pommerenke H, Durr F, Nebe B, Rychly J. Mechan-
ical stressing of integrin receptors induces enhanced tyrosine
phosphorylation of cytoskeletally anchored proteins. J Biol
Chem 1998;273:5081}5.
[100] Salter DM, Robb JE, Wright MO. Electrophysiological re-
sponses of human bone cells to mechanical stimulation: evidence
for speci"c integrin function in mechanotransduction. J Bone
Miner Res 1997;12:1133}41.
[101] Carvalho RS, Scha!er JL, Gerstenfeld LC. Osteoblasts induce
osteopontin expression in response to attachment on "bronec-
tin: demonstration of a common role for integrin receptors in the
signal transduction processes of cell attachment and mechanical
stimulation. J Cell Biochem 1998;70:376}90.
[102] Ziambaras K, Lecanda F, Steinberg TH, Civitelli R. Cyclic
stretch enhances gap junctional communication between os-
teoblastic cells. J Bone Miner Res 1998;13:218}28.
[103] Jorgensen NR, Geist ST, Civitelli R, Steinberg TH. ATP- and
gap junction-dependent intercellular calcium signaling in os-
teoblastic cells. J Cell Biol 1997;139:497}506.
[104] Rawlinson SC, Pitsillides AA, Lanyon LE. Involvement of di!er-
ent ion channels in osteoblasts' and osteocytes' early responses
to mechanical strain. Bone 1996;19:609}14.
[105] Davidson RM, Lingenbrink PA, Norton LA. Continuous mech-
anical loading alters properties of mechanosensitive channels in
G292 osteoblastic cells. Calcif Tissue Int 1996;59:500}4.
[106] Banes AJ, Tsuzaki M, Yamamoto J, et al. Mechanoreception at
the cellular level: the detection, interpretation, and diversity
responses to mechanical signals. Biochem Cell Biol 1995;
73:349}65.
[107] Lo CM, Glogauer M, Rossi M, Ferrier J. Cell}substrate separ-
ation: e!ect of applied force and temperature. Eur Biophys
J Biophys Lett 1998;27:9}17.
[108] Garcia AJ, Ducheyne P, Boettiger D. E!ects of surface reaction
stage on "bronectin-mediated adhesion of osteoblast-like cells
to bioactive glass. J Biomed Mater Res 1998;40:48}56.
[109] Garcia AJ, Ducheyne P, Boettiger D. Quanti"cation of cell
adhesion using a spinning disc device and application to sur-
face-reactive materials. Biomaterials 1998;18:1091}8.
[110] Bowers VM, Fisher LR, Francis GW, Williams KL. A micro-
mechanical technique for monitoring cell}substrate adhesive-
ness: measurements of the strengh of red blood cell adhesion to
glass and to polymer test surfaces. J Biomed Mater Res 1989;
23:1453}73.
[111] Hochmuth RM. Measuring the mechanical properties of indi-
vidual human blood cells. J Biomech Eng 1993;115:515}9.
[112] Sato M, Ohshima N, Nerem RM. Viscoelastic properties of
cultured porcine aortic endothelial cells exposed to shear stress.
J Biomech 1996;29:461}7.
[113] Thoumine O, Ott A. Comparison of the mechanical properties of
normal and transformed "broblasts. Biorheology 1997;34:
309}26.
[114] Corry WD, Defendi V. Centrifugal assessment of cell adhesion.
J Biochem Biophys Meth 1981;4:29}38.
[115] Thoumine O, Ott A, Louvard D. Critical centrifugal forces
induce adhesion rupture or structural reorganization in cultured
cells. Cell Motility Cytoskel 1996;33:276}87.
[116] Lotz MM, Burdsal CA, Erickson HP, McClay DR. Cell ad-
hesion to "bronectin and tenascin: quantitative measurements
of initial binding and subsequent strengthening response. J Cell
Biol 1989;109:1795}805.
[117] Glogauer M, Ferrier J, McCulloch CA. Magnetic "elds
applied to collagen-coated ferric oxide beads induce stretch-
activated Ca
2`
#ux
in "broblasts. Am J Physiol 1995;269:
C1093}104.
[118] Bausch AR, Ziemann F, Boulbitch AA, Jacobson K, Sackmann
E. Local measurements of viscoelastic parameters of adherent
cell surfaces by magnetic bead microrheometry. Biophys
J 1998;75:2038}49.
[119] Mege JL, Capo C, Benoliel AM, Bongrand P. Determination of
binding strength and kinetics of binding initiation. A model
study made on the adhesive properties of P388D1 macrophage-
like cells. Cell Biophys 1986;8:141}60.
[120] Truskey GA, Proulx TL. Relationship between 3T3 cells spread-
ing and the strength of adhesion on glass and silane surfaces.
Biomaterials 1993;14:243}54.
K. Anselme / Biomaterials 21 (2000) 667}681
679

[121] Burmeister JS, Vrany JD, Reichert WM, Truskey GA. E!ect of
"bronectin amount and conformation on the strength of endo-
thelial cell adhesion to HEMA/EMA copolymers. J Biomed
Mater Res 1996;30:13}22.
[122] Anselme K, Lanel B, Gentil C, Hardouin P, Marie PJ, Sigot-
Luizard MF. Bone organotypic culture method: a model for
cytocompatibility
testing
of
biomaterials.
Cells
Mater
1994;4:113}23.
[123] Horbett TA, Waldburger JJ, Ratner BD, Ho!man AS. Cell
adhesion to a series of hydrophilic}hydrophobic copolymers
studied with a spinning disc apparatus. J Biomed Mater Res
1988;22:383}404.
[124] Thoumine O, Ott A. Time scale dependent viscoelastic and
contractile regimes in "broblasts probed by microplate manip-
ulation. J Cell Sci 1997;110:2109}16.
[125] Yamamoto A, Mishima S, Maruyama N, Sumita M. A new
technique for direct measurement of the shear force necessary to
detach a cell from a material. Biomaterials 1998;19:871}9.
[126] Jones DB, Nolte H, Scholubbers JG, Turner E, Veltel D. Bio-
chemical signal transduction of mechanical strain in osteoblast-
like cells. Biomaterials 1991;12:101}10.
[127] Burmeister JS, Olivier LA, Reichert WM, Truskey GA. Applica-
tion of total internal re#ection #uorescence microscopy to study
cell adhesion to biomaterials. Biomaterials 1998;19:307}25.
[128] Urist MR. Bone formation by autoinduction. Science 1965;
150:893}9.
[129] Sakou T. Bone morphogenetic proteins: from basic studies to
clinical approaches. Bone 1998;22:591}603.
[130] Yasko AW, Lane JM, Fellinger EJ, Rosen V, Wozney JM, Wang
EA. The healing of segmental bone defects, induced by recom-
binant human bone morphogenetic protein (rhBMP-2). J Bone
Jt Surg 1992;74-A:659}70.
[131] Cook SD, Dalton JE, Tan EH, Whitecloud TS, Rueger DC. In
vivo evaluation of recombinant human osteogenic protein
(rhOP-1) implants as a bone grafts substitute for spinal fusions.
Spine 1994;19:1655}63.
[132] Cook SD, Ba!es GC, Wolfe MW, Sampath KT, Rueger DC,
Whitecloud TS. The e!ect of recombinant human osteogenic
protein-1 on healing of large segmental bone defects. J Bone Jt
Surg 1994;76-A:827.
[133] Gerhart TN, Kirker-Head CA, Kriz MJ, et al. Healing seg-
mental femoral defects in sheep using recombinant human
bone morphogenetic protein. Clin Orthop Relat Res 1993;293:
317}26.
[134] Johnson EE, Urist MR. One-stage lengthening of femoral
nonunion augmented with human bone morphogenetic protein.
Clin Orthop Relat Res 1998;347:105}16.
[135] Dee KC, Rueger DC, Andersen TT, Bizios R. Conditions which
promote mineralization at the bone-implant interface: a model
in vitro study. Biomaterials 1996;17:209}15.
[136] Dee KC, Bizios R. Mini-review: Proactive biomaterials and bone
tissue engineering. Biotechnol Bioeng 1996;50:438}42.
[137] Qian JJ, Bhatnagar RS. Enhanced cell attachment to anorganic
bone mineral in the presence of a synthetic peptide related to
collagen. J Biomed Mater Res 1996;31:545}54.
[138] McDavid PT, Boone ME, Kafrawy AH, Mitchell DF. E!ect of
autologous marrow and calcitonin on reactions to a ceramic.
J Dent Res 1979;58:1478}83.
[139] Nade S, Armstrong L, McCartney E, Baggaley B. Osteogenesis
after bone and bone marrow transplantation. Clin Orthop Relat
Res 1983;181:255}63.
[140] Ohgushi H, Goldberg VM, Caplan AI. Heterotopic osteogenesis
in porous ceramics induced by marrow cells. J Orthopaed Res
1989;7:568}78.
[141] Wittbjer J, Palmer B, Rohlin M, Thorngren K-G. Osteogenic
activity in composite grafts of demineralized compact bone and
marrow. Clin Orthop Relat Res 1983;173:229}38.
[142] Ohgushi H, Goldberg VM, Caplan AI. Repair of bone defects
with marrow cells and porous ceramic. Acta Orthop Scand
1989;60:334}9.
[143] Grundel RE, Chapman MW, Yee T, Moore DC. Autogeneic
bone marrow and porous biphasic calcium phosphate ceramic
for segmental bone defects in the canine ulna. Clin Orthop Relat
Res 1991;266:244}58.
[144] Werntz JR, Lane JM, Burstein AH, Justin R, Klein R, Tomin E.
Qualitative and quantitative analysis of orthotopic bone regen-
eration by marrow. J Orthopaed Res 1996;14:85}93.
[145] Owen M. Marrow stromal stem cells. J Cell Sci 1988;10:63}76.
[146] Caplan AI. Mesenchymal stem cells. J Orthopaed Res 1991;
9:641}50.
[147] Bab I, Howlett CR, Ashton BA, Owen ME. Ultrastructure of
bone and cartilage formed in vivo in di!usion chambers. Clin
Orthop Relat Res 1984;187:243}54.
[148] Bab I, Passi-Even L, Gazit D, et al. Osteogenesis in in vivo
di!usion chamber cultures of human marrow cells. Bone and
Mineral 1988;4:373}86.
[149] Holden CM, Bernard GW. Ultrastructural in vitro characteriza-
tion of a porous hydroxyapatite/bone cell interface. J Oral
Implantol 1990;XVI:86}95.
[150] Bagambisa FB, Joos U. Preliminary studies on the phenom-
enological cultured on hydroxyapatite ceramics. Biomaterials
1990;11:50}5.
[151] Cheung HS, Haak MH. Growth of osteoblasts on porous cal-
cium phosphate ceramic: an in vitro model for biocompatibility
study. Biomaterials 1989;10:63}7.
[152] Uchida A, Nade S, McCartney E, Ching W. Growth of bone
marrow cells on porous ceramics in vitro. J Biomed Mater Res
1987;21:1}10.
[153] Casser-Bette M, Murray AB, Closs EI, Er#e V, Schmidt J. Bone
formation by osteoblast-like cells in a three dimensional cell
culture. Calcif Tissue Int 1990;46:46}56.
[154] Schoeters G, Leppens H, Van Gorp U, Van Den Heuvel R.
Haemopoietic long-term bone marrow cultures from adult mice
show osteogenic capacity in vitro on 3-dimensional collagen
sponges. Cell Prolif 1992;25:587}603.
[155] Begley CT, Doherty MJ, Hankey DP, Wilson DJ. The culture of
human osteoblasts upon bone graft substitutes. Bone 1993;
14:661}6.
[156] Faucheux C, Bareille R, Rouais F, Amedee J, LiebendoKrfer A,
Dard M. Biocompatibility testing of a bovine hydroxy-apatite
ceramic material with the use of osteo-progenitor cells isolated
from human bone marrow. J Mater Sci Mater Med 1994;5:635}9.
[157] Ishaug SL, Crane GM, Miller MJ, Yasko AW, Yaszemski MJ,
Mikos AG. Bone formation by three-dimensional stromal os-
teoblast culture in biodegradable polymer sca!olds. J Biomed
Mater Res 1997;36:17}28.
[158] Lu JX, Flautre B, Anselme K, et al. Study of porous interconnec-
tions of bioceramic on cellular rehabitation in vitro and in vivo.
In: Sedel L, Rey C, editors. Bioceramics 10. Paris: Pergamon,
Elsevier Science, 1997. p. 583}6.
[159] Nakahara H, Goldberg VM, Caplan AI. Culture-expanded hu-
man periosteal-derived cells exhibit osteochondral potential in
vivo. J Orthopaed Res 1991;9:465}76.
[160] Gundle R, Joyner CJ, Tri$tt JT. Interactions of human osteo-
progenitors with porous ceramic following di!usion chamber
implantation in a xenogeneic host. J Mater Sci Mater Med
1997;8:519}23.
[161] Hayesworth SE, Goshima J, Goldberg VM, Caplan AI. Charac-
terization of cells with osteogenic potential from human mar-
row. Bone 1992;13:81}8.
[162] Nakahara H, Goldberg VM, Caplan AI. Culture-expanded per-
iosteal-derived cells exhibit osteochondrogenic potential in por-
ous calcium phosphate ceramics in vivo. Clin Orthop Relat Res
1992;276:291}8.
680
K. Anselme / Biomaterials 21 (2000) 667}681

[163] Frayssinet P, Autefage A. MateHriaux hybrides de substitution du
tissu osseux. Rev Rhum [Ed Fr] 1993;60:342}51.
[164] Frayssinet P, Primout I, Rouquet N, Autefage A, Guilhem A,
Bonnevialle P. Bone cell grafts in bioreactor: a study of feasibil-
ity of bone cell autograft in large defects. J Mater Sci Mater Med
1991;2:217}21.
[165] Yoshikawa T, Ohgushi H, Tamai S. Immediate bone forming
capability of prefabricated osteogenic hydroxyapatite. J Biomed
Mater Res 1996;32:481}92.
[166] Goshima J, Goldberg VM, Caplan AI. The osteogenic potential
of culture-expanded rat marrow mesenchymal cells assayed
in vivo in calcium phosphate ceramic blocks. Clin Orthop Relat
Res 1991;262:298}311.
[167] Ishaug-Riley SL, Crane GM, Gurlek A, et al. Ectopic bone
formation by marrow stromal osteoblast transplantation using
poly(
DL
-lactic-co-glycolic acid) foams implanted into rat mesen-
tery. J Biomed Mater Res 1997;36:1}8.
K. Anselme / Biomaterials 21 (2000) 667}681
681
Wyszukiwarka
Podobne podstrony:
Increased osteoblast adhesion on nanophase metals
Adhesive capsulitis (2009)
Effective antibacterial adhesive coating on cotton fabric using ZnO
Adhesion
Pressure Sensitive Adhesives
Adhesive Compounds
Adhesive capsulitis (2009)
Osteoblast response
Influence of titanium surfaces on attachment of osteoblast
The proliferation and phenotypic expression of human osteobl
Sp Sh 02 Pawns B Adhesive
CSB 1055 2 Dust Under Main Lens Double Sided Adhesive
Adhesion of bioactive
A Confocal Laser Scanning Microscope investigation of different dental adhesives bonded to root cana
więcej podobnych podstron