0000051 3

V/LJ»Ł/ / AA
oczach i normalnie wykształconych skrzydłach (typ dziki, ponieważ Tymczasem błąd w
czasie pierwszego podziału zygoty (mitotycznego!) prowadzi do utraty chromosomu ojcowskiego przez jeden z dwóch blastomcrów I-szcgo rzędu. Ten z prawej zawiera więc prawidłowy, heterozy-gotyczny układ X"X" **, drugi zaś X’"*. Ze względu na nieskomplikowaną mechanikę rozwojową taka asymetria utrzyma się w czasie dalszego rozwoju. Tak więc w połowic ciała dorosłej muszki komórki będą miały zmniejszoną liczbę chromosomów, w drugiej zaś ich liczba będzie normalna.
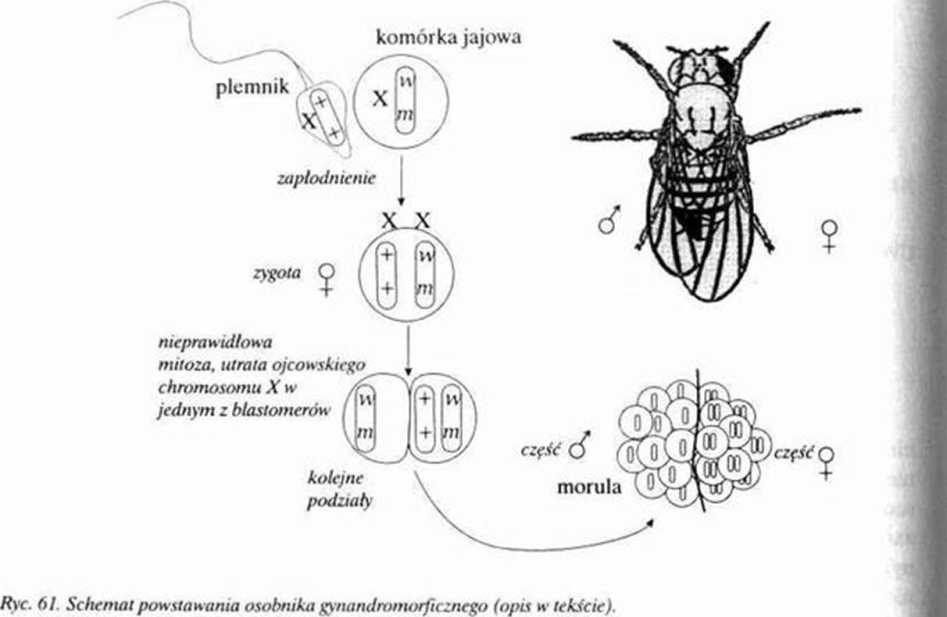
Z „pierwszej — lewej" części zarodka (X0) rozwinie się fenotypowo męski fragment ciała
' - - - —
(jednak jego niedorozwinięte gonady nie będą produkowały gamet), z „drugiej — prawej" żeński fragment (XX) o normalnych funkcjach fizjologicznych. Potwierdzeniem różnic wynikających z
Z analizy genetycznej gynandromorfów można wyprowadzić też wniosek, że część genów determinujących cechy budowy ciała dla obu płci leży na chromosomie X. natomiast sposób ich ekspresji zależy od składu heterosomów. Czy tylko? Prace Amerykanina Bridgesa prowadzone w
dymorfizmu płciowego są inne cechy fenotypowe takiego osobnika. Lewa strona ciała ma białe oczy i zredukowane skrzydła (ujawniają się więc cechy reccsywne). prawa zaś ma „normalnej* 1 cechy dominujące (oczy czerwone i skrzydła typowych rozmiarów).
latach 20-tych doprowadziły do szeregu ciekawych odkryć.
PLEĆ U DROSOPHIl^A JEST WYNIKIEM RÓWNOWAG1 POMIĘDZY GENAMI CHROMOSOMÓW X, A GENAMI AUTOSOMALNYM1
Bridges badał liczne mutanty muszki owocowej określając w nich stopień wykształcenia płci
w stosunku do wzorcowych samców oraz samic. Stwierdził, że u tego gatunku bardzo ważne są

proporcje chromosomów pici do autosomów (por. Ryc. 62). Jego oryginalna teoria zostaia dzisiaj nieco uaktualniona. Zakłada ona, iż u wielu gatunków zwierząt determinowanie jednej z pici, tzw. podstawowej przebiega niejako samorzutnie, wg pierwotnego wzorca ekspresji genów. Z kolei wykształcenie tzw. płci dominującej wymaga dodatkowego sygnału „przckierunkowującego roz-svój osobniczy (nieco inna ekspresja genów). U muszki owocowej podstawową jest płeć meska. która powstaje wówczas, gdy osobnik ma tylko jeden komplet genów sprzężonych z X. „Siła” ekspresji tych ok. 1000 genów jest na tyle niewielka (współczynnik X/A = 0,5), że autosomalnc cechy rozwijają się wg wzorca męskiego. Dopiero obecność drugiego chromosomu X u samic wzmacnia efekt działania pewnych genów na tyle, że wzorzec zmienia kierunek rozwoju w stronę żeńską (współczynnik XJA = 1). W uproszczeniu można powiedzieć, żc „siła” ekspresji 2000 genów jest wystarczająca do wytłumienia autosomalnych genów męskich.
O tym, jak ważna jest proporcja heterosomów do autosomów świadczy fakt, że np. z zygot o kariotypic 3A + XX wyrastały interseksy — osobniki o pośrednich cechach płciowych, mimo iż posiadały dwa chromosomy X! (współczynnik XJA = 0,67). Jeśli zygoty miały 3A+XY. wyrastały / nich tzw. nadsamce (współczynnik XJA = 0,33), mimo posiadania normalnego, „samczego” garnituru heterosomów. Co ciekawsze, z zygot 2A + XXX (współczynnik XJA = 1,5) wyrastały tzw. nadsamice. Wbrew obiecującej nazwie, muszki takie są sterylne, słabe i mało żywotne, chociaż mają wyraźnie podkreślone cechy płciowe. Wszystko to dowodzi, że odchylenia współczynnika XJA zarówno w dół od 0,5, jak i w górę powyżej 1 prowadzą do nasilających się zaburzeń rozwojowych (por. oś na Ryc. 62).
2n = 2A + XX normalna samica
2X
2A
= 1
2n - 1 = 2A + X0 sterylny samiec
I* =05
2A 10
IX
2n = 2A + XY normalny samiec (jeśli jest Y)
!2L -0<5
2A U"
3n - 1 = 3A + XY « 0.33
nadsamiec
3n - 1 = 3A + XX interseks
2X_ _ n <7
3A - 0 67
0.33
0,67
?
nadsamica
1,5
_l_
2n + 1 = 2A + XXX ~ = 1.5
nadsamica
2n + 1 - 2A + XXY ^ = 1
0.5
normalny
samice
normalna
samica
samica
3X
3n = 3A + XXX = 1
triploidalna samica
Ryc. 62. Płeć u Drosophila jako stan równowagi heterosomów X i autosomów (X — liczba chromosomów X. A — liczba kompletów autosomów). Zwróć szczególną uwagf na osobniki 2n - 1 = 2A + X0 oraz 2n + l = 2A + XXY.
101
Wyszukiwarka
Podobne podstrony:
fiz teoria 3 yl c Hon /ł/f7V tę’ ^ŁJ) £ /Vi £ 2-t^O *--/ «= 1i2?fW i^^n/rfyZAM
h5 UW? Od)k- j (MJjAf d <M Pechu <&UMKdfy /tW^Ć(hłjJ C ’l,AA .^ / \vJjtX
Image4726 g _ Ą £ = ^2^4 ĄĄ £ Ą + Ą CĄ + Ą) CĄ + Ą)
img680 UiaJS Otft1 Q,u a c
repetytorium (15) Metoda zasięgówMetoda znaków ruchu _) LJ_ <£ CsćL CD CD LJLJ CD Metoda zasięgów
41347 SWScan00045 (2) rt^k /ąaly pnes/nen/ (* n i d) A;W> >o £ /AA ^ $ dęji*
skanowanie0023 2 -CA^ifyycIlA l~ o<Aa. rvap«e^yc^ap^ - k^j nekJC v>r Cjd)^.
mechanika (49) 1 WmmMm Ą Ak^szó-I^ &^Iłł-feoS:-: £■* aA I ( -o(H-* 53hi HUfk^j rT^ M fiu/<w
egzamin 13 f UWDAMf MToWAUiE t Sev. W , 24 .W, 10A3 ( \jfi£j<ł«^lł( 24^żl*i* 1 /ł] Okopie <JlU
ekonomia067 c a 4 ££)3 ?aa - -mi ci 0^ - 4 pi = tfr 3 ! .. d yf J-0 3 A “ 3 GO czO ci I X Pa
więcej podobnych podstron