
Abstract The present study investigated how muscle fa-
tigue influences single degree-of-freedom elbow flexion
movements and their associated patterns of phasic mus-
cle activation. Maximal unfatigued voluntary isometric
elbow flexor and extensor joint torque was measured at
the beginning of the experiment. Subjects then per-
formed elbow flexion movements over two distances as
fast as possible, and movements over the longer distance
at an intentionally slower speed. The slower speed was
close to what would become the maximal speed in the
fatigued state. Subjects then performed a fatiguing proto-
col of 20 sustained isometric flexion contractions of 25 s
duration with 5 s rest at 50% maximal unfatigued volun-
tary force. After a recovery period they repeated the
movements. The fatigue protocol was successful in in-
ducing muscle fatigue, the evidence being decreased iso-
metric maximal joint torque of over 20%. Fatigued
movements had lower peak muscle torque and speed.
Our principal finding was of changes in the timing of the
phasic patterns of fatigued muscle activation. There was
an increase in the duration of the agonist burst and a de-
lay in the timing of the antagonist muscle as measured
by the centroid of the EMG signals. We conclude that
these changes serve as partial but incomplete, centrally
driven compensation for fatigue induced changes in
muscle function. An additional, unexpected finding was
how small an effect fatigue had on movement perfor-
mance when using a recovery time of 10 min that is long
enough to allow muscle membrane conduction velocity
to return to normal. This raises questions concerning the
behavioral significance of classical laboratory studies of
human fatigue mechanisms.
Keywords Motor control · Fatigue · Electromyography ·
Movement · Neural control
Introduction
Motor control models help us relate changes in move-
ment task to predictable changes in EMG pattern. For
example, movements of longer distances or with heavier
loads are associated with longer and larger agonist EMG
bursts and delayed antagonist muscle activation (Berardelli
et al. 1984; Gottlieb et al. 1989; Pfann et al. 1998).
Movements performed over the same distance that are
made more quickly are associated with larger, more
steeply rising EMG bursts and earlier antagonist activa-
tion (Mustard and Lee 1987; Corcos et al. 1989). These
studies changed movement by instruction or external
conditions such as load, target position or target size.
Movement also changes when muscles fatigue, a condi-
tion deliberately avoided in the studies cited above. Here
we raise the question of whether, to reduce the kinematic
consequences of muscle fatigue, there are compensatory
neural adaptations that modify muscle activation pat-
terns. If so, are those changes predictable from studies of
unfatigued movement?
There is much research on the neural mechanisms that
underlie muscle fatigue (Gandevia et al. 1995b). Most
D.M. Corcos (
✉
) · J. Wilding
School of Kinesiology (M/C 194),
University of Illinois at Chicago, 901 West Roosevelt Road,
Chicago, IL 60680, USA
e-mail: dcorcos@uic.edu
Tel.: +1-312-3551708, Fax: +1-312-3552305
D.M. Corcos
Department of Psychology, University of Illinois at Chicago,
Chicago, IL 60680, USA
D.M. Corcos
Department of Neurological Sciences, Rush Medical College,
Chicago, IL 60612, USA
H.-Y. Jiang
Department of Speech-Language Pathology,
University of Toronto, 6 Queen's Park Crescent West, Toronto,
Ontario M5S 3H2, USA
J. Wilding
Department of Physical Therapy and Human Movement Sciences,
Northwestern University Medical School, Chicago, IL 60611,
USA
G.L. Gottlieb
NeuroMuscular Research Center, Boston University,
19 Deerfield Street, Boston, MA 02215, USA
Exp Brain Res (2002) 142:1–12
DOI 10.1007/s00221-001-0904-9
R E S E A R C H A R T I C L E
Daniel M. Corcos · Hai-Ying Jiang · Janey Wilding
Gerald L. Gottlieb
Fatigue induced changes in phasic muscle activation patterns
for fast elbow flexion movements
Received: 3 January 2001 / Accepted: 7 September 2001 / Published online: 20 November 2001
© Springer-Verlag 2001

studies examine changes in the electromyogram during
steady state isometric contractions (Bigland-Ritchie et al.
1983b; Marsden et al. 1983; Garland et al. 1994; cf.
Enoka and Stuart 1992), whereas relatively few studies
have addressed changes that occur during movement.
Berardelli and colleagues (1984), and Tschoepe et al.
(1994) found fatigue induced slowing and increased the
duration of the first agonist EMG burst. They suggested
that the increase in the duration of the first agonist burst
partially compensates for the decrease in maximal moto-
neuron firing frequency that had been observed in iso-
metric contractions (Bigland-Ritchie et al. 1983a). Some
recent studies, however, suggest that motor unit firing
rates can increase during fatigue (Miller et al. 1996), and
fire at very short interspike intervals (Griffin et al. 1998).
Lucidi and Lehman (1992) found that although the kine-
matics of the movement after an hour of recovery were
not distinguishable from those made before the fatiguing
task, there remained an increase in the width of the first
agonist burst. All three studies that investigated the time
course of the agonist EMG suggest that fatigue causes
changes in the temporal profile of the agonist electromy-
ogram and, if the fatigue is great enough and the recov-
ery interval is not too long, a slowing of the movements.
The present study was intended to test three hypothe-
ses related to the effects of muscle fatigue on patterns of
muscle activation and movement performance. The first
is that the way the CNS compensates for fatigue-induced
muscle weakness is similar to its compensation for a
heavier load. In both cases, we postulate that there are
similar changes in the patterns of muscle activation to in-
crease or maintain force output. These changes are pro-
longation of agonist activation, delay of antagonist acti-
vation and an increase in the peak amplitude of the ago-
nist EMG with no change in the rate of rise of agonist
muscle activation. This strategy, which we have termed a
speed insensitive strategy (Gottlieb et al. 1989) for con-
trolling movement distance as well as controlling load
changes in unfatigued muscle, does not preserve move-
ment time and therefore is an incomplete compensation
for changing task conditions. The rationale for this hy-
pothesis is based in part on previous studies showing fa-
tigue induced temporal changes in agonist and antagonist
EMG waveforms. We tested a second hypothesis that the
EMG compensation under fatigued conditions would be
greater for short movements than for long movements
but that the reduction in peak velocity would be greater
for long movements than for short ones. The rationale is
that because short movements need lower forces and use
less muscle activation, additional motor units might be
available for compensatory recruitment. This is based on
previous studies that have shown EMG increases for
submaximal isometric contractions (Kirsch and Rymer
1987), and additional motor units being recruited in sub-
maximal isotonic tasks (Miller et al. 1996). We tested a
third hypothesis that the increase in agonist duration ob-
served during fatigue would not be observed for unfa-
tigued movements that were intentionally slowed to a fa-
tigued speed. The rationale for this hypothesis is that the
neural control signals associated with weakness induced
by neuromuscular fatigue are different from those asso-
ciated with intentional reductions in movement speed.
Materials and methods
Subjects
Eight male subjects were used in this study. Males were selected
because in laboratory protocols, they fatigue more rapidly than fe-
males (Scalzitti 1994). Our subjects were between the ages of 21
and 33 years, in good health, and without any history of joint or
neuromuscular disease. They performed elbow flexion isometric
and isotonic contractions and elbow extension isometric contrac-
tions with their right arms in the horizontal plane. All subjects
gave informed consent according to University IRB protocols be-
fore participation in the experiment.
Equipment
A manipulandum was used to support the subject's forearm and re-
strict movement to one degree of freedom. A capacitative transducer
on the axis of rotation of the manipulandum measured angular dis-
placement. Joint acceleration was measured by a piezoresistive ac-
celerometer mounted 47.6 cm from the center of rotation. A torque
transducer was attached to the manipulandum. A torque motor was
used to move the manipulandum so that the moment of inertia of
each subject's forearm could be measured. Joint velocity was com-
puted from the measured angle. Pairs of pediatric EKG electrodes
were placed 2 cm apart over the bellies of the biceps brachii, and the
lateral and long heads of triceps to measure the EMG signals that
were amplified (
×
1600) and band pass filtered (60–300 Hz). Joint
angle, acceleration and the EMG signals were digitized with 12-bit
resolution by a data acquisition computer at a rate of 1000/s.
Procedure
The subject sat in a chair with his right arm abducted 90° away
from his body on the manipulandum on which he grasped a verti-
cal handle. The elbow joint was aligned with the rotational axis of
the manipulandum. The manipulandum was locked in place for
isometric contractions and rotated freely when movements were
performed. A weight of 20 lb was added to the end of the manip-
ulandum to increase the moment of inertia of the manipulandum to
2.28 kg.m
2
in order to increase the force requirements during the
movement and thus accentuate the effects of neuromuscular fa-
tigue. Pilot experiments had shown that if the force requirements
of the task are low, fatigue had little effect on either mechanical or
EMG parameters. In addition, our previous work has shown that
the EMG patterns of movements performed against both small and
large inertias are qualitatively the same. The quantitative differ-
ence is that the EMG bursts are longer and larger for larger iner-
tias, and the antagonist is delayed (Gottlieb et al. 1989).
A computer monitor was located in front of the subject. There
was a cursor on the monitor to display the angular position of the
manipulandum and give the subject feedback about the movement.
A narrow green marker on the screen represented the starting posi-
tion. A broad red marker was located as a target at the desired an-
gular distance. The width of the broad marker corresponded to 9°
of angular elbow rotation in all the experiments reported here.
Subjects were instructed that when a computer-generated tone
sounded, they should accurately move to the target zone as quick-
ly as possible. They were asked to perform the following tasks.
Maximal and 50% of maximal isometric contractions
The manipulandum was locked in place at 90°. The subject per-
formed four isometric flexions and four isometric extensions at
2

100% of his maximal voluntary contraction (MVC), and then four
isometric flexions and four isometric extensions at 50% of the just
measured maximal torque. The purpose of measuring 100% MVC
torque was to determine the extent to which fatigue reduces maxi-
mal voluntary torque. The purpose of measuring 50% MVC was
to be able to determine whether contractile fatigue has occurred.
Contractile fatigue would result in an increase in EMG at a given
level of torque (Kirsch and Rymer 1987).
Fast unfatigued flexion
The subject performed 11 voluntary elbow flexions over 20°
(55–75°, 0° being full elbow extension) and over 60° (55–115°) as
fast as possible. The purpose of this was to determine the unfa-
tigued mechanical and EMG parameters of fast voluntary move-
ments over two fixed distances.
Intentionally slowed unfatigued flexion
The subject performed 20 flexions of 60° at a speed that was 10%
less than the unfatigued maximum velocity for the 60° distance.
The purpose of this was to collect data in which speed was inten-
tionally reduced in order to compare these data with movements in
which speed was reduced by fatigue. Pilot studies had shown that
our fatigue protocol for the longer movement distance reduced
peak movement velocity by approximately 10%. The effect of fa-
tigue on peak velocity was less than 10% for the shorter move-
ments, and so we chose not to conduct this experiment at the
shorter distances. To assist the subject, we monitored peak veloci-
ty and reported its value to the subject after each movement, along
with encouragement, if necessary, to move faster or slower.
Isometric fatigue protocol repetition 1
The fatigue protocol consisted of 20 repetitions of a 50% MVC
isometric flexion at the elbow joint for 25 s, followed by 5 s of
rest between the repetitions.
Rest period 1
After the fatigue protocol, the subject rested for ten minutes to al-
low muscle membrane conduction velocity to return to normal
values (Kirsch and Rymer 1987). In another group of four sub-
jects, we used only a 2-min recovery period. We used two recov-
ery time periods so that we could both minimize the effects of re-
covery time on motor performance (2-min recovery protocol), and
collect data in which the EMG signal is not affected by changes in
conduction velocity (10-min recovery protocol). Since the shorter
recovery period causes ambiguities in interpreting EMG changes,
the EMG signal is not analyzed for this recovery time period.
However, the magnitude of the kinematic changes was larger for
the shorter recovery interval and allows us to demonstrate the ef-
fectiveness of this fatigue protocol.
Fast fatigued flexions at distance 1
The subject performed 11 voluntary elbow flexion movements as
fast as possible either over 20° or over 60°. The order in which the
distances were performed was counterbalanced such that half of
the subjects performed the 20° movement before the 60° move-
ment. These movements were analyzed to determine the mechani-
cal and EMG parameters of fatigued muscle when completing vol-
untary movements.
Isometric fatigue protocol repetition 2
The subject repeated the fatigue protocol but did only 11 repeti-
tions since the protocol was quite painful. These 11 repetitions
were intended to restore the muscle's fatigued state.
Rest period 2
The subject rested for the same interval as in Rest period 1 above.
Fast fatigued flexions at distance 2
The subject performed 11 voluntary elbow flexion movements as
fast as possible over either 20° or 60°, whichever distance was not
performed under Fast fatigue flexions above.
Fatigued maximal isometric and 50% isometric
The subject again performed four isometric flexions and four iso-
metric extensions at 100% of his maximal voluntary contraction
(MVC), and then at 50% of his unfatigued MVC.
The protocol developed by Kirsch and Rymer (1987) produces
significant muscle fatigue. Fatigue causes a fall in the mean fre-
quency of the EMG spectrum as a consequence of changes in con-
duction velocity in the muscle fibers. However, Kirsch and Rymer
(1987) showed that 10 min of rest following the fatigue protocol
returns the mean power frequency of the EMG signal to the pre-
fatigue levels in both the biceps and the brachialis muscles. Thus,
after 10 min, any changes in the electromyogram induced by fa-
tigue can be attributed to factors other than changes in conduction
velocity.
Subjects practiced the whole experimental protocol once be-
fore they took part in the experiment. The time interval between
practice and experiment was at least 48 h. Subjects did not do any
intensive exercise before they participated in the experiment.
Data analysis
The digitized EMG signals were full wave rectified and filtered
with a 10-ms moving average window for plotting the EMG time
series data (Fig. 1, Fig. 2, Fig. 5). The data in these figures were
all aligned with respect to the onset of the agonist EMG. The fol-
lowing parameters were calculated.
Isometric parameters
1. Maximal elbow torque (Nm): the maximal elbow torque in the
isometric contraction.
2. Integrated EMG (arbitrary unit): the EMG was integrated over
200 ms centered about the time of peak isometric elbow
torque. We chose this time interval since it was the longest
time interval that all subjects maintained a steady-state maxi-
mum contraction in the fatigued condition.
3. Torque/EMG ratio: the peak of the torque in the 50% isometric
condition divided by the EMG integrated over 200 ms centered
about the time of peak isometric elbow torque. One data set
was lost to equipment malfunction, and so these data were only
collected on seven subjects.
Movement parameters
1. Movement time (ms): the time interval from 1% of peak accel-
eration to the time when the velocity falls to 5% of peak veloc-
ity.
2. Peak velocity (V
max
–deg/s): The largest value of movement ve-
locity.
3

3. Peak elbow torque (Nm): for voluntary movement, elbow
torque was the maximum muscle torque during the accelera-
tion phase of the movement. Elbow torque was calculated by
multiplying acceleration by the effective moment of inertia
(forearm plus manipulandum).
4. Q
30
(arbitrary unit): the integral of the agonist EMG signal
from the visually marked onset to 30 ms thereafter. This pa-
rameter is used to characterize the initial slope of the agonist
EMG burst.
5. Q
ag
(arbitrary unit): the integral of the agonist EMG from the
marked onset to the time of peak velocity. This parameter is
used to characterize the area of the first agonist EMG burst
which is responsible for the limb accelerating towards the tar-
get.
6. Q
ant
(arbitrary unit): the integral of the antagonist EMG from
the marked onset of the agonist burst to the end of the move-
ment (the distance at which velocity drops below 5% of V
max
).
This parameter is used to characterize the area of the antago-
nist burst.
7. Agonist EMG peak amplitude (arbitrary unit): the EMG peak
amplitude was measured as the maximal value in the filtered
and averaged agonist burst.
8. C
ant
ms: the centroid of the antagonist burst. This value is cal-
culated by the following equation:
(1)
u (t)=1 if emg(t)
≥
K emg
max
u (t)=0 if emg(t)<K emg
max
MT is movement time, t
0
is the time of start of acceleration,
emg(t) is the EMG signal in the lateral head of triceps, K is
0.75, emg
max
is the peak EMG of the lateral head of triceps.
This equation resolves the location of the burst and ignores
low level activity (less than Kemg
max
). The algorithm is simi-
lar to locating the peak of the EMG burst but is less sensitive
to the details of the EMG waveform.
9. C
ag
ms: the centroid of the agonist burst. It is computed by
equation 1 with the integration interval bounded by the time of
peak velocity, and K is the same value as used for computing
C
ant
(0.75). The centroid of the agonist burst is a measure of
the duration of the biceps EMG burst.
Statistical analysis
For the maximal voluntary contractions, a paired t-test was per-
formed to examine the effects of fatigue on the maximal elbow
torque and the EMG integral for both flexion and extension con-
tractions. A paired t-test was also used to compare the
torque/EMG ratio in the non-fatigued condition and the fatigued
condition. For the isotonic movements, a two-way repeated-mea-
sures ANOVA was used to examine the effects of fatigue and
movement distance. A paired t-test was performed to compare the
intentionally slowed unfatigued movements with the fatigued
movements.
Results
The results are divided into four parts. Part 1 describes
the effects of fatigue on isometric muscle torque and
EMG. Part 2 describes the effects of fatigue on move-
ment kinematics and EMG patterns. Part 3 compares the
EMG patterns of intentionally slowed unfatigued move-
ments with those of fatigue-slowed movements. Part 4
compares the kinematic effects of a 2-min recovery in-
terval with that of a 10-min recovery interval.
Changes in muscle torque and EMG in maximal
voluntary contractions
On average there was a statistically significant decline of
21.2% in flexion torque [mean±SE pre-fatigue=
69.1±4.2
Nm, fatigued=54.5±4.3
Nm, t(7)=5.98,
P=0.001]. From this fact we conclude that the protocol
developed by Kirsch and Rymer (1987) was effective in
producing fatigue in the agonist biceps muscle. This re-
duction in maximum flexion torque is shown for a repre-
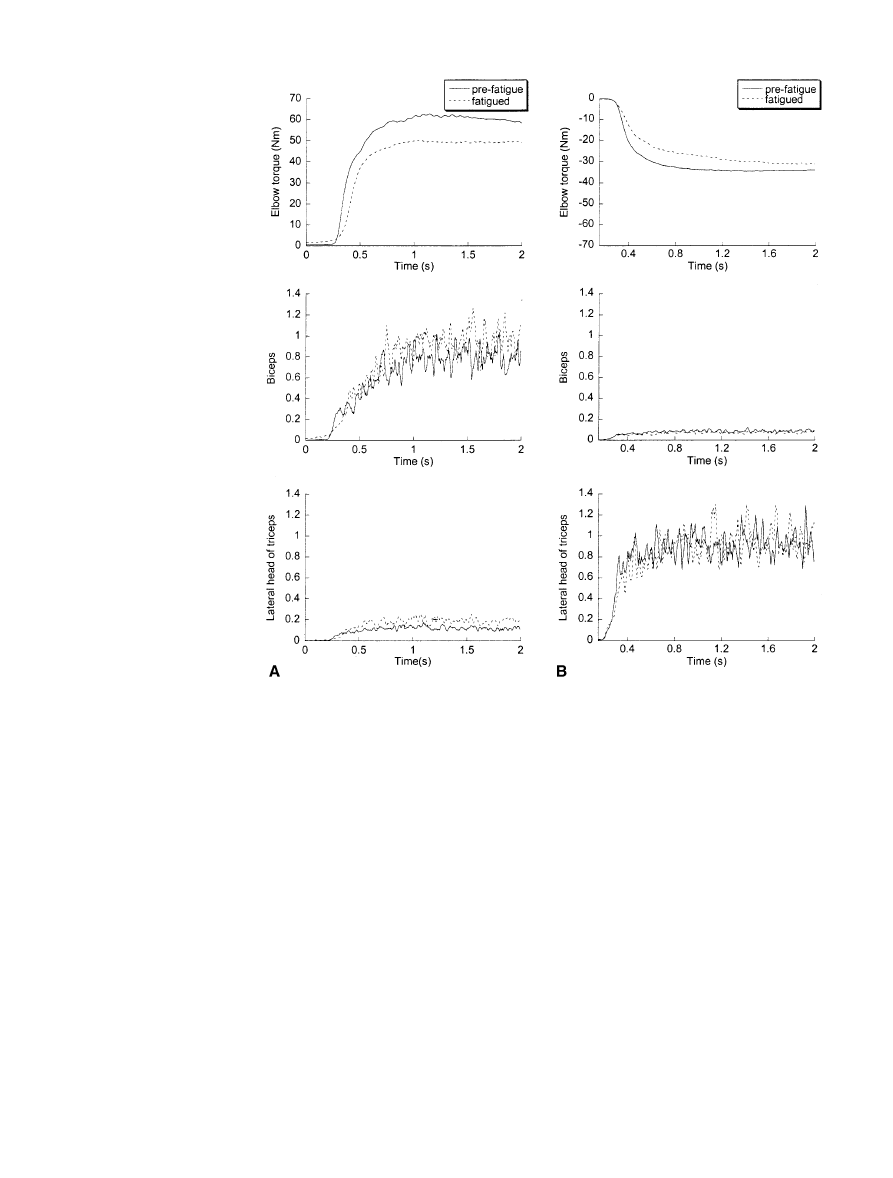
sentative subject in Fig. 1A. Even though the fatigue
protocol did not call for strong contraction of the exten-
sor muscles, maximum extension torque was reduced
4.1% following the fatigue protocol as shown in Fig. 1B,
but the decline was not statistically significant [mean±
SE pre-fatigue=43±2.66 Nm, fatigued=41.2±3 Nm,
t(7)=1.18, P=0.278]. The integrated EMG during MVC
activity was not statistically significantly different be-
tween the fatigued and unfatigued conditions for either
the biceps muscle in flexion [t(7)=1.99, P=0.087] or the
triceps muscle in extension [t(7)=1.82, P=0.111]. In the
agonist muscle, the ratio of torque to EMG, the measure
usually considered the defining characteristic of physio-
logical fatigue, was significantly smaller in the fatigued
than the unfatigued state in the 50% MVC condition
[mean ratio±SE pre-fatigue=85.96±13.32; mean ratio fa-
tigued=55.12±10.3, t(6)=2.73, P=0.03].
Comparison between fatigued movements
and unfatigued movements
Fatigue decreased movement velocity and increased
movement time. The data from one representative sub-
ject are shown in Fig. 2. The peak elbow torque in the
acceleration phase of the movement decreased. The ini-
tial rising phase of the EMG (Q
30
) in the agonist is simi-
lar. However, the rate of rise was not sustained with fa-
tigue and, as a consequence, the EMG peak amplitude of
the biceps muscle decreased. These observations apply
to both 20° and 60° movements. The late component of
the antagonist burst (beginning approximately 160 ms
after the agonist onset) is delayed in both the lateral head
of triceps and the long head of triceps as a consequence
of fatigue.
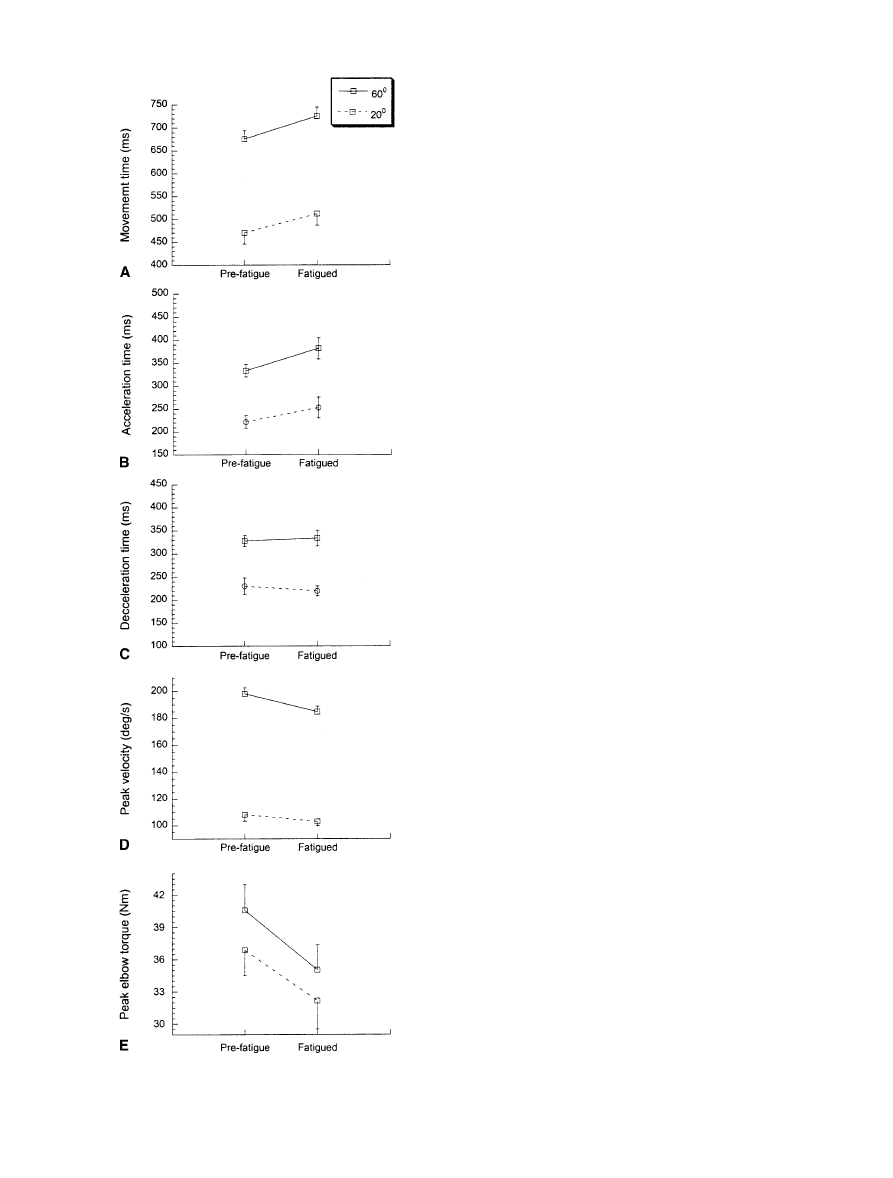
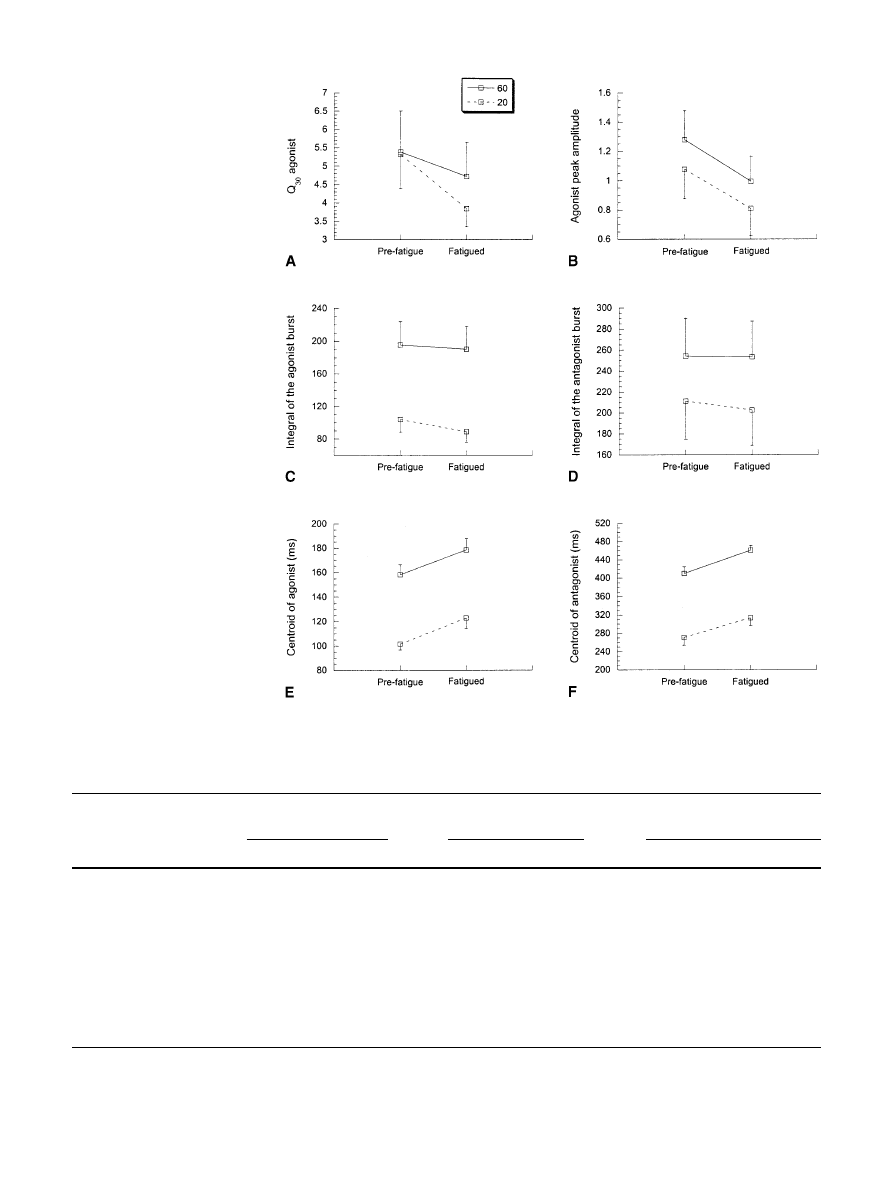
These findings are summarized in Fig. 3 and Fig. 4
and in Table 1 for all eight subjects. There was no
change in movement amplitude with respect to fatigue or
distance. Movement time significantly increased by
7.38% (averaged over 20° and 60°), and was longer for
longer movements. Movement time can be partitioned
into both acceleration time and deceleration time. Accel-
eration time increased significantly while deceleration
time was unchanged. There was a statistically significant
interaction between fatigue and distance for peak move-
ment velocity. As such, paired t-tests were performed on
both the 20° movements and the 60° movements. This
analysis showed that fatigue significantly decreased peak
velocity in the 60° movements by 7.2% [t(7)=–4.33,
4
P=0.003] while the decrease in peak velocity in the 20°
movements (4.89%) did not quite reach statistical signif-
icance [t(7)=–2.16, P=0.067]. Peak elbow torque
dropped significantly (by 15.22% averaged over 20° and
60°). Q
30
dropped by 24.95% (averaged over 20° and
60°) as a result of fatigue but this result was not statisti-
cally significant. Inspection of the data of individual sub-
jects showed that Q
30
dropped by as much as 50% in one
subject, and not at all in other subjects. The agonist peak
amplitude dropped significantly (by 30.53% averaged
over 20° and 60°). The integrals of the agonist burst
(6.83% decrease) and antagonist EMG burst (2.06% de-
crease), averaged over 20° and 60°, did not change sig-
nificantly. However, the timing of the centroid of the ag-
onist burst (14% change) and the antagonist burst (12%
change), averaged over 20° and 60°, occurred signifi-
cantly later in the fatigued condition.
Comparison between the intentionally slowed unfatigued
movements and fatigued movements
Before performing the fatigue protocol, subjects per-
formed 20 movements over 60° at a peak velocity that
was 10% less than their unfatigued maximal speed. From
this set of 20 movements, we later selected those that
were closest to the maximal speed of the fatigued move-
ments. The average number of trials that were selected as
the “intentionally slowed pre-fatigued movements” was
10 (range 6–17).
Intentionally slowed pre-fatigued movements were ki-
nematically indistinguishable from fatigued movements
but the patterns of muscle activation differed as shown in
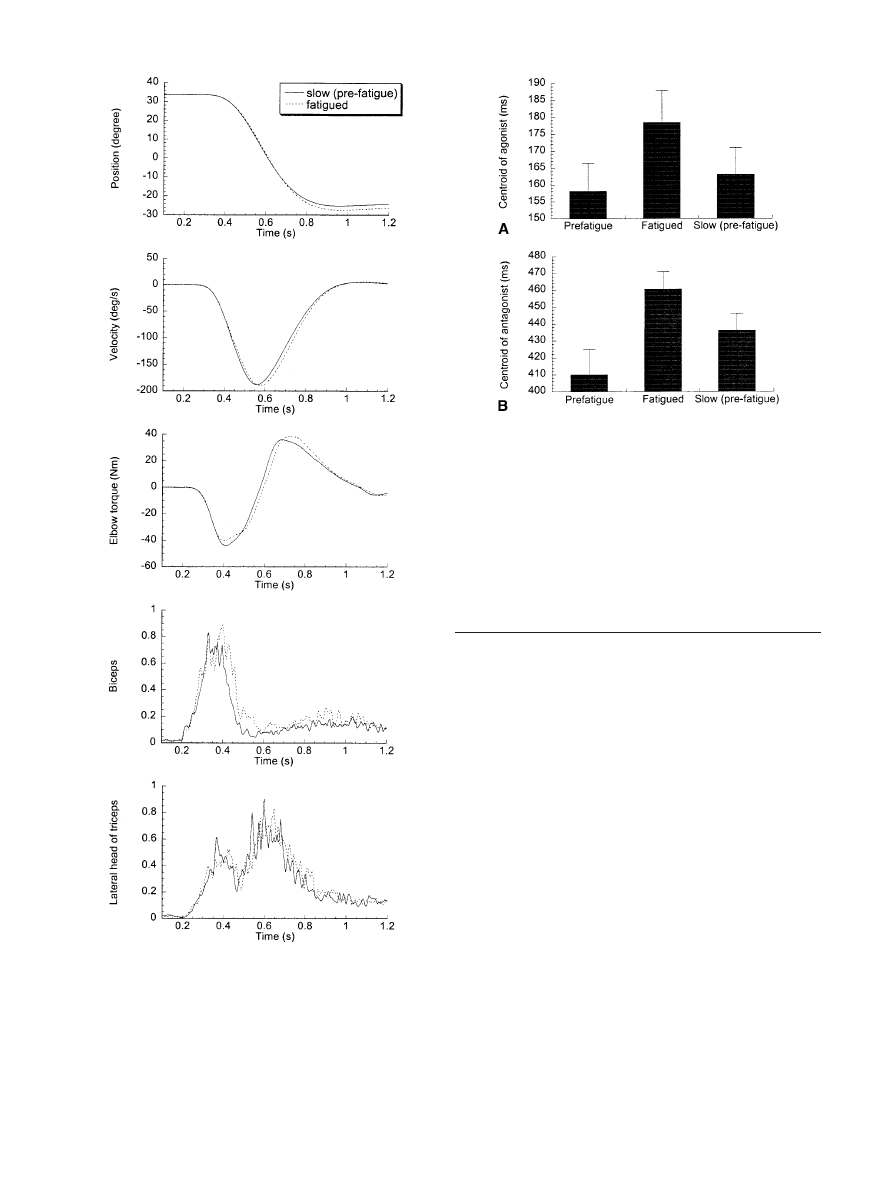
the time series plot for one subject in Fig. 5.
A paired t-test was used to determine if there were
significant differences in selected movement and EMG
parameters between fatigued and intentionally slowed
pre-fatigued movements. There were no significant dif-
ferences in movement amplitude, movement time, peak
velocity, Q
30
, the peak of the agonist burst or the integral
5
Fig. 1 Averaged maximum
voluntary isometric contrac-
tions in flexion (A) and exten-
sion (B) for a representative
subject. The data depict elbow
torque, biceps EMG, and later-
al head of triceps EMG. The
data are from subject 7
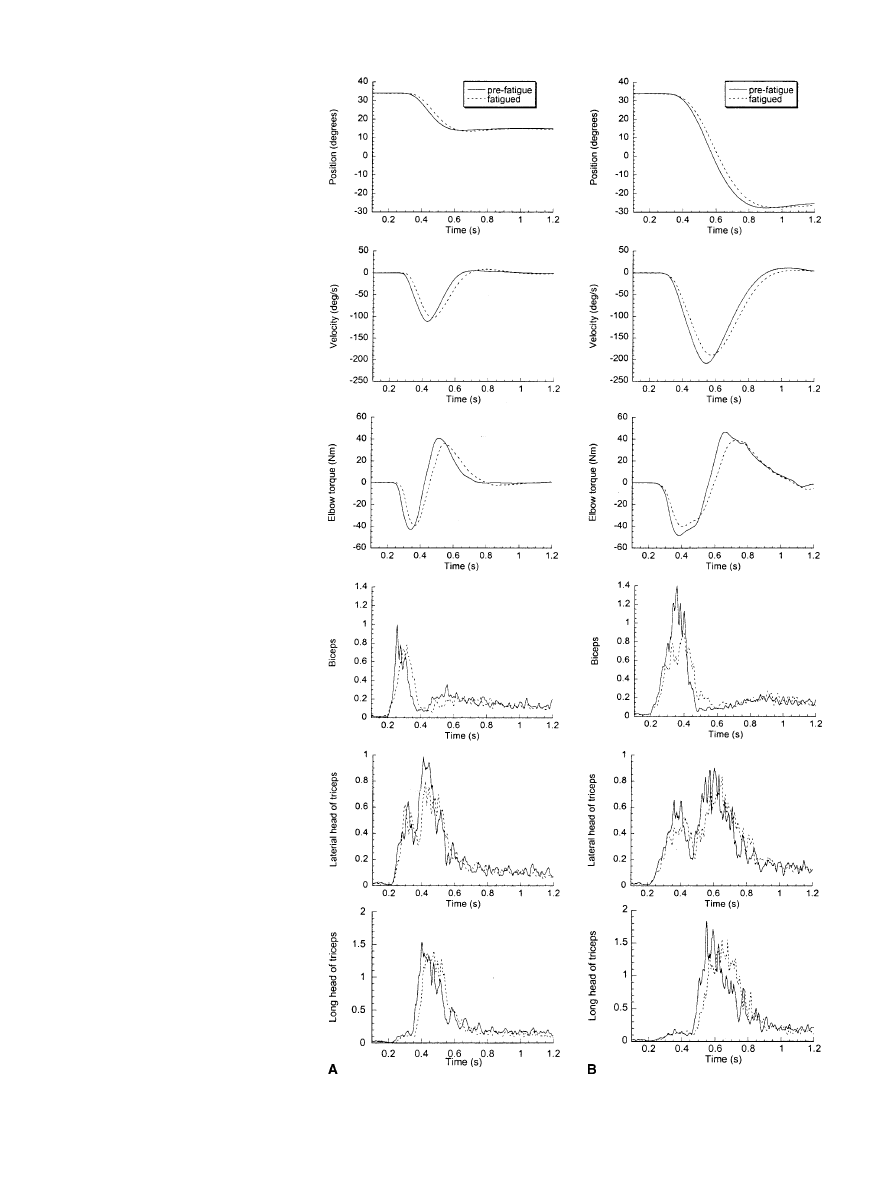
6
Fig. 2 Averaged position, ve-
locity, elbow torque, biceps
(agonist) EMG, lateral head of
triceps (antagonist), and long
head of triceps EMG for move-
ments over 20° (A) and 60°
(B). The data are averaged over
11 trials. Movements were per-
formed prior to the fatiguing
protocol (pre-fatigue) and fol-
lowing the fatigue protocol (fa-
tigued). The data are from sub-
ject 4
of the antagonist burst as shown in Table 2. However,
the integral of the agonist burst was significantly larger
in the fatigued movements. The centroid of the fatigued
agonist burst (C
ag
) was significantly later which indi-
cates an increase in EMG burst duration, and is consis-
tent with the fact that burst area increased although burst
peak amplitude did not. The centroid of the antagonist
burst (C
ant
) was significantly later in the fatigued move-
ments than the intentionally slowed pre-fatigued move-
ments. This is consistent with the fact that it was an in-
crease in acceleration time that produced an increase in
movement time in the fatigued movements. The data in
Fig. 6 depict C
ag
(part A) and C
ant
(part B) in the pre-
fatigue condition, the fatigued condition and in the inten-
tionally slowed pre-fatigued condition.
Effect of fatigue recovery interval
All the fatigue measures above were made after the sub-
jects had a 10 minute recovery period so that muscle
conduction velocities would return to normal (Kirsch
and Rymer 1987). To confirm the return of membrane
conduction velocity to pre-fatigue levels, a power spec-
trum analysis was performed on agonist EMG data from
the isotonic movements. There is a methodological issue
in performing a power spectrum analysis on EMG data
from isotonic movements. Normally, median frequency
is calculated on steady state data (e.g. Kirsch and Rymer
1987). The EMG bursts of isotonic movements are not
stationary and rarely exceed 300 ms in duration. We cal-
culated median frequency using 300 ms of data starting
from the marked agonist onset and padded with 700 ms
of zeros. This method examines frequency changes in
the agonist burst with a resolution of 1 Hz (DeLuca
1985). Consistent with the findings of Kirsch and Rymer
(1987), with 10 min of recovery, we found no statistical
difference in median frequency when comparing pre-fa-
tigued to fatigued movements [mean 20°: pre=72.4 and
post=73.1; mean 60°: pre=71.0 and post=73.1; F(1,7)=
0.22, P=0.65].
We also performed a study on four subjects with only
two minutes of recovery (Jiang 1996). All of the kinemat-
ic effects described above were larger in this group of
four subjects. Peak velocity of 60° movements dropped
by 25% after 2 min of recovery but only by 7.2% after
10 min as shown in Fig. 3D. For 20° movements, the
drop was 11.9% after 2 min of recovery and 4.89% after
10 min. These results show that 10 min of recovery al-
lows not only recovery in muscle fiber conduction veloci-
ty but also substantial recovery in kinematic performance.
Reduction of muscle fiber conduction velocity in-
creases the magnitude of the recorded EMG waveform.
7
Fig. 3 Movement time (A), acceleration time (B), deceleration
time (C), peak movement velocity (D) and peak elbow torque (E)
for 20° (dashed line) and 60° (solid line) movements are shown in
pre-fatigue and fatigued states. The data are averaged over eight
subjects. The data are mean±SE
8
Fig 4 The integral of the first
30 ms of the agonist EMG (A),
the agonist peak amplitude (B),
the integral of the agonist burst
(C), the integral of the antago-
nist burst (D), the centroid of
the agonist (E), and the antago-
nist (F) for 20° (dashed line)
and 60° (solid line) movements
pre-fatigue and fatigued. The
data are averaged over eight
subjects. The data are
mean±SE
Table 1 Effects of fatigue and distance. The results of two-way
factorial repeated measures ANOVA on eight subjects comparing
pre-fatigue movements and fatigued movements and movements
over two different distances. All degrees of freedom for the statis-
tical analysis are 1, 7
Fatigue
Distance
Interaction
Pre vs Post
20° vs 60°
Fatigue by Distance
F
P
F
P
F
P
Movement amplitude
0.38
0.555
7942
0.000
0.029
0.869
Movement time
35.14
0.001
187.1
0.000
0.46
0.520
Peak velocity
12.79
0.009
2002
0.000
26.68
0.001
Acceleration time
11.85
0.011
81.81
0.000
0.46
0.52
Deceleration time
0.02
0.882
90.99
0.000
0.26
0.63
Peak elbow torque
74.63
0.000
14.55
0.007
0.91
0.371
Q
30
3.33
0.111
0.46
0.519
2.95
0.130
Agonist peak
13.66
0.008
8.22
0.024
0.01
0.925
Q
ag
1.30
0.291
39.35
0.000
0.28
0.614
Q
ant
0.32
0.591
10.61
0.014
0.26
0.624
Centroid agonist
14.94
0.006
111.9
0.000
0.05
0.833
Centroid antagonist
46.84
0.000
183.1
0.000
0.35
0.572
This increase could be confused with an increase in re-
cruitment or firing frequency. Therefore, even though the
changes in EMG reported after 10 min of recovery were
also seen after 2 min, we have not presented those re-
sults, since their interpretation is open to question.
Discussion
Our first hypothesis was that the rules that the CNS uses
for compensating for a fatigue-weakened muscle are the
same as those for compensating for a larger inertial load.
Our reasoning is that it is the change in the ratio of re-
quired to available force that drives the compensatory
strategy. This implies that it does not matter whether the
ratio changes because the muscle gets weaker or the load
gets heavier. This strategy has three principal rules: pro-
longed agonist activation, delayed activation of the late
component of the antagonist burst, and no change in the
rate at which the agonist EMG burst rises. This hypothe-
sis was confirmed by the data. One additional finding is
that the peak agonist EMG is reduced which was not pre-
dicted. This change would tend to reduce muscle force
and therefore speed, which would be kinematically non-
compensatory. On the other hand, by reducing muscle
activation, this would tend to slow the progression of fa-
tigue, an effect that might be desirable but is incompati-
ble with kinematic compensation.
Our second hypothesis was that the EMG changes
would be greater for short movements than for long
movements but that the kinematic effects of fatigue
would be greater for long movements than for short
ones. We found that the degree of slowing was indeed
9
Fig. 5 Averaged position, velocity, elbow torque, biceps EMG
(agonist) and lateral head of triceps EMG (antagonist) for inten-
tionally slowed pre-fatigue movements and fatigued movements.
The data are from the same subject as in Fig. 2
Fig. 6 The centroid of the agonist (A) and the centroid of the an-
tagonist (B) in the pre-fatigue condition, the fatigued condition
and in the intentionally slowed pre-fatigue movement condition.
The data are mean±SE

greater for longer distance movements than for shorter
distance movements. However, the EMG did not show
greater changes for shorter movements than for longer
movements, which is in contrast to the findings of
Berardelli and colleagues (1984). This hypothesis, there-
fore, was not confirmed by the data.
We tested a third hypothesis that the EMG patterns
associated with fatigue-induced slowing would differ
from those of intentional slowing. This hypothesis was
confirmed by the data.
These findings question whether the reduction in
torque during movement is exclusively a consequence of
“peripheral, contractile fatigue”, i.e. a decrease in the ca-
pacity of the biceps to generate force. It might also re-
present a change in the way the CNS activates the mus-
cle that would serve to slow the progression of fatigue.
An additional interesting finding is the fact that
movement velocity was reduced by less than 10% after
ten minutes of recovery. This finding is consistent with
that of Raastad and Hallén (2000), who showed a
12–14% reduction in isokinetic performance following a
high-intensity exercise protocol, and a 6–7% reduction
after a moderate intensity protocol with a recovery time
of 5–20 min. The finding is also supported by the work
of Miller et al. (1987), who showed that a 4-min fatigu-
ing protocol can reduce MVC to less than 10% but that it
returns to almost 90% after 10 min of recovery. We also
used a large load to increase the effects of fatigue as
much as possible during the movement. Thus we believe
that we achieved a level of fatigue that was typical of
what has been done by many others. These results all
demonstrate that despite the fact we know how to fatigue
a muscle in order to produce an arbitrary decrement in
isometric force, it is exceedingly difficult to produce
substantial decrements in movement speed, while also
allowing sufficient time for conduction velocity to return
to normal.
Peripheral fatigue
We can draw conclusions similar to those of Kirsch and
Rymer (1987), and Griffin et al. (1998) about decreases
in dynamic torque/EMG ratios from our isometric and
movement data. When fatigued, our subjects showed an
increase in EMG during a 50% MVC isometric contrac-
tion. Fatigue also produced a decrease in peak elbow
torque during isotonic movements without a significant
change in the area of the agonist burst. Both findings re-
veal a decrease in the torque/EMG relationship. Such ob-
servations are also consistent with a number of other
studies (Edwards and Lippold 1956; Hagberg 1981;
Maton and Gamet 1989; Garland et al. 1994; Miller et al.
1996; Potvin 1997). These findings define the presence
of a peripheral fatigue component, a diminished ability
of muscle to produce force.
Central fatigue and rules for muscle activation
The presence of peripheral fatigue does not rule out cen-
tral fatigue as an additional factor. Central fatigue during
exercise has been defined by Gandevia et al. (1995a) as:
“The decrease in muscle force attributable to a decline in
motoneuronal output” (p. 281). Three of our measures of
the strength of activation, Q
30
, Q
ag
and Q
ant
were slightly
reduced by fatigue, but none of the changes reached sta-
tistical significance. The largest and most variable reduc-
tion was in Q
30
, which suggests that some subjects re-
duce the initial excitation of the muscle when fatigued
but others do not. The peak of the agonist burst was sig-
nificantly reduced by fatigue and this could have reduced
peak muscle force. Hence, using Gandevia's definition,
there is evidence for central fatigue in only one of our
four measures of motoneuronal output.
However, the timing of the EMG bursts in both ago-
nist and antagonist muscles was changed by fatigue to a
degree that could not be predicted by the way subjects
intentionally slow their movements. Fatigue prolonged
the agonist burst. If the level of muscle activation is un-
changed, this increases the force output of the muscle.
Were the CNS not to do this, the movement would be
even slower. Hence, this prolongation is compensatory
for the effects of peripheral fatigue and is consistent with
the changes seen when moving a heavier inertial load in
the unfatigued state. However, the compensation is not
complete and the fatigued movement is, never the less,
slower than the unfatigued movement.
Since the fatigued movement time is greater than that
of an unfatigued movement, delay of the antagonist is
10
Table 2 A comparison of fa-
tigued movements and inten-
tionally slowed pre-fatigue
movements. The results of
paired sample t-tests on the da-
ta of eight subjects when com-
paring the intentionally slowed
pre-fatigue movements with the
fatigued movements. The data
are averaged over eight sub-
jects (mean±SE)
Intentionally slowed
Fatigued
Significance
Mean±SE
Mean±SE
t
P
Movement amplitude
62.18±0.41
62.8±0.61
–1.15
0.287
Movement time
711±15
725±19
1.68
0.137
Peak velocity
183±4
185±4
0.93
0.386
Q
30
4.00±0.623
4.72±0.934
1.41
0.201
Peak of agonist burst
0.999±0.198
0.995±0.171
0.04
0.968
Integral of agonist burst
156.1±22.8
190.1±27.9
2.44
0.045
Integral of antagonist burst
245.1±39.3
253.6±33.8
0.71
0.499
Centroid of agonist burst
163±8
178±9
3.56
0.009
Centroid of antagonist burst
436±10
461±10
2.78
0.027

biomechanically appropriate. However, fatigue delayed
the antagonist burst to a degree that exceeded the amount
we would expect from the antagonist timing of unfa-
tigued movements of a similar speed. Furthermore, since
the fatigued movement's prolongation is due almost en-
tirely to prolongation of the acceleration time, and this is
not true of unfatigued movements with equal movement
times, it is also appropriate that the antagonist burst that
leads to decelerating torque be slightly more delayed.
This additional delay of the antagonist burst, like the
prolongation of the agonist burst, tends to increase
movement speed and may prevent stopping the move-
ment too soon. The limb's final resting position depends
on coactivation of the flexor and extensor muscles to
create a position of equilibrium. To perform an accurate
movement, the point at which movement speed reaches
zero during the deceleration phase should coincide with
this equilibrium position. This requires a delay in brak-
ing and hence a delay in antagonist onset.
How should we describe these changes? They do not
fit Gandevia's definition of central fatigue since there is
no overall decline in motoneuronal output. In fact, they
do not represent central fatigue if by that term, we wish
to imply something that diminishes motor performance.
We suggest that these changes represent a “central fa-
tigue strategy.” By this we mean that the CNS changes
the patterns of muscle excitation in order to reduce the
effects of peripheral fatigue (as in agonist prolongation)
and prevent moving incorrect distances due to peripheral
fatigue (as in antagonist delay). The reduction seen in
peak agonist EMG might also be considered part of a
central fatigue strategy. This reduction in peak agonist
EMG could be attributable to lower motoneuron firing
rates, so-called muscle wisdom (Marsden et al. 1983),
that are sufficient to fully activate a muscle in the fa-
tigued state secondary to the concurrent reduction seen
in muscle fiber relaxation rate when fatigued. This might
serve to prevent neuromuscular transmission failure.
It is also worth noting something that we did not find
to be part of a central fatigue strategy. We know that sub-
maximal isometric torque can be preserved in a fatigued
muscle by stronger activation of the muscle. Our second
hypothesis raises the question of whether subjects have a
reserve of performance that they can use, despite instruc-
tions to move as fast as possible. This reserve of perfor-
mance can usually be exploited only by extensive prac-
tice (Corcos et al. 1993). If true, then in the presence of
peripheral fatigue, subjects could, in theory, compensate
more fully by harnessing that reserve. To do this they
would increase the initial firing rates and number of re-
cruited motor units and this would be observed as an in-
crease in Q
30
. We had predicted that if this were the case,
compensation would be greater for shorter movements
than longer ones. This was based on the rationale that
shorter movements require lower forces and therefore
less muscle activation, leaving a reserve of motor units
that could be recruited. We did not find this. Hence, ei-
ther our subjects had a reserve that they were not suffi-
ciently motivated to use, or contrary to our supposition,
the reserve does not exist. These experiments do not al-
low us to decide this issue. That the reserve does not ex-
ist is supported by the fact that Q
30
was the same for
both short and long movements in the unfatigued state.
Additionally, it has been shown that subjects can pro-
duce maximum or near maximum voluntary activation of
their muscles under laboratory conditions (see Gandevia
et al. 1998), thus suggesting that under these conditions,
a reserve does not exist.
Finally, what is the relationship between the fatigue
we have studied and the fatigue that is experienced as
the consequence of sustained hard work or exercise? One
possibility is that exercise fatigue (as we might call it) is
simply greater and less well compensated. If so, were we
to repeat our fatigue protocol enough times, we would
get larger and more significant effects. The protocol we
used was difficult and unpleasant so such an experiment
would not be easy to perform. Another possibility is that
using a less intense and noxious protocol over a longer
period of time would produce more profound and less
compensated fatigue. This should be explored. A third
possibility is that the behavioral consequences of exer-
cise fatigue are different from the slowing of elbow flex-
ions that we are measuring here. This would suggest that
exercise fatigue is not simply a loss of muscle strength
due to muscular and neural factors, but a loss of coordi-
nation among muscles, a very different effect, and one
that is not expressed by the study of single-joint move-
ment. This too should be explored.
Acknowledgements This study was supported in part by the Na-
tional Institute of Arthritis and Musculoskeletal and Skin Diseases
Grant R01-AR 33189 and by the National Institute of Neurologi-
cal and Communicative Disorders and Stroke Grants K04-NS
01508, R01-NS 28127 and RO1-NS40902. We would also like to
acknowledge the valuable comments of Dr. Ziaul Hasan, and the
advice of Dr. Paolo Bonato and Dr. David Vaillancourt.
References
Berardelli A, Rothwell JC, Day BL, Kachi T, Marsden CD (1984)
Duration of the first agonist EMG burst in ballistic arm move-
ments. Brain Res 304:183–187
Bigland-Ritchie B, Johansson R, Lippold OCJ, Smith S, Woods JJ
(1983a) Changes in motoneurone firing rates during sustained
maximal voluntary contractions. J Physiol (Lond) 340:335–346
Bigland-Ritchie B, Johansson R, Lippold OCJ, Woods JJ (1983b)
Contractile speed and EMG changes during fatigue of sus-
tained maximal voluntary contractions. J Neurophysiol
50:313–324
Corcos DM, Gottlieb GL, Agarwal GC (1989) Organizing princi-
ples for single-joint movements: II. A speed-sensitive strategy.
J Neurophysiol 62:358–368
Corcos DM, Jaric S, Agarwal, GC, Gottlieb GL (1993) Principles
for learning single-joint movements I. Enhanced performance
by practice. Exp Brain Res 94:499–513
De Luca CJ (1985) Myoelectric manifestations of localized mus-
cular fatigue in humans. CRC Crit Rev Biomed Eng 11:251–
279
Edwards RG, Lippold OC (1956) The relation between force and
integrated electrical activity in fatigued muscle. J Physiol
132:677–681
Enoka RM, Stuart DG (1992) Neurobiology of muscle fatigue. J
Appl Physiol 72:1631–1648
11

Gandevia SC, Allen GM, McKenzie DK (1995a) Central fatigue:
critical issues, quantification and practical implications. In:
Gandevia SC, Enoka RM, McComas AJ, Stuart DG, Thomas
CK (eds) Fatigue: neural and muscular mechanisms. Plenum
Press, New York, pp 281–294
Gandevia SC, Enoka RM, McComas AJ, Stuart DG, Thomas CK
(eds) (1995b) Fatigue: neural and muscular mechanisms. Ple-
num Press, New York
Gandevia SC, Herbert RD, Leeper JB (1998) Voluntary activation
of human elbow flexor muscles during maximal concentric
contractions. J Physiol: 512.2:595–602
Garland SJ, Enoka RM, Serrano LP, Robinson GA (1994) Behav-
ior of motor units in human biceps brachii during a submaxi-
mal fatiguing contraction. J App Physiol 76:2411–2419
Gottlieb GL, Corcos DM, Agarwal GC (1989) Organizing princi-
ples for single-joint movements: I. A speed-insensitive strate-
gy. J Neurophysiol 62:342–357
Griffin L, Garland SJ, Ivanova T (1998) Discharge patterns in hu-
man motor units during fatiguing arm movements. J App
Physiol 85:1684–92
Hagberg, M. (1981) Muscular endurance and surface electromyo-
gram in isometric and dynamic exercise. J App Physiol: Resp
Env Exer Physiol 51:1–7
Jiang, H. Gottlieb, GL, Corcos, DM (1996) Effects of fatigue on
voluntary elbow flexion movements. Soc Neurosci Abstr
22:129
Kirsch RF, Rymer WZ (1987) Neural compensation for muscular
fatigue: evidence for significant force regulation in man. J
Neurophysiol 57:1893–1910
Lucidi CA, Lehman SL (1992) Adaptation to fatigue of long dura-
tion in human wrist movements. J App Physiol 73:2596–2603
Marsden CD, Meadows JC, Merton PA (1983) “Muscular wis-
dom” that minimizes fatigue during prolonged effort in man:
peak rates of motoneuron discharge and slowing of discharge
during fatigue. In: Desmedt JE (ed) Motor control mechanisms
in health and disease. Raven Press, New York, pp 169–211
Maton B, Gamet D (1989) The fatigability of two agonistic mus-
cles in human isometric voluntary submaximal contraction: an
EMG study. II. Motor unit firing and recruitment. Eur J App
Physiol 58:369–374
Miller KJ, Garland SJ, Ivanova T, Ohtsuki T (1996) Motor-unit
behavior in humans during fatiguing arm movements. J Neu-
rophysiol 75:1629–1636
Miller RG, Giannini D, Milner-Brown HS, Layzer RB, Koretzky
AP, Hooper D, Weiner, MW (1987) Effects of fatiguing exer-
cise on high-energy phosphates, force and EMG: evidence for
three phases of recovery. Muscle Nerve 10:810–821
Mustard BE, Lee RG (1987) Relationship between EMG patterns
and kinematic properties for flexion movements at the human
wrist. Exp Brain Res 66:247–256
Pfann KD, Hoffman DS, Gottlieb GL, Strick PL, Corcos DM
(1998) Common principles underlying the control of rapid,
single degree-of-freedom movements at different joints. Exp
Brain Res 118:35–51
Potvin JR (1997) Effects of muscle kinematics on surface EMG
amplitude and frequency during fatiguing dynamic contrac-
tions. J App Physiol 82:144–151
Raastad T, Hallén J (2000) Recovery of skeletal muscle contractil-
ity after high- and moderate-intensity strength exercise. Eur J
Appl Physiol 82:206–214
Scalzitti DA (1994) Effects of age and gender on quadriceps fem-
oris muscle force production and anaerobic metabolism. Mas-
ter thesis, University of Illinois at Chicago
Tschoepe BA, Sherwood DE, Wallace SA (1994) Localized mus-
cular fatigue duration, EMG parameters and accuracy of rapid
limb movements. J Electromyogr Kinesiol 4:218–229
12
Wyszukiwarka
Podobne podstrony:
Developing Your Intuition With Distant Reiki And Muscle Test
Bigorexia Bodybuilding and Muscle Dysmporphia
Abductor Pollicis Longus Muscle and Extensor Pollicis Brevis Muscle Support KT method
Cthulhu Dark Ages Fatigue, Exposure, Fire and More
Middle Trapezius Muscle and Infraspinatus Muscle KT method
Middle Trapezius Muscle and Infraspinatus Muscle tapeSP
Notch and Mean Stress Effect in Fatigue as Phenomena of Elasto Plastic Inherent Multiaxiality
7 Things You Must Do to Add Muscle, Gain Weight, and Increase Strength
Proteus 8 5 installation and activation
Hedonic Tone and Activation Level
Cistus incanus and Cistus monspeliensis inhibit the contractile response in isolated rat smooth musc
effects of kinesio taping on the timing and ratio of vastus medialis obliquus and lateralis muscle f
Vista, Windows 7, and Server 2008 Activation Script
Transient TLR Activation Restores Inflammatory Response and Ability To Control Pulmonary Bacterial I
Step by step instructions activation of all brands of machines for EasyDiag and completion of the sc
Design Fatigue Test and NDE of a Sectional Wind Turbine Rotor Blade
więcej podobnych podstron