1
Cykl kwasu cytrynowego
Cykl kwasu cytrynowego (cykl Krebsa) jest głównym ośrodkiem
metabolicznym komórki. Wprowadza do metabolizmu tlenowego
wszystkie cząsteczki, które mogą zostać przekształcone
w grupy acetylowe albo w kwasy dikarboksylowe. Cykl ten nie
tylko dostarcza komórce cząsteczek będących zapasową formą
paliwa komórkowego, ale stanowi źródło elementów budulcowych
wielu cząsteczek, takich jak aminokwasy, zasady nukleotydowe,
cholesterol i porfiryna.
Cykl kwasu cytrynowego obejmuje szereg reakcji utleniania
i redukcji, w wyniku których grupa acetylowa zostaje utleniona
do dwóch cząsteczek dwutlenku węgla.
Cykl kwasu cytrynowego jest końcowym, wspólnym szlakiem
utleniania substratów energetycznych: aminokwasów,
kwasów tłuszczowych i węglowodanów.
Większość tego paliwa wchodzi do cyklu w postaci
acetylokoenzymu A
.
aminokwasy
kwasy
tłuszczowe
glukoza
pirogronian
szczawiooctan
kwas cytrynowy
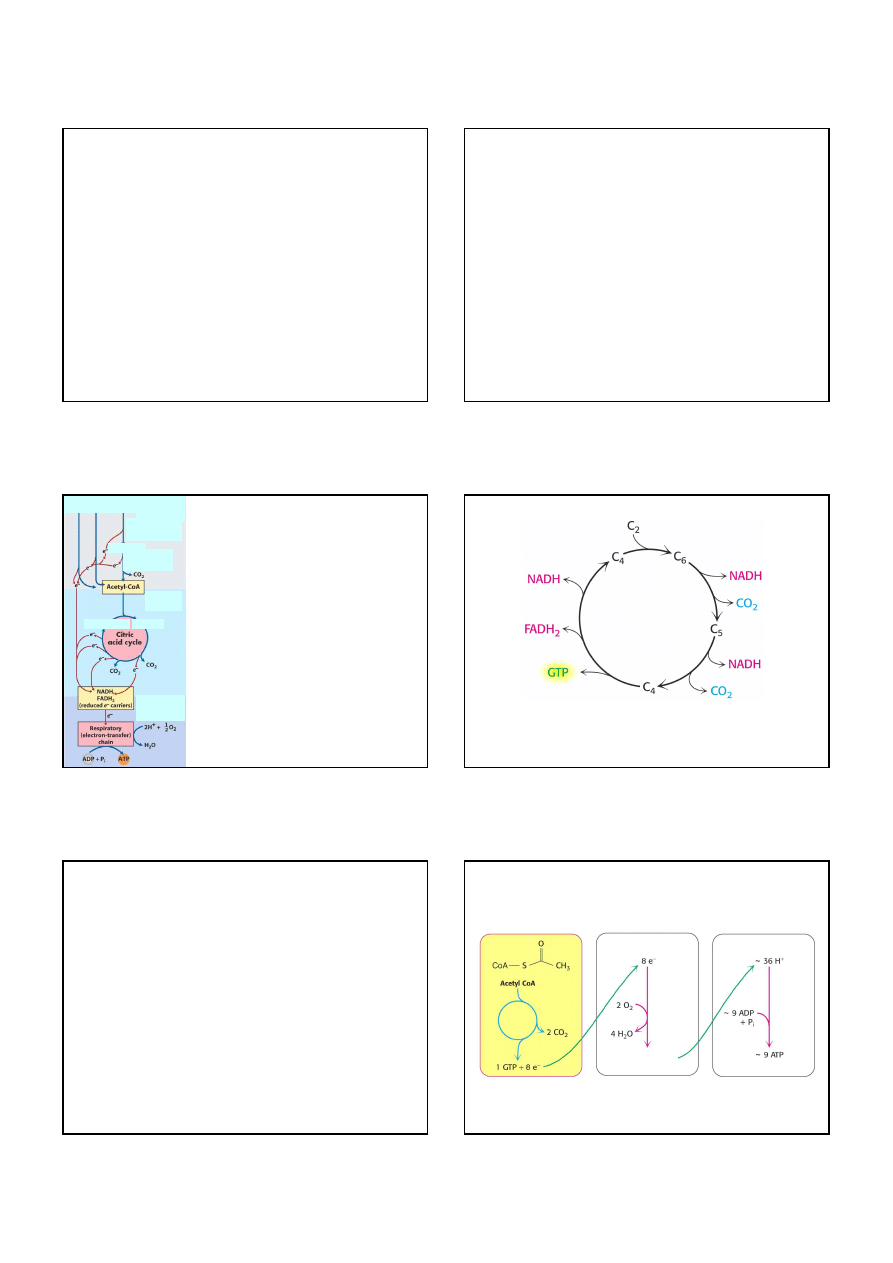
Etap 1:
wytworzenia
acetylo-CoA
Etap 2:
utlenienie
acetylo-CoA
Etap 3:
przeniesienie
elektronów
i fosforylacja
oksydacyjna
dehydrogenaza
pirogronianowa
glikoliza
Katabolizm białek, kwasów tłuszczowych
i węglowodanów w trzech etapach
oddychania komórkowego
Ogólny schemat cyklu kwasu cytrynowego
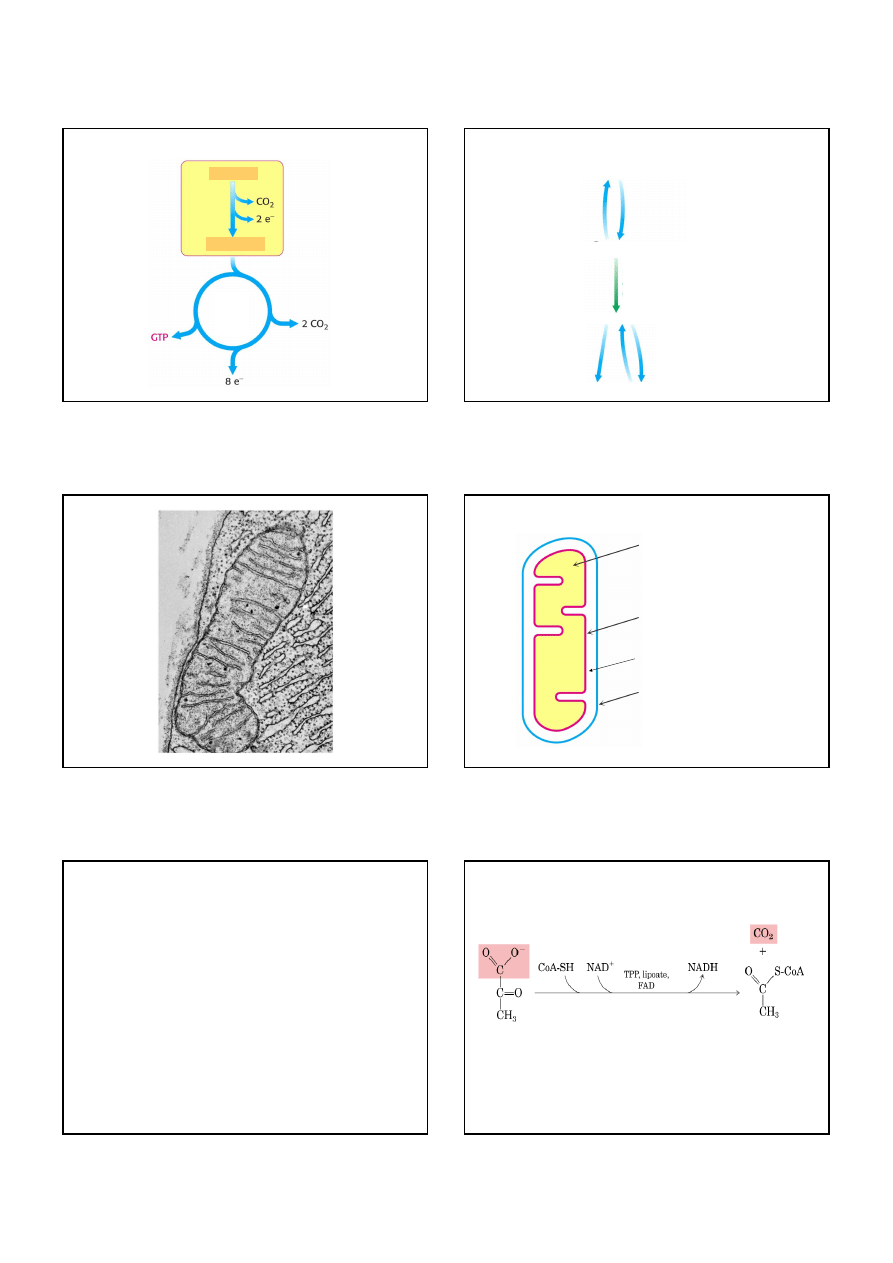
Cykl kwasu cytrynowego utleniając jednostki dwuwęglowe
wytwarza 2 cząsteczki CO
2
, 1 cząsteczkę GTP i wysokoenergetyczne
elektrony w postaci NADH i FADH
2
.
(zredukowany NADH)
Cykl kwasu cytrynowego nie wytwarza dużej ilości ATP, natomiast
usuwa elektrony z acetylo-CoA i wykorzystuje je do tworzenia
NADH i FADH
2
.
Te elektrony są następnie uwalniane w czasie utleniania NADH
i FADH
2
w procesie
fosforylacji oksydacyjnej
,
i przepływają przez szereg białek błonowych
(
łańcuch transportu elektronów
), dzięki czemu w poprzek
błony tworzy się gradient protonowy.
Przepływ protonów przez ATP-azę typu F generuje ATP,
który jest głównym źródłem energii użytecznej.
cykl kwasu
cytrynowego
fosforylacja oksydacyjna
łańcuch
transportu
elektronów
syntaza
ATP
gradient
protonowy
(ok. 36 H
+
)
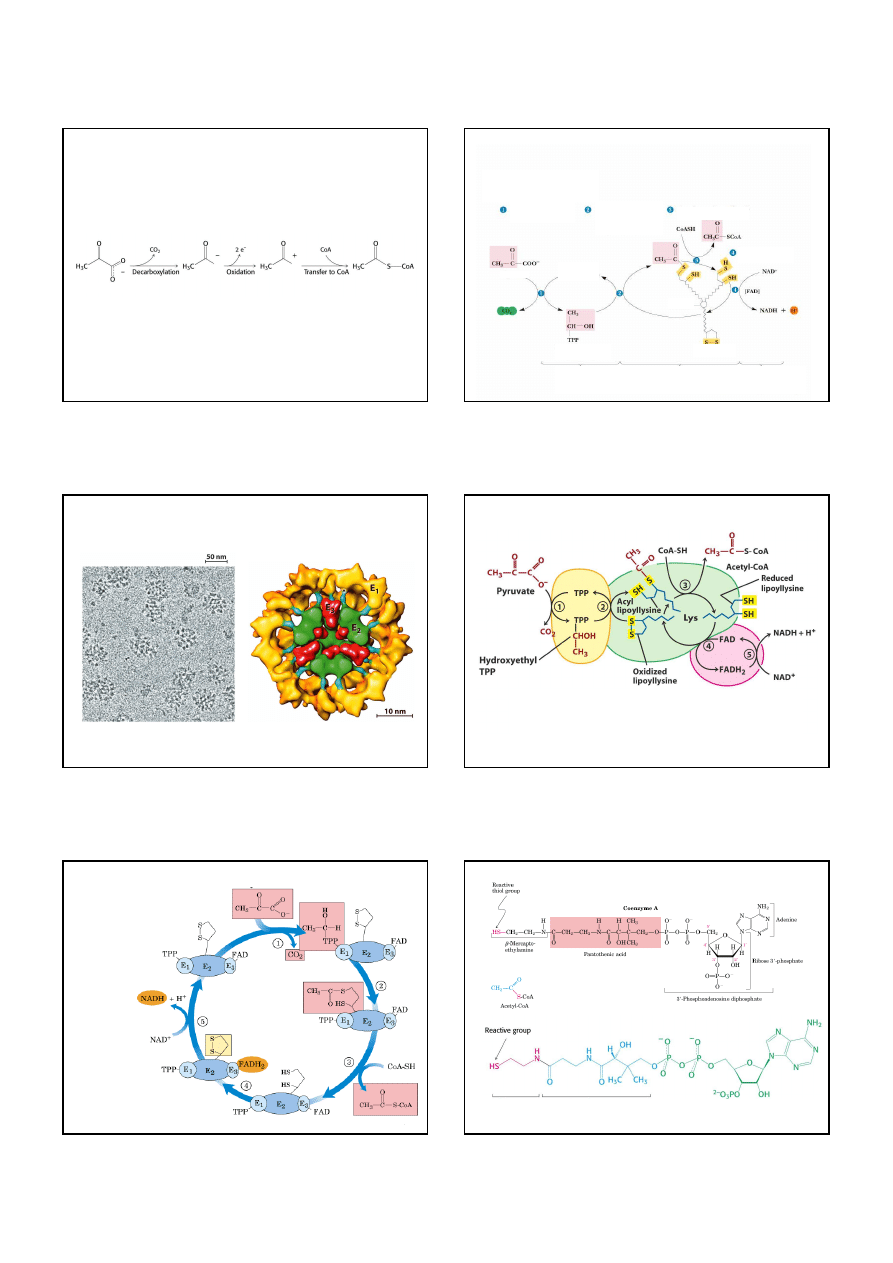
Cykl kwasu cytrynowego jest pierwszym etapem oddychania komórkowego
W cyklu kwasu cytrynowego wysokoenergetyczne elektrony są odbierane z paliwa
komórkowego. Te elektrony redukują O
2
, dzięki czemu powstaje gradient protonowy,
który jest wykorzystywany do syntezy ATP w procesie
fosforylacji oksydacyjnej.
2
Połączenie glikolizy z cyklem kwasu cytrynowego
pirogronian
acetylo-CoA
cykl
kwasu
cytrynowego
acetylo-CoA
glukoza
pirogronian
Kompleks
dehydrogenazy
pirogronianowej
CO
2
lipidy
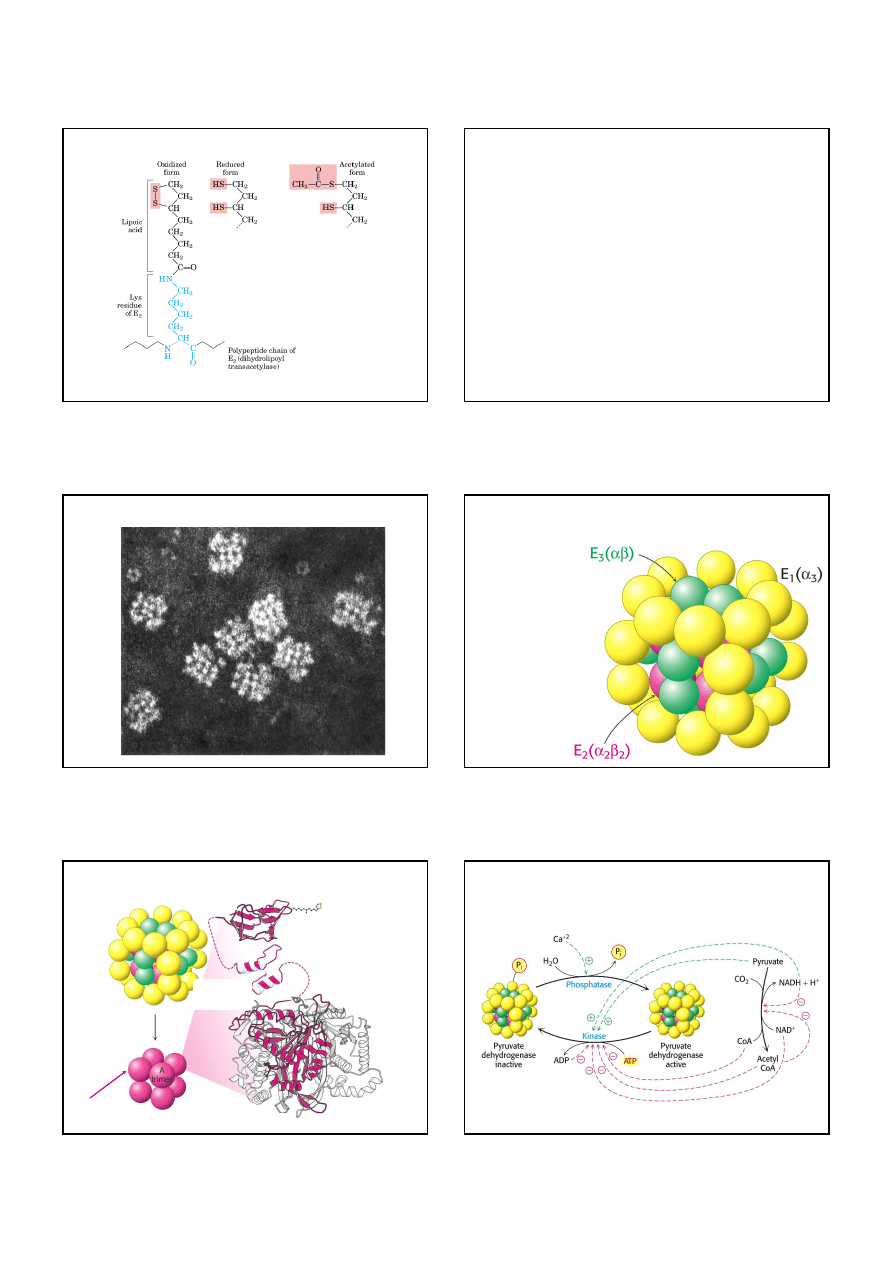
Tworzenie acetylo -CoA z pirogronianu jest kluczową reakcją
nieodwracalną
SCHEMAT MITOCHONDRIUM
Matriks
dekarboksylacja oksydacyjna
pirogronianu i kolejne reakcje cyklu
kwasu cytrynowego zachodzą w
matriks mitochondrialnej
wewnętrzna błona mitochondrialna
zewnętrzna błona mitochondrialna
przestrzeń międzybłonowa
tu gromadzą się jony H
+
,
pompowane z matrix
Pirogronian powstaje w wyniku glikolizy w cytoplazmie komórki.
Aby wejść do cyklu kwasu cytrynowego,
pirogronian musi być przeniesiony do matriks mitochondrium.
Dokonuje tego białko przenoszące kwasy monokarboksylowe
(MCT, monocarboxylic acid transporter), scharakteryzowane
po raz pierwszy w roku 2003.
REAKCJA KATALIZOWANA PRZEZ KOMPLEKS DEHYDROGENAZY
PIROGRONIANOWEJ
tlenowa dekarboksylacja pirogronianu
pirogronian
acetylokoenzym A
(acetylo-CoA)
kompleks
dehydrogenazy pirogronianowej
(E1+E2+E3)
3
pirogronian
acetylo-CoA
1. Dekarboksylacja 2. Utlenianie 3. Przeniesienie do CoA
Przekształcenie pirogronianu w acetylo-CoA zachodzi w 3 etapach
dehydrogenaza
pirogronianowa
acetylotransferaza
dihydrolipoanionowa
dehydrogenaza
dihydrolipoanionowa
Pirogronian
traci CO
2
i powstaje
hydroksyetylo-TPP
hydroksyetylo-TPP
(TPP: pirofosforan
tiaminy)
Grupa hydroksyetylowa
zostaje przeniesiona
na kwas liponowy
i utleniona
do acetylolipoamidu
Przeniesienie
grupy acetylowej
na CoA
Reoksydacja
dihydrolipoamidu
kwas liponowy
Reakcje zachodzące w czasie przekształcenia pirogronianu w acetylo-CoA
pirogronian
pirofosfataza
tiaminowa
Kompleks dehydrogenazy pirogronianowej składa się z 3 enzymów
Trzy enzymy wchodzące w skład
kompleksu dehydrogenazy pirogronianowej
dehydrogenaza
pirogronianowa
E
1
transacetylaza
dihydroliponianowa
E
2
dehydrogenaza
dihydroliponianowa
E
3
TPP: pirosforan tiaminy (witamina B
1
)
Tlenowa dekarbosylacja pirogronianu
pirogronian
pirogronian
acetylo-CoA
utleniona
lipolizyna
utleniona
lipolizyna
zredukowana
lipolizyna
E1 dehydrogenaza
pirogronianowa
E2 acetylotransferaza
dihydroliponioanowa
E3 dehydrogenaza
dihyroliponianowa
Reszta
ββββ
-merkaptoetyloaminy
Reszta kwasu
pantotenowego
Acetylo-CoA
4
KOMPLEKS DEHYDROGENAZY PIROGRONIANOWEJ:
kompleks 3 enzymów ściśle współpracujących z sobą
Enzym
składnik o aktywności
dehydrogenazy
pirogronianowej
acetylotransferaza
dihydroliponioanowa
dehydrogenaza
dihyroliponianowa
Symbol
E1
E2
E3
Liczba
łańcuchów
24
24
12
Grupa
prostetyczna
TPP (B1)
lipoamid
FAD
Katalizowana
reakcja
oksydacyjna
dekarboksylacja
pirogronianu
przeniesienie grupy
acetylowej na CoA
regeneracja
utlenionej formy
lipoamidu
Mikrografia elektronowa kompleksu dehydrogenazy pirogronianowej
Model kompleksu dehydrogenazy pirogronianowej
E1: składnik
dehydrogenazy
pirogronianowej
E2: acetylotransferaza
dihydroliponianowa
E3: dehydrogenaza
dihydroliponianowa
Struktura rdzenia tworzonego
przez acetylotransferazę dihydrolipoanionową (E
2
)
trimer
złożony
z 3 podjednostek E
2
lipoamid
domena
lipoamidowa
domena
oddziałująca
z komponentem E
3
domena
acetylotransferazowa
Aktywność kompleksu dehydrogenazy pirogronianowej
jest regulowana przez
odwracalną fosforylację
5
szczawiooctan + acetylo-CoA
→
→
→
→
cytrynylo-CoA
→
→
→
→
cytrynian + CoA
1 etap cyklu kwasu cytrynowego:
syntaza cytrynianiowa
tworzy cytrynian ze szczwiooctanu i acetylo-CoA
Syntaza
cytrynianowa
kwas szczawiowy
1 etap cyklu kwasu cytrynowego:
syntaza cytrynianiowa
tworzy cytrynian ze szczwiooctanu i acetylo-CoA
Syntaza cytrynianowa zmienia konformację podczas wiązania
szczawiooctanu
2. Izomeracja cytrynianu do izocytrynianu przez akonitazę
(
akonitaza)
Produktem pośrednim jest cis-akonitan
cytrynian
cis-akonitan
izocytrynian
2. Izomeracja cytrynianu do izocytrynianu katalizowna przez
akonitazę
Akonitaza
jest białkiem żelazo-siarkowym:
centrum aktywne zawiera 4 atomy żelaza, niewbudowane w
grupę hemową, które tworzą kompleks z 4 atomami siarki
nieorganicznej i 3 atomami siarki cyteiny.
6
3.
Dehydrogenaza izocytrynianowa
katalizuje utlenianie i
dekarboksylację izocytrynianu do
α
αα
α
-ketoglutaranu
izocytrynian
szczawiobursztynian
α
αα
α
-ketoglutaran
Dehydrogenaza
izocytrynianowa
Produktem pośrednim jest szczawiobursztynian (niestabilny
ββββ
-ketokwas).
W reakcji powstaje dwutlenek węgla.
kwas glutarowy
3. Utlenianie i dekarboksylacja
izocytrynianu do
α
αα
α
-ketoglutaranu
Produktem pośrednim jest szczawiobursztynian (niestabilny
ββββ
-ketokwas).
W reakcji powstaje dwutlenek węgla.
α
αα
α
-ketoglutaran
bursztynylo-CoA
4. Oksydacyjna dekarboksylacja
α
αα
α
-ketoglutaranu prowadzi do
powstania bursztynylokoenzymu A
Dehydrogenaza
α
αα
α
-ketoglutaranowa
W reakcji powstaje dwutlenek węgla i NADH.
kwas bursztynowy, krótszy o 1 C od kwasu
glutarowego
4.
Dehydrogenaza
α
αα
α
-ketogluranowa
katalizuje oksydacyjną
dekarboksylację
α
αα
α
-ketoglutaranu, w wyniku czego powstaje
bursztynylokoenzym A
bursztynylo-CoA
bursztynian
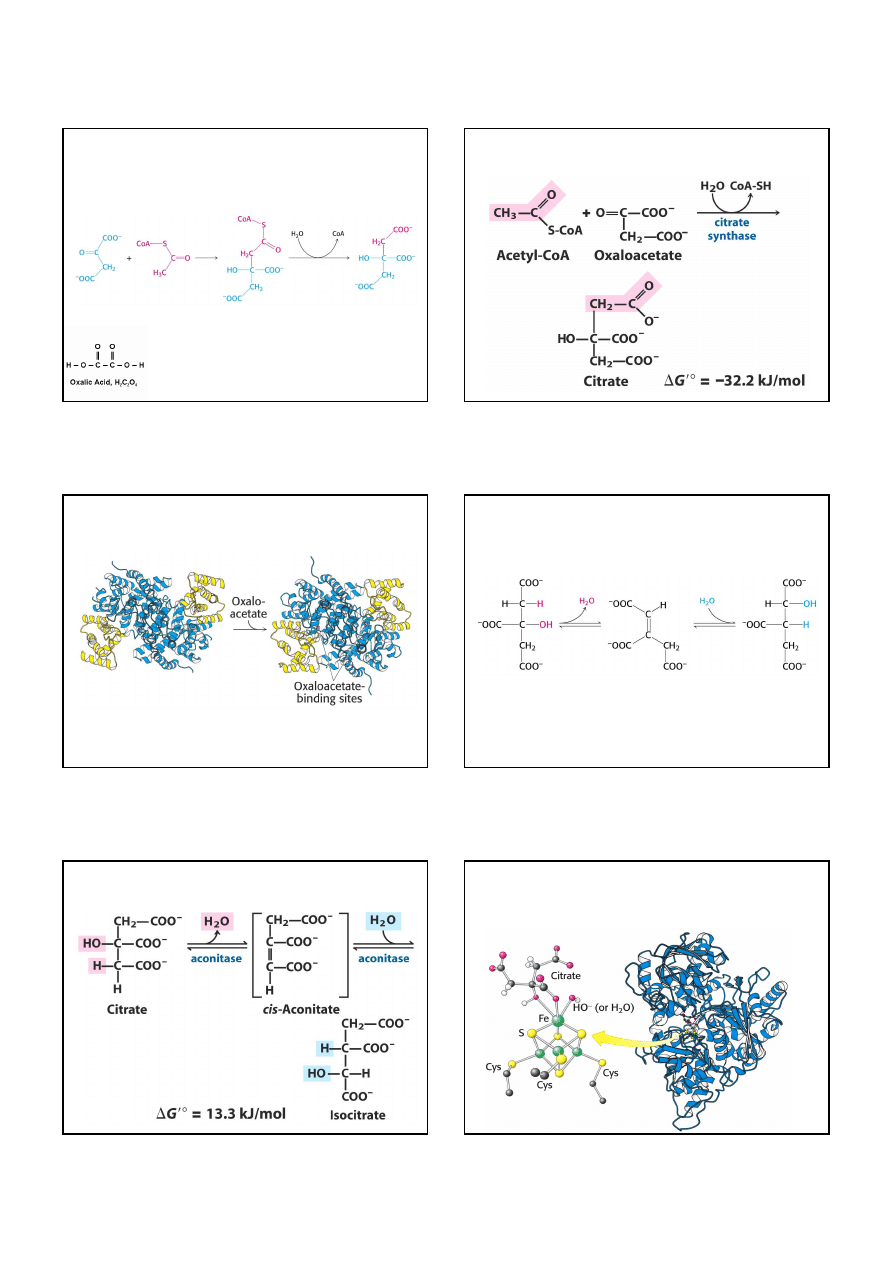
5. Kosztem bursztynylo-CoA powstaje wysokoenergetyczne
wiązanie fosforanowe (fosforylacja substratowa)
Syntetaza
bursztynylo-CoA
Rozerwanie wiązania tioestrowego w bursztynylo-CoA jest sprzężone
z fosforylacją GDP. Grupa fosforanowa GTP może być łatwo
przeniesiona ma ADP, tworząc ATP, w reakcji katalizowanej
przez kinazę nukleozydodifosforanową.
5.
Syntetaza bursztynylo-CoA
przekształca bursztynylo-CoA w
bursztynian z wytworzeniem wysokoenergetycznego wiązania
fosforanowego (fosforylacja substratowa).
7
Mechanizm reakcji katalizowanej przez syntetazę bursztynylo-CoA:
reszta histydynowa enzymu przyłącza grupę fosforanową, dzięki
czemu powstaje bursztynian. Fosforan zostaje przeniesiony na GDP
z wytworzeniem GTP.
bursztynylo-CoA
bursztynian
bursztynian
fumaran
jabłczan
szczawiooctan
6, 7, 8. Szczawiooctan jest regenerowany przez utlenianie
bursztynianu
6. Dehydrogenaza
bursztynianowa
7. Fumaraza
8. Dehydrogenaza
jabłczanowa
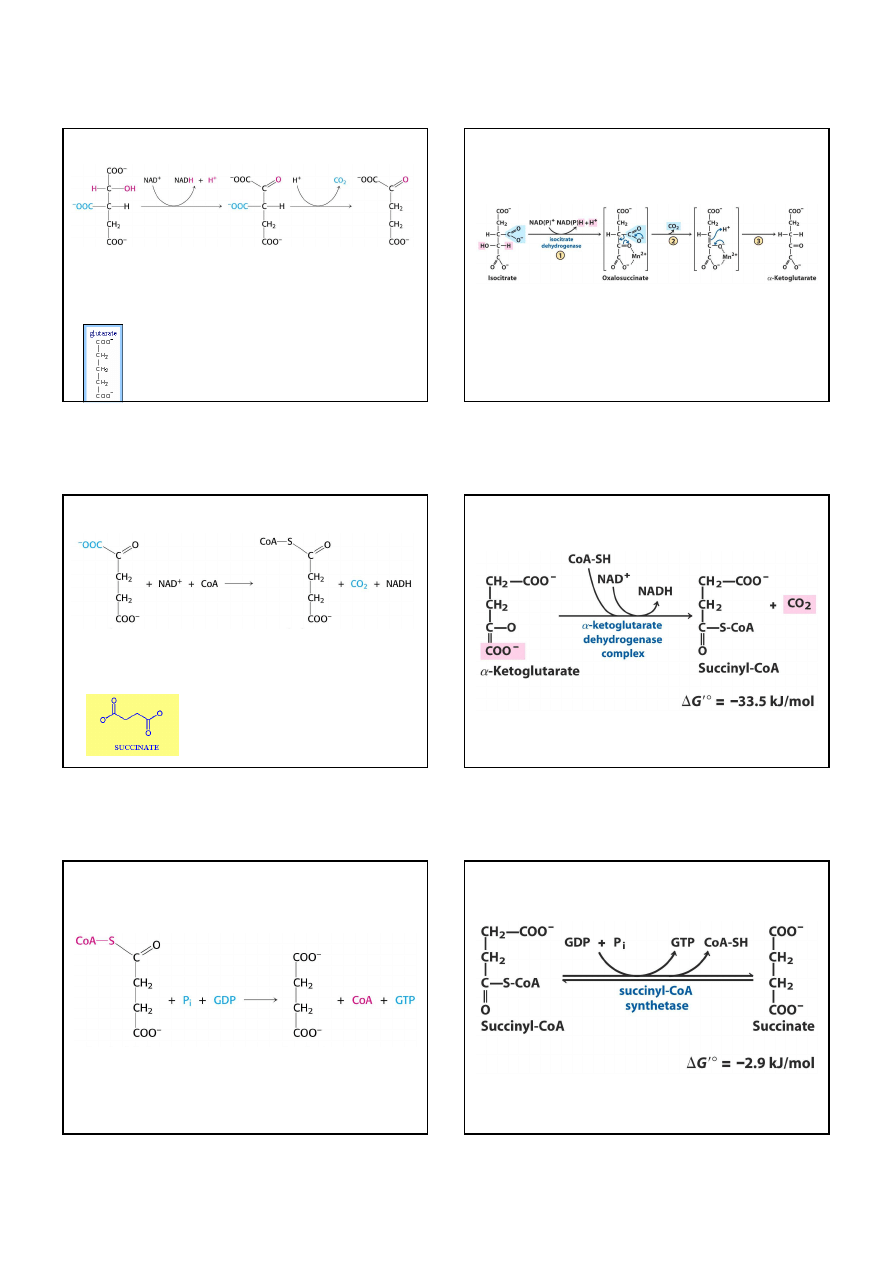
Utlenianie bursztynianiu do fumaranu jest katalizowane przez
dehydrogenazę bursztynianową
. Akceptorem wodoru jest FAD,
ponieważ zmiana energii swobodnej tej reakcji jest niewystarczająca
do zredukowania NAD
+
. W procesach utleniania, w których z substratu
usuwane są 2 atomy wodoru, prawie zawsze akceptorem jest FAD.
Dehydrogenaza bursztynianowa
jest białkiem żelazowo-siarkowym,
i różni się od innych enzymów cyklu kwasu cytrynowego tym, że jest
integralną częścią wewnętrznej błony mitochondrialnej
,
i stanowi pomost między cyklem kwasu cytrynowego i syntezą ATP.
Końcowym akceptorem elektronów z FADH
2
jest tlen cząsteczkowy.
6.
Dehydrogenaza bursztynianiowa
utlenia bursztynian
do fumaranu. Powstaje FADH
2
.
Dehydrogenaza bursztynianowa
jest białkiem żelazowo-siarkowym,
i różni się od innych enzymów cyklu kwasu cytrynowego tym, że jest
integralną częścią wewnętrznej błony mitochondrialnej
,
i stanowi pomost między cyklem kwasu cytrynowego i syntezą ATP.
Końcowym akceptorem elektornów z FADH
2
jest tlen cząsteczkowy.
7.
Fumaraza
katalizuje uwodnienie fumaranu do jabłczanu.
Reakcja jest stereospecyficzna: tylko izomer trans fumaranu
zostaje uwodniony do L-jabłczanu.
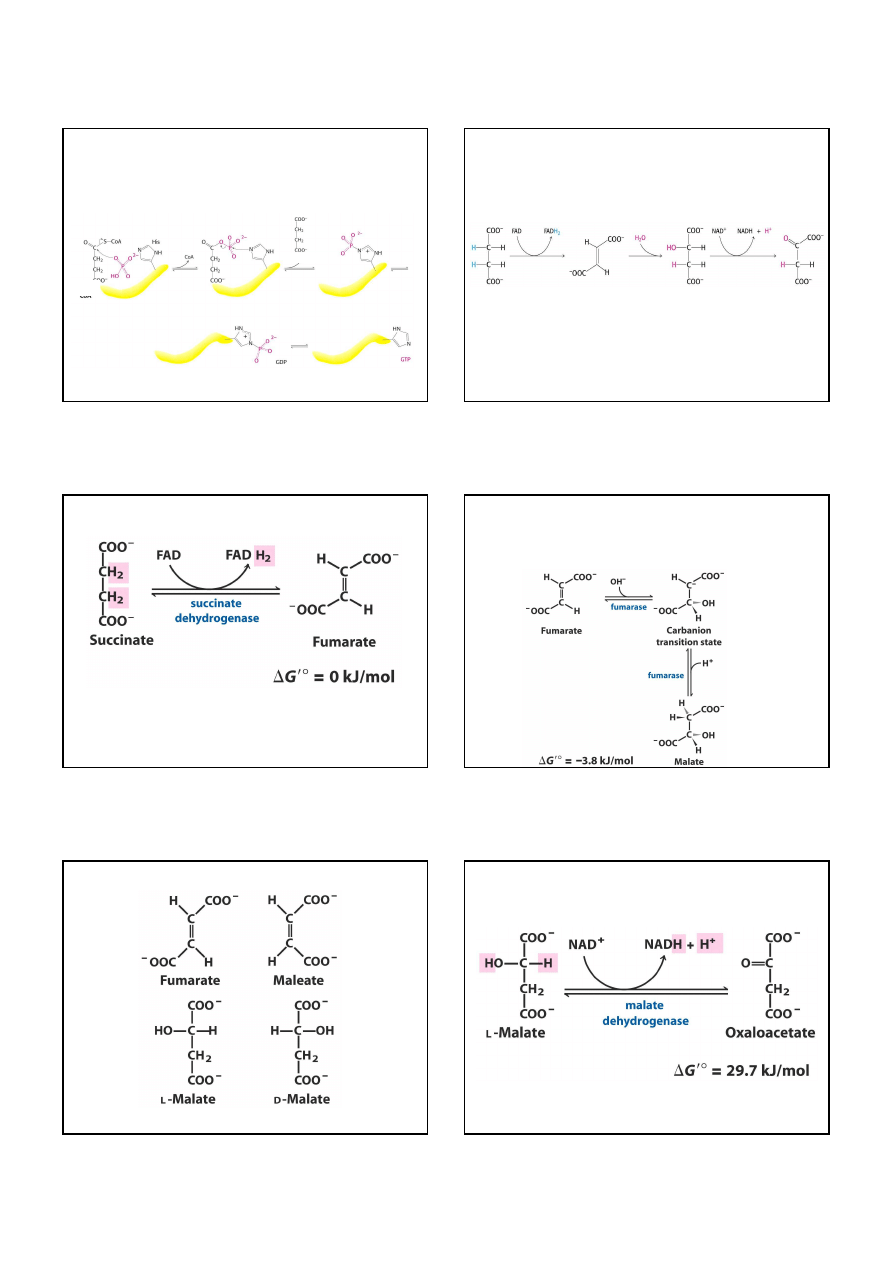
fumaran (izomer trans) maleinian (izomer cis)
L-jabłczan D-jabłczan
8.
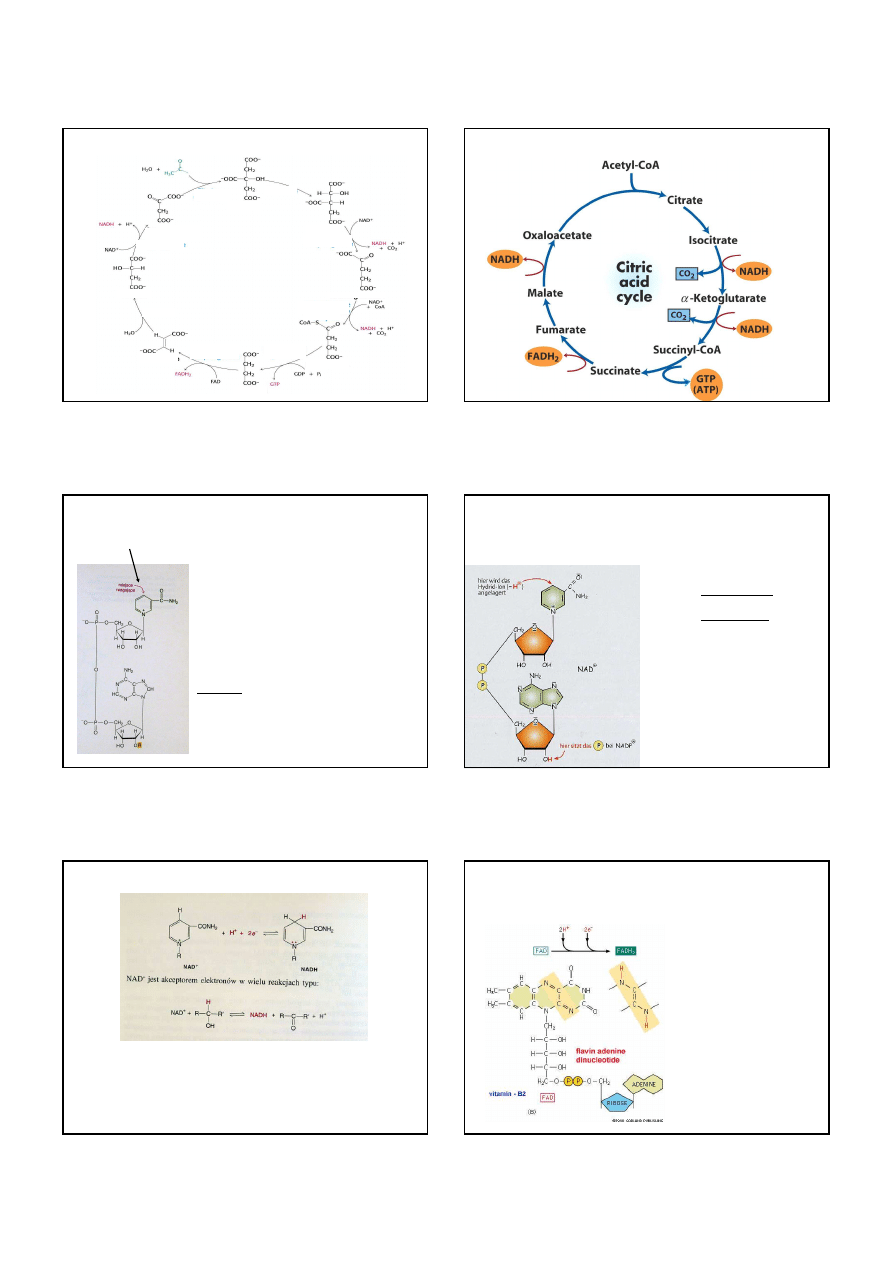
Dehydrogenaza jabłczanowa
utlenia jabłczan do szczawiooctanu
Szczawiooctan jest gotowy do przyłączenia reszty acetylowej
w ramach 1 etapu cyklu kwasu cytrynowego.
8
Cykl kwasu cytrynowego
cytrynian
izocytrynian
α
αα
α
-ketoglutaran
bursztynylo-CoA
bursztynian
fumaran
jabłczan
szczawiooctan
acetylo-CoA
1. syntaza
cytrynianowa
2. Akonitaza
3. Dehydrogenaza
izocytrynianowa
4. Dehydrogenaza
α
αα
α
-ketoglutaranowa
5. Syntetaza
bursztynylo-CoA
6. Dehydrogenaza
bursztynianowa
7. Fumaraza
8. Dehydrogenaza
jabłczanowa
Produkty cyklu kwasu cytrynowego: 3 NADH, 1 FADH
2
, 1 GTP, 2 CO
2
Powstają 3 cząsteczki
zredukowanej
formy
NAD (NADH)
i jedna cząsteczka
FADH
2
NADH i FADH
2
są głównymi przenośnikami elektronów w
procesie utleniania „paliwa molekularnego”
Struktura utlenionej formy dinukleotydu
nikotynamidoadeninowego (NAD
+
) i
fosforanu dinukleotydu
nikotynamidoadeninowego (NADP
+
).
NAD
+
:
R
= H
NADP
+
:
R
= PO
3
2-
NAD jest głównym akceptorem elektronów
w reakcjach utleniania substratów oddechowych.
Może istnieć w formie utlenionej (z usuniętym
elektronem) (NAD
+
),
albo zredukowanej (NADH).
Pierścień nikotynamidowy (niacyna,witamina PP)
NAD
+
(dinukleotyd nikotynamidoadeninowy)
i NADP (fosforan dinukleotydu nikotynamidoadeninowego)
są koenzymami, które przenoszą elektrony w reakcjach
oksydoredukcyjnych: NAD
+
+ H
+
+ 2e
-
→
→
→
→
NADH (
forma zredukowana
)
NAD
+
jest częściej używany
w reakcjach katabolicznych.
NADP jest częściej używany
w reakcjach anabolicznych.
Kwas nikotynowy
(inaczej:
niacyna, witamina B
3
albo PP
)
jest witaminą, jej brak
powoduje pelagrę (objawy:
zmiany skórne, biegunka
i demencja). Dawniej powszechna
choroba w regionach gdzie je się
dużo kukurydzy.
Obecny w owocach i warzywach.
Wiązanie elektronów przez NAD
+
W tej reakcji odwodorowania jeden atom wodoru z cząsteczki
substratu jest przenoszony bezpośrednio do NAD
+
, natomiast
inny pojawia się w roztworze jako proton.
Oba elektrony z substratu przenoszone są bezpośrednio do
pierścienia nikotynamidowego.
FAD (dinukleotyd flawinoadeninowy) również jest przenośnikiem
elektronów: FAD + 2H
+
+ 2e
-
→
→
→
→
FADH
2
.
Ryboflawina (witamina B
2
)
jest witaminą,
jej brak powoduje
upośledzenie wzrostu.
Obecna w mleku i jajach.
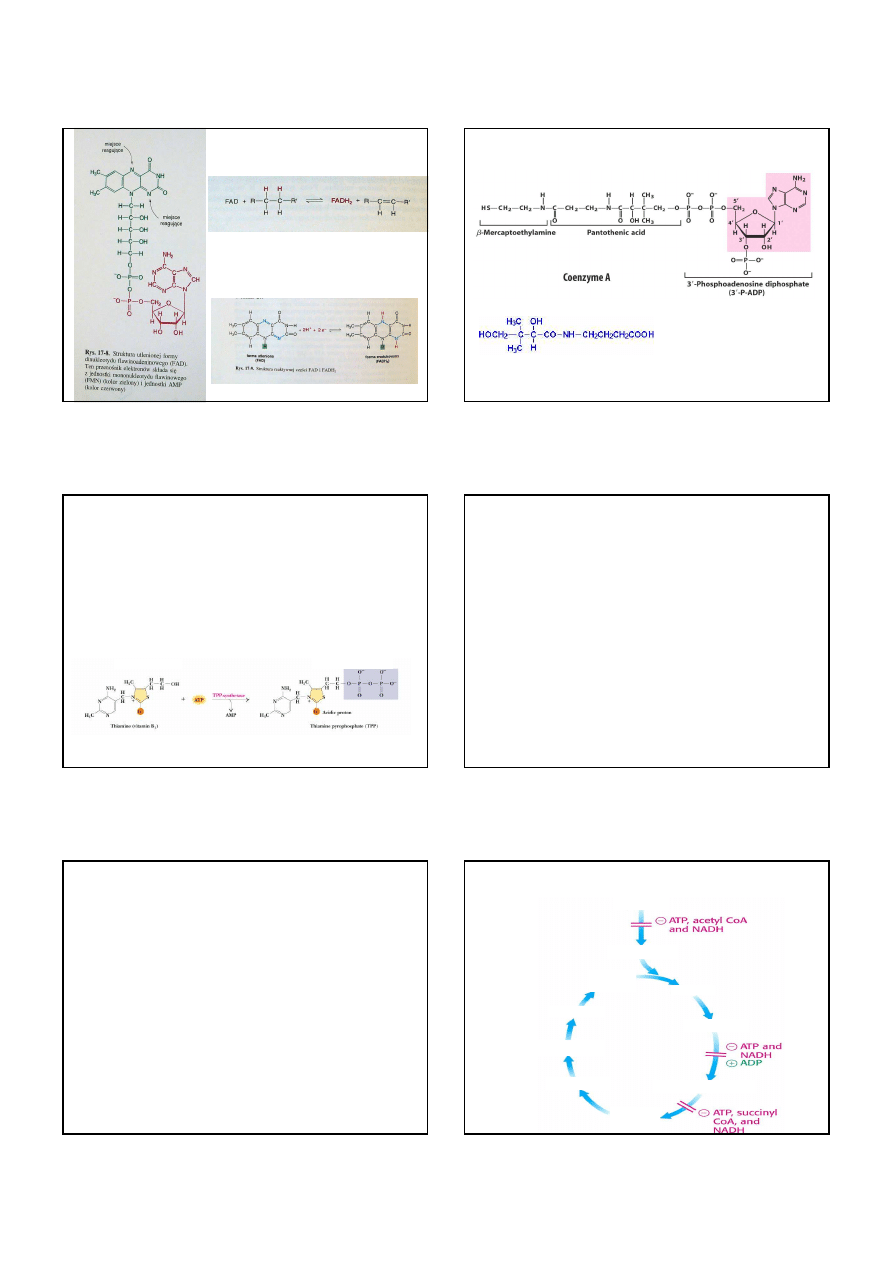
9
Dinukleotyd flawinoadeninowy,
akceptor elektronów w reakcjach
typu:
Przyjmuje 2 elektrony, ale w przeciwieństwie
do NAD
+
, wiąże proton tak samo jak jon
hydroniowy
mononukleotyd
flawinowy
(ryboflawina, B
2
)
AMP
Przenośnikiem grup acetylowych w cyklu kwasu cytrynowego
(i nie tylko) jest koenzym A (CoA).
W jego skład wchodzi
kwas pantotenowy
(witamina B
5
).
Niedobór witaminy B
5
występuje bardzo rzadko.
Obecny wszędzie.
Zakłócenie metabolizmu pirogronianu jest przyczyną choroby beri-beri
tiamina
(witamina B
1
)
pirofosforan tiaminy (TPP)
Beri-beri (owce) jest chorobą spowodowaną brakiem
witaminy B
1
(tiaminy)
w pożywieniu.
Objawy: uszkodzenia obwodowego układu nerwowego,
objawiające się drżeniem rąk i nóg, osłabieniem mięśni i chorobowymi
zmianami na skórze. Ponadto powiększenie serca i objawy niewydolności
krążenia.
U chorych stwierdza się podwyższenie stężenia pirogronianu
i
α
αα
α
-ketoglutaranu. Układ nerwowy jest zdany na glukozę jako jedyne
źródło energii (inne tkanki mogą wykorzystywać tłuszcz).
Pirofosforan tiaminy (TPP) jest grupa prostetyczną 3 enzymów:
dehydrogenazy pirogronianowej, dehydrogenazy
α
αα
α
-ketoglutaranowej
i transketolazy.
Wspólna cecha tych reakcji enzymatycznych:
przenoszenie aktywowanych jednostek aldehydowych.
Związki rtęci i arsenu działają hamująco na te enzymy,
powodując objawy zbliżone do beri-beri.
U nałogowych alkoholików często występują objawy niedoboru
witaminy B
1
z powodu niewłaściwej diety.
Witamina B
1
jest obecna w pełnym ziarnie zbóż.
W diecie wysowęglowodanowej konieczne jest przyjmowanie
podwyższonych ilości witaminy B
1
.
Stechiometria cyklu kwasu cytrynowego
acetylo-CoA + 3 NAD
+
+ FAD + GDP + P
i
+ 2H
2
O
→
→
→
→
2 CO
2
+ 3 NADH + FADH
2
+ GTP + 2H
+
+ CoA
NADH i FADH
2
utworzone w cyklu kwasu cytrynowego
są utleniane w łańcuchu oddechowym. Przeniesienie elektronów
z tych przenośników na O
2
powoduje tworzenie się gradientu
protonowego w poprzek wewnętrznej błony mitochondrialnej.
Powstała w ten sposób siła protonomotoryczna dostarcza energii
do tworzenia ATP: z 1 jednostki octanowej powstaje 10 cząsteczek ATP.
W warunkach beztlenowych w wyniku glikolizy tworzą się
cząsteczki ATP z 1 cząsteczki glukozy.
Glikoliza może zachodzić w warunkach beztlenowych.
Cykl kwasu cytrynowego przebiega
wyłącznie w warunkach tlenowych
(NAD
+
i FAD mogą być zregenerowane w mitochondriach
tylko przez transport elektronów do tlenu cząsteczkowego).
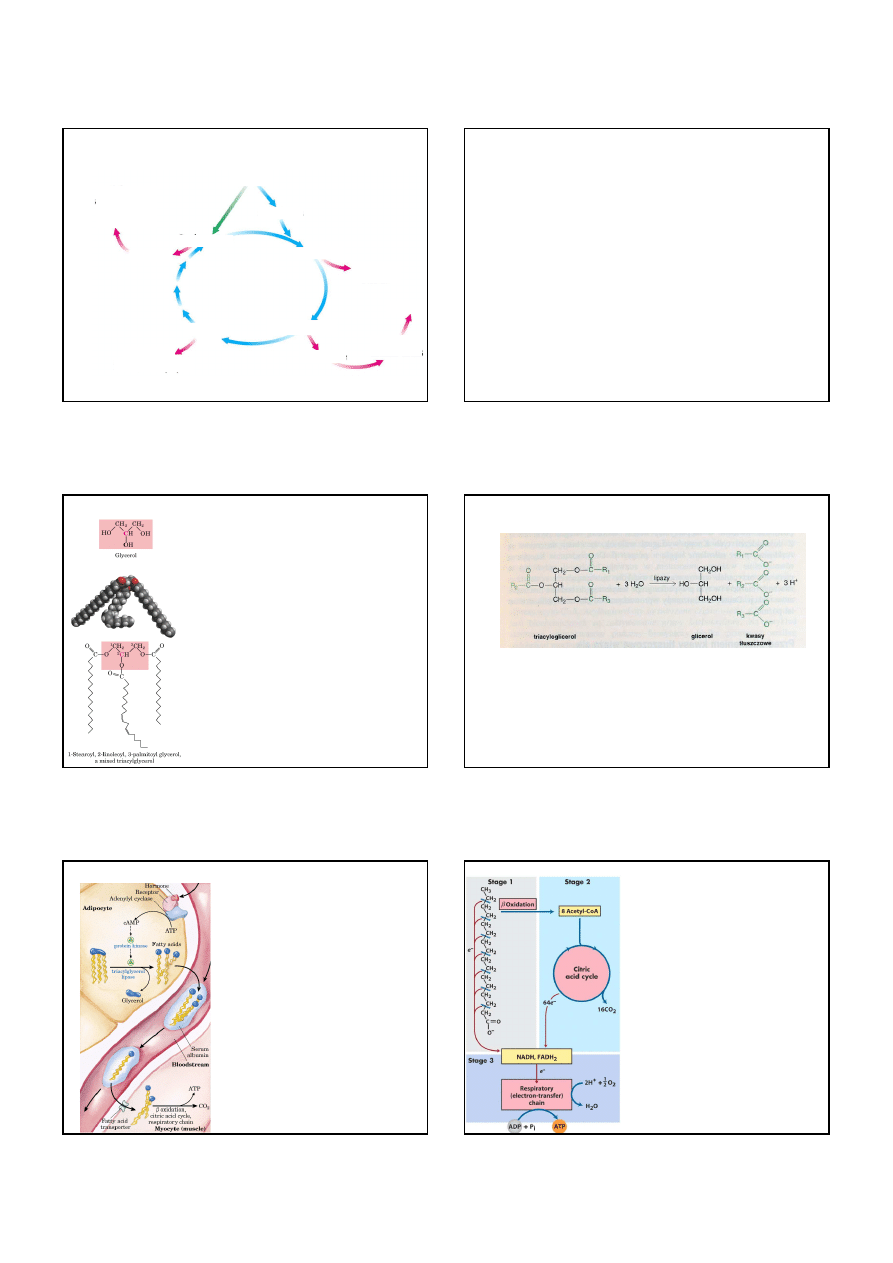
Cykl kwasu cytrynowego i oksydacyjna dekarboksylacja
pirogronianu jest kontrolowana przez stężenia ATP i NADH
pirogronian
acetylo-CoA
cytrynian
izocytrynian
α
αα
α
-ketoglutaran
bursztynylo-CoA
bursztynian
fumaran
jabłczan
szczawiooctan
10
Cykl kwasu cytrynowego jest źródłem prekursorów potzrebnych do
biosyntez
pirogronian
acetylo-CoA
cytrynian
kwasy
tłuszczowe
puryny
inne
aminokwasy
glutaminian
α
αα
α
ketoglutaran
byrsztynylo-CoA
porfiryny, hem, chlorofil
asparaginian
szczawiooctan
inne aminokwasy,
puryny, pirymidyny
Cząsteczki NADH i FADH
2
, utworzone podczas glikolizy,
utleniania kwasów tłuszczowych oraz w cyklu kwasu cytrynowego
są bogate w energię, ponieważ zawierają pary elektronów
o wysokim potencjale przenoszenia.
Energia swobodna uwalniana w znacznej ilości podczas przenoszenia
tych elektronów na tlen cząsteczkowy zostaje wykorzystana
do syntezy ATP.
Fosforylacja oksydacyjna
: proces syntezy ATP zachodzący w wyniku
przeniesienia elektronów z NADH lub FADH
2
na O
2
przez
szereg przenośników elektronów.
Przepływ elektronów z NADH lub FADH
2
na O
2
przez kompleksy białkowe
umiejscowione w wewnętrznej błonie mitochondriów powoduje
wypompowywanie protonów z matrix mitochondrialnej.
Synteza ATP zachodzi na skutek powrotnego przepływu protonów
przez kompleks enzymatyczny(syntaza ATP) do matrix mitochondrialnej.
Katabolizm kwasów tłuszczowych
Triacyloglicerole są magazynami skondensowanej
energii dlatego, że sa zredukowane
i występują w postaci nieuwodnionej.
Utlenienie 1 g kwasu tłuszczowego wyzwala ok.
38 kJ, a utlenienie takiej samej masy
cukrowców albo białek daje ok. 17 kJ.
Ponadto, triacyloglicerole mają charakter
niepolarny, dzięki czemu są magazynowane
praktycznie w postaci
bezwodnej.
Przeciętnie człowiek o masie 70 kg ma zapas
energii:
-
triacylglicerole: 418 000 kJ (ok. 11 kg)
-
białka: 104 000 kJ
-
glikogen: 2500 kJ
-
glukoza: 167 kJ
Gdyby energia ta była przechowywana w formie
glikogenu, człowiek ważyłby o 55 kg więcej.
Hydroliza tłuszczowców: pierwszy etap w ich wykorzystaniu
jako źródła energii
Lipaza triacyloglicerolowa, enzym regulowany hormonalnie.
Adrenalina, noradrenalina, glukagon, hormon adrenokortykotropowy stymulują
cyklazę adenylową w komórkach tłuszczowych.
Regulacja lipolizy
Niski poziom glukozy we krwi powoduje
wydzielanie adrenaliny i glukagonu.
Hormony te wiążą się z receptorami
na powierzchni komórek tłuszczowych
(adipocytów), stymulując cyklazę adenylową.
cAMP uwaktywnia kinazę, która fosforyluje
lipazę.
Ufosforylowana lipaza hydrolizuje
triacyloglicerole.
Uwolnione kwasy tłuszczowe są przenoszone
do krwi, gdzie wiążą się niekowalencyjnie z
albuminą.
Albumina przenosi kwasy tłuszczowe
do komórek.
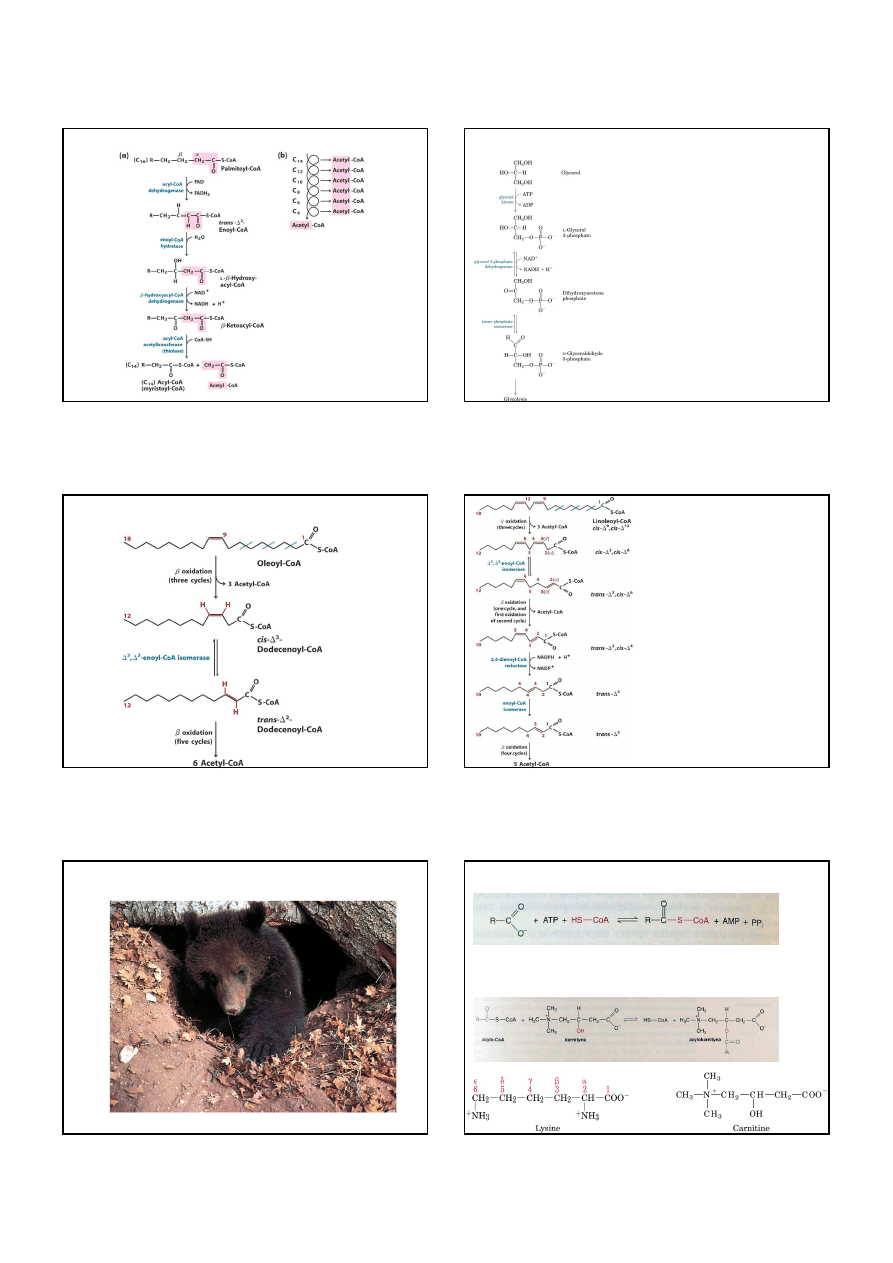
Kwasy tłuszczowe wchodzą
do cyklu kwasu cytrynowego
w wyniku
ββββ
-oksydacji
3 etapy:
1.
ββββ
-oksydacja: usunięcie dwuwęglowych
jednostek w postaci acetylo-CoA.
2. Grupy acetylowe zostają utlenione
do CO
2
w cyklu kwasu cytrynowego.
3. Elektrony uwolnione w czasie utleniania
w etapie 1 i 2 przechodzą na tlen
przez łańcuch oddechowy
w mitochondrium.
11
W wyniku
ββββ
-oksydacji kwasu palmitynowego powstaje acetylo-CoA
Glicerol powstały w procesie lipolizy ulega fosforylacji i utlenieniu
i wchodzi do glikolizy.
glicerol
3-fosfoglicerol
fosforan dihydroksyacetonu
aldehyd 3-fosfoglicerynowy
Oksydacja jednonienasyconych kwasów tłuszczowych wymaga obecności
izomerazy enoilo-CoA
Oksydacja wielonienasyconych
kwasów tłuszczowych wymaga
ponadto reduktazy
dienoilo-CoA
Niedźwiedź przeprowadza
ββββ
-oksydację w czasie snu zimowego
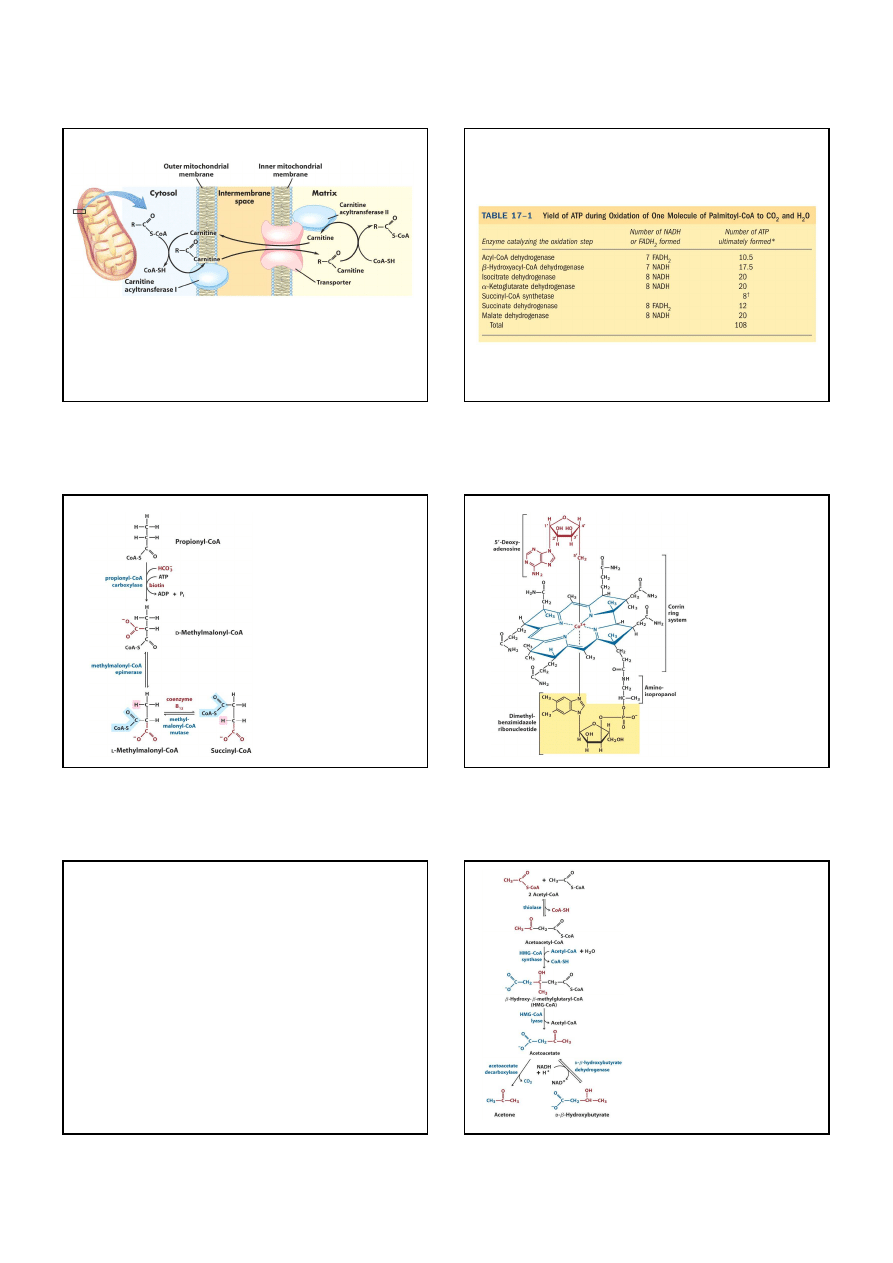
Przed utlenieniem kwasy tłuszczowe wiążą się z koenzymem A
Syntetaza acylo-CoA (tiokinaza kwasów tłuszczowych)
Zaktywowane kwasy tłuszczowe wiążą się z karnityną
Acylotransferaza karnitynowa I
12
Karnityna przenosi zaktywowane kwasy tłuszczowe
do matriks mitochondrialnej
Acylotransferaza
karnitynowa I
Acylotransferaza
karnitynowa II
translokaza
Cząsteczki acylo-CoA o długich łańcuchach nie przenikają
łatwo przez wewnętrzną błonę mitochondrialną
⇓
⇓
⇓
⇓
potrzebny jest specjalny mechanizm transportu.
W wyniku utlenienia cząsteczki kwasu palmitynowego
do CO
2
i H
2
O powstaje 108 cząsteczek ATP.
Kwasy tłuszczowe o nieparzystej
liczbie atomów węgla wymagają
przemiany propionylo-CoA
w bursztynylo-CoA.
W reakcji tej niezbędna jest
witamina B
12
(kobalamina).
Bursztynylo-CoA
wchodzi do cyklu
kwasu cytrynowego.
Koenzym B
12
dostarcza
wolnych rodników do katalizy
wewnątrzcząsteczkowych
przesunięć wodoru
Witamina B
12
jest wytwarzana wyłącznie
przez mikroorganizmy: zwierzęta i rośliny są niezdolne
do jej syntezy.
W żołądku wydziela się glikoproteina zwana
czynnikiem wewnętrznym
(IF, intrinsic factor)
, która w świetle jelita wiąże kobalaminę.
W wiązaniu kobalaminy bierze też udział transkobalamina II.
Najbardziej wrażliwy na brak kobalaminy jest układ krwiotwórczy,
ponieważ krwinki mają krótki okres życia.
Skutkiem braku witaminy B
12
jest niewystarczająca synteza
puryn i tyminy.
Przyczyną niedokrwistości złośliwej
jest niedobór czynnika wewnętrznego (IF).
Kiedy acetylo-CoA nie może być
w całości przetworzony w CO
2
w cyklu kwasu cytrynowego,
powstają
„ciała ketonowe”
:
acetooctan, aceton i
ββββ
-hydroksymaślan
.
Wejście acetylo-CoA w ten cykl zależy
od dostępności szczawiooctanu,
ponieważ po przereagowaniu
z nim tworzy się cytrynian.
W okresie głodu lub w cukrzycy
szczawiooctan jest zużywany
do syntezy glukozy.
W takich warunkach acetylo-CoA
jest kierowany do tworzenia
acetylooctanu i
ββββ
-hydroksymaślanu.
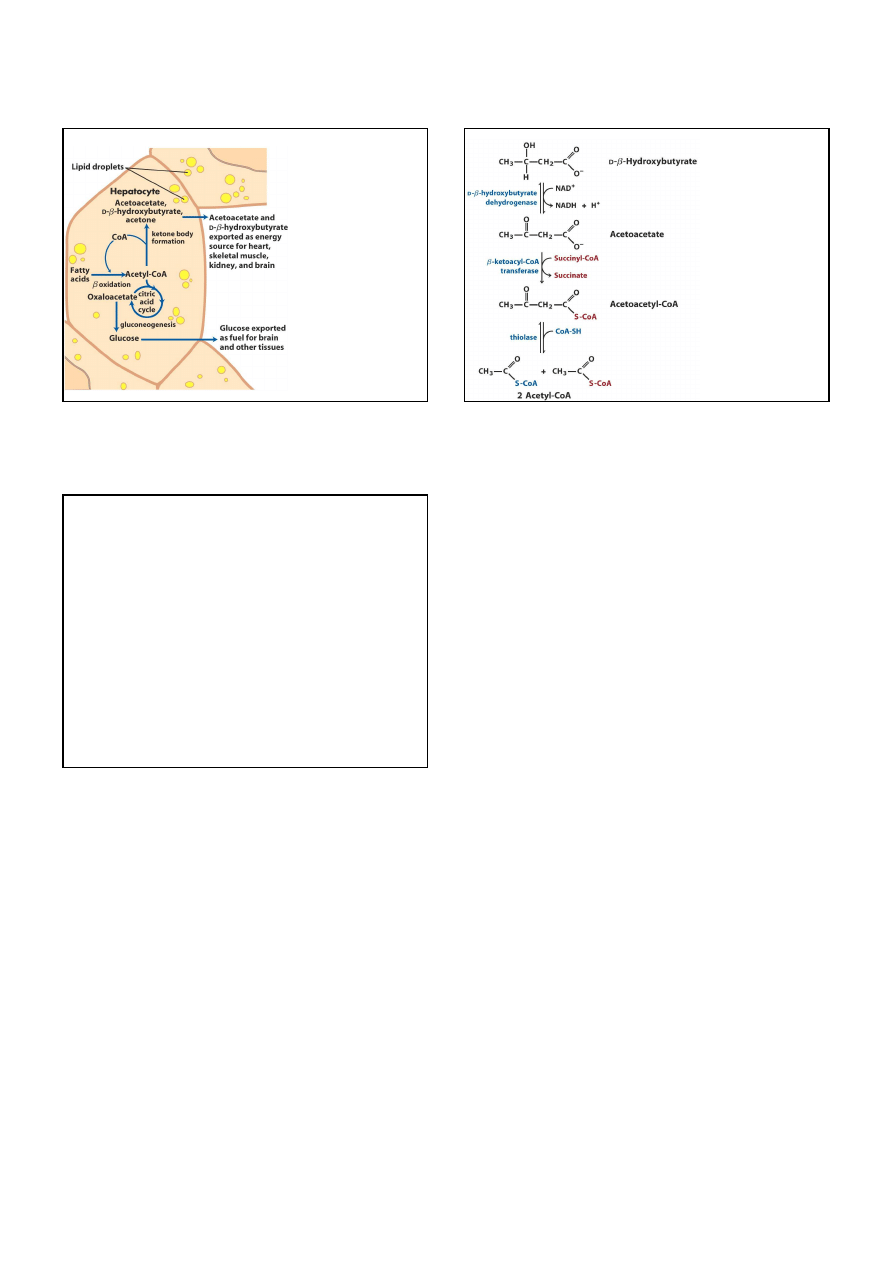
13
W warunkach wymuszonej
glukoneogezy
(np. w cukrzycy
albo w czasie postu)
cykl kwasu cytrynowego
ulega spowolnieniu
z powodu zużywania
szczawiooctanu
do syntezy glukozy.
Ciała ketonowe mogą
być zużywane jako paliwo
przez serce, mięśnie,
nerki i mózg.
ββββ
-hydroksymaślan
albo acetyloaceton
mogą być paliwem
w tych komorkach,
które nie otrzymują
wystarczającej ilości
glukozy (np. w mózgu
w czasie postu).
ββββ
-hydroksymaślan
ulega rozkładowi
do acetylo-CoA.
12. Biochemia - cykl kwasu cytrynowego i
ββββ
-oksydacja kwasów
tłuszczowych
Tematy do zapamiętania
1. Cykl kwasu cytrynowego: schemat, znaczenie.
2. 3 etapy procesu oddychania komórkowego.
3. Rola cyklu kwasu cytrynowego w katabolizmie i biosyntezie.
4. Rola NAD i FADH
2
.
5. Rozkład kwasów tłuszczowych, ciała ketonowe.
Wyszukiwarka
Podobne podstrony:
14 BIOCHEMIA cykl kwasu cytrynowego 2id 15387 ppt
12 BIOCHEMIA cykl kwasu cytrynowegoid 13424 ppt
Biochemia 7 Cykl kwasu cytrynowego
Biochemia 7K Cykl kwasu cytrynowego
Oleksyszyn, Biochemia II, zagadnienia do cykl kwasu cytrynowego
Biochemia wykład 14 Cykl kwasu cytrynowego
Biochemia 7K Cykl kwasu cytrynowego
Cykl Krebsa cykl kwasu cytrynowego
Cykl Krebsa, cykl kwasu cytrynowego
Cykl kwasu cytrynowego
Cykl kwasu cytrynowego Krbsa
CYKL KWASU CYTRYNOWEGO
glikoliza i cykl kwasu cytrynowego reakcje
więcej podobnych podstron