31400 justy005

źródeł różni się rodzajem rozgałęzień, liczbą jednostek w łańcuchu oraz właściwościami.
Skrobię można przeprowadzić do glukozy w drodze kwaśnej hydrolizy lub enzymatycznie. Występują trzy rodzaje enzymatycznej degradacji glukanów: 1) fosforoliza, 2) hydroliza, 3) transglikozylacja.
Fosforoliza. Przekształcenie skrobi, glikogenu i podobnych wielocukrów do glukozo-1-fosforanu jest katalizowane przez a-1,4-glukanofosforylazę (fosforylazy). Chociaż reakcja jest odwracalna, przebiega ona wewnątrzkomórkowe jedynie w katabolizmie wielocukrów i nie ma znaczenia dla syntez. Fosforylacja rozpoczyna się od wolnego nieredukującego końca łańcucha amylozy, z uwolnieniem kolejnych reszt glukozo-1-fosforanu. W przypadku amylopektyny fosforoliza zatrzymuje się przy rozgałęzieniach w pozycji 1,6 i postępuje dalej dopiero po rozszczepieniu tych ostatnich przez amylo-l,6,-glukozydazę. Fosforylazy odgrywją ważną rolę w aktywowaniu i wykorzystywaniu wewnątrzkomórkowych wielocukrów zapasowych (glukanów).
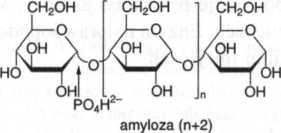
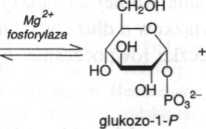
amyloza (n + 1)
Hydroliza. Wielocukry wewnątrz komórki są rozszczepianie hydrolitycznie przez amylazy. a-Amylaza występuje u roślin, zwierząt i drobnoustrojów. Szybko upłynnia ona skrobię, równocześnie atakując wiele wiązań 1,4-glikozydowych, łącznie z wiązaniami w środku łańcucha (jest zatem nazywana również endoamylazą). Uwalnia ona maltozę, glukozę oraz oligomery zawierające od 3 do 7 reszt glukozy. Z powodu szybkiej degradacji struktury makrocząsteczkowej lepkość roztworu oraz jego zdolność wchodzenia w reakcję z jodyną również szybko maleją, z równoczesnym stopniowym pojawieniem się cukrów ulegających fermentacji (glukoza, maltoza, maltotrioza). W obecności enzymów odcinających rozgałęzienia, jak pullulanaza lub izoamylaza, zachodzi również degradacja dekstryn (ryc. 14.3).
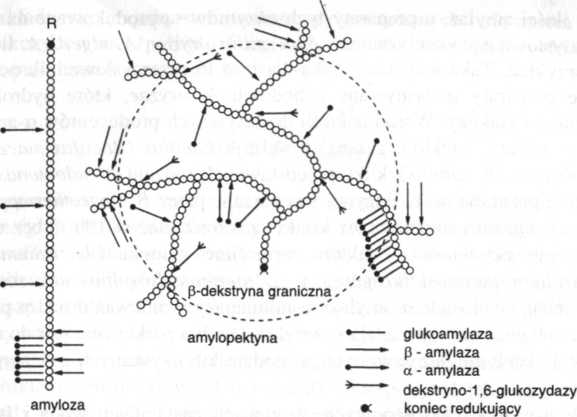
Ryc. 14.3. Miejsce działania enzymów na amylozę i amylopektynę
/3-amylazy występują przede wszystkim w roślinach (np. w jęczmieniu, pszenicy), lecz również w bakteriach. W przeciwieństwie do a-amylazy, przejawiają aktywność od wolnych nieredukujących końców makrocząsteczki. Ich działanie na skrobię prowadzi do szybkiej akumulacji cukrów z zachowaniem zdolności reagowania z jodem. Hydroliza zatrzymuje się w miejscach rozgałęzień, a pozostające reszty noszą nazwę granicznej /i-dekstryny. Gdy miejsca rozgałęzień ulegną enzymatycznemu rozszczepieniu, następuje całkowite przekształcenia substratu do maltozy, która z kolei na zewnątrz komórki może być rozkładana przez maltazę. W obecności odpowiednich permeaz maltoza i inne oligomery są transportowane do wnętrza komórki i tam rozszczepiane fosfory li tycznie.
Transglikozylacja. Schardinger odkrył w podłożach zawierających skrobię, w których hodowano Bacillus macerans, pewne związki o charakterze krystalicznym. Składają się one z pierścieniowo zamkniętych łańcuchów reszt glukozy połączonych wiązaniami a-l,4-glikozydowymi. Pierścienie tych a-, P- lub y-cyklodekstryn składają się z sześciu, siedmiu lub ośmiu reszt glukozy i powstają ze skrobi w wyniku działania transglikozylaz.
Zarówno grzyby, jak i bakterie wytwarzają a-amylazy. Zdolność rozkładania skrobi przez zewnątrzkomórkowe enzymy amylolityczne jest wśród drobnoustrojów szeroko rozpowszechniona; trudno mówić o mikroflorze swoiście degradującej skrobię. Wiele grzybów glebowych wytwarza
511
Wyszukiwarka
Podobne podstrony:
justy001 rozkładu jest znacznie bardziej rozgałęziony niż w beztlenowym łańcuchu pokarmowym. Z tego
wywiad; wyjaśnia czym rożni się plan szczegółowy od planu ramowego oraz redaguje szczegółowy plan ut
e2 MIĘSIEŃ SERCOWY Mięsień sercowy jest rodzajem mięśni poprzecznie prążkowanych, lecz różni się
metody 2 F 1. Co to jest kojarzenie i jakie znasz rodzaje kojarzeń 2. Czym różni s
skanuj0012 (235) MISTRZ powstawszy wznosząc z tej pożoi że sit tak jak się grozi jedyna Liczba która
skanuj0025 (229) i ju egzaminu. Końcowy test różni się w zależności od tradycji. Czasami szaman — na
farma zwierzeta Twoje imię:_ Pokoloruj obrazek,, któiy różni sie od innych.
File0055 Znajdź w każdym rzędzie jeden element, który różni się od pozostałych.
Image102 Bramka z otwartym obwodem kolektora Bramka z otwartym obwodem kolektora różni się od typowy
więcej podobnych podstron