Cechy mięśnia sercowego
Mięsień sercowy składa się z 2 mięśni: mięśnia przedsionków i mięśnia komór, które są od siebie odgrodzone pierścieniami włóknistymi, otaczającymi ujście przedsionkowo-komorowe. Mięsień sercowy zbudowany jest z komórek mięśniowych, które posiadają włókienka mięśniowe i podstawowe elementy kurczliwe, tworzące sarkomery. Błony komórkowe sąsiednich komórek mięśniowych silnie do siebie przylegają w miejscach występowania prążków Z, tworząc pozazębianą błonkę zwaną wstawką. Dzięki nim i połączeniom komunikującym, pobudzenie przenosi się z jednej komórki na drugą i mięsień sercowy stanowi syncytium fizjologiczne. Odpowiada na bodziec maksymalnym skurczem, zgodnie z prawem „wszystko albo nic”.
Mięsień sercowy należy do mięśni wolno kurczących się i ma względnie małą aktywność, jego miocyty są zależne od metabolizmu tlenowego, a tym samym od stałego dopływu tlenu. Ich skurcz wymaga dopływu Ca ze zbiorników końcowych oraz zewnątrzkomórkowego. W przeciwieństwie do innych mięśni prążkowanych, kurczy się całkowicie, niezależnie od naszej woli, co jest możliwe dzięki tkance rozrusznikowej. Serce kontynuuje swoje skurcze po przecięciu wszystkich nerwów, dzieje się tak dzięki wyspecjalizowanej tkance rozrusznikowej tworzącej układ przewodzenia, który w normalnych warunkach rozprzestrzenia impulsy na całe serce. Cechą tej tkanki jest niestabilny potencjał błonowy. Pomiędzy kolejnymi impulsami nie ma stałych wartości potencjału, lecz po każdym potencjale czynnościowym występuje stałe obniżenie potencjału błonowego, aż do poziomu krytycznego, przy którym następuje wyzwolenie kolejnego potencjału czynnościowego. Ta wolna depolaryzacja między potencjałami nazywa się potencjałem rozrusznika. Im bardziej krzywe jest nachylenie krzywej potencjału rozrusznika, tym szybszy jest rytm wyładowań tkanki rozrusznika.
Spoczynkowy potencjał błony miocytu wynosi -90 mV. Jest to tk pobudliwa, potencjł czynnościowy warunkuje zmiany w stężeniu K wewnątrz i Na na zewnątrz. Pobudzenie wywołuje potencjał czynnościowy trwający 250 ms. Odpowiedź skurczowa zaczyna się zaraz po depolaryzacji i trwa ok. 1,5 raza dłużej niż potencjał czynnościowy. Okres refrakcji bezwzględnej tra podczas faz 0,1,2 oraz przez część fazy 3 aż potencjał błony nie osiągnie -50 mV. Okres refrakcji względnej trwa do czasu rozpoczęcia się fazy 4. Skurcz tężcowy jest niemożliwy w mięśniu sercowym, co stanowi zabezpieczenie przed śmiercią spowodowaną dłuższym skurczem tężcowym. Mięsień jest obficie unaczyniony i unerwiony z wysoką zawartością mioglobiny. Siła skurczu m. sercowego zwiększa się pod wpływem krążących amin katecholowych. Wzrost siły skurczu występuje bez zmiany długości mięśnia.
Układ bodźco-przewodzący serca
Prawidłowa czynność serca zależy od generowania stanu czynnościowego w strukturach serca w sposób ściśle uporządkowany w czasie i przestrzeni, co jest wynikiem zróżnicowanych właściwości morfologicznych i czynnościowych poszczególnych części serca, tj. układu bodźco-przewodzącego mięśnia przedsionków i komór.
Układ ten generuje impulsy stanu czynnościowego w sercu bez dopływu czynników zewnętrznych, odpowiada więc za automatyzm serca i przewodzi impulsy w sposób ściśle zsynchronizowany i uporządkowany. Zbudowany jest ze skupisk komórek: węzłów i pęczków.
W układzie tym wyróżnia się:
węzeł zatokowo-przedsionkowy (ujście żyły głównej)
Jest pierwszorzędowym ośrodkiem automatyzmu, a występującą tu grupę komórek odpowiedzialnych za wytwarzanie impulsów nazywamy rozrusznikiem serca. Struktura ta narzuca swój rytm całemu sercu. Typową cechą komórek węzła jest brak potencjału spoczynkowego. Zaraz po zakończonej repolaryzacji poprzedniego pobudzenia następuje przesuwanie potencjału błonowego w kierunku dodatnim, jest to tzw. powolna spoczynkowa depolaryzacja. Kolejna depolaryzacja rozpoczyna się od potencjału mniej ujemnego, a szybkość rozrastania potencjału i jego amplituda są niewielkie (brak plateau). Następnie zachodzi repolaryzacja i znów spoczynkowa depolaryzacja. Im bardziej stromy jest przebieg krzywej powolnej spoczynkowej depolaryzacji, tym szybszy jest rytm pobudzeń narzucany sercu przez jego rozrusznik. Na szybkość rytmu wpływa też zmiana potencjału progowego rozrusznika. W komórkach tego węzła występują grupy komórek o różnej własnej częstotliwości pobudzeń. Właściwy rozrusznik serca jest w głowie węzła, ale niedokrwienie, pobudzenie nerwów serca, zatrucia, mogą powodować zmianę jego położenia, tzw. wędrowanie rozrusznika
węzeł przedsionkowo-komorowy (przegroda międzyprzedsionkowa)
Można wyróżnić tu 3 strefy: przedsionkowo-węzłowa, węzłowa, węzłowo-pęczkowa. W strefie przedsionkowo-węzłowej (złącze) komórki cechuje nieznaczny ujemny potencjał spoczynkowy, brak właściwości automatycznych, powolne narastanie i mała amplituda potencjału czynnościowego, brak wyraźnego plateau, potencjały mogą się sumować, dzięki czemu przewodzenie jest wolne, co opóźnia aktywację komór w stosunku do przedsionków. W strefie węzłowej komórki węzła przedsionkowo-komorowego mają swój automatyzm, jednak przebieg powolnej spoczynkowej depolaryzacji jest wolniejszy niż w węźle zatokowo-przedsionkowym, ale gdy zostanie on zablokowany, same stają się właściwym rozrusznikiem i swój rytm (węzłowy). Strefa węzłowo-pęczkowa stanowi początek pęczka Hisa
pęczek przedsionkowo-komorowy Hisa
włókna Purkiniego (komorowy układ przewodzący)
Są to komórki mięśniowe, przewodzące serca, mają mało włókien mięśniowych, dużą zawartość glikogenu, dużą średnicę, są zebrane w pęczki oddzielone od siebie otoczkami tkanki łącznej, są one izolowane od pozostałych części komór z wyjątkiem zakończeń komórek przewodzących. Układ przewodzący komór rozpoczyna się od części węzła przedsionkowo-komorowego (pęczka Hisa), który stanowi jedyne połączenie między komorami a przedsionkami (połączenie mięśniowe). Pęczek ten dociera do górnej części przegrody międzykomorowej i dzieli się na 2 gałązki: lewą prawą. Obie przechodzą odpowiednie powierzchnie przegrody międzykomórkowej i leżą tuż pod wsierdziem. Gałązka prawa dzieli się na kilka odnóg, przebijających światło prawej komory i kończących się z mięśniami brodawkowatymi. Gałązka lewa dzieli się u samej góry na 2 lub więcej części, z których najbardziej tylna przebija światło komory, tworzy połączenia z mięśniami brodawkowatymi
ścięgna rzekome
Są to odgałęzienia układu przewodzącego komór, które przecinają ich światło, odgrywają dużą rolę przy aktywacji komór, gdyż wysyłają drogę impulsów biegnących do poszczególnych części komór, a tym samym skracają ich całkowity czas pobudzenia
Właściwości i potencjały czynnościowe komórek układu bodźco-przewodzącego
Rozrusznikiem dla potencjałów czynnościowych mięśnia sercowego, które wytwarzają skurcz jest tkanka przewodząca. Pierwszorzędowy ośrodek automatyzmu stanowi węzeł zatokowo-przedsionkowy, który ma zdolność powolnej spoczynkowej depolaryzacji, potencjał czynnościowy nie posiada fazy plateau. Faza repolaryzacji jest stroma, a powolna spoczynkowa depolaryzacja wynosi od -60 do -50 mV. Mięsień sercowy jest podporządkowany węzłowi zatokowo-przedsionkowemu, który jest rozrusznikiem serca, może działać bez układu nerwowego. Pobudzenia w węźle sięgają częstości 60-90/min, średnio ok. 72 pobudzenia/min, pobudzenie jest tu najszybsze i najszybciej opada. Drogi łączące węzeł zatokowo-przedsionkowy z przedsionkowo-komorowym to 3 pęczki międzywęzłowe: przedni Bachmana, środkowy Wenkeneacha i tylny Thorela. Impulsy docierają do strefy przedsionkowo-węzłowej za pomocą tych pęczków oraz pęczka pierścieniowatego zatokowo-przedsionkowego i innych dróg. Poza tym impulsy rozchodzą się także na przedsionek lewy i prawy. W strefie przedsionkowo-węzłowej impulsy ulegają sumowaniu i osiągają amplitudę umożliwiającą dalsze przewodzenie. Następuje tu większe zwolnienie przewodzenia. Pęczek przedsionkowo-komorowy Hisa jest jedynym połączeniem mięśniowym między mięśniem przedsionków i komór, za jego pośrednictwem depolaryzacja jest przenoszona do komór. W obrębie komór pęczek dzieli się na 2 odnogi, przechodzące pod wsierdziem w komórki mięśniowe sercowe przewodzące- włókna Purkiniego. Wcześniej pobudzenie obejmuje odnogę prawą. Depolaryzacja przewodzona przez pęczek Palladino-Hisa szerzy się na komórki mięśnia komorowego, począwszy od przegrody międzykomorowej, przez mięśnie brodawkowate, do koniuszka serca. Po dotarciu tu(do koniuszka), pobudzenie obejmuje mięśnie od strony wewnętrznej- podwsierdziowe i zewnętrznej- podosierdziowe, następnie pobudzenie rozszerza się ku podstawie serca. Najpóźniej pobudzeniu ulega część podosierdziowa, gdyż nie ma tam (jak w części podwsierdziowej) włókien Purkiniego. Pobudzenie to w części podosierdziowej następuje po 80-90 ms. Pomiędzy poszczególnymi komórkami przewodzenie obywa się za pomocą wstawek. Są to 2 warstwy błony i prążków Z sąsiadujących komórek, są pofałdowane, stanowią czynnościowe połączenia między komórkami. W miejscu styku po zewnętrznej(?) stronie błony komórkowej znajdują się tzw. plamki przylegania (zbity materiał prążka), odcinki pomiędzy nimi noszą nazwę obwódki przylegania. Co kilka sarkomerów wstawki przesuwają się o 1 sarkomer i przebiegają na poziomie następnego sarkomeru. Wstawki biegnące na 2 różnych poziomach są połączone tzw. złączami ścisłymi niskooporowymi, co pozwala na przewodzenie impulsów między komórkami.
Potencjały spoczynkowe i czynnościowe komórek roboczych
Potencjał spoczynkowy wynosi 90 mV. W spoczynku jony potasowe stale wychodzą z komórek mięśnia sercowego przez kanały potasowe w błonie komórkowej na zewnątrz- odkomórkowy wolny prąd jonów potasowych. Jednocześnie jony sodowe stale wchodzą do komórek przez kanały sodowe- dokomórkowy wolny prąd jonów sodowych. Przesunięcia w rozmieszczeniu jonów K i Na wyrównywane są przez pompę sodowo-potasową. W czasie pobudzenia komórek mięśnia komór serca, występuje czynnościowy potencjał składający się z 4 faz. Rozpoczyna się fazą 0- bardo szybko zachodzącą depolaryzacją, po której występuje nieznaczna repolaryzacja, czyli faza 1, przechodzącą w fazę 2, charakteryzującą się utrzymywaniem stałej depolaryzacji w czasie ok. 300 ms. W fazie 3 następuje powrót do potencjału spoczynkowego, faza 0 potencjału czynnościowego jest wywołana dokomórkowym szybkim prądem jonów Na i w niewielkim stopniu przez dokomórkowy wolny prąd jonów Ca. Faza 1 w komórkach mięśnia przedsionków wiąże się z odkomórkowym prądem K, natomiast w kom mięśnia komór jest wywołana przez dokomórkowy prąd jonów Cl. W czasie fazy 2 występuje równowaga pomiędzy dokomórkowym prądem jonów Ca i odkomórkowym prądem jonów K. W fazie 0, 1, 2 jony Ca napływają do sarkoplazmy z zewnętrznej powierzchni sarkolemy, gdzie są magazynowane w glikokaliksie i tylko częściowo ze zbiorników sarkoplazmatycznych. W fazie 3 następuje przewaga odkomórkowych prądów jonów dodatnich, głównie wywoływana przez jony K, i powrót ujemnego potencjału w komórce. Okres bezwzględnej niewrażliwości (bezwzględna refrakcja), obejmuje depolaryzację oraz fazy 1,2 repolaryzacji. Okres względnej niewrażliwości (względna refrakcja), przypada na fazę 3 repolaryzacji. Łącznie oba te okresy niewrażliwości są dłuższe od czasu skurczu mięśnia sercowego. Dzięki temu w warunkach prawidłowych w mięśniu tym nie występują skurcze tężcowe. Siła skurczu m. sercowego zależy od początkowej długości jego komórek, podobnie jak to występuje w m. poprzecznie prążkowanych szkieletowych. Przy optymalnym wypełnieniu jam serca i optymalnym rozciągnięciu komórek m sercowego występują max skurcze zgodne z tzw. prawem Starlinga. Energię potrzebną do skurczów m sercowy czerpie ze składników odżywczych, którymi są: 60% kw tłuszczowych, których połowę stanowią wolne kwasy tłuszczowe, 35% węglowodany oraz aminokwasy, ciała ketonowe, kwas mlekowy, kwas pirogronowy.
Sprzężenie elektromechaniczne w mięśniu sercowym
Molekularny mechanizm działania układów kurczliwych mięśnia sercowego w zasadzie nie różni się od tych mechanizmów w mięśniach szkieletowych. Przebieg zjawisk jest wolniejszy, co powoduje wolniejsze narastanie siły skurczu oraz mniejszą maksymalną prędkość skracania. Czynnikiem aktywującym układy kurczliwe pobudzanej komórki jest pojawienie się w sarkoplazmie zjonizowanego wapnia w stężeniu nadprogowym, ale poszczególne sprzężenia różnią się od tych w mięśniu szkieletowym, co ma duże znaczenie dla jego funkcji. Mięsień sercowy w odróżnieniu od szkieletowego do procesów sprzężenia używa nie tylko wapnia wewnątrzkomórkowego, ale również nie może obyć się bez wapnia zewnątrzkomórkowego. Usunięcie tych jonów ze środowiska powoduje rozkojarzenie elektromechaniczne, co świadczy o wymianie Ca pomiędzy wnętrzem komórek, a środowiskiem zewnątrzkomórkowym. Struktury błonowe odpowiedzialne za wydzielanie i wychwytywanie Ca są różne niż w mięśniu szkieletowym. Zbiorniki końcowe siateczki śródplazmatycznej tworzą ścisłe wiązania nie tylko z cewkami T, ale również z zewnętrzną błoną komórkową. Ma to duże znaczenie w przedsionkach, gdzie cewki T są bardzo rzadkie. W czasie pobudzenia komórki, do jej wnętra napływają jony Ca. Część z nich bezpośrednio dociera do układów kurczliwych i uczestniczy w ich aktywacji, a część jest odkładana w tzw. zbiornikach pośrednich. Są to niektóre elementy siateczki śródplazmatycznej. W czasie spoczynku są przekazywane jony ze zbiorników pośrednich oraz jony znajdujące się w komórce, do zbiorników końcowych, które są w sąsiedztwie błony komórkowej lub cewek T. Im dłuższa jest przerwa między pobudzeniami, tym więcej jonów Ca zostaje przekazanych do zbiorników końcowych. W czasie kolejnego pobudzenia ze zbiorników końcowych zostaje uwolniony wapń, który jest głównym aktywatorem skurczu- ok. 75% siły skurczu. Pozostałe 25% siły skurczu zależy od napływu wapnia zewnątrzkomórkowego przez błonę. Jony Ca są uwalniane ze zbiorników końcowych na skutek gwałtownych zmian pola elektrycznego powstających w ich pobliżu. W czasie każdego cyklu ilość jonów Ca, podobna do ilości jonów napływających do komórki, zostaje wydalona na zewnątrz oraz osiąga swój szczyt pod koniec plateau potencjału czynnościowego. Rozkurcz służy do wychwytu jonów Ca przez kanaliki podłużne siateczki śródplazmatycznej i transportowane są do zbiorników końcowych. Ten typ sprzężenia powoduje zależność siły skurczu od rytmu oraz od przebiegu w czasie potencjałów komórkowych.
Zależność siły skurczu od przebiegu potencjału czynnościowego
wydłużenie czasu trwania potencjałów na drodze elektrycznej powoduje wzmocnienie aktywowanego przez niego skurczu oraz znaczne wzmocnienie skurczu następnego w wyniku zwiększonego napływu jonów Ca w czasie wydłużonego plateau.
Zależność siły skurczu od rytmu
skrócenie przerwy między poprzedzającym, a trwającym pobudzeniem powoduje obniżenie siły skurczu, a wydłużenie przerwy spowoduje zwiększenie siły skurczu. Im więcej czasu upłynie, tym wypełnienie zbiorników końcowych jest mniejsze. Skurcze dodatkowe są tym słabsze, im wcześniej w cyklu występują. Maksymalną siłę skurczu uzyskuje się po przerwie ok. 1 s. Siła skurczu po skurczu dodatkowym jest większa niż innych skurczów, tzw. wzmocnienie o skurczu dodatkowym, co jest spowodowane nowym napływem jonów wapniowych. Zbiorniki końcowe, które nie zdążyły wypełnić się wydzielają mało Ca, całkowita ilość Ca w komorze wzrasta. Po skurczu dodatkowym są transportowane do kanalików końcowych i wypełniają je w większym stopniu niż w czasie normalnego rytmu.
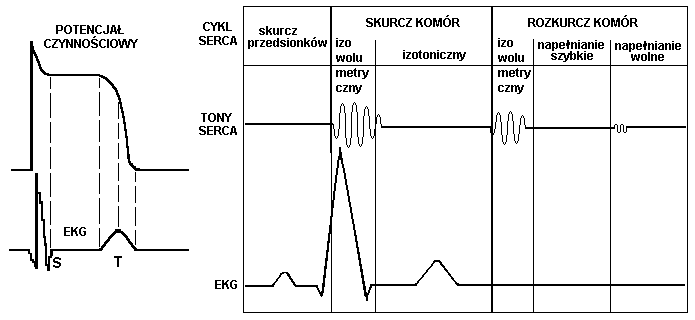
Cykl hemodynamiczny serca
Cykl pracy serca (cykl hemodynamiczny serca) jest indukowany przez układ bodźco-przewodzący serca, który pobudza kardiomiocyty do skurczu w odpowiedniej kolejności wymuszając przepływ krwi. Na układ bodźco-przewodzący wpływa impulsacja z układu autonomicznego regulując rytm serca i dostosowując go do aktualnych potrzeb ustroju. Za początek cyklu pracy serca powszechnie przyjmuje się pauzę. W czasie pauzy przedsionki i komory serca są w stanie rozkurczu i krew pod wpływem gradientu (różnicy) ciśnień przelewa się z żył głównych i płucnych do przedsionków, a stamtąd do komór. Następnie dochodzi do skurczu przedsionków, zwiększając ciśnienie w przedsionkach i powodując dopchnięcie jeszcze porcji krwi do komór, objętość komór po skurczu przedsionków nazywa się objętością późnorozkurczową, a ciśnienie panujące w komorach ciśnieniem późnorozkurczowym lub obciążeniem wstępnym. Ciśnienie w komorach wzrasta powyżej ciśnienia w przedsionkach i następuje zamknięcie zastawek, odpowiednio trójdzielnej po prawej i mitralnej po lewej stronie serca i uderzenie krwi o zastawki od strony komór. Zamknięcie zastawek wywołuje efekt akustyczny w postaci pierwszego tonu serca. Następnie rozpoczyna się skurcz komór nie powodujący zmiany objętości krwi zawartej w komorach jest to tzw. skurcz izowolumetryczny. W czasie skurczu izowolumetrycznego narasta napięcie ścian komór serca, co powoduje wzrost ciśnienia w komorach. Gdy ciśnienie przekroczy ciśnienie odpowiednio w pniu płucnym i aorcie następuje faza wyrzutu i pewna objętość krwi zostaje wypchnięta do pnia płucnego i aorty, jest to tzw. objętość wyrzutowa. Po fazie wyrzutu ciśnienie w komorach zaczyna spadać co powoduje zamknięcie zastawek pnia płucnego i aortalnej i wywołuje drugi ton serca.
W komorach po wyrzucie pozostaje zawsze pewna ilość krwi jest to objętość późnoskurczowa, a ciśnienie panujące w komorze nazywane jest ciśnieniem późnoskurczowym. Rozpoczyna się rozkurcz komór. W początkowej fazie rozkurczu ciśnienie w komorach jest jeszcze wyższe niż w przedsionkach i zastawki przedsionkowo-komorowe są zamknięte, ta faza rozkurczu nazywana jest rozkurczem izowolumetrycznym. Gdy ciśnienie w komorach spadnie poniżej ciśnienia w przedsionkach zastawki otwierają się i krew przelewa się z przedsionków do komór i cały cykl powtarza się.
Zjawiska akustyczne serca
Drgania towarzyszące pracy serca są niejednorodnej częstotliwości. Można je podzielić na stale występujące w warunkach fizjologicznych, czyli tony serca, oraz na patologiczne szmery. W każdym cyklu pracy serca występują tony:
1 ton serca- wywołany zamykaniem się zastawek przedsionkowo-komorowych i początkiem skurczu serca. Trwa ok. 150 ms i obejmuje drgania o częstotliwości 25-45 Hz
2 ton serca- powstaje w czasie zamykania się zastawek aorty i pnia płucnego. Trwa 120 ms i 50 Hz
3 ton serca- występuje w rozkurczu, w okresie wypełniania się komór krwią napływającą z przedsionków. Spowodowany jest przez wibrację krwi, szybko wypełniającej jamy obu komór. Trwa najkrócej
W warunkach patologicznych występują dodatkowo w czasie pracy serca szmery, różniące się od tonów. Są one zazwyczaj spowodowane nieprawidłowymi warunkami przepływu krwi pomiędzy przedsionkami, komorami i zbiornikami tętniczymi. Zjawiska akustyczne występujące w sercu można zarejestrować, otrzymując zapis w postaci fonokardiogramu. Do skóry w okolicy serca przystawia się specjalny przetwornik. Wzmacnia się prądy elektryczne wzbudzane w przetworniku, przez drgania towarzyszące pracy serca i rejestruje się je za pomocą elektrokardiografu.
8. Objętość wyrzutowa.
Objętość wyrzutowa serca (SV) - objętość krwi wyrzucanej z prawej i lewej komory odpowiednio do tętnicy płucnej i aorty. Objętości wyrzutowe obu komór mogą się nieco różnić w czasie poszczególnych skurczów, jednak średnia ich wartość jest jednakowa, wynosi ona średnio 80ml w spoczynku i 200 ml podczas wysiłku.
Po zakończeniu wyrzutu pozostaje w komorach pewna ilość krwi, jest to krew zalegająca - zależną od niej objętość komory nazywamy objętością późnoskurczową.
Objętość późnoskurczowa = objętość późnorozkurczowa - obj. wyrzutowa.
Stosunek objętości wyrzutowej do objętości późnorozkurczowej nazywamy frakcją wyrzutu (określa ona, jaka część krwi, znajdującej się w komorach na początku skurczu, została wyrzucana do tętnic; wynosi 60-70%; jest wskaźnikiem stanu czynnościowego mięśnia sercowego)
9. Pojemność minutowa serca i jej rozkład na poszczególne narządy w spoczynku i w czasie wysiłku.
Pojemność minutowa serca (CO) - jest to ilość krwi przepompowywanej przez serce w ciągu jednej minuty. Jest to jednocześnie ilość krwi , która przepływa przez krążenie i transportuje rożne związki do tkanek czy z tkanek.
Poj. Minutowa = obj. wyrzutowa*liczba skurczów serca na minutę
wynosi średnio 5 - 5,5 litra
zmienia się zależnie od masy ciała, wieku, stanu metabolicznego
istnieje powiązanie między powierzchnią ciała a pojemnością minutową:
poj. minutowa/pow. Ciała = wskaźnik sercowy (3,5 litra/m2/min)
Prawo serca Starlinga - siła skurczu (i objętość wyrzutowa) jest wprost proporcjonalna
od wstępnego rozciągnięcia włókien mięśnia serca (stopnia wypełnienia komór)
→ prawidłowa adaptacja serca do wzmożonego zapotrzebowania na tlen odbywa się przez zwiększenie objętości wyrzutowej, a nie przyśpieszenie akcji serca.
Oprócz tego pojemność minutowa może się zwiększać przez przyspieszenie akcji serca (mniej korzystne dla organizmu - pogarsza krążenie wieńcowe, a w skrajnych przypadkach może prowadzić do niewydolności krążenia - trzepotanie lub migotanie komór.).
Pojemność minutowa zwiększa się podczas:
wysiłku fizycznego (nawet 5 razy)
stanów stresowych
podniesienia temperatury ciała
zmianie pozycji ciała z leżącej na siedzącą
pobudzenie układu współczulnego (zmniejszenie przy pobudzeniu przywspółczulnego)
10. Elektrokardiografia - zasady wykonania, rodzaje odprowadzeń.
Elektrokardiografia polega na rejestracji zmian potencjałów powstających na powierzchni ciała lub w jego wnętrzu pod wpływem depolaryzacji i repolaryzacji serca.
Umieszczając elektrody na końcach i rejestrując różnice potencjałów występujące pomiędzy elektrodami odbiera się czynność bioelektryczną mięśnia sercowego, czyli elektrokardiogram - EKG, za pomocą tzw. odprowadzeń kończynowych.
Trzy odprowadzenia kończynowe dwubiegunowe, klasyczne Einthovena, uzyskuje się rejestrując różnice potencjałów pomiędzy:
prawym przedramieniem (R ) a lewym przedramieniem (L) - I odprowadzenie kończynowe
prawym przedramieniem (R ) i lewą golenią (F) - II odprowadzenie kończynowe
lewym przedramieniem (L) i lewą golenią (F) - III odprowadzenie kończynowe
Elektrody przystawione do kończyn zachowują pewne stałe przestrzenne położenie w stosunku do mięśnia sercowego. Elektrody, które spełniają tylko rolę przewodników elektrycznych, odbierają z różnych stron czynność bioelektryczną serca. Zapis EKG zarejestrowany z każdego z trzech odprowadzeń kończynowych jest inny, ponieważ stanowi wypadkową potencjałów elektrycznych serca występujących w przestrzeni pomiędzy tymi elektrodami.
Odprowadzenia jednobiegunowe:
Za ich pomocą rejestruje się różnice potencjałów pomiędzy elektrodą aktywną i elektrodą nieaktywną (jedna przystawiona do klatki piersiowej jest aktywna). Zapis EKG odbierany za pomocą odprowadzeń przedsercowych różni się od EKG odebranego za pomocą odprowadzeń kończynowych.
Stosuje się 6 jednobiegunowych odprowadzeń przedsercowych (Wilsona): V1, V2, V3, V4, V5, V6. Elektroda I przedsercowego odprowadzenia V1 jest przystawiona do skóry w czwartym międzyżebrzu po stronie prawej mostka, pozostałe po lewej stronie.
Odprowadzenia kończynowe jednobiegunowe
zawierając bezpośrednio dwie elektrody kończynowe, uzyskuje się z trzeciej elektrody aktywnej EKG o załamkach o wyższej amplitudzie. Mamy trzy odprowadzenia kończynowe EKG jednobiegunowe nasilone (Goldbergera):
aVR - elektroda aktywna na prawym przedramieniu
aVL - elektroda aktywna na lewym przedramieniu
aVF - elektroda aktywna na lewej goleni
Porównywalność zapisów EKG wymaga nie tylko stosowania typowych odprowadzeń, lecz również jednakowych warunków rejestracji. Aparaty - elektrokardiografy - rejestrują krzywą EKG na podstawie czasu 2 sekundy, prędkość przesuwu papieru 25 lub 50 mm/s i wzmocnieniu odpowiadającemu 1mV=10mm
11. Analiza krzywej EKG.
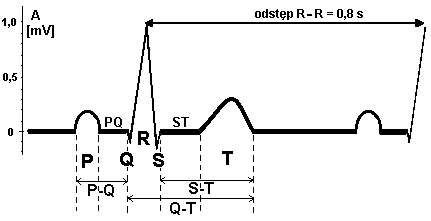
EKG zarejestrowany za pomocą II odprowadzenia kończynowego dwubiegunowego ma 5 załamków (P,Q,R,S,T).
Analiza krzywej EKG obejmuje:
odcinki - czas trwania linii izoelektrycznej między załamkami
załamki dodatnie i ujemne, ich amplitudę, czas trwania, częstotliwość występowania i kształt
odstępy - łączny czas trwania załamka i odcinka
-załamek P (100 ms) - czas przewodzenia depolaryzacji w mięśniu przedsionków
-odcinek PQ (50ms) - czas przejścia depolaryzacji przez węzeł przedsionkowo-komorowy i pęczek przedsionkowo-komorowy
-odstęp PQ (150 ms) - czas przewodzenia od węzła zatokowego do mięśnia komór
-zespół QRS (90 ms) - czas szerzenia się depolaryzacji w mięśniu komór
-odcinek ST (120 ms) - okres depolaryzacji mięśnia komór
-załamek T (120 ms) - szybka repolaryzacja komór
-odstęp ST (280 ms) - czas wolnej i szybkiej repolaryzacji mięśnia komór)
-odstęp QT (370 ms) potencjał czynnościowy mięśnia komór (depolaryzacja i repolaryzacja)
-odstęp RR (800 ms) - czas trwania jednego cyklu serca
Czas trwania poszczególnych załamków, odstępów i odcinków zależy od czasu trwania potencjałów czynnościowych mięśni serca.
Amplituda załamków zależy od oporu elektrycznego tkanek znajdujących się pomiędzy elektrodą i sercem, budowy ciała i położenia serca w klatce piersiowej.
Amplituda poszczególnych załamków w II odprowadzeniu:
-załamek P 0.1-0.25 mV
-zespół QRS 0.7-1.8 mV
-załamek T 0.26-0.6 mV
Nieprawidłowy kształt i amplituda załamków oraz nieprawidłowe wartości odnoszące się do odcinków i odstępów wskazują na zaburzenia w czynności bioelektrycznej mięśnia sercowego.
wydłużenie odcinka PQ - świadczy o zwolnieniu przewodzenia stanu czynnego przez układ przewodzący
brak załamka S, uniesienie odcinka ST ponad linię izoelektryczną lub inne zniekształcenia zespołu QRST mogą wskazywać na zaburzenia w krążeniu krwi w naczyniach wieńcowych serca na skutek np. zaczopowania odgałęzienia jednej z tętnic wieńcowych.
12. Wpływ układu współczulnego i amin katecholowych na serce.
Układ współczulny unerwia wszystkie części serca oddając dużo włókien do mięśni komór.
Pobudzenie włókien przywspółczulnych lub podanie noradrenaliny lub adrenaliny powoduje:
dodatni efekt chronotropowy (zwiększenie częstości rytmu zatokowego)
dodatni efekt dromotropowy (zwiększenie szybkości przewodzenia)
dodatnie działanie inotropowe (zwiększenie kurczliwości mięśni przedsionków i komór)
dodatni efekt batmotropowy (zwiększenie pobudliwości serca; w stanach patologicznych, oraz w przypadku bardzo silnego pobudzenia układów współczulnych aż do arytmii komorowej i migotania komór włącznie; adrenalina i NA nie mają wpływu na rozkurczowy próg pobudliwości, wywołana przez nie arytmia jest spowodowana pobudzeniem automatyzmu obwodowych (pozazatokowych) ośrodków)
13. Wpływ układu przywspółczulnego i acetylocholiny na serce.
Układ przywspółczulny unerwia głównie węzeł zatokowo-przedsionkowy i przedsionkowo-komorowy, w mniejszym stopniu mięśnie przedsionków, niemal wcale nie unerwia mięśni komór.
Przy pobudzeniu układu przywspółczulnego biegnącego do serca następuje uwalnianie acetylocholiny (Ach) z zakończeń włókien nerwu błędnego.
Działanie acetylocholiny na serce:
ujemne działanie chronotropowe (zwalnia lub całkowicie hamuje rytm zatokowy oraz rytm węzła przedsionkowo - komorowego)
ujemne działanie dromotropowe (zmniejszenie szybkości przewodzenia, aż do jego całkowitego zniesienia w strefie przedsionkowo-węzłowej)
ujemne działanie inotropowe (zmniejsza kurczliwość mięśni przedsionków; nie wpływa bezpośrednio na kurczliwość mięśnia komór, jednak hamuje inotropowy wpływ katecholamin)
brak wpływu na automatyzm komórek przewodzących, ale hamuje wpływ katecholamin na przebieg ich powolnej spoczynkowej depolaryzacji
Przy bardzo silnym pobudzeniu nerwów błędnych może dojść do całkowitego zatrzymania rytmu węzła zatokowo-przedsionkowego lub do całkowitego zablokowania przewodzenia stanu czynnego w połączeniach przedsionkowo-komorowych.
W obu tych przypadkach impulsy przestają być przekazywane do komór
→ komory przestaną się kurczyć
stan ten trwa nie dłużej niż 4-10 sekund, po czym w pewnym miejscu na przebiegu włókien Purkinjego, zwykle w części obwodowej pęczka przedsionkowo-komorowego, rozwinie się własny rytm (15-40 uderzeń/min). Zjawisko to nazywany ucieczką komór.
14. Przedsionkowy czynnik natriuretyczny (ANF)
Przedsionkowy czynnik natriuretyczny (ANF) to jeden z hormonów wpływających na czynność kanalików nerkowych. Prekursor ANF jest wytwarzany przez komórki mięśniowe przedsionków serca. Aktywne fragmenty prekursora są uwalniane do krwi po rozciągnięciu ścian przedsionków serca (przede wszystkim pod wpływem zwiększonej objętości krwi krążącej - TBV).
Rola ANP:
zwiększa wydalanie przez nerki jonów sodowych (oraz innych jonów) i wody, jednocześnie obniża ciśnienie tętnicze krwi
powoduje rozkurczanie mięśni gładkich naczyń krwionośnych (rozszerzanie naczyń)
hamuje działanie wazopresyny i układu hormonów: renina-angiotensyna-aldosteron
Efekt działania: zmniejszenie objętości płynów ustrojowych (krwi) i zniesienie nadmiernego rozciągnięcia przedsionków.
Wyszukiwarka
Podobne podstrony:
Fizjologia krwi notatka, ~FARMACJA, I rok, anatomia - fizjologia, fizjo
KREW, ~FARMACJA, I rok, anatomia - fizjologia, fizjo
2uklad krazenia, ~FARMACJA, I rok, anatomia - fizjologia, fizjo
Anatomia i fizjologia uklad nerwowy 3 nots, ~FARMACJA, I rok, anatomia - fizjologia, fizjo
uk nerwowy cz 2 ściąga, ~FARMACJA, I rok, anatomia - fizjologia, fizjo
Układ krążenia notatka, ~FARMACJA, I rok, anatomia - fizjologia, fizjo
skrypt skryptu do fizjo egzamin )), ~FARMACJA, I rok, anatomia - fizjologia, fizjo
Wielka ściąga uk nerwowy III, ~FARMACJA, I rok, anatomia - fizjologia, fizjo
Wielka Ściąga na Anatomię i Fizjologię uk. nerwowy cz 1, ~FARMACJA, I rok, anatomia - fizjologia, fi
Fizjologia serca notatka, ~FARMACJA, I rok, anatomia - fizjologia, fizjo
ANATOMIA ZALICZONKO - ściąga, ~FARMACJA, I rok, anatomia - fizjologia, fizjo
Kolokwium I sem - lekarski(2), I rok, Anatomia, cxrtsjxcgvhbjnkmugjyfghkjl, Anatomia egzamin, Semest
sciaga1-mikroby (podloza, farma 2 rok, mikrobiologia, fizjologia bakterii
anatomia ustne 1, Medycyna ŚUM, Rok 1, Anatomia, Testy kolokwia egzaminy
anatomka, Położnictwo - studia - Łódź, I rok, Anatomia, Wejściówki, kolokwia, egzamin
układ stawowy i układ mięśniowy, Pielęgniarstwo rok I i inne, Anatomia i Fizjologia
więcej podobnych podstron