W jaki sposób można wykorzystać w hodowli niepłodne mieszańce międzygatunkowe lub międzyrodzajowe? Znaczenie poliploidyzacji spontanicznej i sztucznej - przy pomocy kolchicyny.
posiadanie lub przejmowanie od form dzikich poszczególnych cech lub kompleksowych właściwości= introgresja
często sa formami allopoliploidalnymi.
wyjasniają stosunki pokrewienstwa
wywołuja resynteze gatunków- odtworzenie ich pochodzenia
umozliwiają syntezę nowych gatunków
KOLCHICYNA- działanie polega na zablokowaniu mechanizmy wrzeciona podziałowego. Po podłużnym podziale wrzeciona nie rozchodzą się na bieguny i nie tworzą się komórki potomne. Jeżeli stężenie kolchicyny jest za wysokie lub jej działanie trwa zbyt długo,nastepuja dalsze podziały chromosomów prowadzące do śmierci komórki na skutek przekroczenia poziomu ploidalności.
Znaczenie ujemne:
na skutek zwiekszenia liczby chromosomów jądra komórkowego jak i całe komórki powiększają się przez co zmienia się ilościowy stosunek plazmojądrowy, objetość komórki zwieksza się znaczniej niż jej powierzchnia.
Przemiana materii , podziały komórkowe zachodzą wolniej na skutek czego również wzrost i rozwój jest wolniejszy w porównaniu z formami diploidalnymi.
Czasami obniżenie odporności
Często liczba kwiatów w kwiatostanach jest mniejsza
Zaburzenia plonu nasion, który może być obniżony z powodu zaburzeń w gametogenezie i w procesie zapłodnienia.
Ogólna produkcja swieżej masy może być większa, lecz w przeliczeniu na jednostke czasu jest z reguły mniejsza niż u diploidów.
Wyższa zawartośc wody w świeżej masie.
Znaczenie pozytywne:
< wszystkie części rosliny sa większe
opóźnienie drewnienia tkanek
bujniejsze rosliny- wieksz produkcja swieżej masy
owoce i nasiona sa większe
bardziej sztywne pedy
Jakie korzyści z punktu widzenia hodowlanego wnoszą mieszańce somatyczne? Jakie znaczenie mogą mieć mieszańce asymetryczne?
Mieszańce somatyczne mogą być: symetryczne, asymetryczne cybrydy
Korzyści:
przeniesienie odpornosci na patogeny, np. do ziemniaka form dzikich na bakterie ervinia carotovora, nicienie
zwiększenie wartości odzywczej i trwałosci roslin pastewnych poprzez otrzymanie nowego gat, trawy
otzrymanie cytoplazmy roslin męskosterylnych ziemniaka, kapusty
znaczenie:
wieksza zmiennosć genetyczna
otrzymujemy różne proporcje genów
miszance mogą posłuzyc do krzyżowań.
Wykorzystanie kultur tkankowych do otrzymywania materiałów hodowlanych.
utrzymanie i rozmnożenie danego materiału
mozliwość otrzmania mutantów
mozliwość dokonania selekcji invitro
źródło zmienności somaklonalnej
możliwość prowadzenia kultur pylników
przy barierach pre i post zygotycznych wykorzystujemy do otrzymania mieszanców międzygatunkowych
prowadzenie somatycznej embriogenezy- otzrymywanie sztucznych nasion
możemy dokonac transformacji komórki
łatwiejsze wykonanie mutacji niż w warunkach naturalnych
łatwośc wykonania rozmnożenia klonalnego, co gwarantuje uzyskanie wiekszej liczby osobników z nowa własciwościa.Jakie znaczenie może mieć zastosowanie sztucznych nasion? Omów procedurę ich otrzymywania.
Sztuczne nasiona: somatyczne zarodki uzyskane w hodowli in vitro, otoczone sztuczną okrywą zawierająca materiały odżywcze oraz regulatory wzrostu. Są 2 technologie wytwarzania sztucznych nasion:
mokra- zarodki są kapsułkowane w hydrożelach
sucha- zarodki po osiągnięciu stadium dojrzałości poddawane są suszeniu.
Proces uzyskiwania ma 4 fazy:
hodowla tkanki proembrionalnej w kulturach in vitro
masowa produkcja synchronicznie rozwijających się zarodków somatycznych
przygotowanie uzyskanych zarodków do kiełkowania przez ochładzanie lub podsuszanie
otoczkowanie w hydrożelach lub kapsułowanie otrzymanych zarodków.
Zastosowanie: ogrodnictwo, lesnictwo, sadownictwo, sztuczne nasiona mogą być jedynym sposobem rozmnażania roslin poddanych różnym manipulacjom genetycznym, sztuczne nasiona: modrzew europejski, świerk biały, pospolity, orzech woski
Jakie związki mogą występować między hodowlą roślin a biotechnologią?
Hodowcy często uciekaja się do metod biotechnologii aby udoskonalić rosliny. Udoskonalenie to polega na wszczepieniu do genomu rosliny obcych genów, w wyniku czego powstająrosliny transgeniczne. Zaczęły się pojawiac w latach 80'. Izolowany przenoszony gen może pochodzic od zwierzat, mikroorganzmów lub innych roslin. Aby przeniesc gen musimy: wyizolowac go, znac jego ekspresje, tak go wzbogacic w inne geny, aby mógł funkcjonowac w organixmie do którego go przenosimy.
Wektory i przenosniki genu: najcz ęściej jest to plazmid Ti z bakterii agrobacterium tumefaciens powodującej przy porazeniu roslin powstawanie tumorów na korzeniach. Plazmid jest to kolista cząsteczka DNA wystepująca poza głównym bakteryjnym DNA. W plazmidzie sa geny kodujące enzymy potzrebne do wnikniecia do rosliny i odpowiedzialne za powstawanie tumoru. Biotechnolodzy wycięli geny tumoru i wprowadzili geny, o które chca wzbogacić rosline, geny markerowe, oraz geny ułatwiające transformacje.
Plazmid Ri- z jednej z bakterii agrobacterium powodującej powstawanie wielu korzeni przybyszowych
Metoda bidistyczna- małe czasteczki metalu (złoto, tytan) opłaszczone DNA i wstzreliwane do genomu rosluiny. Metoda ta jest stosowana, gdynie możemy wykorzystac bakterii.
Możemy wprowadzic do rosliny 1-3 gemy, więc nie możemy wprowadzac poligenów. Przenosimy geny odpowiadające za: odpornosć na herbicydy, wirusy, szkodniki, skład, czas dojrzewania.
Proces wprowadzania genu: pobieramy tkanke roslinna , najlepeij z młodej rosliny , eksplantaty przenosimy na szalke , na sztuczna pozywke, stwarzamy dobre warunki infekcji, mieszamy eksplantaty z zawiwesiną bakterii, bakteria swobodnie penetruje do tkanki, po 10 min wyjmujemy eksplantaty, osaczamy i przenosimy na pozywke z antybiotykiem niszczącym bakterie, która wprowadziła nam już DNA. Ze stransformowanych komórek regenerujemy całe rosliny, jeżeli zastosowaliśmy dobry plazmid i dobry antybiotyk to jest duże prawdopodobienstwo, że otzrymana roslina będzie transgeniczna. Inna metoda jest kaleczenie dorosłej rosliny, infekcja bakterią, a nastepnie izolacja komórek z tumoru i regeneracja z nich całej rośliny, jednakze jest to metoda mniej wydajna i rzadziej stosowana. Możemy również pod ciśnieniem transformować pakikwiatowe w celu transformacji komrki jajowej, pyłku, powstałe nasiona wysiewamy na pożywkę z antybiotykiem i przeprowadzamy selekcje. W plazmidzie Ti oprócz genów replikacji, sa inne geny, np. opi- wksaźnik czy gen został już wprowadzony do komórki. Cechy które wprowadzono (patrz poprzednie pyrania). Rośliny transgeniczne nie oddziaływują źle na rosliny, środowisko czy człowieka.
Biotechnologia umozliwia również produkcje sztucznych nasion, jednak dotychczas stworzone sztuczne nasiona nadaja się tylko do doświadczeń laboratoryjnych, nie udało się stworzyc takich, które moznaby wykorzystac w uprawie.
Kultura in vitro jest to kultura eksplantatów , polega na wyizolowaniu eksplantatów i umieszczeniu ich w sterylnym środowisku- pozywce. Pozwala na to totipotencja komórkowa, czylizdolność pojedynczych komórek roślinnych do odróznicowania i regeneracji całego organizmu roslinnego. Zastosowanie:
utrzymanie bądx rozmnożenie cennego materiału
otzrymanie mutantów
selekcja in vitro
wykorzystanie smienności somaklonalnej
otrzymanie haploidów dzieki kulturze mikrospor, pylików, zalążni, zalążków
otrzymanie mieszanców międzygatunkowych umozliwia kultura zarodków.
Zapylenie in vitro (jak jest niezgodnośc prezygotyczna)
Otrzymywanie mieszańców somatycznych powstałych z poacenia 2 komórek somatycznych.
Otrzynmanie zarodków somatycznych przez wykorzystanie embriogenezy somatycznej.
Omów zasady sporządzania map chromosomowych. Czy mapy takie mogą być przydatne w hodowli?
Tomasz Morgan zaobserwował, ze częscstość z jaką pojawiają się rekombinanty, jest stała dla danej pary genów, co wskazywało, że geny zajmują na chromosomie stałe miejsca(loci). Pojawienie się rekombinantów jest wynikeim pekania chromatyd chromosomów homologicznych i ich ponownego łaczenia się w nowym układzie. Proce ten to crossing-over. Morgan załozył również, że częstoś crossing-over jest funkcją odległości pomiędzy loci genów na chromosomie: im dalej od siebie sa połozone, tym wieksza szansa na zajście crossing-over miedzy nimi. Słuszność założeńMorgana została potwierdzona. Dla wszystkich organizmów będacych przedmiotem intensywnych prac genetycznych, oraz dla wielu organizmów ważnych z punktu widzenia użytkowego, sporzadzono dokładne mapyy gebetyczne poszczególnych chromosomów.. Mapy te obrazuja kolejnośc ułozenia genów na chromosomieoraz odległości między genami wyrazone w jednostkach rekombinacji. W celu sporzadzenia takiej mapy wykonuje się serię krzyzówek testowych, zwykle krzyżówek trzy lub więcej punktowych, tzn. takich w których bada się jednoczesnie segrgacje trzech lub więcej genów. W wyniku krzyżowki powstaje 8 klas potomstwa, wszystkie rozrózniane na podstawie fenotypów. O ile wszystkie trzy geny sa sprzężone, to klasy rekombinantów będą mniej liczne nizklasy typu rodzicielskiego. Najmniej liczne będą klasy postałe w wyniku podwójnego c/o, gdyz prawdopodobienstwo zajscia podójnego c/o = iloczynowi prawdopodobieństw zajscia pojedyńczych c/o. Ponieważ w wyniku podwójnego c/o tylko gen środkowy zmienia swoja pozycje względem pozostałych genów, to liczebność klas daje nam informację nie tylko o odległościach pomiędzy genami, ale również o ich wzajemnym położeniu na chromosomie. ! jednostka rekombinacji (centy morgac cM) = 1% rekombnacji.
Mapy chromosomowe sa przydatne w hodowli ponieważ uzyskujemy z nich informacje o sile sprzężeń między genami i prawdopodobieństwie uzyskania konkretnych rekombinantów. Podcas zakładania hodowli mamy co do niej ustalone plany i wiemy co chcemy uzyskać. Korzystając z map chromosomowych różnych gatunków danej rolsinymożemy wybrać te gatunki, które przekazuja jak największej liczbie potomstwa korzystne dla nas cechy, a poza tym geny warunkujące te cechy nie sa sprzężone z genami dla nas niekorzystnymi czy tez sa sprzęzone, ale w mniejszym stopniu. Możemy tez przewidzieć jaki procent potomstwa będzie
Wykład 5. Systemy krzyżowania i selekcji. Hodowla kombinacyjna i transgresyjna.
Typy selekcji i
Czym różni się „selekcja" od „metody hodowlanej"?
Selekcja jest najstarsza i najprostsza metoda hodowlaną, oparta na wyborze z naturalenj populacji poządanych biotypów. W metodzie tej rola hodowcy ograniczona jest do wykorzystania już isniejącej zmienności. Jednakże selekcja, jako metoda hodowlana, nie powinna być utożsamiana z samym tylko wyborem korzystnych wariantów. Wybór taki stanowi z reguły nieodzowny element techniczny każdej metody hodowli niezaleznie od tego czy zmiennosć materiału wyjściowego została w ten czy inny sposób poszerzona czy nie.
Selekcje dzielimy na selekcję roslin samopylnych i obcopylnych. Łatwiejsza i szybsza jest selekcja roślin samopylnych , jest tez bardzo skuteczna, gdyż rosliny zapylaja się swoim własnym pyłkiem, co prowadzi do powstania linii czystych, które już pozostaja niezmienne więc szybko możemy uzyskać osobniki pożądane. Selekcja roślin obcopylnych jest trudniejsza ze względu na swobodne przepylanie się roslin, wobec czego aby otrzymac pożądane formy należy kontrolowac zapylanie, znac formy ojcowskie, izolowac rosliny, zabiegi z tym związane sa pracochłonne i drogie.
Selekcje możemy wykonywać: a) przed kwitnieniemwówczas rosliny niepotrzebne odrzucamy przed kwitnieniem i krzyżuja nam się już wyselekcjonowane lepsze formy b) po kwitbieniu: gdy interesuja nas takie cechy jak ziarniaki u zbóz, owoce. Niestety gdy dotyczy to roslin obcopylnych przekzyżują się one swobodnie co oznacza, że mogą się przkżyżowac z formami lepszymi i gorszymi(przyudatna jest tu metoda rezerw)
Selekcja może być:
pozytywna- wybór i rozmnozenie najlepszych osobników
negatywna- odrzucenie najgorszych form
indywidualna z ocena potomstwa
masowa (populacyjna)
masowo-grupowa- reprodukcja pojedynków w grupach
rozdzielcza- prowadzona w 2 kierunkach, otrzymujemy małe populacje o cechach skrajnie różniących.
Zachowawcza- selekcjonowanie osobników najbardziej trypowych dla odmiany.
W zaleznosci od biologicznych własciwosci danego gatunku stosuje się selekcję klonów,, selekcję masowa lub indywidualna. Selekcja klonów możliwa jest tylko w stysunku do roślin, które można rozmnazać wegetatywnie.w stosunku do roslin rozmnażanychgeneratywnie można stosowac zarówno selekcje masowa jak i indywidualna ale sposób postępowania jest odmienny dla gatunków samo i obcopłodnych..
Selekcja klonów: wybieramy rosliny rozmnażajace się wegetatywnie, ale mające odmienne genotypy. Najcęsciej materiał wyjściowy stanowi generatywne pokolenie F1, uzyskane z nasion powstałych przez samozapylenie lub krzyzowanie. Pierwszy wybór pojedynków jest bardzo istotny ponieważ w wyniku wegetatywnego rozmnażania ich własciwości genetyczne zostaja utrwalone w całym klonie. Rosliny po pierwszej selekcji to klony A, sadzimy je na polu i porównujemy ze standardem, badamy ich cechy jakościowe, ilościowe, termin dojrzewania itp. Wybieramy najlepsze i rozmnazamy (klony B)powtarzamy to osiągając klony c i d na których wykonujemy doswiadczenia polowe, porówawcze i najlepsze klony rozmnażamy. Można stosowac dla roslin rozmnazających się generatywnie jak i wegetatywnie. Wybór klonów i krzyzowanie najlepszych z nich pozwoli otrzymać najlepze formy.
Selekcja masowa: a) rosliny samopłodne - polega na wyborze z naturalnej populacji większej liczby roslin o pożądanym fennotypie i ich wspólnym dalszym rozmnażaniu. Oceny potomstwa nie stosuje się. Zakłada się, że rosliny o poządanym fenotypie jednorodnym, sa też genotypowo jednorodne. Selekcja masowa znajduje zastosowanie w ulepszaniu prymitywnych odmian. Modyfikacje selekcji masowej stanowi selekcja grupowa polegająca natym, że wyselekcjonowane rosliny dzieli się na grupy o okreslonych cechach i rozmnaża je oddzielnie. W ten sposób możemy otrzymac mniejsze grupy roślin o określonych cechach, np. formy jare i ozime. Bywa stosowana jedynie w hodowli zahowawczej.
b)rosliny obcopłodne- rosliny te sa w wiekszym lub mniejszym stopniu heterozygotami i selekcja masowa polega tu na zwiększeniu udziału pożądanych genów w populacji. Efekty sa szczególnie wyraxne w poczatkowym okresie, potem maleja i w końcu sa niezauważalne. Selekcja jest kontynuowana w ciągu kilku pokoleń.
Skuteczność tej metody zalezy od:
odziedziczalności cechy stanowiącej przedmiot selekcji, im mniejszej podlega zmiennosci środowiskowej tym lepiej i tym skuteczniejsza bezie selekcja.
Czy selekcjonowana cecha objawia się przed czy pokwitnieniu
Genetyczny mechanizm przekazywania cechy (recesywna, dominujaca)
Jednoczesnie obok selekcji hodowcy przebiega selekcja naturalna, która może być zgodna z kierunkiem selekcji prowadzonej przez hodowce lub nie.
selekcja indywidualna z ocena potomstwa:
rosliny samopłodne- potomstwo poszczególnych wybranych pojedynków jest rozmnażane oddzielnie i testowane.. populacja składa się z homozygotycznych ro,slin o niejednakowych genotypach; w wyniku selekcji indywidualnej wyprowadza się z niej linie czyste, które mogs róznic się od populacji wyjściowej; dalsza selekcja w obrębie lini czystej jest bezskuteczna.
Stosowana w celu uzyskania komponentów do hodowli kombinacyjnych oraz jako selekcja w obrębie odmian miejscowych dobrze przystosowanych do lokalnych warunków.
W hodowli gatunków u których bonitacja artości uzytkowej może być dokonana dopiero po przekwitnieniu, dobre wyniki daje tzw. Metoda rezerw. Polega ona na tym, że z nasion wybranych rodów czy rodzin tylko część wysiewa się w doświadczeniach porównawczych, podczas gdy reszta stanowi rezerwę. Po dokonaniu wyboru najlepszych rodzin czy rodów na podstawie tych doświadczen hodowcy pozostaja jeszcze do dyspozycji ich nasiona. W nastepnym roku zatem zostają zatem rozmnożone i przekzyzowane te rody, które uzyskały wysoka wartość użytkowa. Najczęsciej powtarza się celem uzyskania peniejszych wyników po czym nasiona najlepszych rodów B sa miesznne i po kilkakrotnym rozmnożeniu mmogąstanowic kwalifikowany materiał siewny. Pozwala na dużą kontrole zapylenia i eliminacje niepożadanych rodów, nawet gdy cecha ujawnia się po kwitnieniu. Można również krzyżowac roslinyy parami, wówczas zobaczymy, które dadzą najlepsze potomstwo.
Jakie znasz metody hodowli kombinacyjnej? (rodowodowa, populacyjna-ramsz, populacji cząstkowych, pojedynczych nasion
Hodowla kombinacyjna jest to metoda hodowli krzyżówkowej roslin samopłodnych. Opiera się na podstawowych regułach dziedziczenia. Począwszy od pokolenia F2 następuje rozszczepienie się cech i pojawienie się ich w nowych kombinacjach. Tym samym już w pokoleniu F2 można zacząć poszukiwania pożadanych rekombinantów. U roslin samopłodnych z pokolenia na pokolenie udział heterozygot zmniejsza się na korzysć homozygot. Z tego względu w hodowli kombinacyjnej można wyodrębnić dwie podstawowe metody, które mogą być ze sobą w różny sposb łączone.
metoda rodowodowa: selekcja rozpoczyna się tu w pokoleniu F2, z populacjiniejednorodnej na skutek rozszcepien wybiera się fenotypowo najlepsze pojedynki i nasionami kazdego z nich obiewa się indywidualnie po 1 poletku. W F3 wybiera się najlepsze potomstwa , a znich najlepsze rośliny, charakteryzujące się pożadanymi kombinacjami cech. W następnych pokoleniach kontynuuje się wybór pojedynków az ich potomstwo w wyniku wzrostu homozygotyczności nie stanie się na tyle jednorodne, aby je traktowac jako wyrównane rody hodowlane. W pokoleniu F7 stopien homozygotyzacji jest już z reguły na tyle wysoki, że rody a można rozmnażać i porównywać w doświadzceniach z powszechnie uprawianymi odmianami wzorcowymi. Dalszy wybór odbywa się wśród rodów B, C, D podobnie jak w selekcji.
Metoda populacyjna czyli ramsz. W odróżnieniu od rodowodowej wybór pojedynków rozpoczyna się tu w późniejszych generacjach. U roslin samopylnych wzrost idziały homozygot jest bardzo szybki w kolejnych pokoleniach w związku z czym przystępując do selekcji indywidualnej w F5-F7 można spodziewac się, ze wybrane z populacji pojedynki, będą w znacznym stopniu homozygotyczne. Tym samym rody A uzyskane z rozmnażania tych pojedynków są bardzo jednorodne pod względem licznych własciwości. Należy zadbac o to aby uzyskac odpowiednią liczbę roślin, wobec tego krzyżujemy tyle razy pary wytypowane na rodzicielskie aby w pokoleniu F2 uzyskać 10 000- 20000 roslin. W pokoleniach F2-F5 stosujemy ramsz , czyli pozwalamy na swobodne przepylanie się roslin. Wzrasta poziom homozygotyczności i zachodzi selekcja naturalna. W pokoleniu F6 wybieramy najlepsze pojedynki, których potomstwo stanowi rody A, następnie selekcjonuje się je i otrzymujemy rody B, na których przeprowadza się mikrodosiadczenia. Na tej podstawie selekcjonuje się rody C, a potem D, są one badane w dużych wielopowtórzeniowych badaniach zaozonych w kilku miejscowościahc. W końcu wybieramy kilka czolowych linii. Dzieki działaniu selekcji naturalnej w F6 pozostaja tylko rośliny dobrze przystosowane do miejscowych warunków. Jednakze należy bacznie obserwowac zachodzącą selekcję, bo może się ona odbywac w wyniku konkurencji pomiędzy poszcególnymi rośłinami. Również dobór naturalny może iść w niechcianym kierunku.
Metoda populacji cząstkowych: stanowi kombinacje wielu metod. Selekcja rozpoczyna się już w pokoleniu F2, podobnie jak w metodzie rodowodowej.wybiera się 3000-5000roslin najbardziej odpowiadających zamierzeniom hodowcy i rozmnaża się je indywidualnie. poPonieważosliny te sa przewaznie heterozygotyczne, ich potomstwa traktuje się jak populacje cząstkowe (ramsze), które rozmnaża się nastepnie w ciągu kilku pokoleń, az do uzyskania wysokiego stopnia homozygotyczności, likwidując ramsze wykazujące niepożądane właściwości. Z kolei w obrębie pozostawionych ramszów przeprowadza się wybóe pojedyńczych roslin i przebieg dalszej hodowli jest taki sam jak w metodzie rodowodowej.
Jakie procesy zachodzące w czasie mejozy są podstawą hodowli kombinacyjnej i transgresyjnej? Czy sprzężenia genów pomagają czy przeszkadzają w hodowli?
Moim zdaniem podstawa jest segregacja chromosomów w Iipodzale mejozy oraz crossing-over zachodzące w I podziale podczas profazy. Dzieki tym procesom powstja 4 różne gamety, zawierające rózne allele tych samych genów. Oczywiście należy pamiętac,że każdy gen może mieć tylko 2 rózne allele, więc mogą powstać 2 różne pod względem danego genu gamety. Ale dochodzi nam tutaj proces crossing-over, który w kazdej dzielacej się komórce i w każdej parze hromosomów homologicznych może zajsć w innym miejscu co daje nam ogromna ilość możliwości jakości zawartosci materiału genetycznego w gametach. W tej ogromnej puli gamet sa te zawierające korzytne dla nasw hodowli allele, które po połaczeniu z korzystnymi gametami drugiego rodzica stworzą rośline o interesujacych nas cechach, rosliny takie wyselekcjonujemy do dalszej hodowli.
Wydaje mi się, że sprzężenia genów sa niekorzystnym dla hodowli zjawiskiem przede wszystkim dla hodowli prowadzonej w celu przeniesienia z jednej rosliny do drugiej tylko jednego genu. Im bliżej geny leżą obok siebie, tym sa bardziej sprzezone i tym rzadziej wystepuje miedzy nimi c/o. Gdy obok genu warunkującego ujawnienie się pozytywnej pod względem hodowlanym cechy połozony jest gen niekorzystny, wiele trzeba wykonac powtórzen krzyzowań, aby usunąć niekorzystny gen (allel) i zastąpić go pozytywnym. Jednakze może być również sytuacja gdzie obok siebie leża same korzystne geny i wówczas sprzężenie nam nie przeszkadza.
Jaki związek ma współdziałanie genów z hodowlą? Jakie znasz typy współdziałania genów?
Współdziałanie genów jest przyczyna typowych odchyleń od mendlowskich rozszczepien.
geny letalne(wywołują bezpośrednio smierć organizmu) i subletalne (przyczyniaja się do osłabienia organizmu i jegowcze,sniejszej śmierci). U myszy biała barwa jest letalna. Gen letalny może być dominujący lub recesywny, w heterozygotach gen letalny jest maskowny, tylko organizmy heterozygotyczne niosa genny letalne, u samopylnych , które są homozygotami nastepuje samoczynne oczyszczenie z tych genów.
Plejotropia: taki sposób dziedziczenia, w którym jeden gen determinuje więcej niż 1 echę.
plejotropia właściwa: gdy jeden gen wpływa bezpośrednio na więcej cech, ale nie maja one ze soba logicznego połaczenia. Np. u pomidora gen L warunkuje jednopiennośc i drobne owoce.
Plejotropia pozorna: gdy 1 gen wpływa bezpośrednio na 1 ceche, która zkolei powoduje zmiane kolejnych, z którymi jest powiązana, np. u kur gen h- brak haczyków piór pióra łamią się i stroszą strata ciepła wieksze spozycie pokarmu i przemiana materii wieksze rozmiary organów wewnętrznych zwiekszony transport słabsze przyrosty niższa mas a ciała, żywotnosć i niosność jaj.
współdziałanie genów różnych loci:
epistaza: rodzaj współdziałania alleli 2 par gdzie 1 gen jest nadrzędny w stosunku do drugiego.gen epistatyczny gdy się ujawni, to nie dopuszcza do ujawnienia się genu hipostatycznego
geny o działaniu jednoznacznym: dany fenotyp jest determinowany jest allelem dominującym róznych loci.
Komplementacja: allele róznych loci się dopełniaja, musza razem wystąpic, aby wywołac pewien efekt(dynia).
Co to jest zjawisko transgresji? Kiedy można uzyskać efekt transgresji? Jakie typy genów warunkują zjawisko transgresji? Przykład - wczesność u pomidorów (jak ją uzyskano w Katedrze, jakie powstały odmiany?)
Transgresja- wystepowanie w pokoleniu F2 i dalszych osobników o wiekszej i mniejszej wartości cech w porównaniu do form rodzicielskich. W hodowli transgresyjnej chodzi o nagromadzenie licznych, a jeśli to możliwe, wszystkich czynników dziedzicznych wpływających pozytywnie na ukształtowanie jednej określonej cechy. Zjawiska transgresji sa z reguły genetycznie kontrolowane przez geny polimeryczne, modyfikacyjne lub komplementarne.dzieki wspóldziałaniu takich genów zmienność poszczegolnych cech może w dalszychgeneracjach znacznie przekroczyc granicę charakterystyczne dla form rodzicielskich, a nawet spowodowac ujawnieni się nowych własciwosci. Transgresi w wyniku korzystengo połączenia czynników genetycznych można oczekiwac szczególnie w wypadku poligenicznych cech ilościowych, takich jak wielkość owoców, wysokosć roślin, niepodatność na wylęganie, zawartość białka itp. Ponieważ genetyczny skład odmiany na ogół nie jest znany, pomyślny wynik zalezy od mniej lub więcej przypadkowego doboru własciwych komponentów krzyzówkowych. Przypadkowość tę można w hodowli transgresyjnej w pewnym stopniu ograniczyc, wykonując na wyczucuie liczne skrzyzowania i wybierając do dalszejselekcji tylko te, w których na podstawie ścisłych obserwacji rodów pokoleń F1 i F2 stwierdzono występownie form wykazujących transgresje pożadancyh własciwości. Pochodzenie krzyzowanych partnerów rodzicelskich z różnych rejonów geograficznych oraz odmienny skład komponentów plonu, u obu tych form mogą również korzystnie wpłynąć na wynik hodowli transgresyjnej. Formy rodzicielskie fenotypowo różniące się znacznie między sobą pod względem danej cechy sa również najprawdopodobniej odmienne genetycnie i tym samym w wiekszym stopniu rokują nadzieję wystapienia transgresji, niż np. blisko spokrewnione odmiany. Tok postępowania w hodowli transgresyjnej jest taki sam jak w hodowli kombinacyjnej, więc mogą tu znaleźć zastosowanie wszystkie metody stosowane w hodowli kombinacyjnej.
Przykład wczesności u pomidorów: skrzyzowano dwie wczesne odmiany- amerykańską Beverlodge i rosyjska Nowosybirskaja- o średnich owocach i dosć dużej odporności na iską temperaturę, obie odmiany były dość podobne. Po skrzyzowaniu otzrymano dużo genotypów, wyselekcjonowano te, które już pod koniec czerwca dawały owoce. oTrzymana odmiane nazwano alfa, została ona jeszcze poprawiona i w końcowym efekcie otrzymano odmiane beta. W pokoleniu F2 wystąpiły różnice , ale niektóre rózniace się fenotypy wychwyono dopiero w pokoleniu F3. Rosliny krzyzowane były rózne (w loci) i dlatego po ich skrzyzowniu otrzymano nowe udoskonalone rosliny.
Poligeny: geny determinujące cechy ilościowe, wpływają na stopien nasilenia cech, im więcej zgromadzonych w zygocie alleli warunkujacych dana ceche, tym w większym nasileniu wystapi dana cecha. Poligeny odkryli: Nilsson-Ehle, East.. poligeny wykazują niewielkie efekty jednostkowe, działają w tym samym kierunku, efekty ich działania sumuja ssię przykłady cech: plon suchy masy, wzrost roslin, zawartość cukru, dł. okresu wegetacji cechy ilosciowe w duzym stopniu ulegaja wpływom środowiska.
Jakie metody hodowlane częściej stosuje się dla roślin samopylnych a jakie częściej dla roślin obcopylnych?
metody hodowli roslin samopłodnych:
rodowodow
ramszu
pojedyńczych nasion
populacji cząstkowych
metody hodowli roślon obcopłodnych:
odmian populacyjnych
rezerw
indywidualno-rodowa
odmian mieszańcowych F1(heterozyjna)
hodowli cyklicznej
hodowli odmian syntetycznych
Prawo Hardego i Weinberga a genetyka i hodowla. Czy jest tu jakiś związek?
Prawo Hardyaeo- Weinberg stanowi podstawe genetyki populacji, która zajmuje się badaniem alleli genów w populacji oraz siłami, które utrzymuja lub zmieniają częstośc poszczególnych alleli i genotypów w populacji. Lokalna krzyzujaca się populacja jest nazwana demem. Naturalne populacje składają się z wielu krzyzujących się osobników, każdy o unikatowej kombinacji alleli i gen ów, lecz dziela wspólna pul genowa. Ewolucja to zmiana częstości alleli w puli genowej populacji, dlatego genetyka populacji ma fundamentalne znaczenie. Hardy i weinberg niezaleznie opublikowali obliczenia pokazując, że niepożadane cechy nie mogłyby stac się powszechne z powodu rzadkich kombinacji stosunkowo powszechnych alleli. Pokazali oni, że częstosc poszczególnych genotypów w populacji rozmnażającego się płciowo, diploidalnego gatunku osiągaja równowage po jednym pokoleniu losowego kojarzenia i zapłodnienia oraz pozostaja stałe, chyba , że coś zmienia czestości allei w populacji. Równowaga hardyego-weinberga wymaga:
panmiksji
nieograniczenia
braku selekcji naturlanej
kojarzenia losowego
braku migracji róznicującej
braku mutacji lub jednakowej częstotliwości mutacji w obie strony
zgodność obserwowanej częstotliwosci genotypów z czestotliwością spodziewaną można sprawdzić za pomoca testu hi kwadrat.
Równanie H-W : p2 + 2pq +q2 = 1
P+H+Q = 1, gdzie p2 = AA, czyli p=A, 2pq= 2Aa, q2 = aa czyli q=a
Co do drugiej częsci pytania:
Związki: genetyka: prawo H/W podstawa genetyki populacji
Hodowla- podczas prowadzenia hodowli możemy utrzymywac pewna kontrole nad pula genów, wiec możemy sobie wyliczyć częstotliwość wystąpienia jakiegoś genotypu pozytywnego lub nie, będziemy mogli przewidziec ich częstotliwość. Prawo może być pomocne przy dobieraniu komponentów do hodowli z jakichś populacji.
Czym różni się zjawisko komplementacji i kumulacji genów?
Komplementacja: allele dominujące róznych loci wzajemnie się uzupełniają, musza wystapic jednoczesnie, aby wystąpił okreslony efekt. Brak któregokolwiek z tych genów sprawia, że dana cecha się wogole nie ujawnia, nawet gdyby były obecne. Stosunki: 9:7; 9:6:1; 9;3:4
Kumulacja genów: nagromadzenie w jedny miejscu dużej liczby genów takich samych lub o podobnym działaniu,np. kilka genów odporności na patogena w jednej roslinie.
Wykład 6. Krzyżowanie wsteczne, typy przenoszonych cech.
Kiedy stosuje się metodę krzyżowania wstecznego w hodowli roślin.
Krzyzowanie wsteczne stosujemy gdy chcemy wprowadzić jedna lub kilka cech, ale cecha musi być monogeniczna. Krzyżowanie to pozwala na ograniczenie liczby genotypów w rozszczepiających się pokoleniach. Krzyzowany jest homozygotyczny mieszaniec pokolenia F1 z jednym z rodziców. Krzyzowanie wsteczne powoduje ograniczenie genotypów i ułatwia wyselekcjonowanie pożadanych rekombinantów. Skutecznosć krzyżowania jest jest zatem wyraźna, gdy liczba genów jest wysoka.
Krzyzowanie wsteczne może być pomocne, gdy celem hodowli sa formy powstałe w wyniku rekombinacji cech sprzęzonych. Krzyżowanie z recesywnym rodzicem ujawnia w pierwszym pokoleniu B1 poządane recesywne rekombinacje, a w F2 przeważają heterozygoty fenotypowo diominujące. Krzyzowanie wsteczne polecane jest gdy udział pożądanych właściwosci u jednego z partenrów rodzicielskich jest wyższy niż u drigiego. Krzyżowanie wsteczne musi być z tym pierwszym. Wielokrotne krzyzowanie wsteczne zaleca się, gdy hodowca zamierza przenieść pojedynczą ceche do istniejącej odmiany z rasy bardzo odległej i różnej od tej odmiany. Rosliny mieszańcowe krzyzuje się wielokrotnie z ta odmianą rodzicielską, do której ma być wprowadzona nowa cecha. To wielokrotne krzyżowanie ma na celu ograniczenie udziału drugij formy rodzicielskiej do przekazania tylko jednej cechy. Metodę tę określa się jako hodowle wypierającą. Najlepiej gdy wprowadzana cecha jest dominująca i uwarunkowana jednoczynnikowo.
Przykład wprowadzenia genu dominującego (Omawiany na przykładzie Ccu - odp. na -parach dyniowatych Cladosporium cucumerinum, lub gen partenokarpii Pc.)
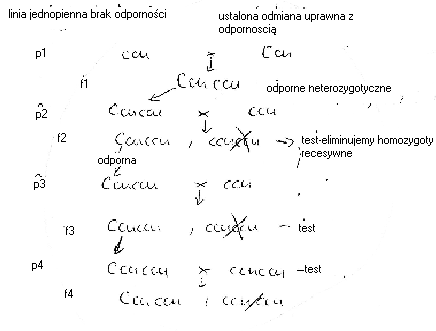
Wprowadzenie genu dominującego następuje szybciej, bo można stale krzyzowac osobnki heterozygotyczne odporne z odnianą czy linia do której wprowadza się cechę odpornosci. Do wprowadzenia genu dominującego potrzeba 6-7 lat.
Przykład wprowadzania genu recesywnego braku goryczy w owocach ogórka b i - brak goryczy. Jakie sprzężenie ułatwia testowanie roślin?
Przy samozapyleniu, tak jak teraz metoda wprowadzania genu jest wolniejsza, ale bezpieczniejsza w przypadku trudnej do testowania cechy i kłopotliwa jeśli test przeprowadzamy na dużych roslinach i zajmuja one duzo miejsca. Cecha goryczy i barwy kolców jest łatwa do testowania, bo testujemy siewki. Rosliny o gorzkich owocach mają gorzkie liście dlatego test może być na siewkach. Cecha goryczy jest sprzężona z barwa kolców i dlatego wybierając rosliny o białych kolcach wybieramy jednoczesnie rosliny pozbawione goryczy. Wprowadzenie tego genu trwa 10-12 lat z założeniem, że testujemy przed kwitnieniem, jeśli po kwitnieniu to liczbna krzyzowań musi być wieksza.
Jakie geny utrudniają postęp hodowlany w krzyżowaniu wstecznym?
Czarne kolce u ogórka waryunkuje gen B, ale sa inne trzy geny (b-2,b-3,b-4) warunkujące występowanie czarnych kolców. Geny t ze soba współdziałaja. Oprócz genu bi istenieje locus Bt, który działa niezależnie od bi. Postępowanie hodowcy będzie zalezało od tego z jakimi genami czarnych kolców i genami goryczy będzie miał do czynienia
Odmiana wieloliniowa . Jaki jest jej skład i kiedy jej otrzymywanie jest uzasadnione?
Odmiana wieloliniowa powstaje w wyniku wielokrotnego krzyżowania wstecznego oraz w wyniku zmieszania kilku linii identycznych lub bardzo podobnych pod względem cech morfologicznych i jakościowych, ale rózniących się odpornoscia na tę samą chorobę (linie izogeniczne). Liczba wybieranych linii:6-18. Otrzymujemy je wtedy kiedy chcemy w krótkim czasie otrzymac odmianę odporną na kilka ras danego patogena. Nie ulega ona szybkiemu zakażeniu, ani całkowitemu zniszczeniu, w rzie pojawienia się nowej razsy patogena zakażeniu ulegaja tylko te rosliny wrazliwe na tą rase. W razie potzreby odmianę porażona można zastapić inna linia. Zaleta odmiany wieloliniowej jest krótki czas jej wytwarzania ok. 5-6 lat. Wykorzystuje się ją w uprawie pszenicy.
Wykład 7 i 8. Hodowla heterozyjna, hodowla cykliczna.
Czym różni się metoda hodowli cyklicznej od metody hodowli heterozyjnej?
Hodowle cykliczna stosuje się gdy cecha lub cechy warunkowane sa wieloma genami o małym jednostkowym działaniu i sa trudne do oceny. Metoda polega na sukcesywnym prowadzeniu cykli hodowlanych, na które składa się przekżyzowanie w populacji i samozapylenie wybranych osobników. Swobodne przekzyżowanie i samozapylenie mogą obejmowac jedno lub kilka pokoleń. Materiałem wyjściowym przy selekcji cyklicznej może być dowolna heterozygotyczna populacja czyli panmiktyczne odmiany populacyjne, syntetyczne lub heterozyjne. Populacje uzyskane tą metoda mogas byś wykorzystane do tworzenia linii wsobnych jako komponenty odmian mieszańcowych lub syntetycznych bądź jako materiał wyjściowy do dalszej selekcji. Selekcja cykliczna może być: zwykła, jako test na ogólna lub swista wartość kombinacyjną lub wzajemna selekcja cykliczna.
Hodowla heterozyjna: w jej wyniku otrzymujemy mieszańce pokolenia F1 o wyższej bujności, zywotności, plenności w stosunku do form rodzicielskich. Do mieszańców heterozyjnych można w krótkiim zasie wprowadzić różne geny odporności, można tez modyfikowac cechy owoców, dobierając odpowiednie formy rodzicielskie. Hodowla heterozyjna rozpoczyna się od otrzymania linii wsobnych homozygotycznych z różnych odmian i populacji. Przy zbyt wąskiej zmienności materiału wyjściowego, można wybrane materiały krzyzowac i dalej prowadzic samozapylenie i selekcje. Samozapyla się zwykle przez 6-10 pokoleń, az do pełnego wyrównania cech. oDrzuca się linie słabe, chore, obarczone defektem.nastepnie wybieramy linie o najlepszej wartości kombinacyjnej (topcross,polycross). Linie najlepsze w teście na ogólna wartość kombinacyjna oceniamy w tescie na swoista wartość kombinacyjną a potem krzyzujemy w układzie diallelicznym. Po tym tescie będziemy wiedziec, która z linii jest lepszą linią mateczna, a która ojcowska.
Teoretyczne podstawy zjawiska heterozji.
Heterozygotycznosć- homozygota dominująca i recesywna. Geny warunkujące plennosc miayby w stanie heterozygotycznym działać silniej niż w homozygotycznym. Inaczej spadek plenności w wyniku sukcesywnego samozapylania nalezałoby przypisac narastajacej homozygotyczności lini wspobnych. Utrwalenie heterozji metodami hodowlanymi wg. tej hipotezy jest niemozliwe. Tłumienie genów szkodliwych. 2. Dominacja- działanie poządanych alleli dominujących rodziców uzupełnia się u mieszańca i może on wykazać wyższą plenność niż materiał wyjściowy. Przeszkadzaja tutaj negatywne sprzężenia genów dominujących z recesywnymi. Rośliny obcopylne tworzą populacje złożone z osobników genetycznie niejednorodnych, w tym również obarczonych szkodliwymi recesywnymi genami, które dzięki heterozygotyczności sa utajone, a ujawniaja się w wyniku samozapylenia powodując wystapienie defektów w liniach wsobnych. 3. Naddominacja: allele danego genu mogą działac niejednakowo w zalezności od tego czy wystepuja w postaci homo czy heterozygotycznej. W stanie heterozygotycznym allele tego samego genu mogą działać silniej.
Kolejne etapy otrzymywania odmiany heterozyjnej (na przykładzie ogórka, pomidora, marchwi, kapusty).
Ogórek- mieszaniec pojedyńczy szklarniowy
(linia mateczna tzw. Twarda ) MMFF x MMFF (linia ojowska tzw. Miękka po azotanie srebra) reczne zap.
F1. MMFF ♀
Mieszaniec pojedyńczy polowy:
Linia mateczna MMFF x MMff linia ojcowska jednopienna ♂♀ zapylenie przez pszczoły
F1: MMFf ♀ mniej stabilny od homozygoty FF. Replikacja formy matecznej odbywa się przez samozapylenie w izolacji przestrzennej
Pomidor:
Jadrowa meska sterylnosć zalezy od genów jądrowych :ms, sl, ps1,ps2,ex - mutanty wykporzystywane w produkcji nasion mieszańcowych. Idealny mutant będacy kiniąmatecznąpowienin charakteryzowac się:
całkowita męska sterylnością w warunkach w których produkowane są nasiona mieszańcowe, zapobieganie samozapyleniu.
Normalna żeńska płodnosć, kwiaty ani owoce nie mogą mieć nasion
Całkowita recesywność alleli steryklkości, nie może być żadnych złych cech przekazywane na mieszańce.
Marchew:
Cytoplazmatyczno jądrowa meska sterylnosć
Linia męskosterylna A SaaBB
Dopełniająca B NaaBB
Płodna(ojcowska) C N_ _ _
Dopełniająca B' NaaBB
Mieszaniec pojedyńczy:
Linia A SaaBB ♀ x linia C ♂N_ _ _ F1 (AxC)
Mieszaniec trójliniowy
P1: ♀linia A SaaBB x ♂linia B' NaaBB F1 (axB') SaaBB - sterylny
P2: ♀(AxB') SaaBB x ♂linia C N_ _ _ F2 (AxB') x C SaaBb
Reprodukcja lini męskosterylnej A
♀linia ASaaBB x ♂linia B dopełniająca NaaBB liniaA SaaBB
linie muszą wykazywac dobra wartość kombinacyjna.
Kapusta
wybór pojedynków z kolekcji odmian- rozmnazanie wegetatywne
po ścieciu główki otrzymujemy odrosty, łatwo się one ukorzeniają, przechodza one jaowizację i poddajemy je samozapyleniu. Zapylenie to nastepuje w stadium paka 2-3 dni przed normalnym zakwitaniem. Rozchylamy pąk i zapylamy ręcznie pyłkim tej samej rosliny. Samozapyleniejest możliwe, ponieważ nie działaja bariery samoniezgodności, uzyskujemy tym sposobem pseudozgodność
miesaniec pojedyńczy dwukierunkowy:
linia A S1S1 x linia B S2S2 F1 (AxB) S1S2
mieszaniec podwójny:
linia A S1S1 x LINIA B S2S2 F1 (AxB) S1S2
LINIA C S3S3 x LINIA D S4S4 F1(CxD) S3S4
F1(AxB) S1S2 x F1(CxD) S3S4 F1(AxB)S1S3, S2S3 x (CxD) S1S4 ,S2S4
Dlaczego w pewnych gatunkach roślin warzywnych mamy wiele odmian heterozyjnych a w innych wcale?
Sałata, groch fasola, bób- brak odmian heterozyjnych. Przeprowadzenie kastracji u roslin z bardzo małymi kwiatami jest trudne i z jednego kwiatu uzyskujemy mało nasion. Nie ma tez odmian mieszańcowych iu pszenicy i jeczmienia. Nowa odmiana mieszańcowa musi być opłacalna, musi dawac wyższy plon lub jakosć, nowa atrakcyjna cecha kwiatu lub liscie ozdobne. Mieszańxe można łatwiej wyhodowac niż linię ustaloną, łatwiej jest wprowadzić geny odporności do odmiany mieszańcwoej.
Podaj typy męskiej niepłodności i ich wykorzystanie u różnych gatunków uprawnych (cebula, marchew, pomidor)
Cebula- cytoplazmatyczno-jądrowa męska sterylność. Wykorzystujemy ten typ do tworzenia mieszańców heteruzyjnych.
pojedyńczy
A (mateczna sterylmna) msms x męska płodna C MsMs
Msms nasiona mieszańcowe
trójliniowy tworzymy aby uniknać problemu pozyskiwania nasion F1, który występuje u mieszańca pojedyńczego.
A msms x B' (płodna) msms AB' msms
AB' x C AB'C
podwójny
A msms x B' msms AB' msms (sterylny)
A msms x C MsMs Msms AC (płodny)
AB' msms x AC Msms F1 (AB'Msms x Acmsms )
marchew 0 sytoplazmatyczno jądrowa męskosterylność, decydują o niej 2 geny jądrowe. Typ łatkowy: pylniki zamienione w płatki korone- zielone. Typ brązowe pylniki- wcześnie zaschnięte pylniki.
pojedyńczy
♀ A x C ♂ AC
trójlioniowy
♀A aa BB x B' aa BB♂ AB' aa BB
♀AB' x C♂ F1 (AB') x C AaBb
pomidor męskasterylność. Homozygoty recesywne dają sterylnosć i sa one komponentem matecznym. Musi być: stabilna sterylność, normalna żeńska płodność, całkowita recesywność alleli sterylności, mnie może być żadnych złych cech przekazywanych na mieszańce.
Cytoplazmatyczna męska sterylnosć (powodowana przez mitochondria)
Typ cytoplazmy: u kukurydzy
N- normalna, pyłek płodny
T,C,S - pyłek sterylny
restorery
u ludz defektyymitochondrialne powodują
Gen Rf- znajdujące się w mitochondrium w genie męskosterylnym przywraca płodność.
Omów geny męskiej niepłodności na przykładzie pomidora. Które z tych genów wykorzystuje się w hodowli odmian pomidora szklarniowego? (ex, ps, sl, ms)
Geny recesywne - wszystkie
ms- ograniczenie wydzielania pyłku i sterylności
sl - kwiat nie ma pręcików, niesie zdeformowanie owoców (nitkowe, stasmione)
ps -, ps2 - pręciki- ich główki mają grubszą ściane niż normalne, nie ulegaja pęknięciu i pyłek nie wysypuje się.
Ex - heterostylia- długość szyjki słupka większa niż normalnie, pyłek nie osypuje się na znamię.
Geny ps,ps2 i ex- dają najlepsze mutanty, normalne ładne kuliste owoce- wprowadzone do lini matecznej
Omów zjawisko samoniepłodności (samobezpłodności) na przykładzie kapusty (jaki jest to typ samobezpłodności ?) W jaki sposób samobezpłodność można wykorzystać w produkcji nasion mieszańcowych kapusty (mieszańce 4 liniowe)?
U kapusty samoniezgodność warunkują allele wielokrotne. Samoniezgodność może być gametofityczna lub sporofityczna. Zjawosko to wykorzystujemy do tworzenia mieszańców heterozyjnych podwójnych .
Hodowla mieszańców:
wybór pojedynków z kolekcji odmian - rozmnan zanie wegetatywne, odrosty łatwo się ukorzeniają, rosna i uzyskujemy pokolenie wsobne S1.
rozmnożenie generatywne: zapylenie w stadium pąka. Poddajemy je samozapyleniu aby pokonac bariery samoniezgodności dokonujemy zapylenia w stadium pąka. Otwieramy jeszcze niedojzałego paka i nanosimy pyłek z rośliny z otwartymi kwiatami -pseudoniezgodność. Lub nansosimy pyłek na dojrzałe kwiaty, gdzie znamiona słupka wczesniej zroszono sola kuchenna.
ocena linii S1 w polu: tak samozapylamy przez 3-4 pokolenia wsobne by była linia. Wykonujemy krzyżowanie w ukladzie diallelicznym. Tam gdzie uzyskamy nasiona uzyskujemy mieszańce próbne. Prowadzimy hodowlę porównawczą i wyprowadzamy nasza linię.
mieszańce podwójne 9czteroliniowe)
I S1S1 (liscie jasnozielone) x S2S2 (l. jasnozielone) S3S3 (l.ciemnozielone) x S4S4 (l. Ciemnozielone)
S1S2 ( liście jasnoziel.) x S3S4 (liscie ciemnoziel.)
S1S3 ; S1S4 ; S2S3 ; S2S4 odmiany mieszańca
Omów wykorzystanie roślin genetycznie żeńskich w produkcji mieszańców heterozyjnych ogórka? Jakie geny warunkują żeńskość ogórka? Jak rozmnaża się linie żeńskie? Jakie pod względem płci mogą być komponenty rodzicielskie mieszańca F1?
Geny u ogórka:
F,f- rosliny z genem F sa żenskie
M,, - gen m - wpływa na powstanie kwiatów obypłciowych
U ogórka wykorzystujemy rozdzielnopłciowosc do tworzenia mieszańców. Nie trzeba kastrować bo linia mateczna wytwarza tylko kwiaty żeńskie. Tworzenie mieszańców:
KRZYŻOWANIE LINII ŻEŃSKICH Z JEDNOPIENNĄ (uprawy polowe)
P: MMFF (linia mateczna) x MMff (linia ojcowska jednopienna) zapylene przez pszczoły
F1: MMFf mieszaniec heterozyjny
KRZYŻOWANIE LINI ŻEŃSKIEJ Z LINIĄ ŻEŃSKA (do szklarni)
P: MMFF (linia mateczna żeńska) x MMFF (linia ojcowska żenska) płeć modyfikowana przez oprysk azotanem srebra
Zapylenie ręczne
F1:MMFF
KRZYŻOWANIE LINII ŻENSKIEJ Z LINIA MĘSKĄ
P: MMFFAA (linia mateczna żenska) x mmffaa (linia ojcowska męska) zapylenie przez pszczoły
F1: MmFfAa mieszaniec żeński
Pojęcie ogólnej i swoistej wartości kombinacyjnej.
Wartość kombinacyjna linii mierzona plennością potomstw F1: im wyższy plon mieszańców tym wyższa jest zdolnosć kombinacyjna danych linii.
ogólna wartość kombinacyjna - zdolność danej linii do wydania bujnego potomstwa przy krzyżowaniu w wieloma innymi liniami topcross.
specyficzna wartośc kombinacyjna - charakteryzująca zdolność dwóch linii do wydania określonego bujnego potomstwa przy wzajemnym przekrzyżowaniu.
Kiedy uzasadniona jest hodowla odmian heterozyjnych?
Zjawisko heterozji jest uzasadnione i ma sens gdy:
gdy krzyżowane formy są możliwie wysokiej homozygotyczności
gdy krzyżowane formy mają wysoka ogólna i swoistą wartość kommbinayjna
gdy po samozapyleniu nie ma dużej deprsji wsobnej
gdy po skrzyżowaniu otrzymamy nasuina F1
gdy uzyskamy dużą bujność i plenność
Odmiany heterozyjne a odmiany syntetyczne - różnice i podobieństwa.
w hodowli syntetycznej nie stosujemy kastracji ani sztucznego krzyżowania
komponentami jest w syntetycznej 6-10 linii najlepszych, zas w heterozji 1-4 linie najlepsze.
W hodowli heterozyjnej stosujemy kastrację i sztuczne krzyżowannie
Produkcja hodowli odmian syntetycznych jest o wiele tańsza
Wybrane linie w odmianie syntetycznej przepylają się wzajemnie
Nasiona F1- to mieszanka kilku linii u odm. Syntetycznej
W odmianie heterozyjnej wykorzystujemy tylko nasiona F1, póxniej efekt heteroji spada
U odmiany syntetycznej efekt heterozji utrzymuje się przez 2-3 pokolenia
W heterozji stosujemy test topcross zaś w syntetycznej polucross.
W oby metodach chodzi o wyższy plon.
Hodowla cykliczna. Na czym polega i kiedy ją stosujemy?
Materiałem wyjsciowym selekscji cyklicznej może być dowolna heterozygotyczna populacja czyli panmiktyczne odmiany populacyjne, syntetyczne lub heterozyjne.. populacje uzyskane w jej wyniku mogą być wykorzystane do róznych celów: do tworzenia linii wsobnych, ale również jako komponenty odmian mieszańcowych lub syntetycznych jako materiał do dalszej selekcji. Selkcja cykliczna znana jest w 4 wariantach:
sel. Cykl. Zwykła
samozapylenie heterozygotycznych pojedynków
rozmnozenie najlepszych pojedynków i krzyżowanie ich potomstw we wszystkich możliwych kombinacjach.
Wysiew populacji uzyskanych w wyniku krzyzowania i samozapylenia wybranych pojedynków az do uzyskania populacji o pożądanych własciwościach.
se. Cykl. Jako test na ogólną wartosć kombinacyjną i
se. Cykl. Jako test na swoistą wartość kombiancyjna
tu właczane jest badanie wartosci kombincyjnej krzyżowanych potomstw pojedynków. Jeśli jako tester występuje genetycznie niejednorodna odmiana lub populacja test daje odpowiedź dotyczacą OWK. Natomiast jeśli testerem jest jednorodna linia wsobna, wynik testyu dotyczy SWK.
wzajemna selekcja cykliczna
uwzględnia zarówno OWK jak i SWK. Materiałem wyjściowym sa dwie różne populacje mozliwie odmiennego pochodzenia . w kazdej z nich pewna liczba roślin zostaje samozapylona. Jednoczesnie te same rosliny sa kerzyżowane AxB i BxA. Nasiona uzyskane z self zostaja w rezerwie do 3 etapu cyklu, natomiast nasiona pochodzace ze skrzyzowania służ a do założenia w 2 roku doświadczenia porównawczego.=> ustalamy, które pojedynki najlepsze na mieszańce. Pozostałe rezerwy zostaja wysiane w 3 roku i przekrzyżowane we wszystkich możliwych kombinacjach. Na tym kończy się pierwszy cykl. W 4 roku rozpoczyna się drugi cykl, w którrym materiałem wyjściowym sa najlepsze populacje A' i B'. Z populacji uzyskanych w 2,3 i jeszcze późniejszym cyklu mogą być wybrane pojedynki, tóre stanowią materiał wyjściowy dla linii wspobnych do krzyzowania pojedyńczego lub podwójnego lub tez populacje te mogą być zmieszane i przekrzyżowane w celu uzyskania nowej populacji w wysokim stopniu heterozygotycznej.
Porównaj metodę hodowli heterozyjnej i transgresyjnej?
Metoda hodowli heterozyjnej |
Metoda hodowli transgresyjnej |
Wybieramy osobniki nie spokrewnione, otrzymanie zróżnicowanych genetycznie linii wsobnych, badanie owk, test pollycross, topcross, swk- krzyżowanie dialleliczne, rozmnozenie linii metoda kojarzenia siostrzanego: wiatropylne, owadopylne. Tworzenie mieszańców z najlepszych lini. |
Przypadkowy wybór komponentów krzyżówkowych czyli wykonanie "„a wyczucie"”licznych krzyzowan i wybór do dlszej selekcji tych , w których na podstawie ścisłej obsdeerwacji rodów pokolenia F1 i F2 stwierdzono wystepowanie form wykazujacych transgresje pożądanych właściwości. Ochodzenie krzyżowanych partnerów rodzicielskich z órznych rejnów geograficznych oraz odmienny skład komponentów plonu u obu tych form mogą również korzystnie wpłynąć na wyniki hodowli tranmsgresyjnej. |
Wykład 9. Wykorzystywanie krzyżowania oddalonego w hodowli roślin.
Omów właściwości i pochodzenie pszenżyta - nowego gatunku otrzymanego przez człowieka.
Triticum aestivum AABBDD x secale cereale RR
AABBDDRR pszenżyto oktoploidalne
Pszenzyto pierwotne:
Pszenica AABB x zyto RR
ABR kolchicyna - AABBRR
AABBDD x RR
ABDR -- > kolchicynowanie AABBDDRR
Stabilnosć cytogenetyczna
Płodność
Plon
Stopien samopłodności
Cechy paszowe
Zimowanie
Wylęganie
odporne
Omów na przykładzie pomidora możliwości wykorzystania gatunków dzikich w hodowli gatunku uprawnego.
Wiele dzikich gatunków pomidora daje się krzyżowac z uprawnymi. Dzięki temu możemy wykorzystywać geny odporności i wprowadzic je do odmian uprawnych. Obecnie najgroxniejszym patogenem w uprawie rego warzywa jest TSMV. Jest on przenoszoy przez wciornastka zachodniego.. ochrona polega na eliminacji wektora tego wirusa, ale jest ona bardzo trudna. Dzięki hodowli możemy przenieść geny odporności z gat. Dzikich krzyzując je z gat. Uprawnymi. Po wprowadzeniu genów odporności poprzez kilkakrotnr krzyżowanie wsteczne z gat. Uprawnym otrzymuje się odmiane plenna.. należy pamietac, ze odpornosć tej odmiany zalezy tez od tego, gdzie gen wprowadzilismy, czy dobrze będzie funkcjonował, czy będzie przekazywany w nast. Epnych pokoleniach w czy nie zostanie odrzucony podczas mejozy. Odmiana Stivens: wysoka odporność na TSWV - w polsce nie ma ras wirusa, które by przełamały tę odporność. Niestety odmiana ta nie jest dobra do krzyżowania.
Podaj charakterystykę mieszańców otrzymanych przez krzyżowanie dyni olbrzymiej z dynią zwyczajną.
P: cukinia(spłaszczona, liści obbłe) x dynia olbrzymia ( okrągły, lisci epowcicnane)
F1:spłaszczony, okrągły i liscie lekko powcinane.
W szerokim stopniu niepłodne, mimo, ze rodizce maja tę samą liczbe chromosomów, wysika homologia, nie ma zywotnych gamet- mieszańce utrzymujemy za pomoca kultury zarodków(omijanie barjery). Krzyzowanie mieszańców z formami rodzicielskimi jest rtudne, cechy trudno prekazywane.
Wykład 10. Hodowla poliploidalna (głównie haploidy)
Podaj kilka przykładów odmian tetraploidalnych otrzymanych przez człowieka. Pod względem jakich cech przewyższają diploidy? (tetraploidalna frezja - większe kwiaty, tetraploidalna koniczyna - większe liście, większy plon masy zielonej, tetraploidałna rzodkiewka - szybko rosnące zgrubienia).
Triticum monococcum x aegiles spelboides AB AABB Triticum diccocum, t. Durum, T. Turgidum
Brassica dereceae x B. Campestris AB AABB B> napus (rzepak)
Odmiany tetraploidalne tworzone sa głównie dlatego, iż otrzymuje się z nich wiekszy plon.
frezja- większe płatki, ale nie pachną
koniczyna- większa masa zielona z hektara, ale trudności z uzyskaniem nasion)
rzodkiewka- grubsza, wolniej parcieje
Podaj kilka przykładów odmian triploidalnych (burak cukrowy - większa zawartość cukru, banan - owoce bez nasion, podobnie kawon, winorośl, śliwa, jabłoń triploidalna), jak są one rozmnażane?
Patrz pytanie 14.
Przydatność haploidów w hodowli roślin na przykładzie (np. szparaga, ziemniaka, jęczmienia lub innych). Wykorzystanie: do badan genetycznych, taksonomicznych, hodowlanych, badanie mejozy, mitozy , rekombinacji.
Umozliwiaja ujawnianie cech warunkowanych genami recesywnymi. Służą do tworzenia lini homozygotycznych.
Diploidy: doskonałe homozygoty, służa do tworzenia mieszańców F1, sa materiałem wyjsciowym do hodowli.
Wykład 11. Hodowla mutacyjna - przykłady praktycznego wykorzystania mutantów w hodowli roślin.
Mutacja genowa: sa to mutacje dotyczace pojedyńczych genów indukowane napromieniowaniemlub czynnikami chemicznymi. W mutacjach punktowych do ich indukcji wykorzystujemy u ogórka iminę. Zmiany po jej zastosowaniu: wzrost 31,3% wyższy, zmiana w chlorofilu 21,3%, w owocach 13,1%, liściach 12,1%, w cechach fizjologicznych 6,5%, w cechach sterylności 5,6%, zmiany płci 4,3%, zmiany w kwiatach i pąkach 4,6%.
Mutacje te badano w F2 ponieważ w F1 nie były jeszcze widoczne, wiąże si ę to z tym, że mutacja jest warunkowana allelm recesywnym dlatego nie ujawnia się gdy mamy heterozygotę.
Wygląd przed mutacja: dojrzała roślina ma długie pędy, ok. 10 owoców, strsze na nasiennik, lezy na podłozu.
Po mutacji: karłowata, krótkie pędy, 4 owoce, krzaczasta, dobra na zbiór mechaniczny, lecz mało owoców.
Prace nad mutantem krzaczastości: wysokie nasilenie żeńskości, partenokarpia, białe kolce, wysmukłe owoce, wartości smakowe, tolerancja na mączniaka
Odmiana krzaczasta to DAR, cechy: zwarty pokrój, ładne, wydłuzone owoce, jedyna krzaczasta odmian
Wykład 12. Hodowla odpornościowa na warunki stresowe, choroby i szkodniki.
Typy odporności na choroby. Czy warto prowadzić hodowlę odpornościową na choroby (podaj przykład)? I
Typy odporności na szkodniki. Czy warto prowadzić hodowlę odpornościową na szkodniki (podaj przykład)?
Typy odpornoscina chroby:
odporność typu niegospodarza, roslina, która nie jest gospodarzem, nie jest porazona
immunia- pełna odporność
odporność pionowa (OPN)
odpornosc pozioma (OPZ)
fitoabksyny - mikrocząsteczkowe związki hamuj ące rozwój mikroorganizmów
odporność indukowana- interferencja wirusowa( mało zjadiwy wiru przeciw bardziej zjadliwemu)
typy odpornosci na szkodniki:
pozorna- spowodowana brakiem szkodnika (szkodliwa)
odporność rzeczywista: brak akceptacji- szkodnik nie akceptuje rosliny i odlatuje; antybioza: niekorzystny wpływ pokarmu roslinnego na rozwój lub przezywalność, np. larw; toleruje - niwelacja uszkodzeń przez dalszy wzrost lub odrastanie.
Metody hodowli:
> odmiany wieloliniowe - wysiew odpornych i wrazliwych
odnalezienie roslin odpornych i wyprowadzenie odmiany odpornej.
Stosowanie dzikiej odpornej podkładki, krzyzowanie z nimi i wyprowadzanie odmiany odpornej.
Za i przeciw wprowadzaniu odporności pionowej i poziomej.
Odporność pozioma: poligeniczne: może ulegać wpływom modyfikacujnym. Częściej wykorzystywana jest odporność pionowa: łatwiejszy mechanizm dziedziczenia odporności, łatwiejszy do wykorzystania w hodowli, różnica pomiędzy formami wrazliwymi a odpornymi ujawnia się już w stadium siewki. Można tez prowadzic testy odpornościowe systemem tasmowym w labolatorium. Odporność pozioma bardziej uniwersalne zabezpieczenie, trwała, niezaleznie od zmian patogena wyklucza to możliwość utraty odpornosci.
Jakie typy odporności można wyprowadzać z gatunków dzikich do pomidora uprawnego? - odpornosć na choroby, pyt. 51
Omów problemy hodowli odpornościowej na przykładzie mączniaka rzekomego u ogórka.
Nie ma całkowitej odporności, jest tylko pewien rodzaj tolerancji. Po 2,5 tygodniach można uzyskać tylko 4% porażenia. Rózne odmiany charakteryzują się różną odpornością. Gdy jest tolerancja0 oprysk można wykonać stosujac okres karencji. W odmianach nieodpornych objawy narastaja bardzo gwałtownie.
Testowanie roslin na obecność mączniaka rzekomego:
testy polowe: naturalna infecja i naturalne warunki do rozwoju patogena
testy laboratoryjne: nie odpowiadaja testom polowym - wykonujemy na lisciach.
Możemy spowolnić rozwój patogena by dojrzały owoce, ale 100% odpornosci nie ma na tego patogena.
Porównaj metodę hodowli odpornościowej na choroby i na szkodniki
Rodzaje odporności:
odporność poziomowa: wyspecjalizowana w stosunku do prototypu, oligogeniczna, często nadwrazliwość, rzadko trwała, ekspresja jakościowa, łatwe wykorzystanie, podlega cyklom rozwoju i zaniku, wystepuje tylko u roslin rocznych.
Ruchliwe patogeny przenoszone z wiatrem- skuteczne pod wrunkiem stosowania w odpowiednim czasie i miejscu populacji wieloliniowcyh i mieszankach z zachowaniem izolacji przestrzennej.
Mao ruchliwe: może być bardzo skuteczna przeciw prototypom zlokalizowanym.
OPM : oporność prionowa: nie wyspecjalizowana w stosunku do protowypu, poligeniczna, brak nadwrazliwosci, ekspresja ilościowa, trudniejsze wykorzystanie, stabilna, trwała, występuje u roslin rocznych i wieloletnich.
Wszystkie patogeny - wymagaja uzupełniania innymi metodami zwalczania chorób.
W hodowli odpornościowej wykorzystuje siękrzyżowanie roslin odpornych z wrazliwyymi. Tło genetyczne odrzucane jest przez krzyżowanie wsteczne. Po każdym krzyżowaniu wykonuje się testy na odporność.
testy laboratoryjne: na siewkach, młodych liściach lub na liściach- zakaza się je i prowadzi obserwcje.
Testy polowe: wprowadzamy roślinyuważane za odporne i zakaza się lub czeka na naturalne infekcje, nastepnie prowadzi się obserwacje.
Testy laboratoryjne muszz abyć potwierdzone testami polowymi.
Wyszukiwarka
Podobne podstrony:
Kolokwia,egzaminy, Dzienni07, Wyniki z zaliczenia ćwiczeń i wykładów z Genetyki i hodowli roślin ogr
egzamin z hodowli, sem. 4, Hodowla roślin
hodowla egzamin, hodowla roślin
hodowla kolos - wyklady, sem. 4, Hodowla roślin
hodowla roslin 3
HODOWLA ROŚLIN ĆWICZENIA
Ściąga 6 z Hodowli Roślin i Nasiennictwa
genetyka molekularna i hodowla roślin, W14R03, Wykłady z genetyki i hodowli roślin ozdobnych, Sulech
Ściąga 2 z Hodowli Roślin i Nasiennictwa
DETARDANTY, sem. 4, Hodowla roślin
HODOWLA ROŚLIN ĆWICZENIA
HODOWLA ROŚLIN-test, Imię i nazwisko……………………
1 5 Rodowód hodowli roślinnych komórek, tkanek i organów
zagadnienia , Ogrodnictwo, Ogrodnictwo UP Wro, ROK II, semestr III, genetyka i hodowla roślin
Ściąga 3 z Hodowli Roślin i Nasiennictwa
hodowla roślin
Hodowla roślin samopylnych
więcej podobnych podstron