67858 lastscan13 (5)
0 20 40 60 80 100 120 140
ciśnienie parcjalne tlenu (mm Hg)

ciśnienie parcjalne tlenu (kPa)
0_5_10 15 _20
|
PC |
wietr |
26 \ | |||||
|
\ | |||||||
|
-O- _ |
w |
oda |
\ |
\ | |||
v- }>t • •:
^ Dwutlenek węgla oddychanie w wodzie
Ilość dwutlenku węgla, powstającego podczas procesów metabolicznych jest ści-ślejzwiązana z ilością zużywanego tlenu. Stosunek wymiany oddechowej dwutlenku "węgtąi tlenu, czyli współczynnik wymiany _ oddechowel (nazywany także ilorazem od-dechowym) zwyk\e utrzymuje się w zakresie 0,7 — 1,0 (patrz rozdział 5). Oprowadzonych tu rozważaniach przyjmujemy, że ten współczynnik wymiany wynosi 1,0, a więc na każdą cząsteczkę pobieranego tlenu jest oddawana dokładnie jedna cząsteczka dwutlenku węgla.
U zwierzęcia oddychającego powietrzem, na przykład ssaka, o współczynniku wymiany gazowej równym 1,0, można łatwo przewidzieć stężenie dwutlenku węgla w wydychanym powietrzu. Jeśli zawartość tlenu w wydychanym powietrzu zmniejsza się z 21 do 16%, to silą rzeczy powietrze to zawiera 5% C02. Rozpatrując natomiast zmianę ciśnienia parcjalnego tych dwóch gazów, możemy powiedzieć, że jeśli ciśnienie tlenu w powietrzu oddechowym zmniejszyło się o 50 mm Hg (6,7 kPa), to ciśnienie dwutlenku węgla musiało wzrosnąć o tę samą wartość. Każda zmiana ciśnienia parcjalnego tlenu w pf^leTTZa oddechowym pociąga za sobą-iaką samą, ale przeciwnicTsk ierowa-ną, zmianę, ciśnienia dwutlenku węgla.
Jeśli współczynnik wymiany-równy jedności zastosujemy do analizy wymiany gazowej w wodzie, ilości tlenu i dwutlenku węgla będą oczywiście jednakowe, lecz zmiany ich ciśnienia parcjalnego będą "się różnic ogromnie, ponieważ tak bardzóToz-na jestlćh rozpuszczalność.
~UNa przykład, jeśli ryba zużyje dwie trze-die^tięnu zawartego w wodzie, zmniejszając -sposób jego ciśnienie ze 150 na
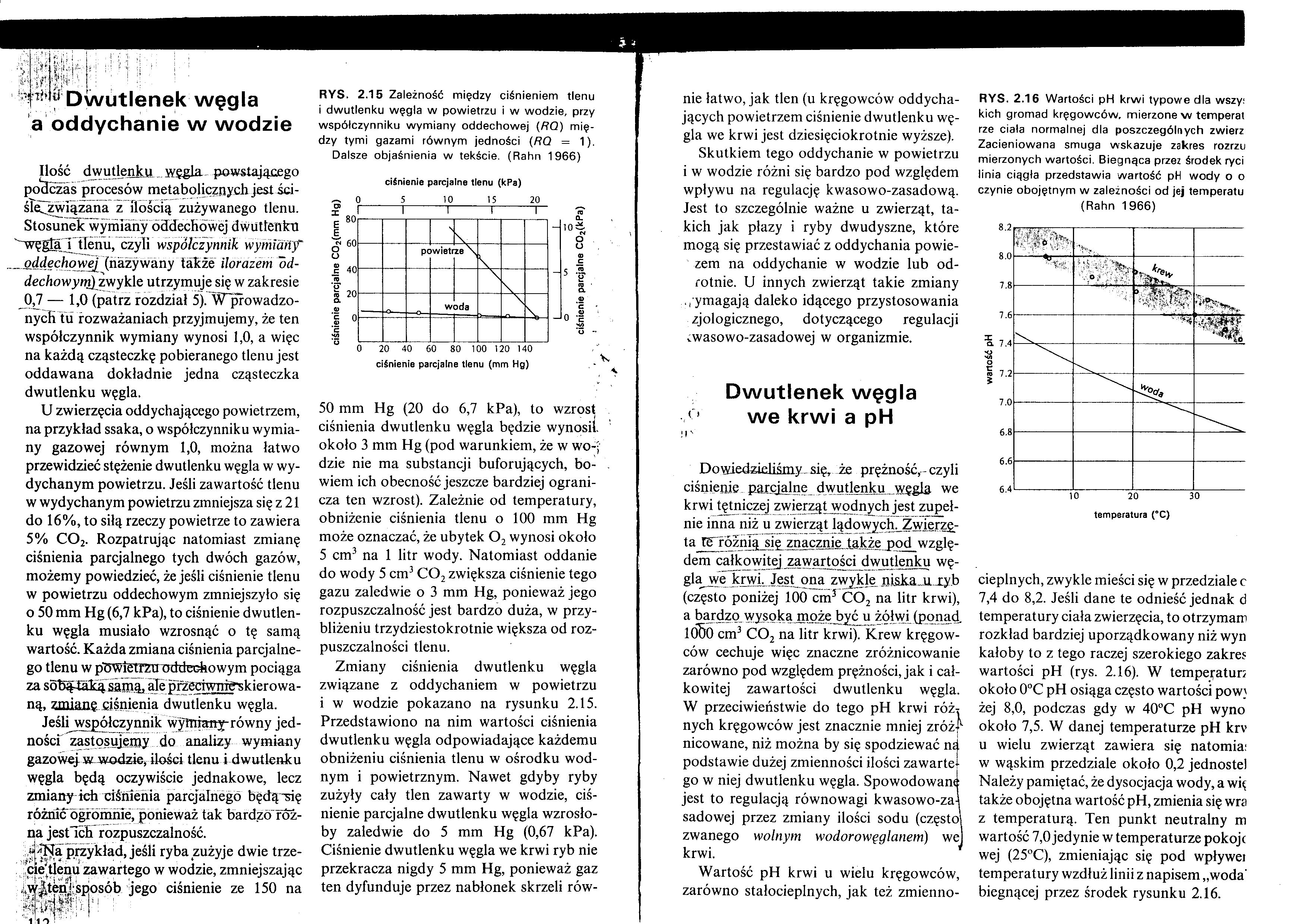
RYS. 2.15 Zależność między ciśnieniem tlenu i dwutlenku węgla w powietrzu i w wodzie, przy współczynniku wymiany oddechowej (RQ) między tymi gazami równym jedności (RQ = 1).
Dalsze objaśnienia w tekście. (Rahn 1966) 50 mm Hg (20 do 6,7 kPa), to wzrost ciśnienia dwutlenku węgla będzie wynosił, około 3 mm Hg (pod warunkiem, że w wo-j dzie nie ma substancji buforujących, bowiem ich obecność jeszcze bardziej ogranicza ten wzrost). Zależnie od temperatury, obniżenie ciśnienia tlenu o 100 mm Hg może oznaczać, że ubytek 02 wynosi około 5 cm3 na 1 litr wody. Natomiast oddanie do wody 5 cm3 C02 zwiększa ciśnienie tego gazu zaledwie o 3 mm Hg, ponieważ jego rozpuszczalność jest bardzo duża, w przybliżeniu trzydziestokrotnie większa od rozpuszczalności tlenu.
Zmiany ciśnienia dwutlenku węgla związane z oddychaniem w powietrzu i w wodzie pokazano na rysunku 2.15. Przedstawiono na nim wartości ciśnienia dwutlenku węgla odpowiadające każdemu obniżeniu ciśnienia tlenu w ośrodku wodnym i powietrznym. Nawet gdyby ryby zużyły cały tlen zawarty w wodzie, ciśnienie parcjalne dwutlenku węgla wzrosłoby zaledwie do 5 mm Hg (0,67 kPa). Ciśnienie dwutlenku węgla we krwi ryb nie przekracza nigdy 5 mm Hg, ponieważ gaz ten dyfunduje przez nabłonek skrzeli równie łatwo, jak tlen (u kręgowców oddychających powietrzem ciśnienie dwutlenku węgla we krwi jest dziesięciokrotnie wyższe).
Skutkiem tego oddychanie w powietrzu i w wodzie różni się bardzo pod względem wpływu na regulację kwasowo-zasadową. Jest to szczególnie ważne u zwierząt, takich jak plaży i ryby dwudyszne, które mogą się przestawiać z oddychania powie-zem na oddychanie w wodzie lub od-rotnie. U innych zwierząt takie zmiany ..ymagają daleko idącego przystosowania zjologicznego, dotyczącego regulacji ewasowo-zasadowej w organizmie.
Dwutlenek węgla (! we krwi a pH
ir
Dowiedzieliśmy^ się, że prężność, - czyli ciśnienie parcjalne dwutlenku węgla we krwi tętniczej zwierząt wodnych jest zupełnie inna niż u zwierząt lądowych. Żwjęrzę_-ta BTróżnia sie znacznie także pod wzglę-dem całkowitej zawartości dwutlenku węgla we krwi. Jest ona zwykle niska u ryb (często poniżej 100 cm3 C02 na litr krwi), a bardzo wysoka może być u żółwi (ponad. 1Ó00 cm3 C02 na litr krwi). Krew kręgowców cechuje więc znaczne zróżnicowanie zarówno pod względem prężności, jak i całkowitej zawartości dwutlenku węgla. W przeciwieństwie do tego pH krwi tóŻt nych kręgowców jest znacznie mniej zróż-*1 nicowane, niż można by się spodziewać nr podstawie dużej zmienności ilości zawarte go w niej dwutlenku węgla. Spowodowani jest to regulacją równowagi kwasowo-za sadowej przez zmiany ilości sodu (częstol zwanego wolnym wodorowęglanem) we] krwi.
Wartość pH krwi u wielu kręgowców, zarówno stałocieplnych, jak też zmienno-
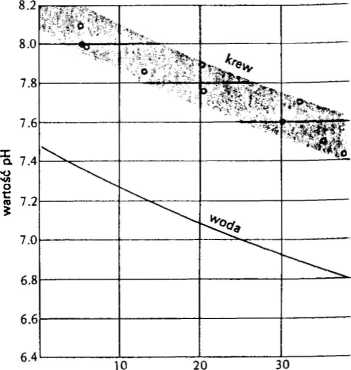
RYS. 2.16 Wartości pH krwi typowe dla wszy! kich gromad kręgowców, mierzone w temperat rze ciała normalnej dla poszczególnych zwierz Zacieniowana smuga wskazuje zakres rozrzu mierzonych wartości. Biegnąca przez środek ryci linia ciągła przedstawia wartość pH wody o o czynie obojętnym w zależności od jej temperatu (Rahn 1966)
temperatura (”C)
cieplnych, zwykle mieści się w przedziale c 7,4 do 8,2. Jeśli dane te odnieść jednak d temperatury ciała zwierzęcia, to otrzymam rozkład bardziej uporządkowany niż wyn kaloby to z tego raczej szerokiego zakres wartości pH (rys. 2.16). W temperatur; około 0°C pH osiąga często wartości pow_' żej 8,0, podczas gdy w 40°C pH wyno około 7,5. W danej temperaturze pH krv u wielu zwierząt zawiera się natomia: w wąskim przedziale około 0,2 jednostel Należy pamiętać, że dysocjacja wody, a wię także obojętna wartość pH, zmienia się wra z temperaturą. Ten punkt neutralny m wartość 7,0 jedynie w temperaturze pokoje wej (25°C), zmieniając się pod wpływei temperatury wzdłuż linii z napisem „woda' biegnącej przez środek rysunku 2.16.
Wyszukiwarka
Podobne podstrony:
image 100 100 Wybrane problemy projektowania anten i szyków antenowych 0 0 20 40 6
IMGD10 20 40 60 80 procentowy stopie
100 200 amin 20 = 300 400 50$ 600 1.03 m/s mDistance (m) O 20 40 60 80 100 120 140 J.60 « Całculated
Databazove systemy 2014 Pocct studcntu 20 40 60 80 100 120 140 160 180 200
A PKM2 high express»on o- PKM2 Iow expression T 0 T" 20 40 60 80 100 4
DSC05810 (2) co daje wartości: c Tablica £ 9 f 20° 40° 60° 80° 90° R
STRUKTURA WYDATKÓW W LATACH 2012 - 2015 2015 2014 2013 2012 0% 20% 40% 60% 80% 100% ■
DSC04183 201) o -20 -40 -60 -80 Sztum Żabno Barie* :e 5 H—~- ~
Wartość Yrms [mm/s] Ioz.6 999 Wartość Yrms [mm/s] loz.5 10 140 160 180 20U 220 Hz I 20 40 60 80 100
O 20 40 60 80 0 20 40 60 80 *ew Sao** Fi f- • M -9- • vy r^ *CQ H *ł w •
acacia21 lilii! 1 I 1 I 1 I 1 I 1 I 1 I 1 I 1 I 1 I 1 iiiiiiiii 0’ 20 40’ 60’ 80’ 100 1 Dm lOm
P1050390 Rozpuszczalność gazów w wodzie II gaz 0° 20° 1_ 40° 60° 80° 100’ 11
00241 tbbf0afd5fc79c2ad0c3ecbce2b5261 Statistical Process Monitoring 243 0 20 40 60 Batch Number 0
200 Wrocławia 09 r (1) jpeg RDZEŃ ŚLĄSKA 20 40 60
Lepkość kinematyczna [mm2/s] 20 40 60 Temperatura
więcej podobnych podstron