
Wykonała Ewelina Ampuła

Służą jako wydajne źródło energii zarówno bezpośrednio, jak i potencjalnie, gdy
odłożone są jako materiał zapasowy w tkance tłuszczowej podskórnej
Tłuszcze w tkance tłuszczowej podskórnej chronią przed nadmiernym wydzielaniem
ciepła i pozwalają na adoptowanie się w niskiej temperaturze
Wewnątrz organizmu utrzymują narządy w stałym położeniu i służą do mechanicznej i
termicznej izolacji.
W tłuszczach zawartych w naturalnych pokarmach znajdują się witaminy rozpuszczalne
w tłuszczach ( A, D, E, K ) oraz niezbędne nienasycone kwasy tłuszczowe
Pełnią funkcję budulcową, są składnikami błon komórkowych
Połączenia tłuszczów z białkami ( lipoproteiny) służą jako środek transportu lipidów w
osoczu.
Steroidy i niektóre fosfolipidy służą jako hormony i drugie przekaźniki

Tłuszcze wchłonięte z pokarmów oraz syntetyzowane w wątrobie i
tkance tłuszczowej muszą być transportowane między różnymi
tkankami i narządami, aby mogły być zużywane i magazynowane.
Długołańcuchowe kwasy tłuszczowe wiążą się z albuminami,
krótkołańcuchowe rozpuszczają się w osoczu, natomiast inne
lipidy transportowane są w kompleksach lipoproteinowych,
które w osoczu krwi występują w kilku rodzajach,
różniących się wielkością i składem:
Chylomikrony, powstające z wchłanianych w jelicie
triacylogliceroli
Lipoproteiny o bardzo małej gęstości (VLDL), formowane
głównie w wątrobie, pełnią funkcję eksportera triacylogliceroli
z tego narządu
Lipoproteiny małej gęstości (LDL), będące przedstawicielem
końcowych produktów katabolizmu VLDL
Lipoproteiny o dużej gęstości (HDL),biorące udział w
metabolizmie VLDL i chylomikronów, a także w przemianie
cholesterolu
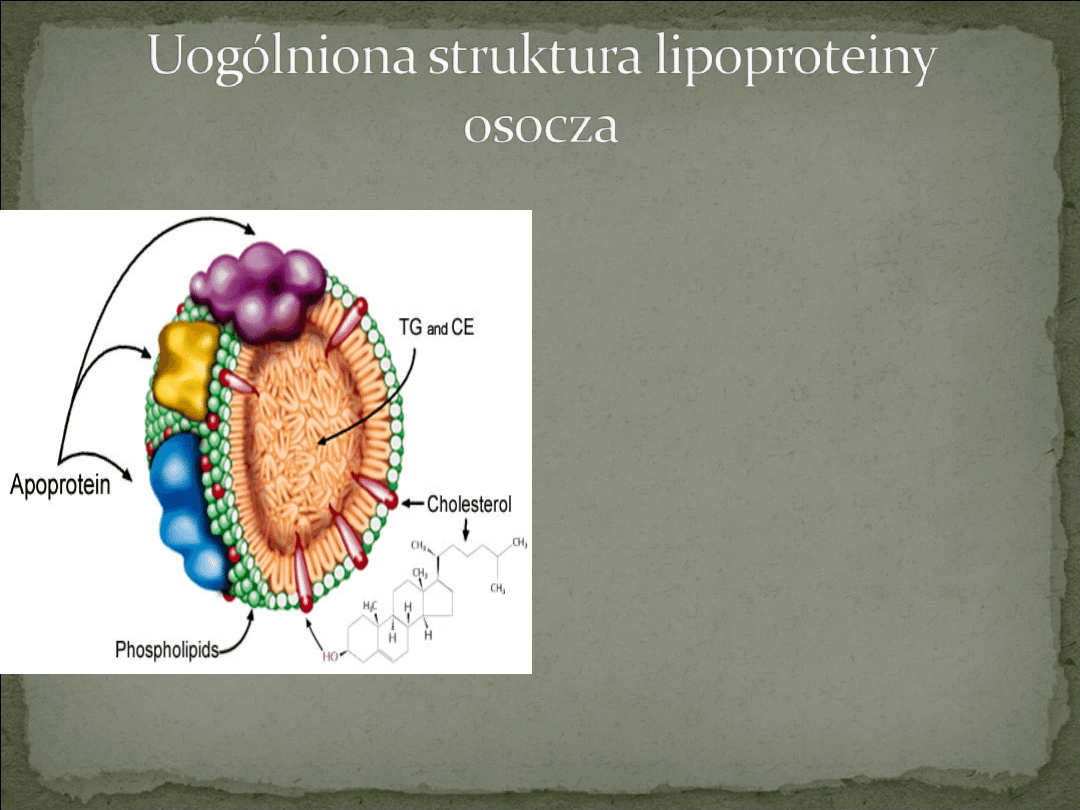
Lipoproteiny są kulistymi agregatami, w
których skład wchodzą lipidy i apoproteiny.
Typowa lipoproteina składa się z rdzenia
lipidowego, zawierającego głównie
niepolarne triacyloglicerole i estry
cholesterolu, otoczonego warstwą
powierzchniową złożoną z amfipatycznych
cząsteczek fosfolipidów i cholesterolu
Białkowa część lipoprotein stanowi od ok.
70% masy niektórych HDL do zaledwie 1%
masy chylomikronów. Rozmieszczenie
apolipoprotein jest charakterystyczne dla
danej danej lipoproteiny
Apolipoproteiny spełniają kilka funkcji:
A.
tworzą część struktury lipoprotein np. apo
B
B.
Są kofaktorami enzymów np. apo C-II
lipazy lipoproteinowej
C.
Działają jako ligandy z receptorami
lipoprotein w tkankach np. apoB-100 i
apoE w interakcji z receptorami LDL

Chylomikrony uczestniczą w transporcie triacylogliceroli z jelita do
tkanek. Na powierzchni śródbłonka naczyń włosowatych tkanek
pozawątrobowych (głównie mięśni i tkanki tłuszczowej), lipaza
lipoproteinowa hydrolizuje większość triacylogliceroli. Uwolnione
kwasy tłuszczowe i glicerol wchłaniane są do komórek, a
chylomikrony przechodzą stopniowo w chylomikrony resztkowe
usuwane z krwi przez wątrobę.
VLDL, IDL, LDL są ze sobą ściśle spokrewnione. Wytwarzane w
wątrobie VLDL transportują triacyloglicerole, cholesterol i
fosfolipidy do innych tkanek. Pod wpływem lipazy lipoproteinowej
przechodzą stopniowo w IDL i LDL Komórki mające zapotrzebowanie
na cholesterol wiążą LDL przez oddziaływanie swoich receptorów
LDL z apoB-100 i wchłaniają całe cząsteczki na drodze endocytozy
HDL również pochodzą z wątroby. Sprowadzają do niej z powrotem z
tkanek cholesterol. Podczas transportu cholesterol ulega acylowaniu
przez LCAT.

Jadalne tłuszcze zwierzęce i oleje roślinne są stosunkowo
dobrze przyswajalne przez człowieka. Większość z nich
przedostaje się z żołądka do dwunastnicy w postaci
niezmienionej, ponieważ ich trawienie w żołądku jest
raczej niewielkie. Tak więc proces hydrolizy i wchłaniania
lipidów przebiega przede wszystkim w dwunastnicy i
początkowym odcinku jelita czczego, gdzie ulegają niemal
całkowitej resorpcji (wchłonięciu).
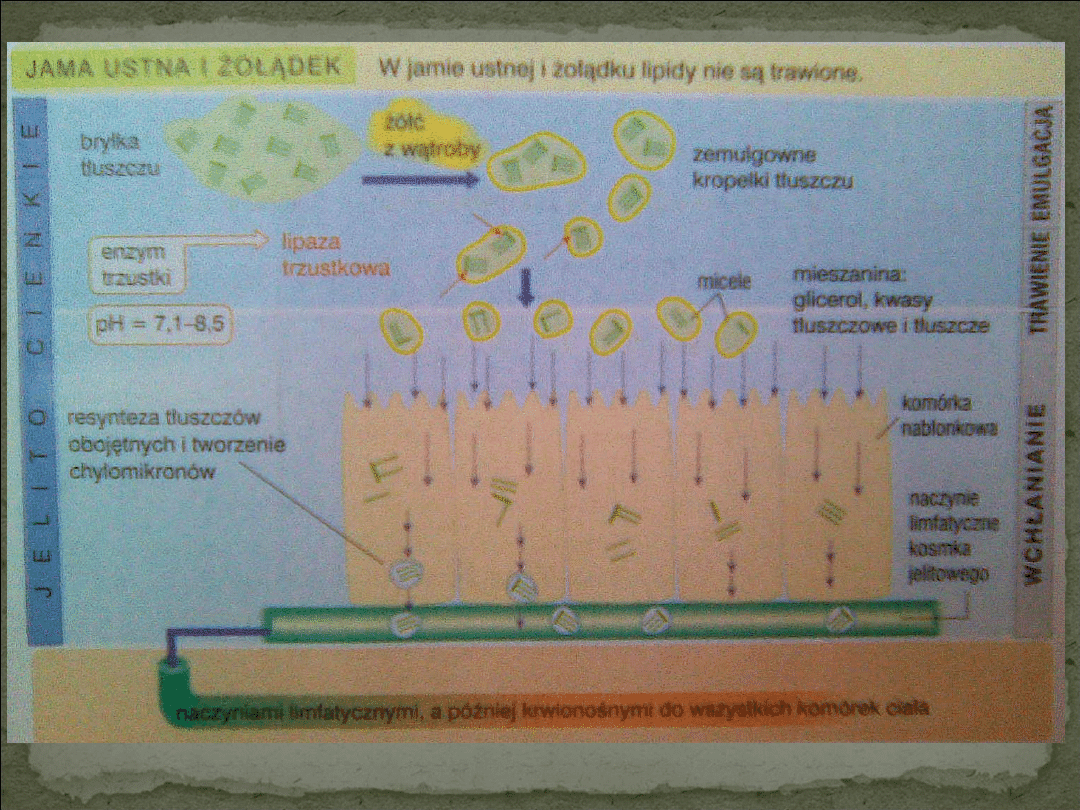
Trawienie lipidów jest poprzedzone ich rozdrobnieniem
dokonującym się z udziałem soli kwasów żółciowych. Sole
te wykazują zdolność do zmniejszania napięcia
powierzchniowego, dzięki czemu lipidy
ulegają zawieszeniu, czyli tworzą emulsję.
Następuje wówczas dyspersja lipidów do kropelek.

pewną aktywność przejawia u ludzi lipaza wydzielana przez ślinianki języka
(lipaza językowa ślinowa). Uwolnione pod wpływem lipazy ślinowej
krótkołańcuchowe kwasy tłuszczowe przenikają przez ścianę żołądka
bezpośrednio do żyły wrotnej, natomiast kwasy o długich łańcuchach
rozpuszczają się w kroplach tłuszczowych zawieszonych w treści żołądkowej,
skąd przedostają się do dwunastnicy.
Trawienie lipidów pokarmowych następuje przede wszystkim w jelicie
cienkim pod wpływem lipazy trzustkowej, która katalizuje hydrolizę prawie
wszystkich triacylogliceroli pokarmowych. Enzym ten działa na granicy fazy
wodnej i tłuszczowej kropelek tłuszczowych w obecności soli kwasów
żółciowych, koliazy, fosfolipidów i fosfolipazy A2, również występującej w
soku trzustkowym. W obecności soli kwasów żółciowych zmniejsza się
aktywność lipazy, ale dzięki kolipazie lipaza odzyskuje pełną zdolność
hydrolizy triacylogliceroli.
Oprócz enzymów w procesie trawienia tłuszczów niezbędne są sole kwasów
żółciowych i wodorowęglany. Pierwsze emulgują tłuszcze i dzięki swoim
micelom rozpuszczają produkty lipolizy w fazie wodnej w jelitach, drugie
utrzymują optymalne pH dla działania enzymów lipolitycznych.
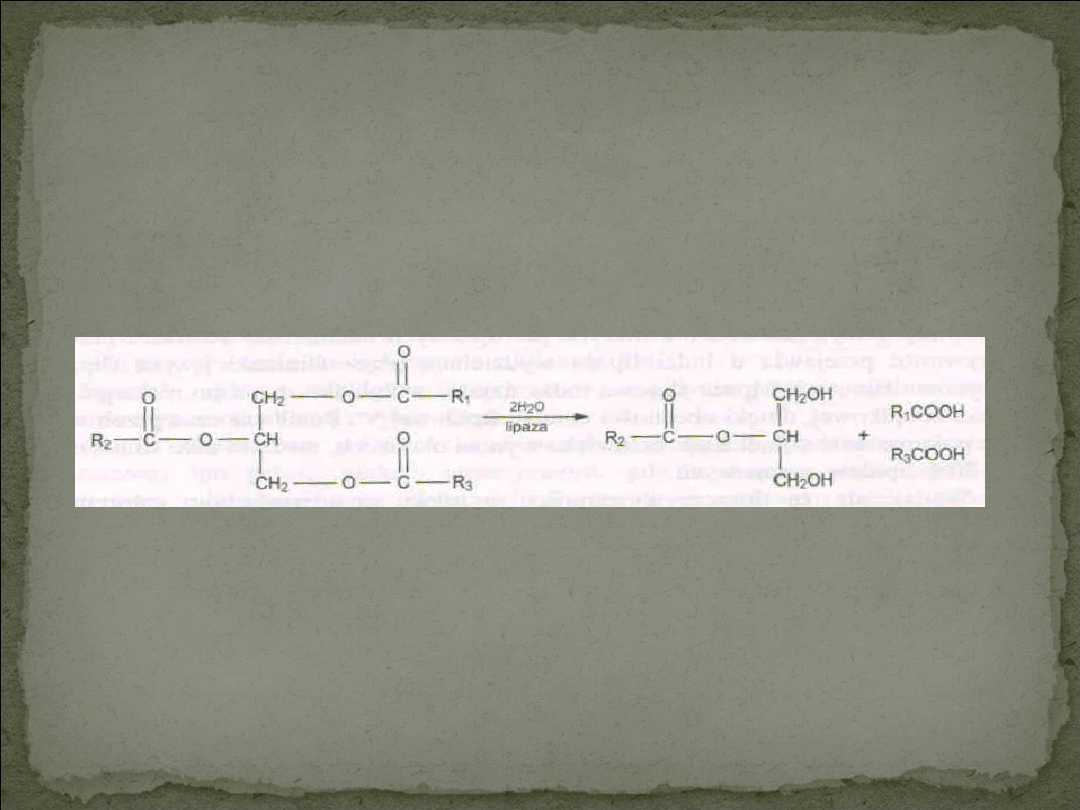
Micele utworzone z kwasów żółciowych pełnią funkcję przenośnika
nierozpuszczalnych w wodzie produktów lipolizy z miejsca ich powstania na
powierzchni kropelek tłuszczowych na powierzchnię błony śluzowej jelita,
skąd w wyniku zwykłej dyfuzji dostają się enterocytów, czyli komórek
chłonnych kosmków jelitowych.
Liapaza trzustkowa ssaków katalizuje hydrolizę wiązań estrowych kwasów
tłuszczowych z pierwszorzędowymi grupami hydroksylowymi glicerolu.
Produktem reakcji są kwasy tłuszczowe i 2-monoacyloglicerol.
Około 75% lipidów pokarmowych ulega resorpcji w postaci 2-
monoacyloglicerolu i kwasów tłuszczowych, reszta zostaje całkowicie
rozłożona przez lipazę 2-monoacyloglicerolową do glicerolu i kwasów
tłuszczowych.

Produkty lipolizy, bezpośrednio po ich wniknięciu
do enterocytów podlegają resyntezie do
triacylogliceroli, które po przyłączeniu innych
lipidów i białka tworzą duże kompleksy zwane
chylomikronami.
Chylomikrony jako specyficzne lipoproteiny, są
następnie transportowane z enterocytów do
układu chłonnego i dalej wraz z chłonką za
pośrednictwem przewodu piersiowego dostają się
do ogólnego krwioobiegu.
W krwioobiegu chylomikrony ulegają bardzo
szybko rozkładowi. Biologiczny okres półtrwania
tych cząsteczek wynosi mniej niż 1h.
Triacyloglicerole chylomikronów są hydrolizowane
przez lipazę lipoproteinową.

Triacyloglicerole są głównymi lipidami tłuszczu zapasowego organizmu i
tłuszczu zawartego w pokarmach.
Glicerol z kwasami tłuszczowymi może tworzyć mono-, di- lub triestry
zwane odpowiednio: triacyloglicerol, diacyloglicerol, monoacyloglicerol.
Triacyloglicerole są estrami alkoholu trójwodorotlenowego–glicerolu i trzech
kwasów tłuszczowych
Trzy reszty acylowe cząsteczki triacyloglicerolu mogą się różnić długością
swych łańcuchów oraz liczbą podwójnych wiązań, czego konsekwencją jest
duża liczba możliwych kombinacji pojedynczych cząsteczek
Nienasycone kwasy tłuszczowe przeważnie znajdują się przy środkowym
atomie węgla glicerolu.

Fosfoglicerole syntetyzowane są albo z 1,2-
diacyloglicerolu albo z fosfatydanu np.
fosfatydyloinozytol, który drogą kolejnych drogą
kolejnych fosforylacji przekształcany jest w
fosfatydyloinozytolo-4,5-bisfosforan.
Fosfoglicerole są głównymi lipidowymi składnikami błon
komórkowych
Dipalmitoilofosfatydylocholina (syntetyzowana krótko
przed porodem) jest głównym składnikiem surfaktantu
płucnego, którego brak u wcześniaków jest przyczyną
zespołu niewydolności oddechowej (RDS) noworodków.
Fosfolipidy inozytolowe w błonie komórkowej są
prekursorami wtórnych przekaźników działania
hormonów –DAG (diacyloglicerol–aktywuje kinazę białek
C) i IP3 (inozytolo-1,4,5-trifosforan–otwiera kanały
Ca2+)
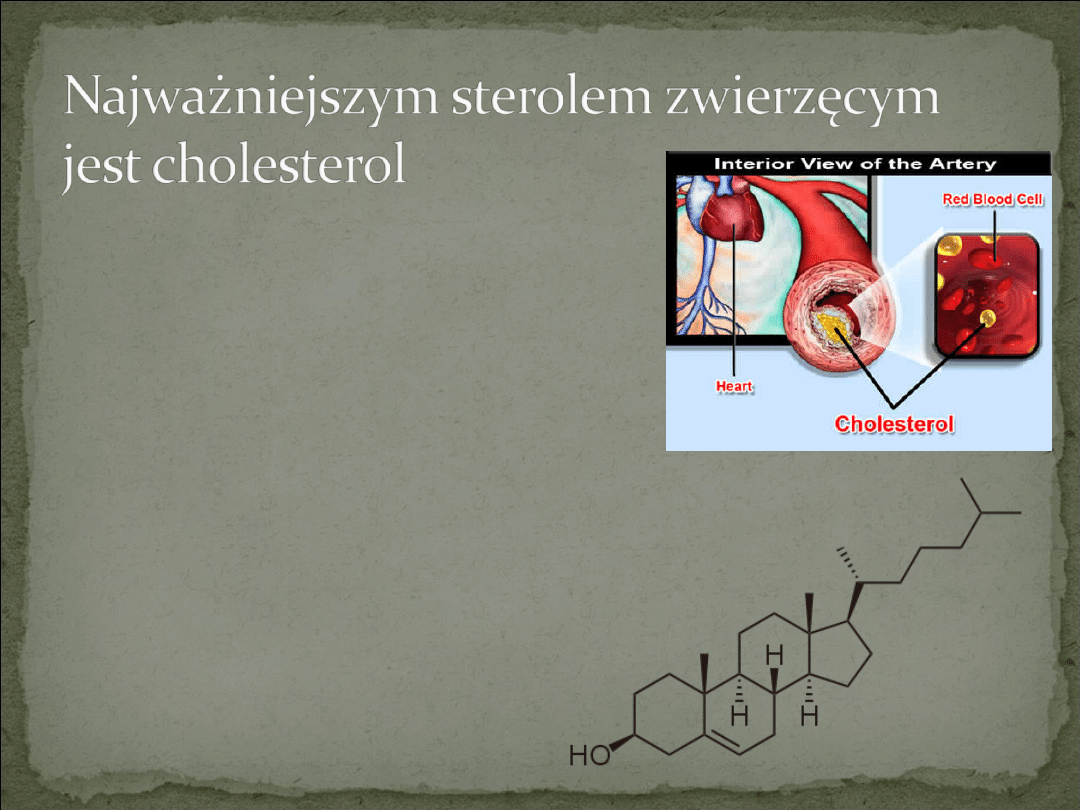
Cholesterol występuje we
wszystkich tkankach oraz w
lipoproteinach osocza jako wolny
cholesterol albo, w połączeniu z
długołańcuchowymi kwasami
tłuszczowymi, jako estry cholesterolu.
Syntetyzowany jest z acetylo-
CoA(wszystkie 27 atomów węgla
cholesterolu z niego pochodzi) i
ostatecznie wydalany z organizmu z
żółcią jako cholesterol lub sole kwasów
żółciowych.
Cholesterol jest typowym produktem
metabolizmu zwierzęcego, więc
znajduje się w pokarmach
pochodzenia zwierzęcego np. żółtkach
jaj, wątrobie, mięsie.

Odgrywa główną role w transporcie i metabolizmie
lipidów:
Ułatwia trawienie i wchłanianie lipidów z przewodu
pokarmowego, dlatego że wytwarza żółć zawierającą
cholesterol i sole kwasów żółciowych syntetyzowane w
wątrobie
Ma aktywne układy enzymatyczne niezbędne do
syntetyzowania i utleniania kwasów tłuszczowych,
syntetyzowania triacylogliceroli, fosfolipidów i
cholesterolu
Syntetyzuje lipoproteiny osocza
Przekształca kwasy tłuszczowe w ciała ketonowe
Odgrywa integrującą rolę w przemianie lipoprotein
osocza

Tkanka tłuszczowa może wytworzyć warstwę izolującą
lub pełnić funkcje amortyzatora, gdy występuje
pomiędzy wewnętrznymi narządami jamy brzusznej.
Szczególnym rodzajem tkanki jest tzw.
Brunatna tkanka tłuszczowa, rozmieszczona pod
skórą (w okolicy grzbietu) lub w sąsiedztwie
niektórych narządów. Zadaniem tej tkanki jest
produkcja znacznej ilości ciepła co chroni młody
organizm przed znacznymi nawet spadkami
temperatury. Pozwala to przetrwać organizmom
niekorzystne warunki. Wykazano, że tkanka ta jest
aktywna u ludzi zdrowych, natomiast u ludzi otyłych
ilość tej tkanki jest znacznie mniejsza bądź jej brak.
Ten fakt tłumaczy dlaczego niektóre osoby
spożywają stosunkowo dużo pokarmu, a nie są otyłe.

Insulina ––hamuje uwalnianie wolnych
kwasów tłuszczowych z tkanki tłuszczowej,
czego następstwem jest zmniejszenie
stężenia WKT w osoczu. Wzmaga lipogenezę,
syntezę acylogliceroli i utlenianie glukozy do
CO2 w szlaku pentozofosforanowym
Glukagon, adrenalina, noradrenalina,
hormony tarczycy–przyspieszają uwalnianie
wolnych kwasów tłuszczowych z tkanki
tłuszczowej dzięki temu, że zwiększają
szybkość lipolizy zapasów triacylogliceroli.
Prowadzi to do zwiększenia stężenia WKT w
osoczu.

„Biochemia kręgowców” PWN 2007
„Biochemia Harpera” PZWL
Document Outline
- Slide 1
- Slide 2
- Slide 3
- Slide 4
- Slide 5
- Slide 6
- Slide 7
- Slide 8
- Slide 9
- Slide 10
- Slide 11
- Slide 12
- Slide 13
- Slide 14
- Slide 15
- Slide 16
- Slide 17
- Slide 18
Wyszukiwarka
Podobne podstrony:
PLAN STRATEGICZNY LOKALU pod lip±, Studia
LIP 6000H
LIP 7000F
Ca,cuk,AK,RR,Miaż,Lip w I odpowiedzi
LIP 2000H
Krajowy system informacji o terenie [12 lip 2001]
LIP 1020H
LIP 1010H
Lip liner
LIP 5000H
Lip?lm
Lip augmentation
Testy z komentarzami do odpowiedzi czerw lip 92
cw lip 23 03 10 dyslipidemie diagnoza jh
Lip gloss
PLAN STRATEGICZNY LOKALU pod lip±, Studia
LIP 6000H
LIP 7000F
Seinfeld 506 The Lip Reader
więcej podobnych podstron