17100 skanowanie0006

V 48
Aktywacja (otwieranie) receptorów-kanałó w wapniowych zalezj nych od IP3 zachodzi w wyniku wiązania tego informatora II rzędU? który jest produktem akty wriości fosfólipązy C rozkładającej błonow<jj pochodne fosfatYdyioinozytoly (patrz niźejli;. Warto wspomnieć, żot funkcja tych receptorów jest silnie hamowana przez alkohol etylowyj Właśnie temu można przypisać utratę koordynacji ruchowej w staniej upojenia alkoholowego.
Rola Ca2+ jako informatora II rzędu
Ca2* jest bardzo ważnym informatorem II rzędu, który przy] udziale kalmodułiny (CaM, białka cytosolowego wiążącego Ca2*) ak^ tywuje wiele kinaz i fosfataz serynowó-trępninovyycłi pkreślanyęl| jako kinazy zależne od kompleksu Ća2*/CaM (Ryc. 4); Wzrost stężej nia Ca2* w cytosolu w wyniku pobudzenia' i otwarcia receptorów! kanałów wapniowych mą więc ogromne znaczenie dla dalszego przef kazywania sygnałów oraz indukcji ostatecznego efektu fizjologiczl nago. 1 . J V
Przykładem roli Ca2 * jako informatora Ił rzędu w aktywacjij procesów komÓrkPwych może być np. jego udział w skurczu mięśnia gładkiego, gdzie zależna od Ca2 * /CaM kinaza f osf oryluje łańcuchy] lekkie miozyny. Innym# fcupełnię odmiennym przykładem jest aktyi wacja limfocytów T zależna! od receptora d|0 antygenu. W tymi przypadku, kompleks Ca**/ĆaM aktywuje fosfatazę sęrynową PP2B; (kalcyneurynę), która defosforylując/czynnik transkrypcyjny NF-ATi umożliwia mu przejście do jądra komórkowego, przyłączenie się do swoistej regulatorowej sekwencji DNA W genie dla interleukiny-2 i w| konsekwencji stymulaćj^ ekspresji tej cytokiny, odgry wającej kluczo- j wą rolę w Odpowiedzi immunologicznej;
Ca2* odgrywa te£ istotną rolę. w aktywacji innej kinazy serynowej, zależnej od diiacylpgtićerblu (DG) kinazy C, aktywuje niektóre ATPazy, bezpośrednio łub pośrednio stymuluje DNazy odpowiedzialne za fragmentatję DNA w przebiegu apoptozy oraz ułatwia fuzję błon komórkowych, np. w przebiegu egzocytozy.
Receptory związane z aktywacją białek G
Przekazywanie informacji przy udziale tych receptorów zależy od wiązania, zmiany konformacji i w rezultacie aktywacji swoistych białek G {cytosolowych, białek wiążących i hydrolizujących GTP).. Pełnią one rolę w transdijkcji sygnału przez błonę aktywując swoisty enzym wzmacniacz. Re&eptory tej grupy mają charakterystyczną
budowę. W ich cząsteczce występuje 7 powtarzających się sekwencji hydrofobowych o budowie {/-helisy, które przechodzą przez błonę tworząc charakterystyczne pętle (Ryc. 5). Poprzyłączeniu ligandu w ” kieszorigg” utworzonej prze2 pętle pozakomórkowe następuje zmiana konformacji receptora, w wyniku której druga i trzecią pętla cytoplazmatyczha wiążą białka q. Związanie białka G prowadzi do jogo aktywacji. Heterogennośę receptorów tej grupy jest związana r aktywacją różnych białek G, a co za tym idzie, różnych enzymów wzmacniaczy i różnych informatorów 11 rzędu.
Mlefsc% wiązania Hgandu
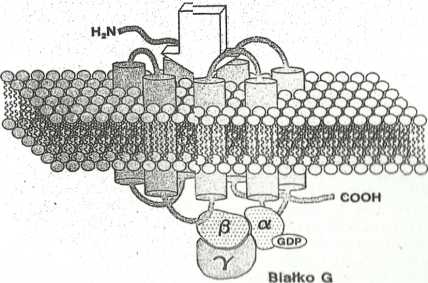
Ryc. B. Schomot budowy roooptor^w zwląitnyoh i iktywao|q blalok 0,
Blotka G aą hotarotrlmara»ni utworionyml pri»/ podjadnoitkl a, 0 I y. Zdolność do wiązania Qtp I Ingo hydroliiy do OOP poiladn podjadnoatka a, Po pnyląo/oriu białka Q do aklywowanago laoap
Wyszukiwarka
Podobne podstrony:
skanowanie0006 V 48 Aktywacja (otwieranie) receptorów-kanałó w wapniowych zalezj nych od IP3 zachodz
skanowanie0022 (48) Kamizelkaz szalowym kołnierzemRozmiary: 40/42 (46/48) Materiał: 750 (900) g brąz
skanowanie0029 (48) Tabela 15. .-śsj Zapotrzebowanie na warzywa i owoce (w porcjac
skanowanie1 (6) 48. Moczowód u mężczyzn uchodzi do: a. cewki moc
phoca thumb l slajd17 (18) Różne typy bodźców wpływają na otwieranie i zamykanie kanałów jonowychKan
skanowanie0025 (48) ■fejl- <20)1 ■ 4 1/2 59 tyłu i lewy przód IMI (29)
skanowanie0014 (48)
skanowanie0015 (48) Biurko pod komputerBiurko powinno: > Być stabilne, >
więcej podobnych podstron