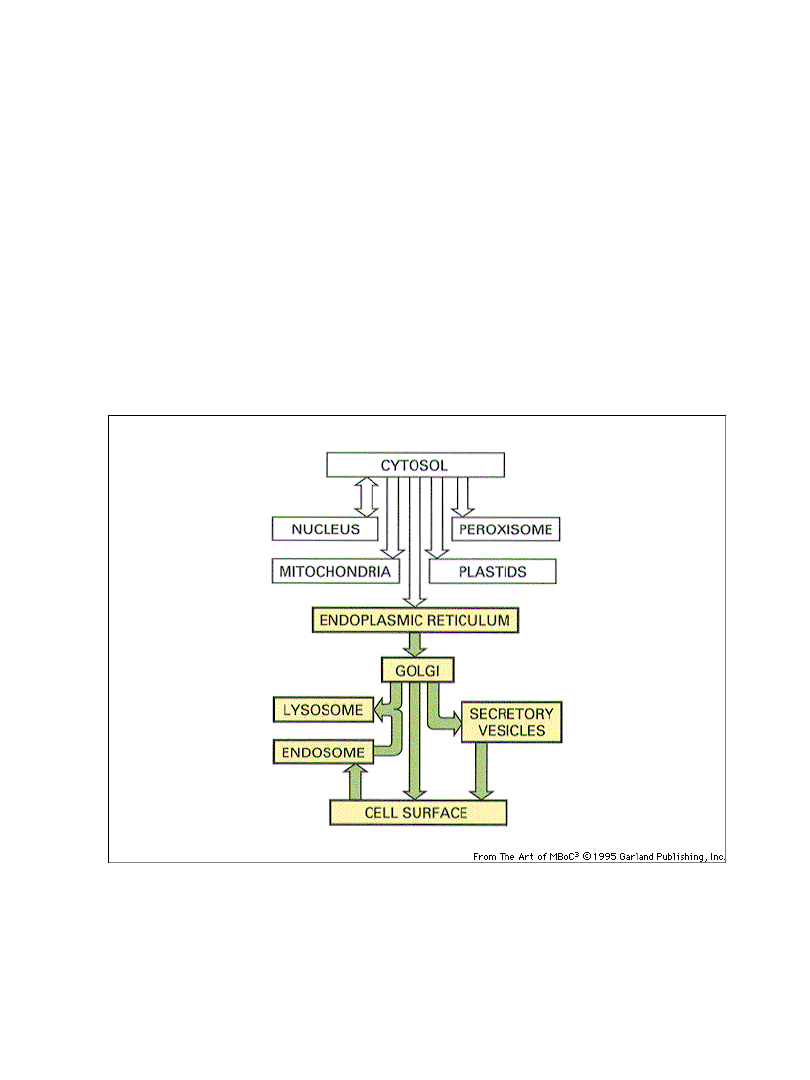
Transport wewnątrzkomórkowy
Transport wewnątrzkomórkowy
pomiędzy cytoplazmą i jądrem
pomiędzy cytoplazmą i jądrem
do/z organelli błonowych
do/z organelli błonowych
Rodzaje błon i organelli błonowych
-błona cytoplazmatyczna
-błona jądrowa
-siateczka śródplazmatyczna (ER) – szorstkie i gładkie
-aparat Golgiego
-lizosomy
-peroksysomy
-endosomy
-inne pęcherzyki
los białek po syntezie w rybosomach zależy od obecności sygnałów
lokalizacyjnych (sortujących) lub ich braku
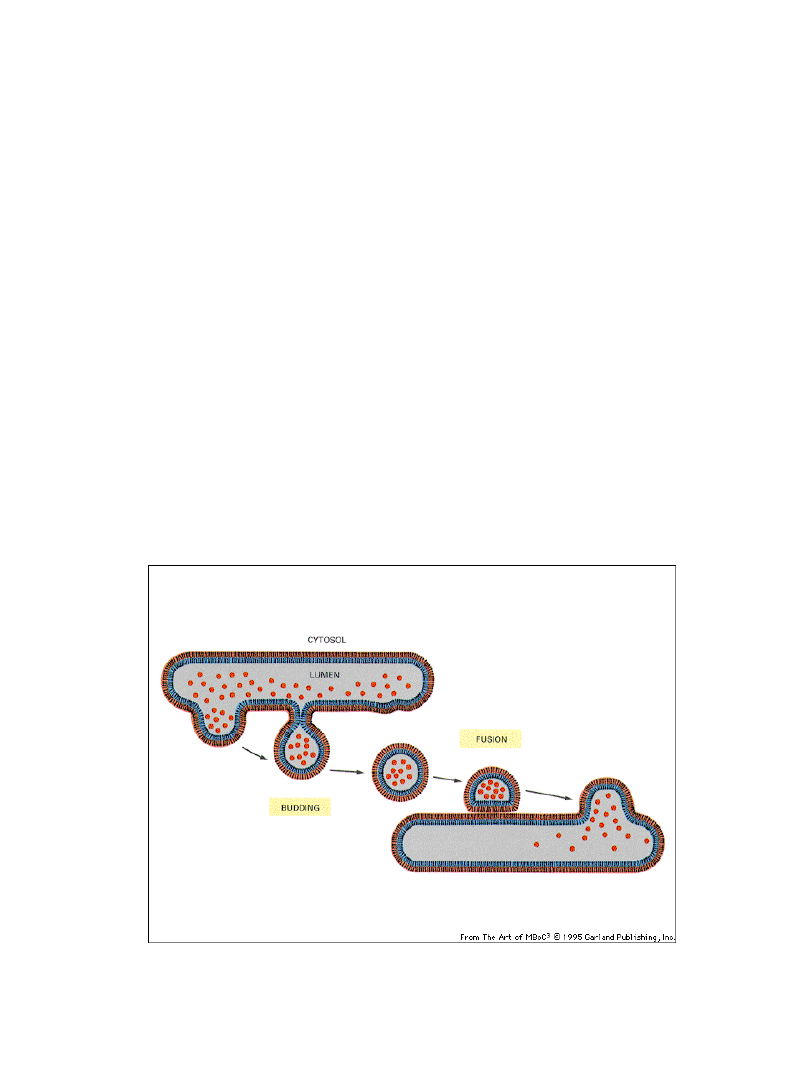
transport m.in. białek związany jest transportem pęcherzykowym
lub bezpośrednim przenoszeniem przez błony (translokacja) z udziałem
wyspecjalizowanych białek transportowych
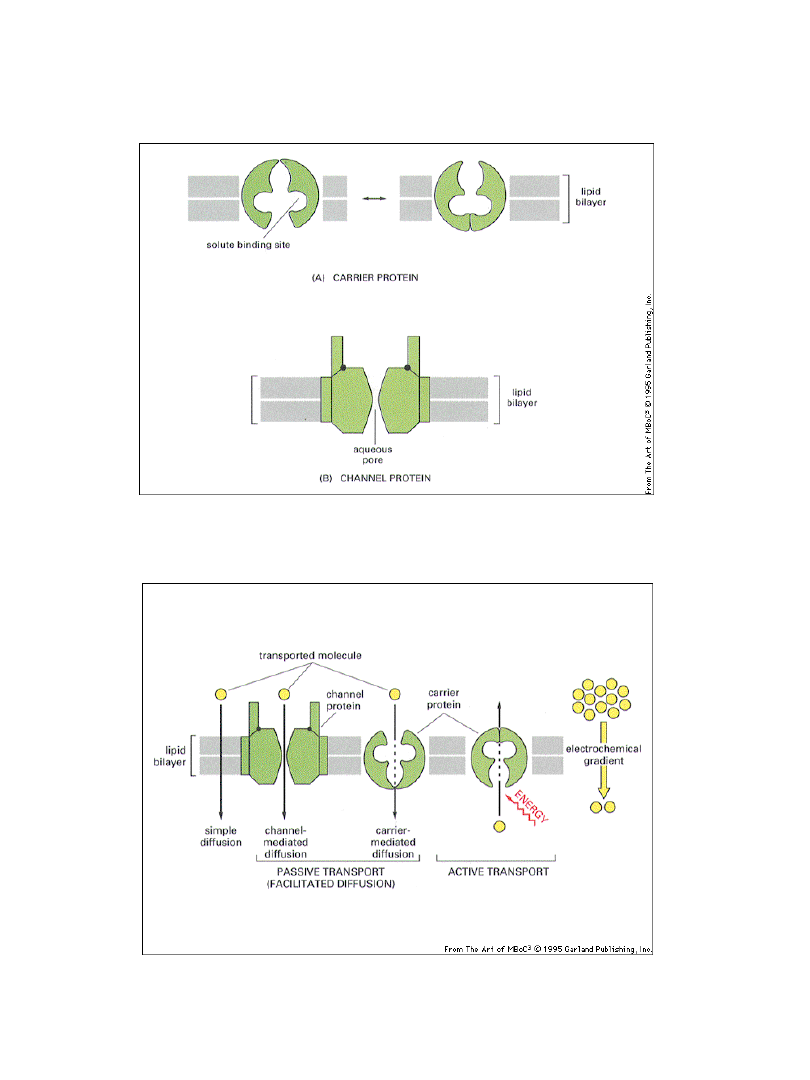
Klasy białek transportowych w błonach
Transport pasywny a aktywny
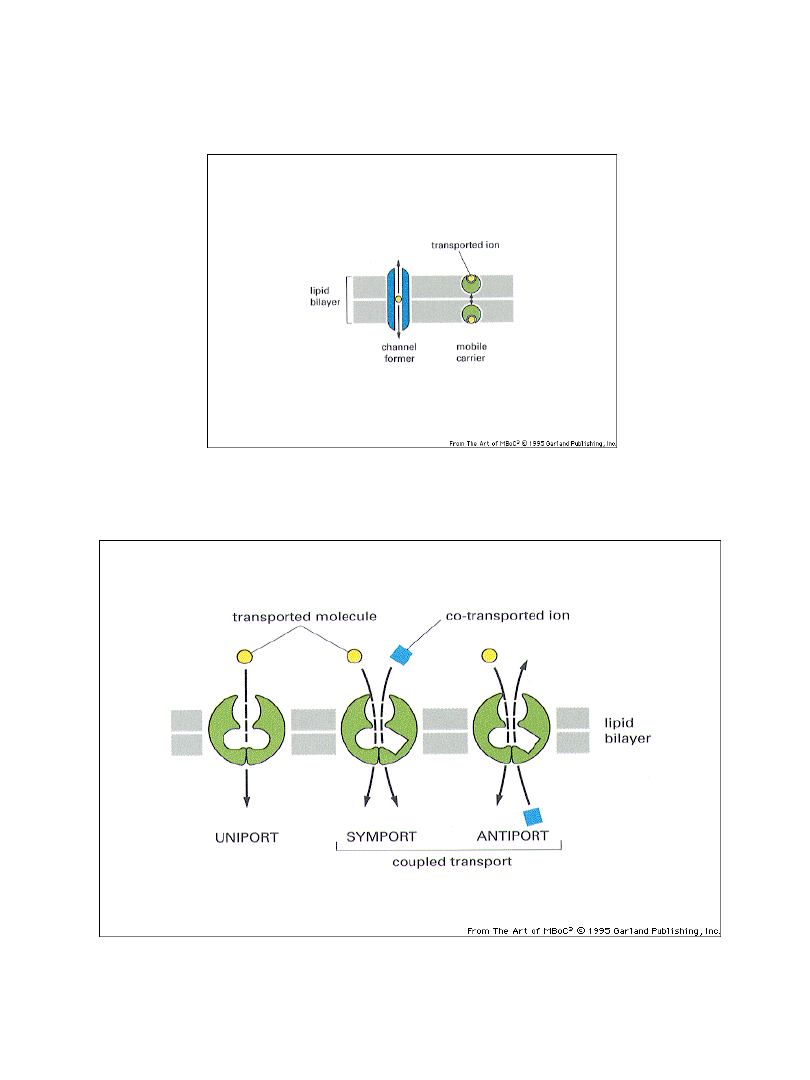
Transport błonowy można badać za pomocą jonoforów
Typy transportu błonowego
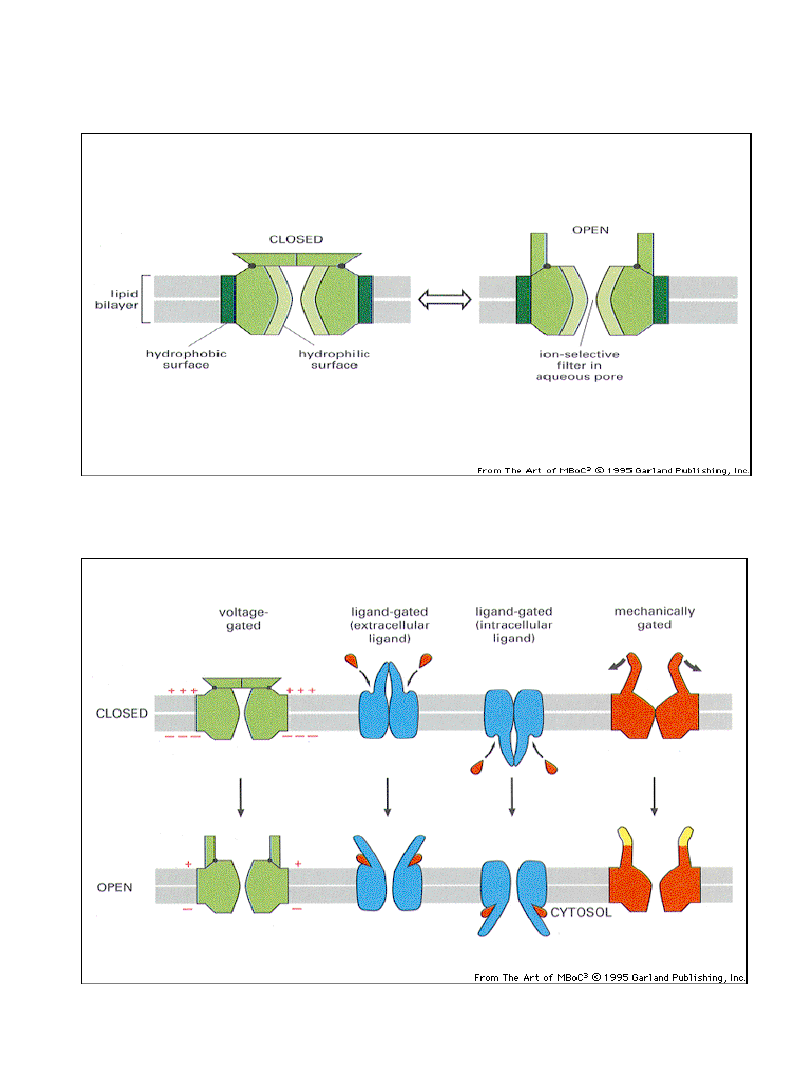
Schemat budowy typowych kanałów jonowych
Sposoby zamykania/otwierania kanałów jonowych
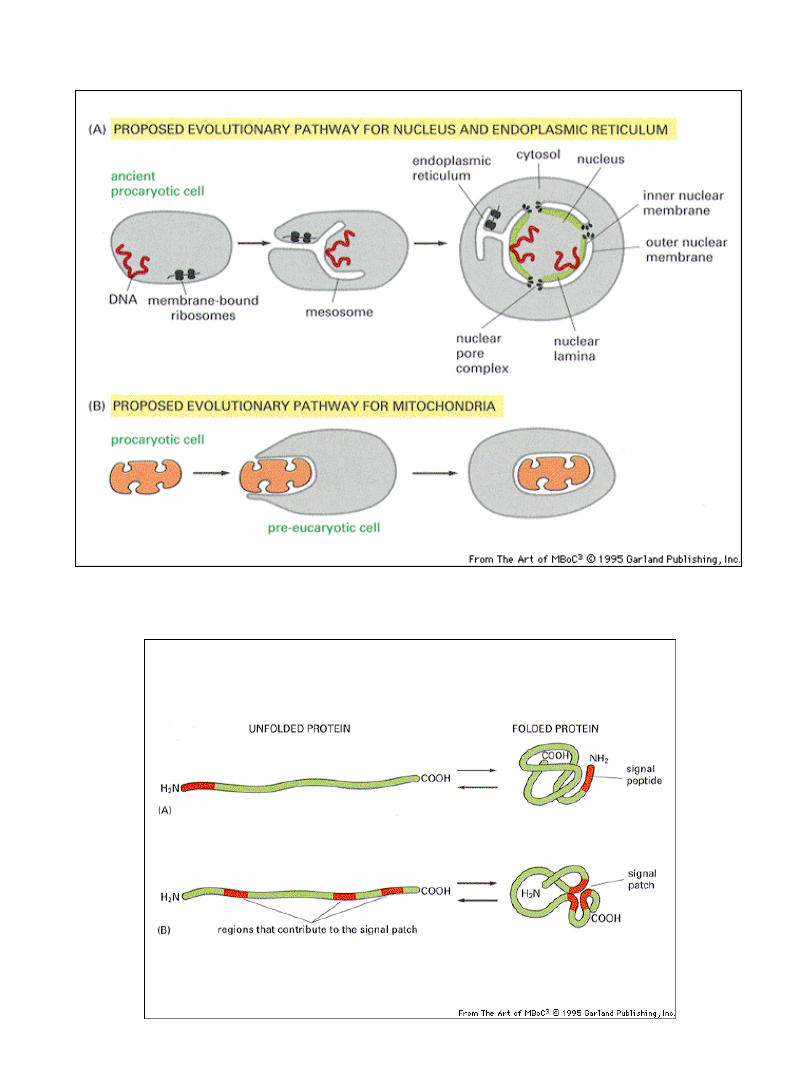
Hipoteza nt pochodzenia organelli błonowych
Sygnał i wzór sortujący
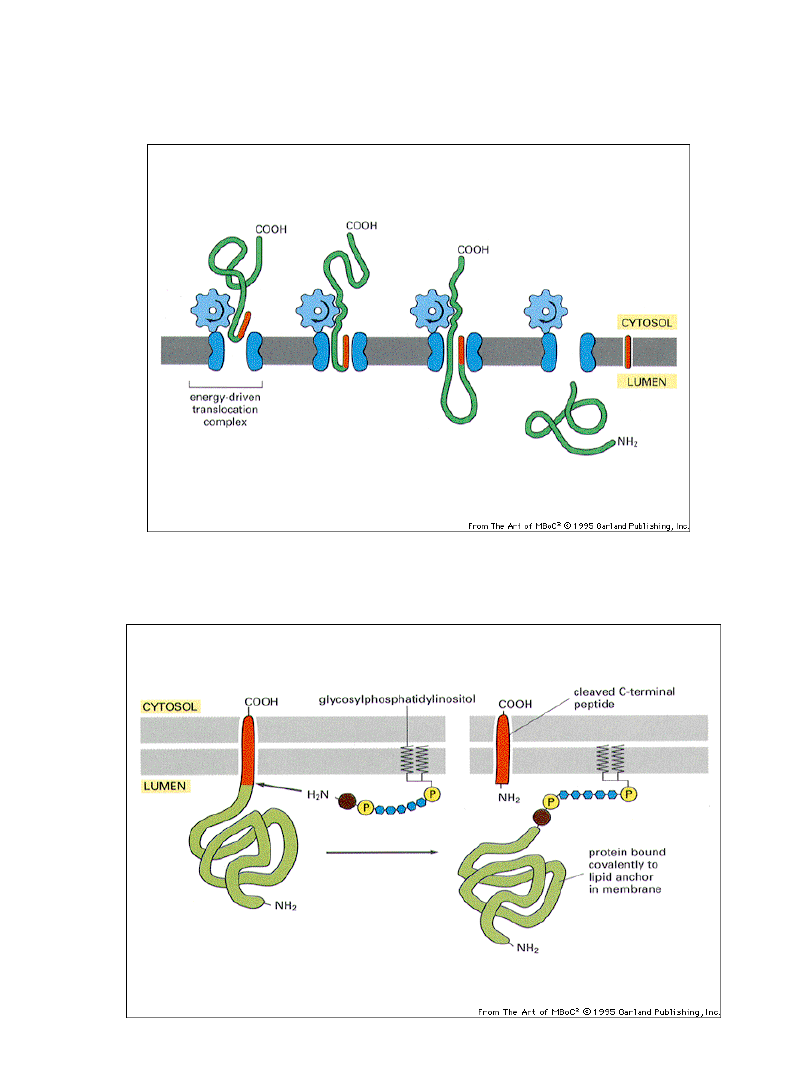
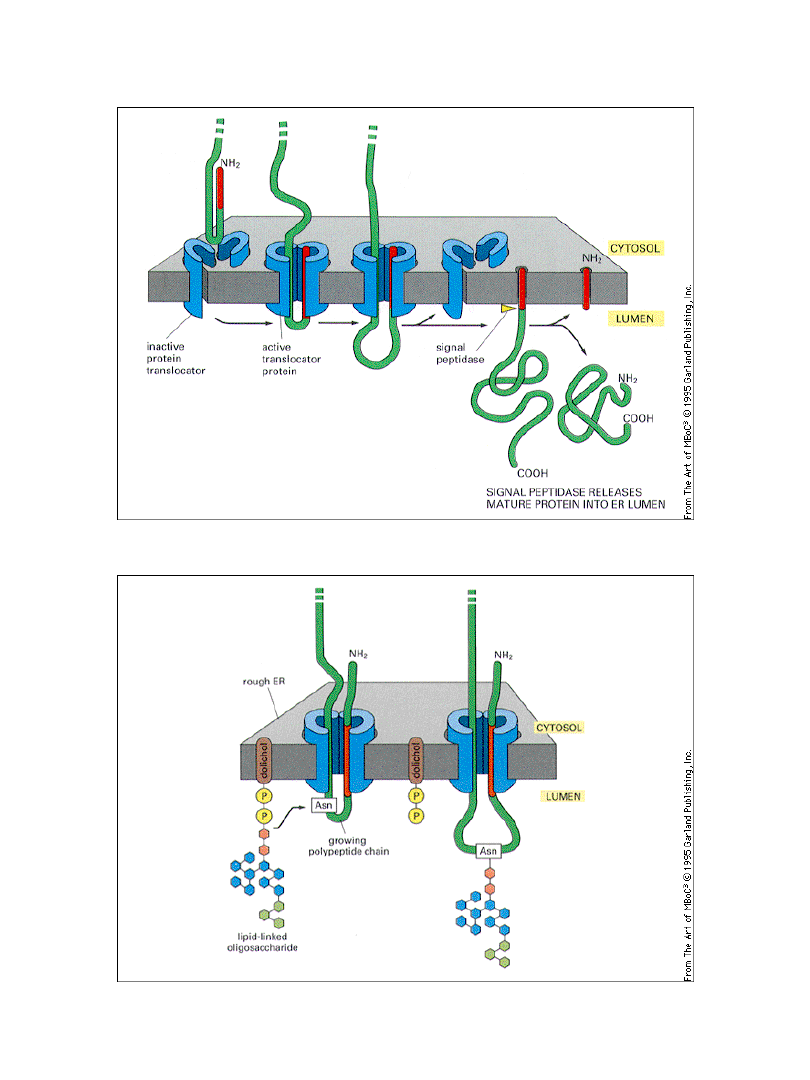
Przeniesienie niektórych białek przez błony
wymaga energii i translokatora
Przyłączanie ”kotwicy” glikozylofosfatydyloinozytolowej
(immobilizacja chemiczna)
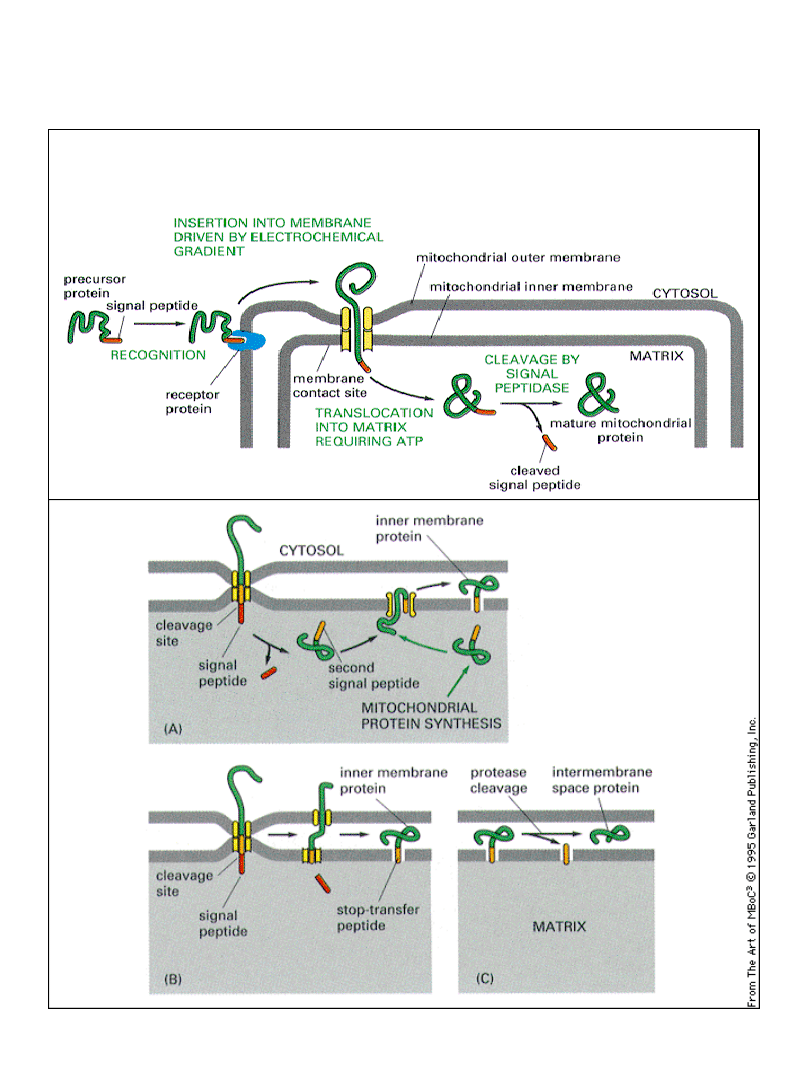
Import białek do matriks mitochondrium
lub przestrzeni międzybłonowej
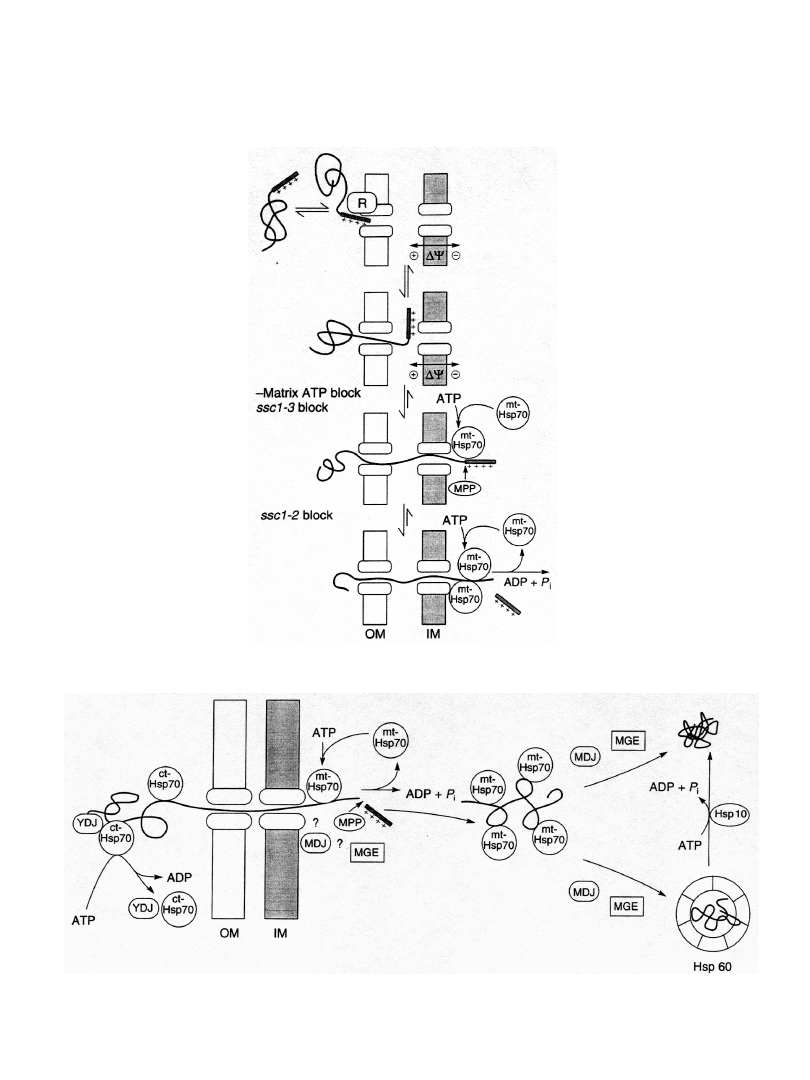
Transport białek przez błony
mitochondrialne
udział potencjału
membranowego
po przejściu przez błony białka muszą być ponownie
zwinięte
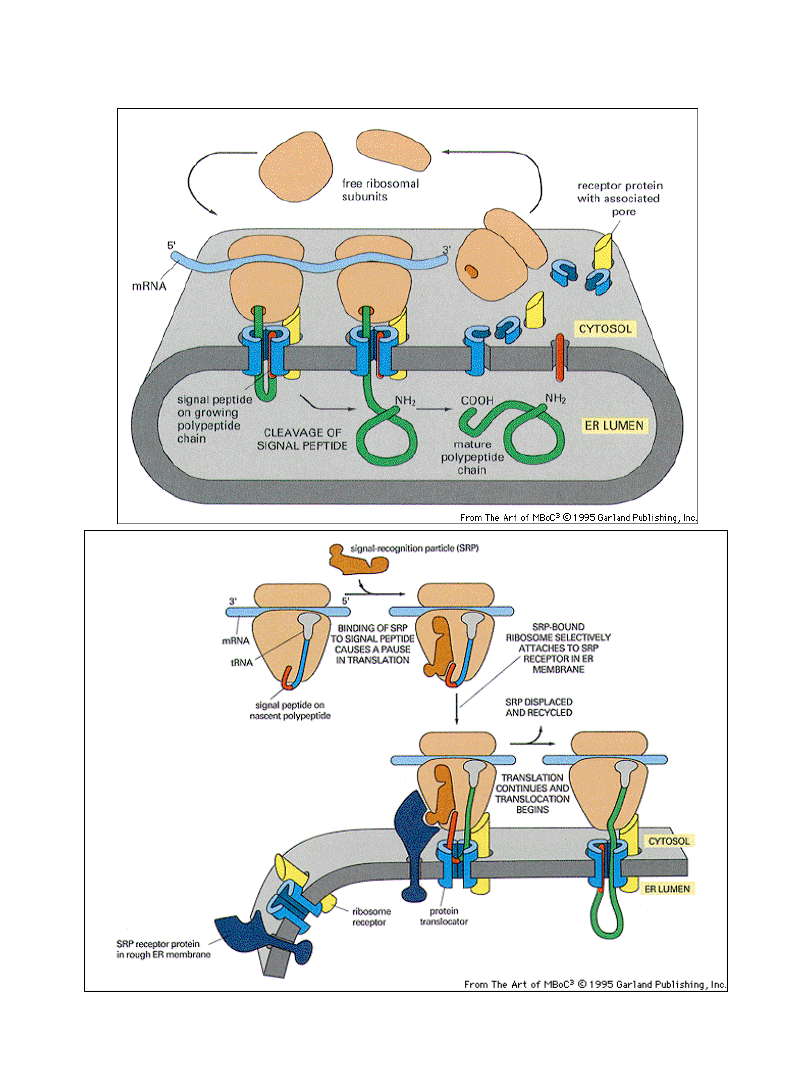
Schemat transportu białek do wnętrza ER
Translokacja białka przez błonę ER
Glikozylacja białek w szorstkim ER
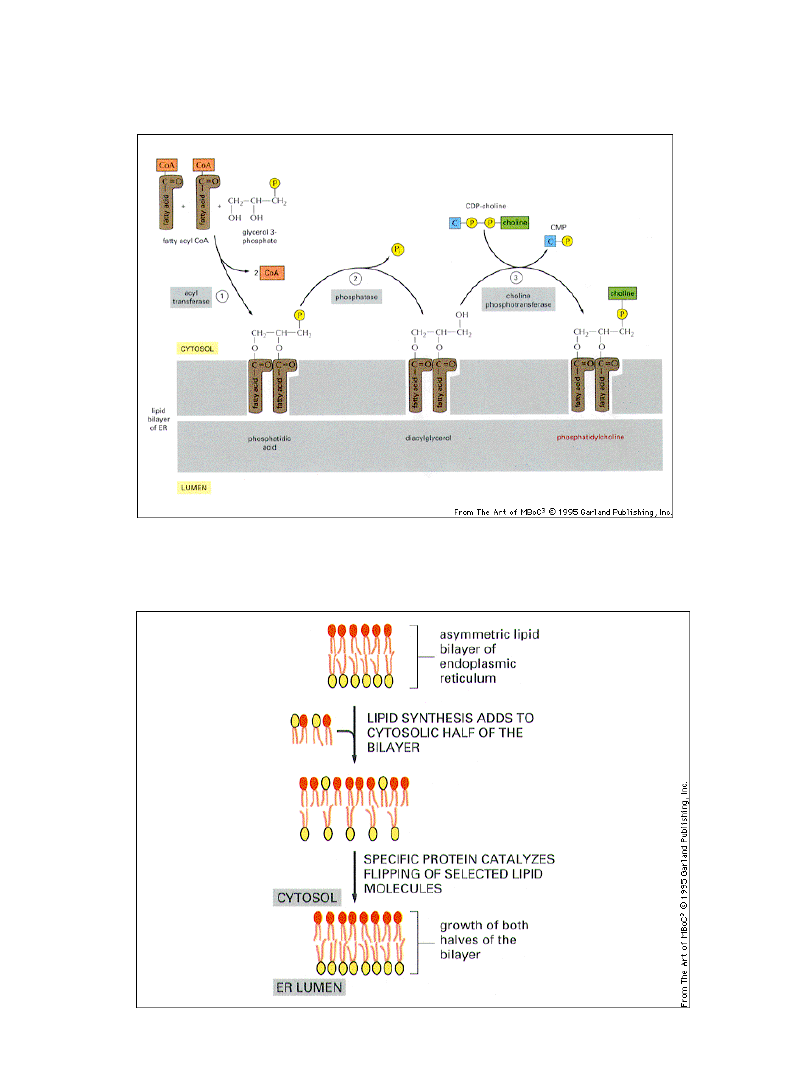
Większość składników błonowych i błon powstaje w ER
Prowadzi to do niesymetrii w rozkładzie fosfolipidów
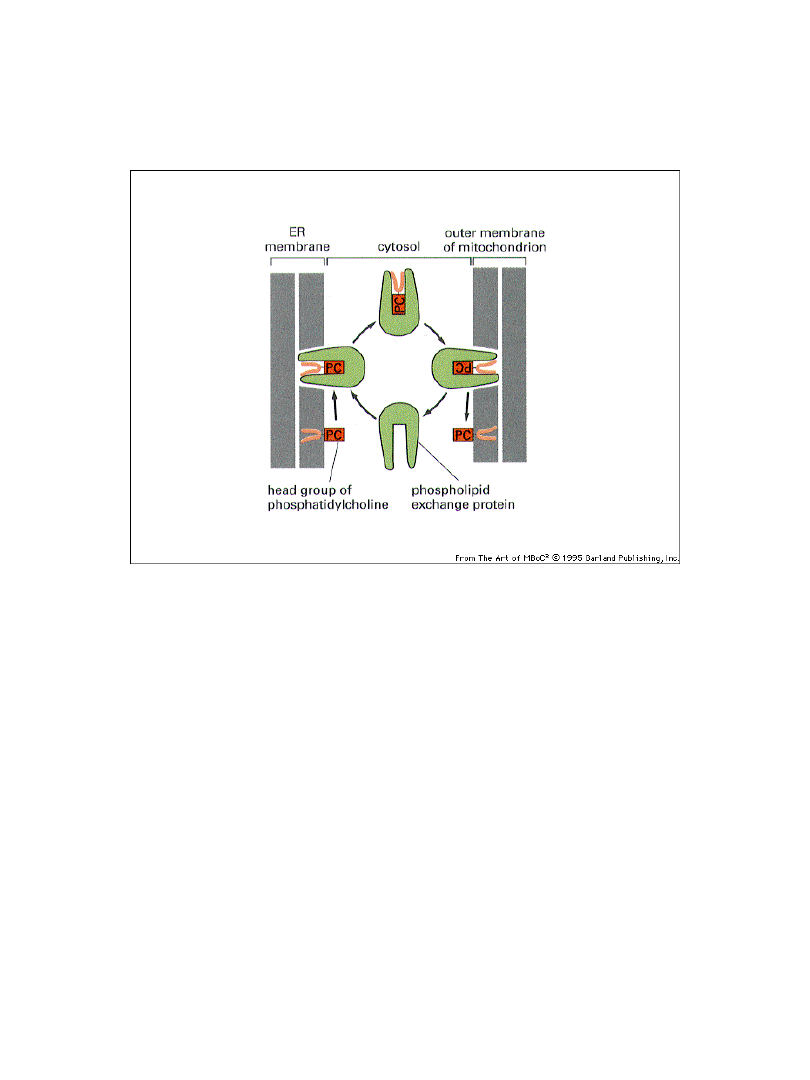
Białko wymiany fosfolipidowej
Sortowanie białek
np. transport z aparatu Golgiego do lizosomów
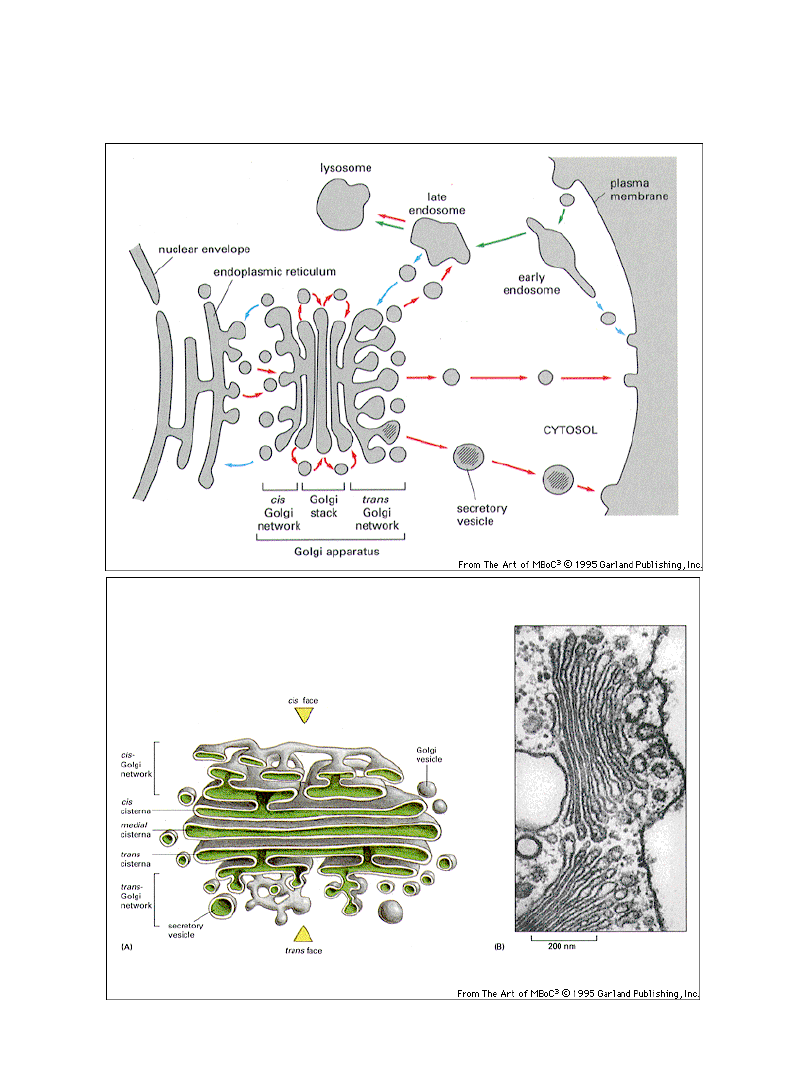
Budowa aparatu Golgiego
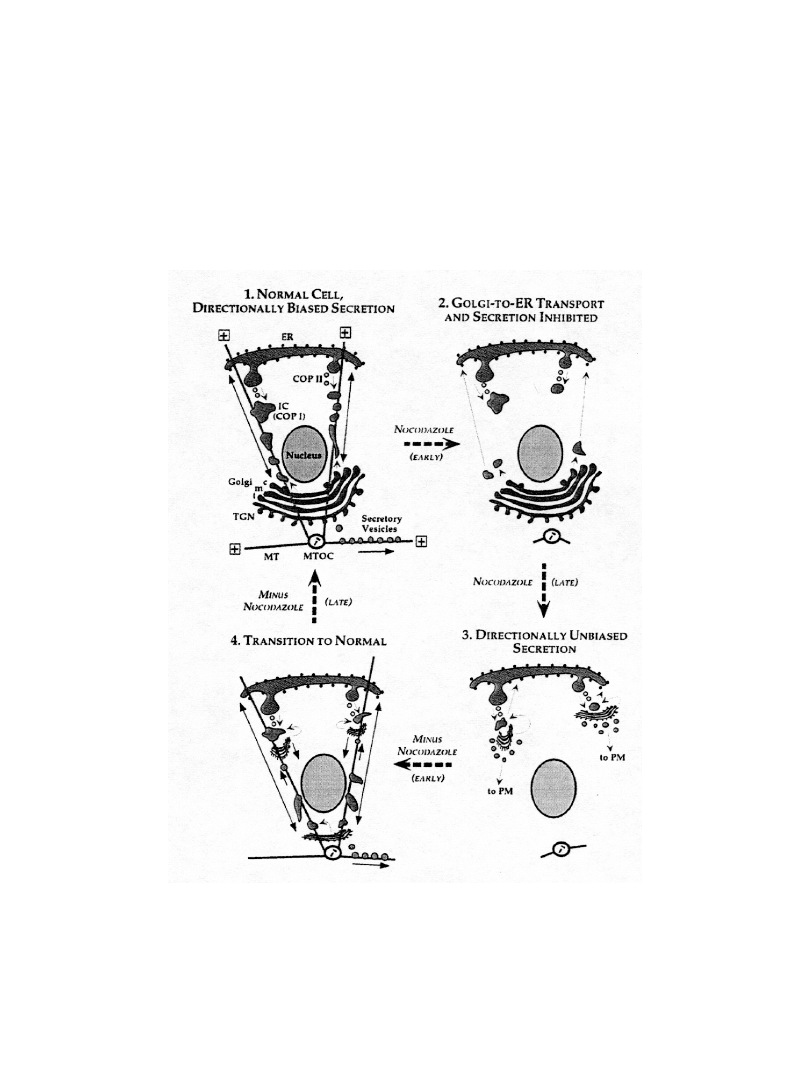
Funkcje mikrotubul w
transporcie substancji wewnątrz
komórki
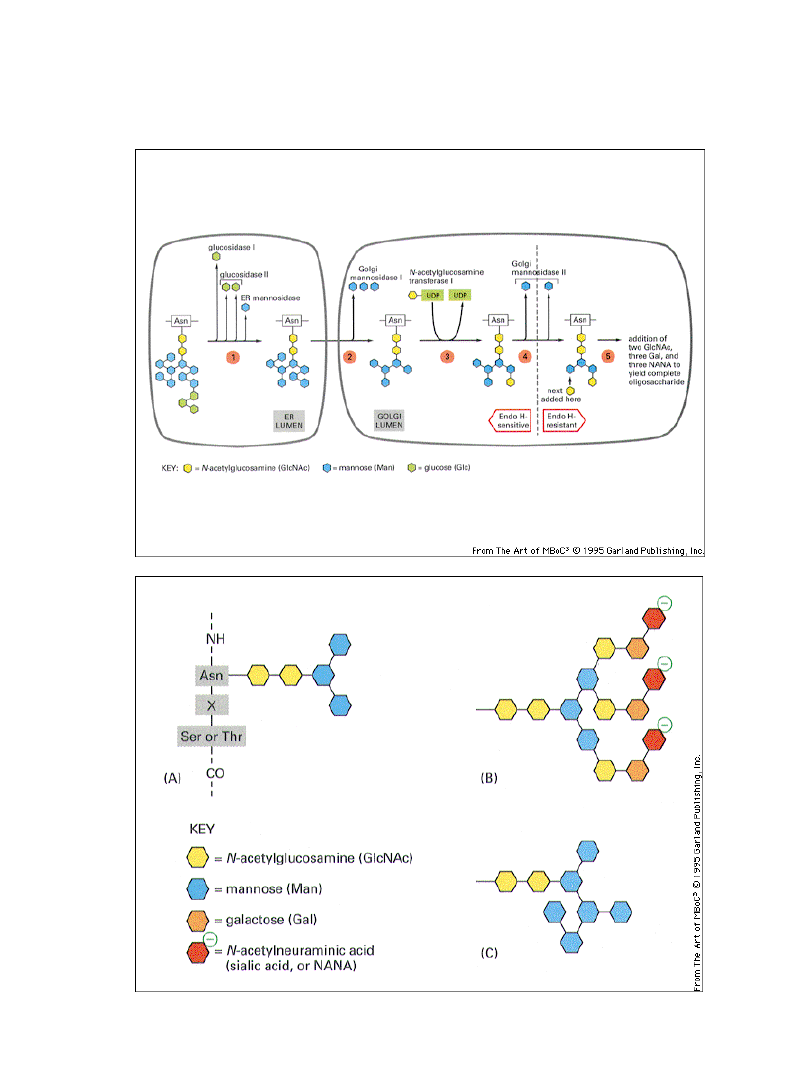
Obróbka oligosacharydów w obrębie
aparatu Golgiego
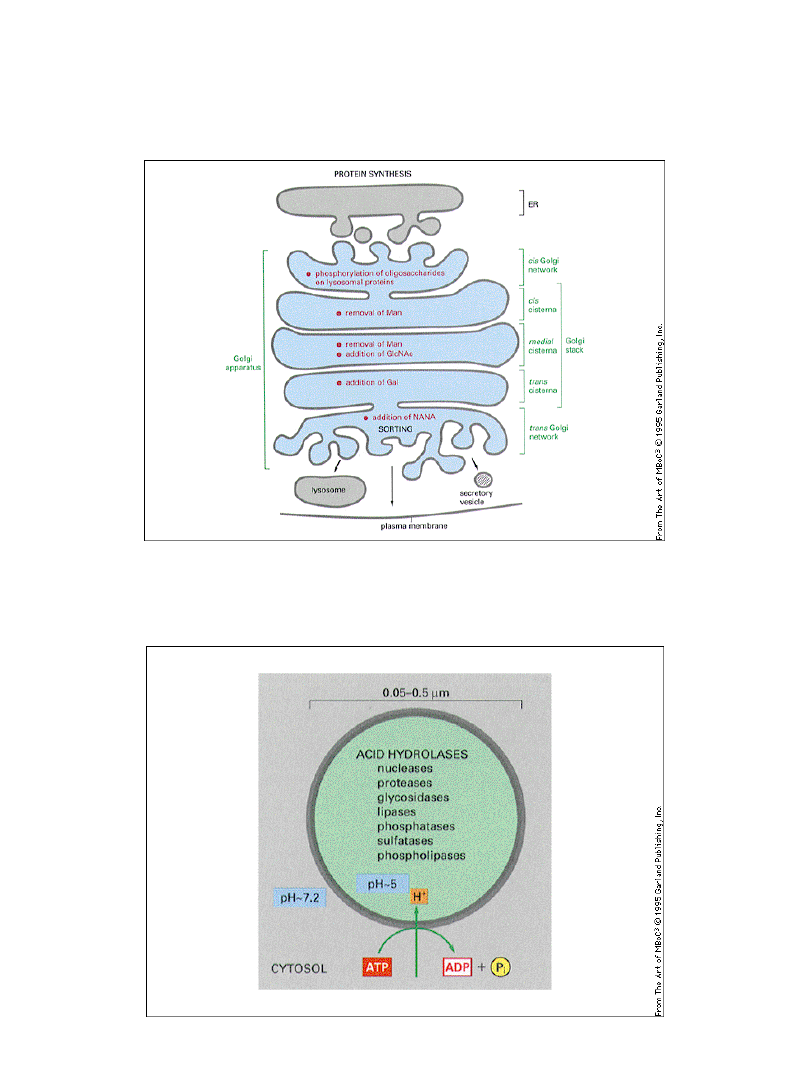
Aparat Golgiego możemy podzielić na
obszary
o różnych funkcjach
Jednym z zadań aparatu Golgiego
jest kierowanie białek do lizosomów
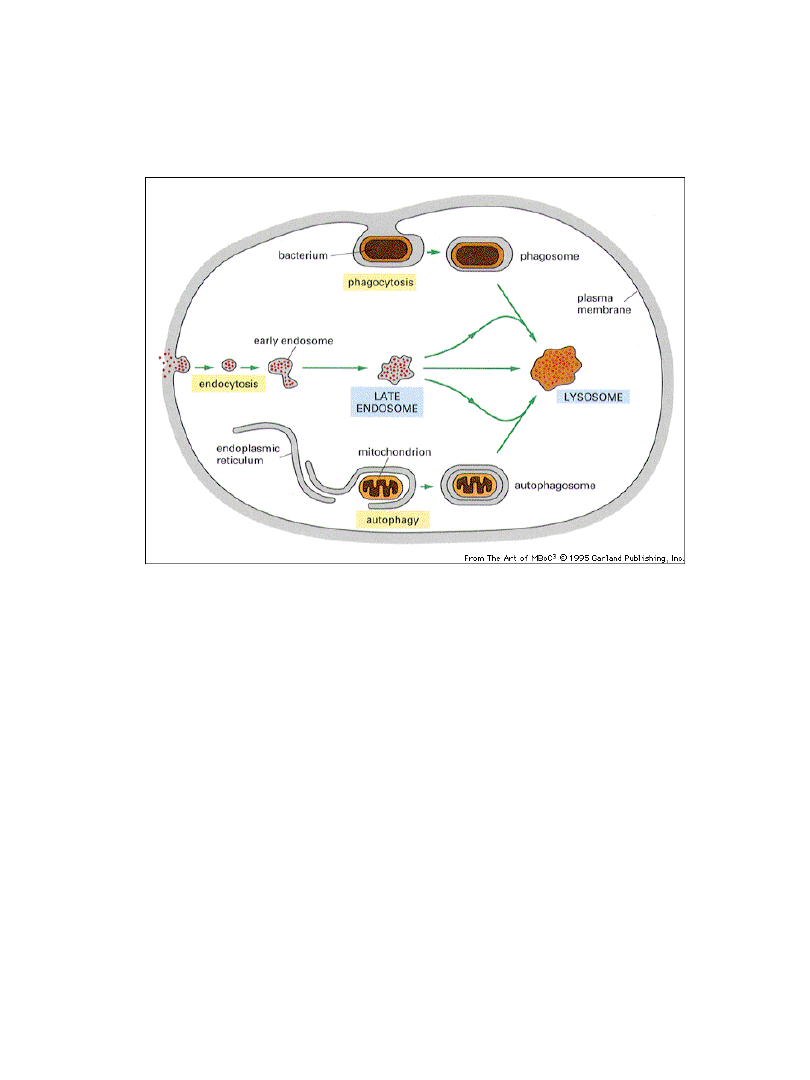
W lizosomach zachodzi degradacja
makromolekuł
Wiele z białek cytozolowych jest transportowana bezpośrednio
do lizosomów aby ulec degradacji
zawierają sekwencję aminokwasów KFERQ
lizyna-fenyloalanina-kwas glutaminowy-arginina-glutamina
bezpośrednie wiązanie z pęcherzykami ??
specjalne białko transportujące ??
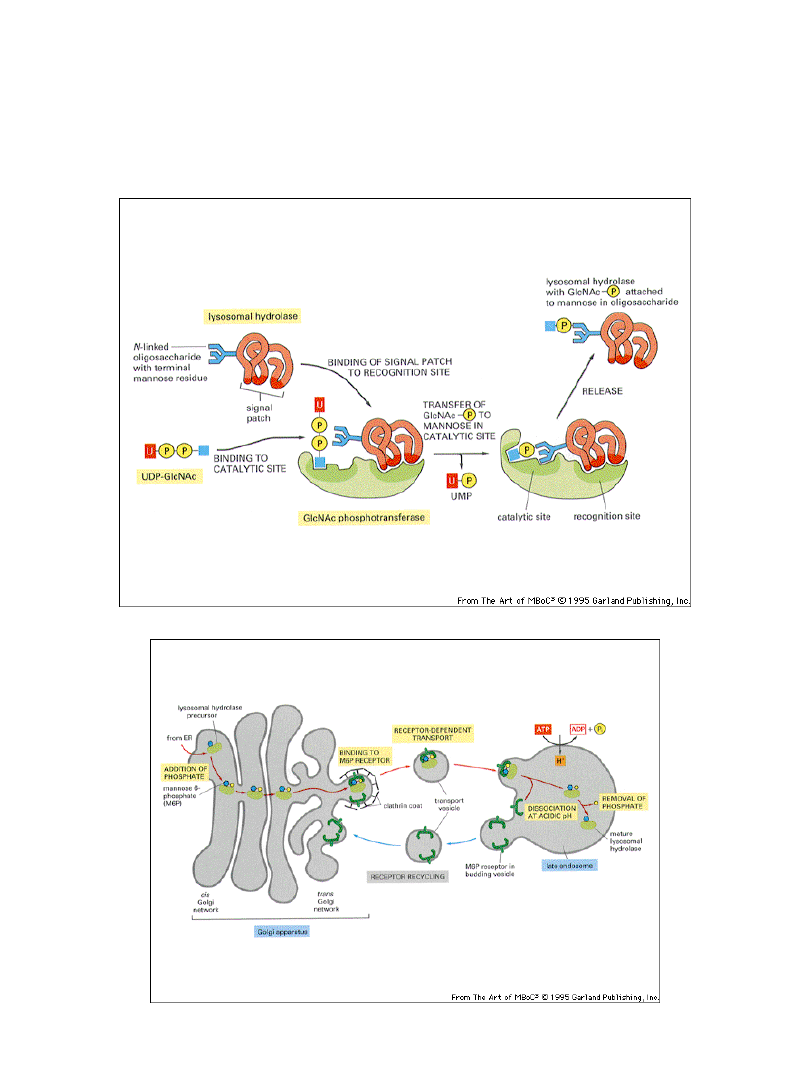
Do transportu nowozsyntetyzowanych
enzymów do lizosomów wymagane jest
przyłączenie grupy mannozo-6-
fosforanu
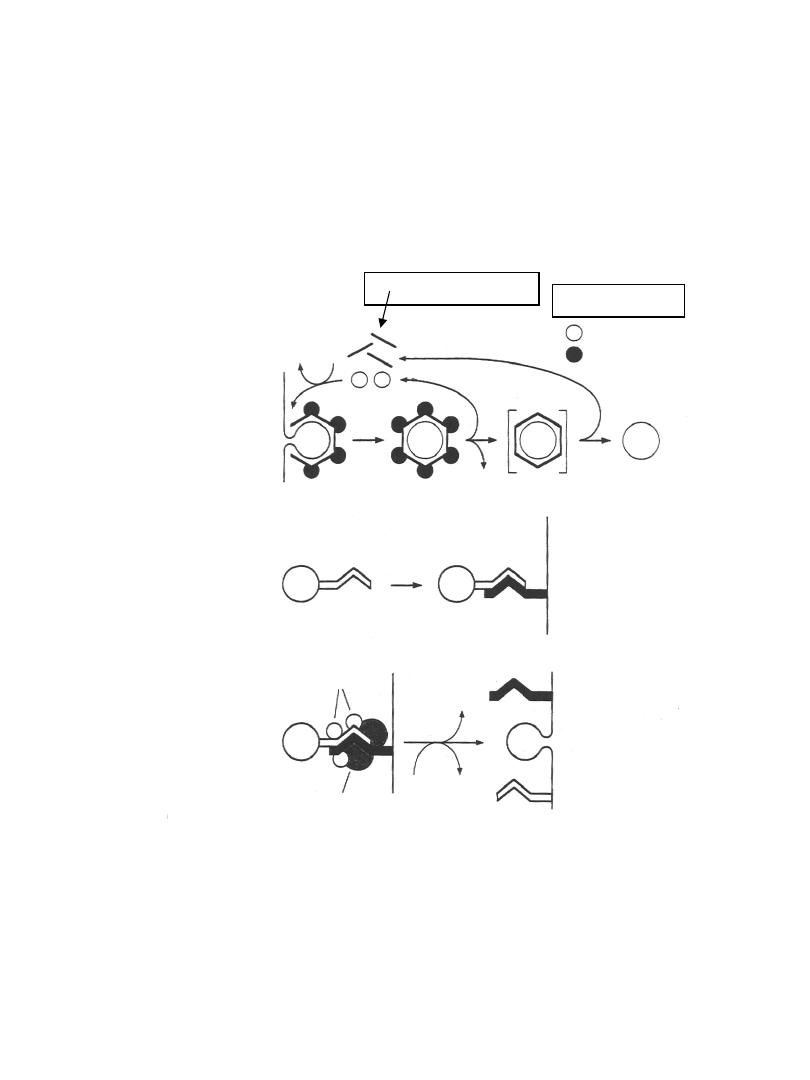
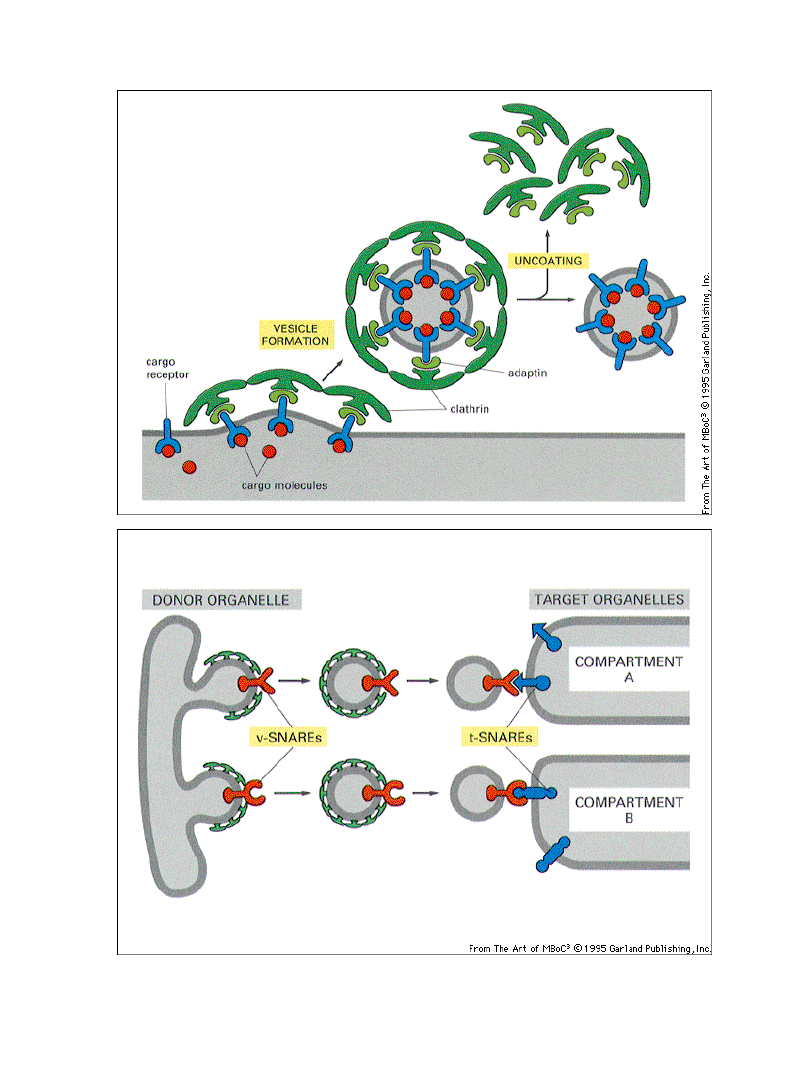
Etapy i składniki potrzebne do
powstawania pęcherzyków
transportowych i kierowania ich
do odpowiednich błon
błona-donor
GDPGTP
białko ARF
związane z GTP
związane z GDP
Pi
białka płaszcza
v-SNARE
t-SNARE
pęcherzyk transportowy
błona-cel
SNAPs
NSF
NSF
SNAPs
ATPADP
+Pi
1. tworzenie pęcherzyka transportowego
2. rozpoznawanie pęcherzyka
3. fuzja pęcherzyka
v-SNARE – identyfikator pęcherzyka
t-SNARE – akceptor pęcherzyka na blonie docelowej
SNAP, NSF – białka wywołujące fuzję pęcherzyka z błoną docelową
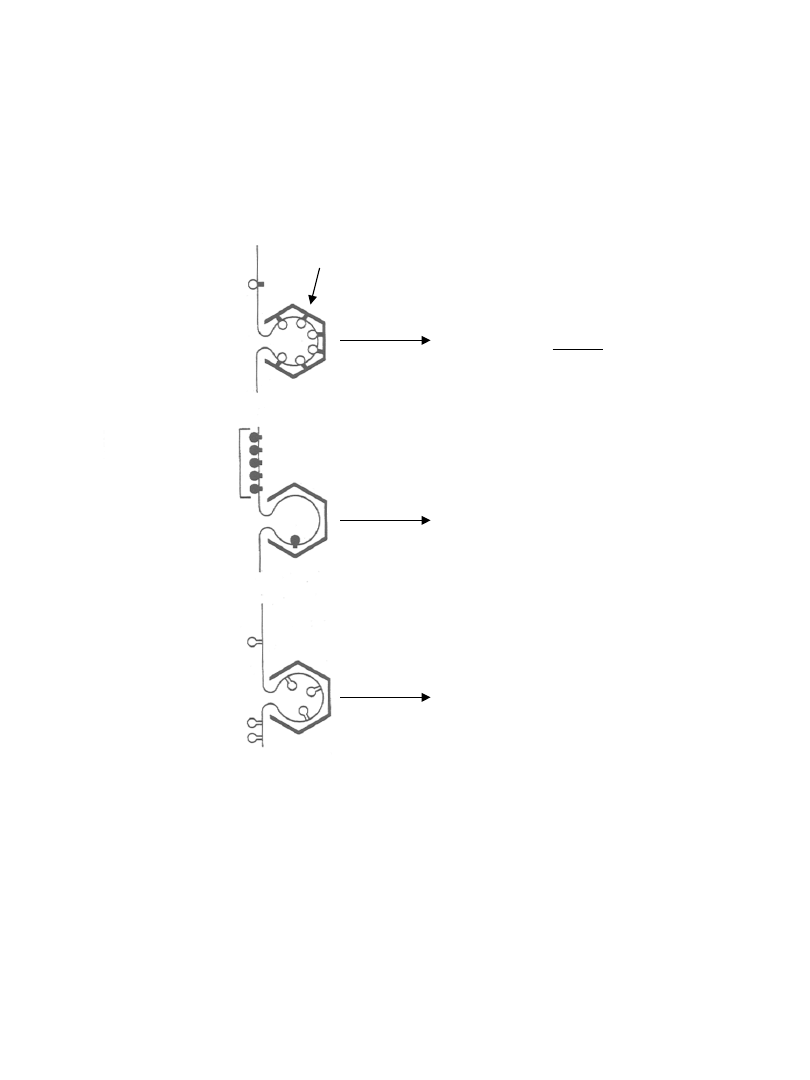
Los białka podczas transportu przez
dany typ pęcherzyka transportowego
białko gromadzi się w pęcherzyku
transportowym tylko gdy posiada
sygnał sortujący
białko nie gromadzi się w pęcherzyku
transportowym ponieważ posiada
sygnał zatrzymujący (ang. retention signal)
wiążący je z błoną-donor
białko przechodzi do wnętrza
pęcherzyka
transportowego przez dyfuzję
gdy nie posiada sygnału
sortującego
i sygnału zatrzymującego
sygnał sortujący wiąże się z białkami płąszcza
sygnał zatrzymujący
ogranicza przechodzenie
białka do wnętrza
pęcherzyka
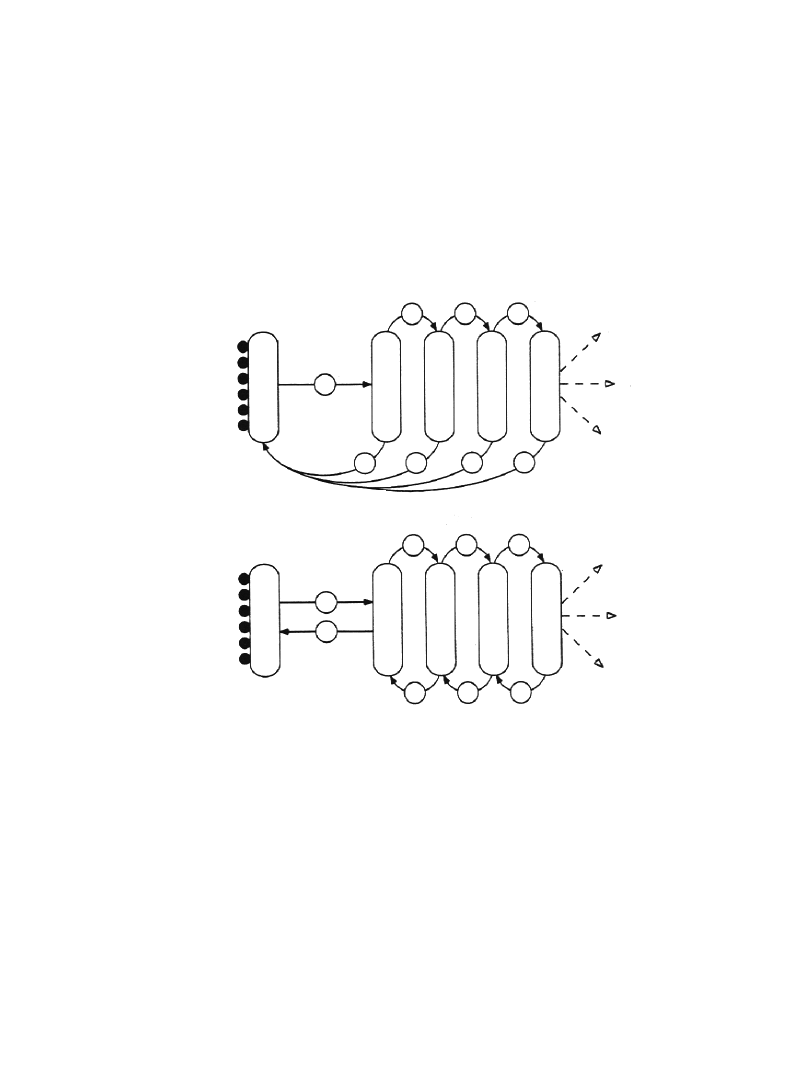
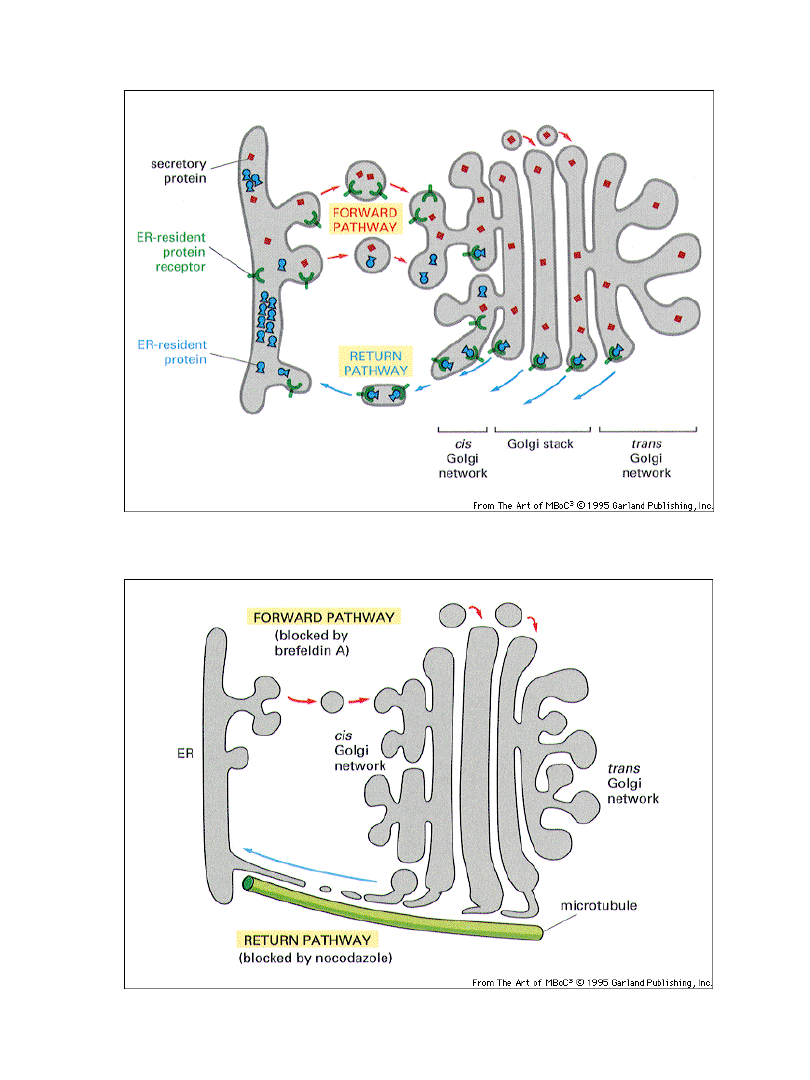
Dwa modele przepływu pęcherzyków
transportowych między aparatem
Golgiego a ER
model 2
model 1
ER
aparat Golgiego
cis
trans
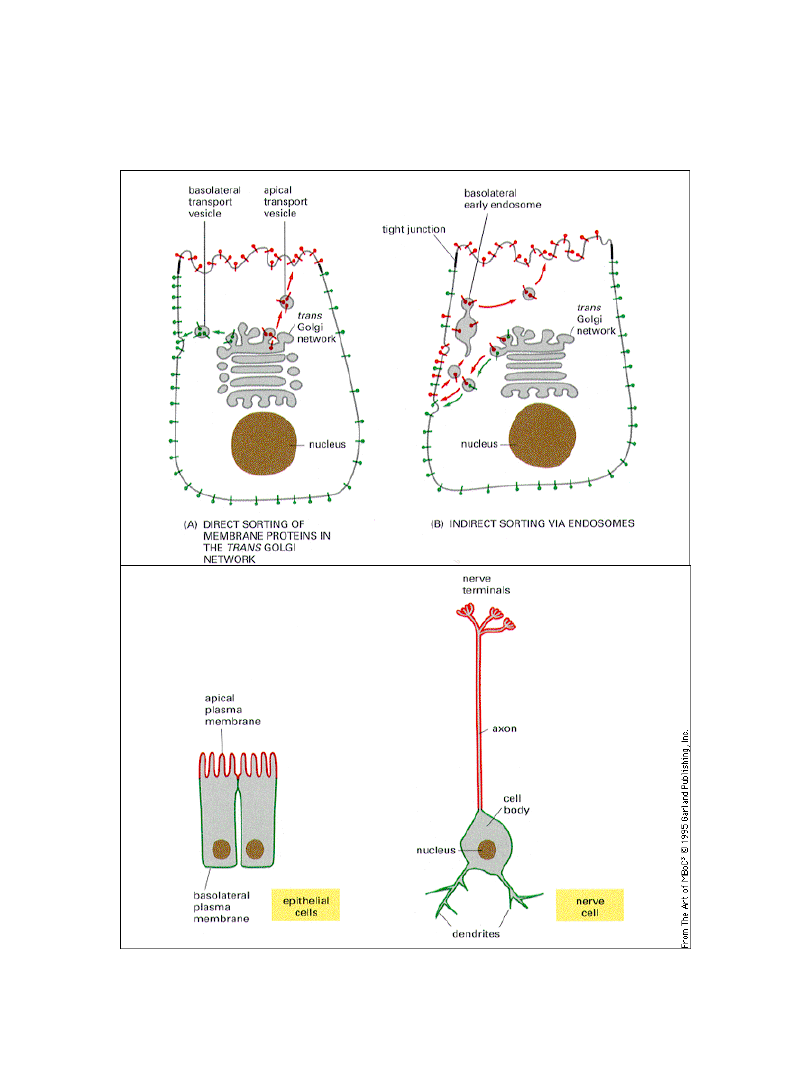
Sortowanie białek może być związane
z polaryzacją komórki
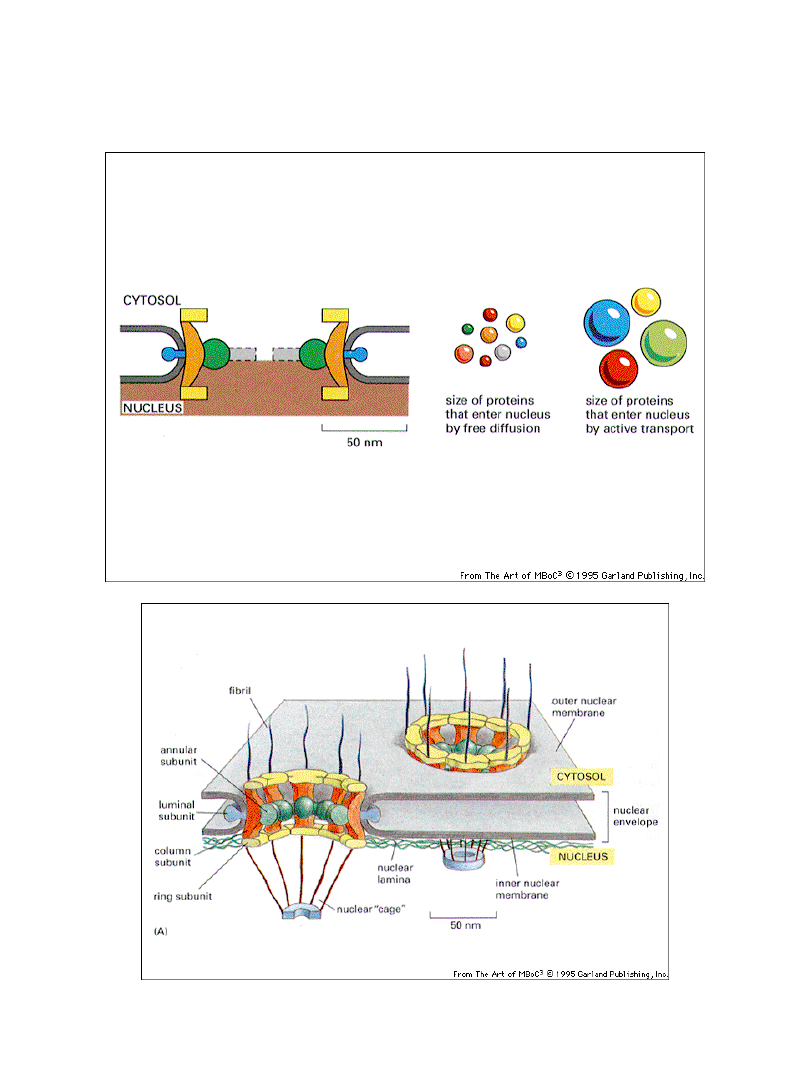
Transport pomiędzy cytoplazmą a jądrem
Kompleksy por jądrowych zapobiegaja biernemu
przemieszczaniu się białek cytoplazmatycznych do jądra
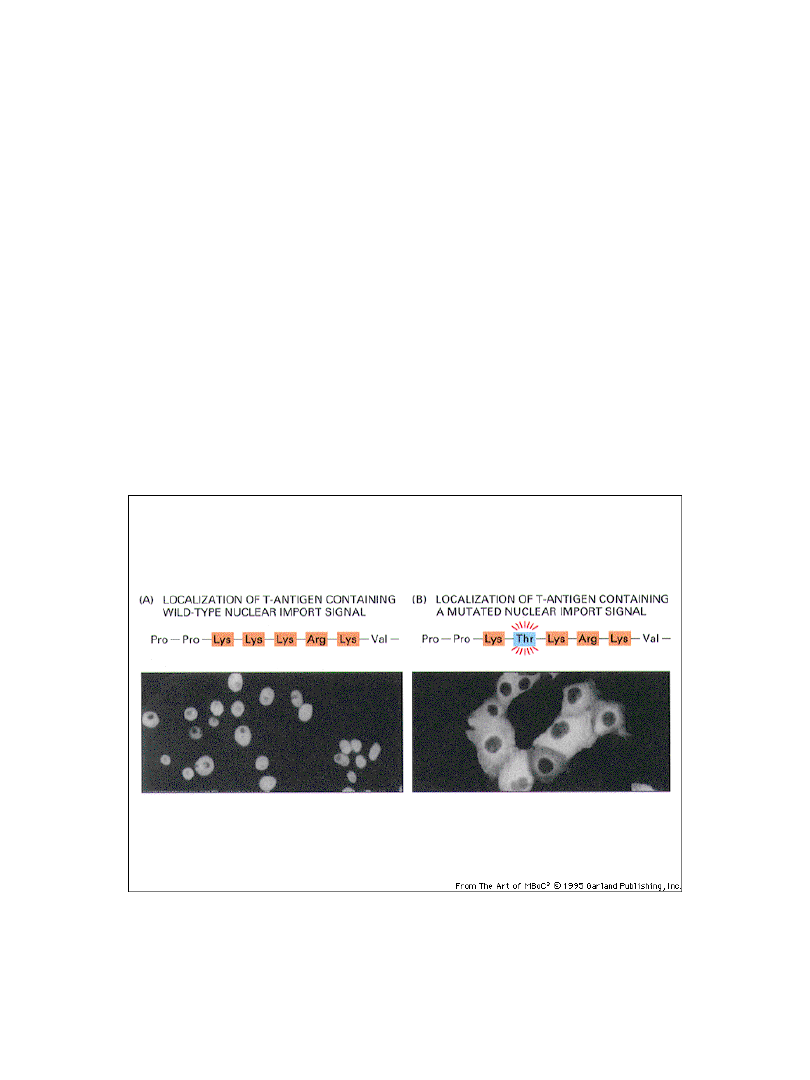
do transportu białka z cytoplazmy do jądra potrzebna jest
sekwencja lokalizacji jądrowej
4-8 aminokwasów naładowanych dodatnio, zwykle zawiera Pro
Funkcjonowanie sekwencji lokalizacji
jądrowej można śledzić za pomocą
technik immunofluorescencyjnych
Część białek przechodzi do jądra na
drodze
tzw. swobodnej dyfuzji
GTP
GDP
P
i
H
2
O
Ran-GDP
Ran-GTP
RanGAP1
RanGEF:RCC1
cytoplazma
jądro
RanGAP1 – Ran-GTP-activating protein 1
RanGEF – Ran-guanine-nucleotide exchange factor
(gł. RCC1)
Cykl GTP-azy Ran
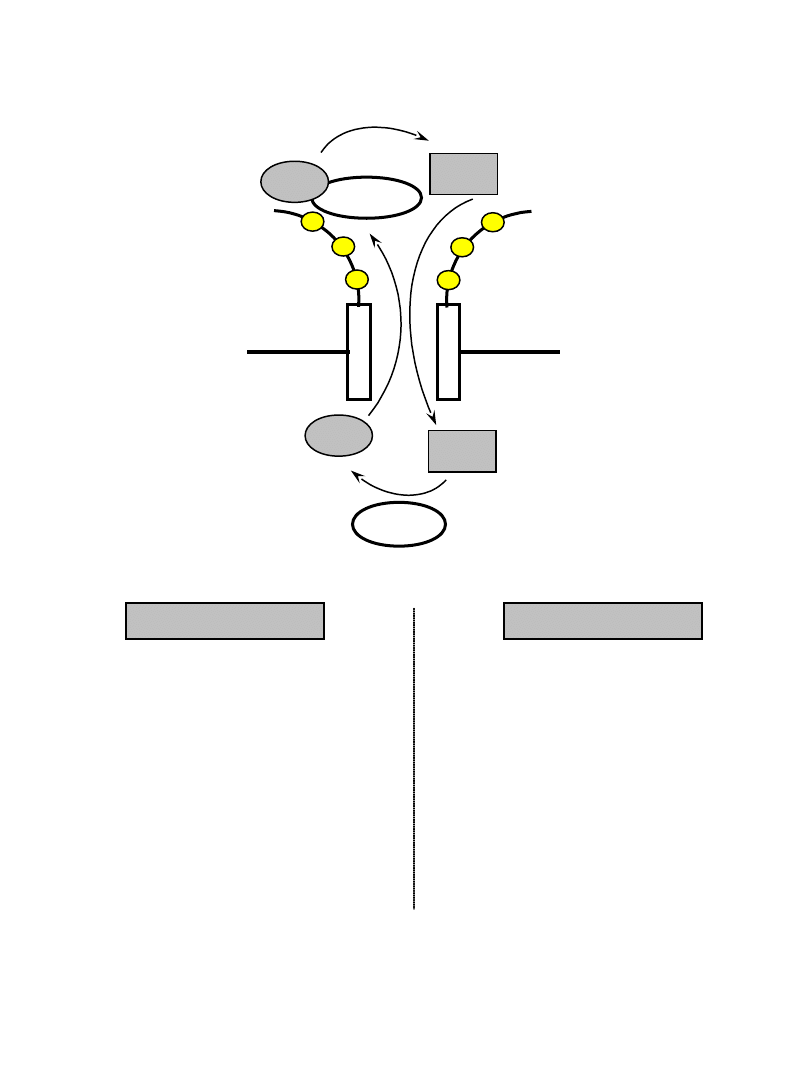
GTP
GDP
GDP
RanGAP
RCC1
GTP
wysokie stężenie GTP
niskie stężenie GDP
wysokie stężenie GDP
niskie stężenie GTP
Ran-GTP
Ran-GDP
cytoplazma
jądro
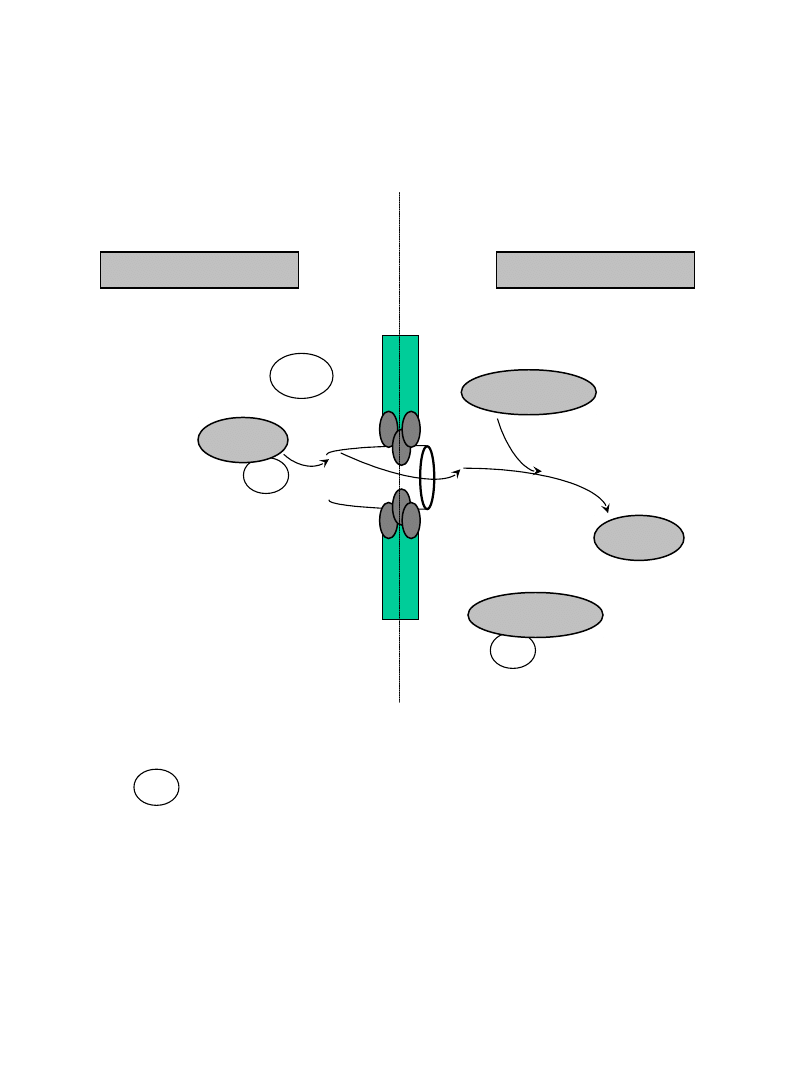
imp
cargo
cargo
Ran-GTP
imp
Ran-GTP
GDP
import
do jądra
NLS –nuclear localization sequence
imp
importin
cytoplazma
jądro
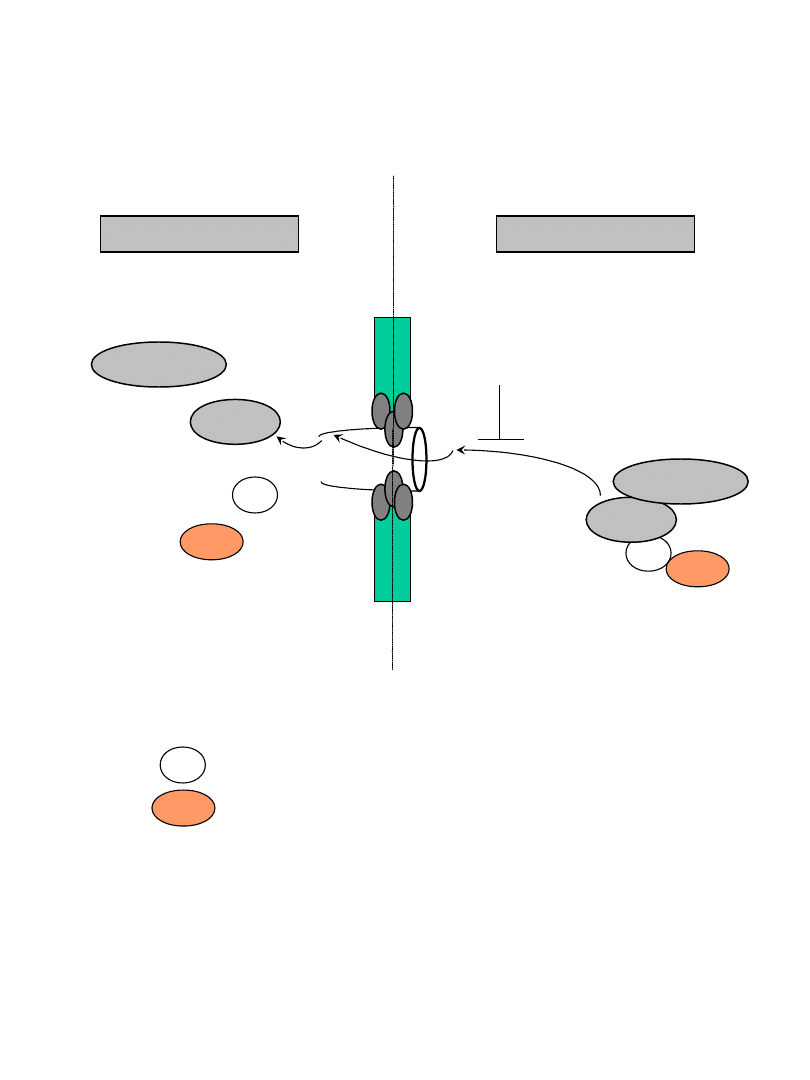
exp
cargo
exp
export
z jądra
cytoplazma
jądro
cargo
RanGAP1
Ran-GTP
Ran-GDP
Crm1
Crm1
leptomycin B
NES –nuclear export sequence
exportin
exp
Crm1
białko pomocnicze Crm1
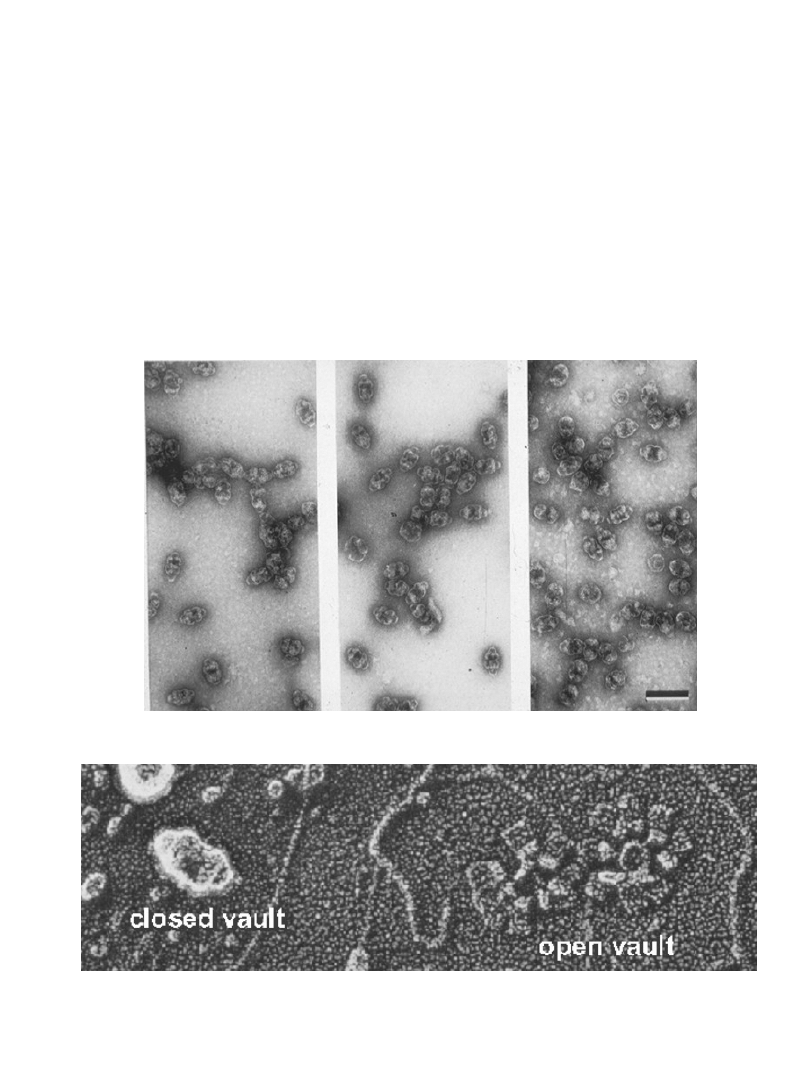
Krypta (ang. vault)– nowa organella
nowa organella odkryta 1986 roku
składa się z:
MVP (główne białka krypty)
ok. 100 kDa
Cap proteins (białka pierścienia centralnego)
240 kDa
vRNA (hgv1-4)
86-98 bp
szczur
żaba
królik
struktura zamknięta
struktura otwarta
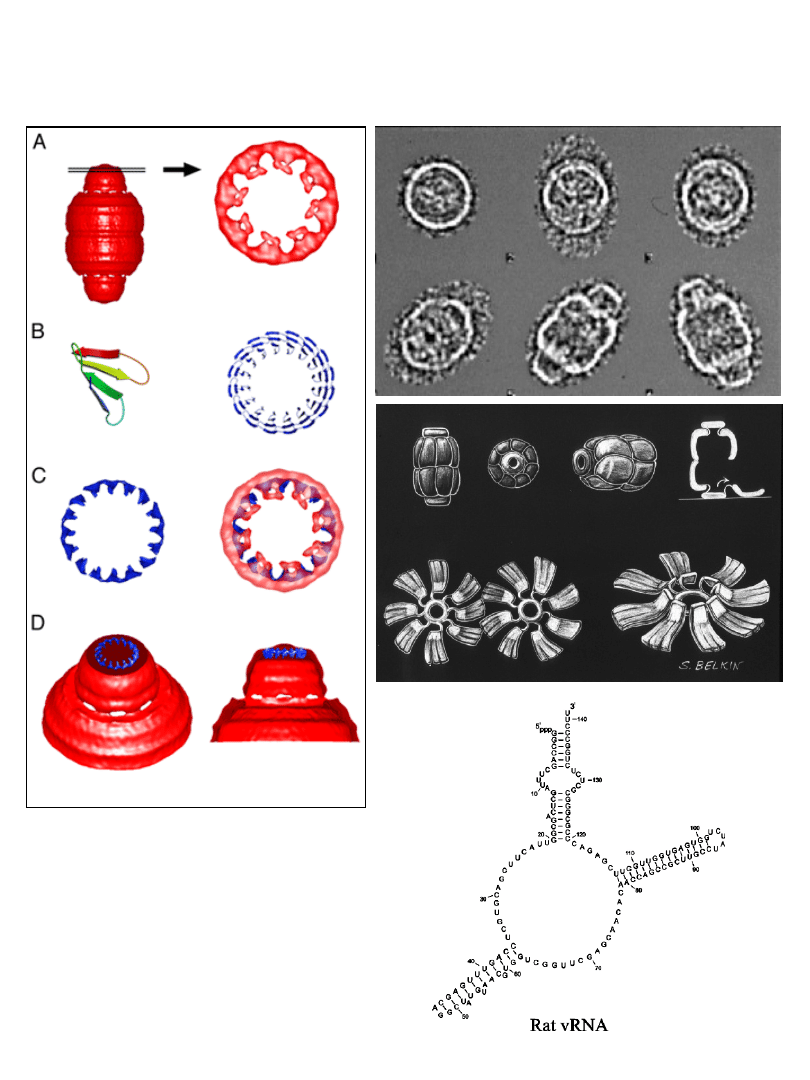
Krypta - struktura
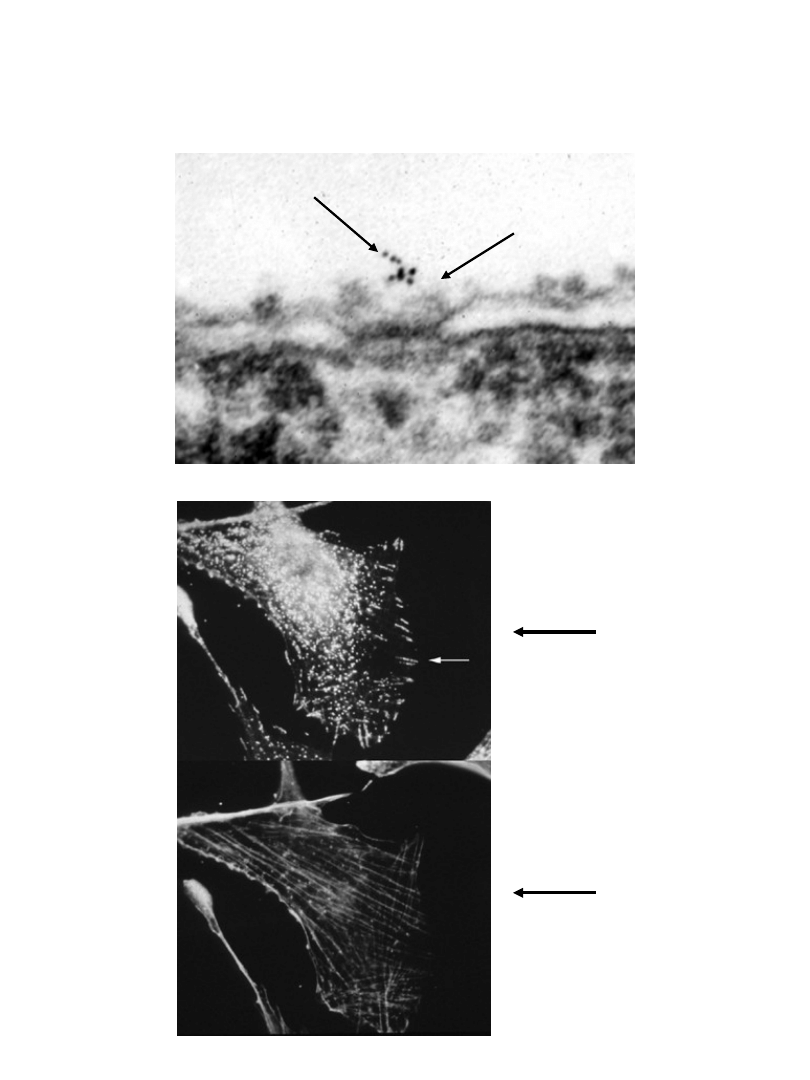
Krypta - rola w transporcie wewnątrzkomórkowym
krypta
aktyna
krypta
kompleks
pory jądrowej
Krypta - rola w oporności na ksenobiotyki
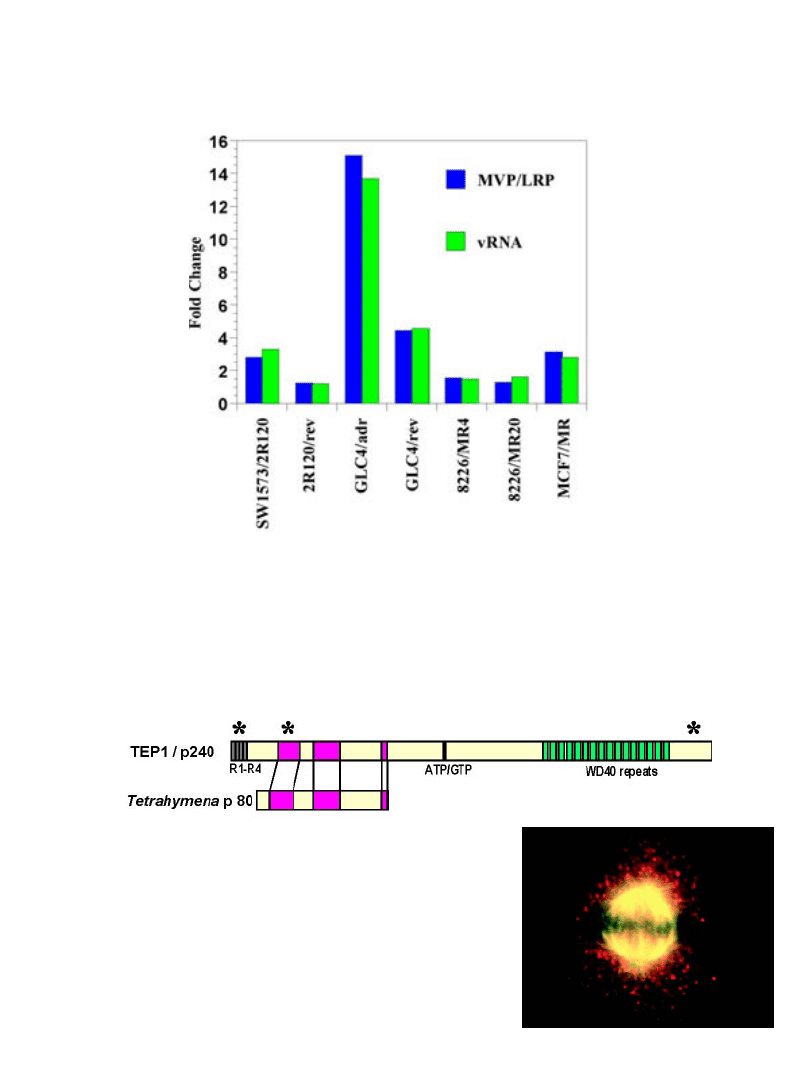
Krypta - niespodzianki
MVP ma homologię z TEP1 (składnik kompleksu telomerazy)
w krypcie odkryto aktywność vPARP
lokalizacja z wrzecionem mitotycznym
Transport RNA wewnątrz komórki
Lokalizacja RNA w określonych miejscach komórki
zależy od funkcji włókien cytoszkieletu (wł. aktynowych,
mikrotubul) oraz obecności klasy białek wiążących RNA
(białek przenoszących tzw. „listonoszy”)
np. Staufen
ZBP-1 (zipcode binding protein 1)
Białka zawierają miejsca rozpoznające np.
RLR – RNA Localization Region
RRM – RNA Recognizng Motif
RTS – RNA Transporting Sequence
dsRNA BM – double stranded RNA Binding Motif
przykłady m-RNA lokalizowanego przy pomocy
białek „listonoszy”:
Miranda
Prospero
Oscar
Inscutable
Numb i inne
ważne szczególnie w procesach różnicowania komórek
i rozwoju zarodkowego
Ważne: m-RNA jest nieaktywne translacyjnie w czasie transportu
(upakowanie/nieczynna konformacja)

Strony domowe nt. transportu wewnątrzkomórkowego
http://vaults.arc.ucla.edu + links
http://mrc-lmb.cam.ac.uk/myosin/myosin.html
http://www.blocks.fhcrc.org/~kinesin/index.html
Document Outline
- Slide 1
- Slide 2
- Slide 3
- Slide 4
- Slide 5
- Slide 6
- Slide 7
- Slide 8
- Slide 9
- Slide 10
- Slide 11
- Slide 12
- Slide 13
- Slide 14
- Slide 15
- Slide 16
- Slide 17
- Slide 18
- Slide 19
- Slide 20
- Slide 21
- Slide 22
- Slide 23
- Slide 24
- Slide 25
- Slide 26
- Slide 27
- Slide 28
- Slide 29
- Slide 30
- Slide 31
- Slide 32
- Slide 33
- Slide 34
- Slide 35
- Slide 36
- Slide 37
Wyszukiwarka
Podobne podstrony:
BKiIg sem 3 wykład 2 Transport wewnątrzkomórkowy
Transport wewnątrzzakładowy składowanie materiałów 2
Wozki jezdniowe z napedem silnikowym, BHP i PPOŻ przygotowanie do szkoleń, Transport wewnątrzzakłado
Wykorzystanie wózkow widłowych, BHP i PPOŻ przygotowanie do szkoleń, Transport wewnątrzzakładowy bhp
Bezpieczne korzystanie z wozków jezdniowych, BHP i PPOŻ przygotowanie do szkoleń, Transport wewnątrz
8 4 4 TRANSPORT WEWNĄTRZ ZAKŁADOWY
W8 Transport wewnatrzkomorkowy
transport wewnątrzszpitalny, ORGANIZACJA I ZARZĄDZANIE W SŁUŻBIE ZDROWIA
transport wewnątrz zakładowy, BHP
kosz podajnika, Transport wewnątrzzakładowy bhp
kubica, biologia z elementami mikrobiologii, Transport wewnątrzkomórkowy i jego znaczeniex
Transport kopalniany jest transportem wewnątrz zakładowym
07-transport wewnątrzzakładowy, Instrukcje BHP, XXXII - TRANSPORT
Transport wewnątrzzakładowy(1), Zachomikowane - bhp, TBP i Analiza Zagrożeń
Transport wewnatrzzakladowy
BKiIg sem 3 wykład 2 Transport wewnątrzkomórkowy
Transport wewnątrzzakładowy(1)
więcej podobnych podstron