
96
P
oradniki ochrony siedlisk i gatunk
ów
Wody s∏odkie i torfowiska
Nizinne i podgórskie
rzeki ze zbiorowiskami
w∏osieniczników
Kod Physis: 24.4: euhydrofityczna roÊlinnoÊç rzek
Definicja
Cieki wodne – nizinne do podgórskich – poroÊni´te przez
zakorzenione w dnie roÊliny zanurzone lub z p∏ywajàcymi
liÊçmi ze zwiàzku
Ranunculion fluitantis lub wodne mszaki.
Charakterystyka
Dna koryt strumieni i rzek, najcz´Êciej (I) – II – V rz´du
o Êrednio intensywnym do intensywnego przep∏ywie wody,
osadach z dominujàcà frakcjà piaszczystà, ˝wirowà lub
drobnokamienistà, zasilane wodami podziemnymi lub
z intensywnà wymianà wód mi´dzy korytem rzeki a jej stre-
fà hyporeicznà poprzez przepuszczalne osady denne. Cie-
ki te przynajmniej we fragmentach poroÊni´te sà przez
p∏aty roÊlin zakorzenionych w dnie i z zanurzonymi w wo-
dzie p´dami, rzadziej wytwarzajàcych równie˝ liÊcie p∏ywa-
jàce po powierzchni wody lub p´dy cz´Êciowo wyniesione
ponad powierzchni´ (cz´ste przy brzegu lub w mikrosiedli-
skach o spowolnionym przep∏ywie wody). W sk∏ad roÊlin-
noÊci siedliska wchodzà równie˝ przytwierdzone do dna
mszaki i makroskopowe glony (zielenice nitkowate, kra-
snorosty i ramienice).
Podzia∏ na podtypy
W zachodnioeuropejskich klasyfikacjach wyró˝nia si´ zwy-
kle kilka (do 6–7) ró˝niàcych si´ strukturà roÊlinnoÊci pod-
typów, w zale˝noÊci od odczynu wody i jej potencja∏u tro-
ficznego (Francja) lub charakteru geologicznego pod∏o˝a
(wapienne – bezwapienne, co koresponduje z odczynem)
i spadku odcinka cieku (Niemcy). Ze wzgl´du na brak lub
niewielkà reprezentacj´ na terenie Polski niektórych z uwa-
runkowaƒ fizycznych stanowiàcych podstaw´ klasyfikacji,
zwykle zubo˝onà struktur´ zbiorowisk roÊlinnych w gra-
nicznym obszarze zasi´gu siedliska na terenie Polski oraz
s∏aby stopieƒ poznania i nieliczne tylko opisy roÊlinnoÊci,
brak jest obecnie podstaw do Êcis∏ego i udokumentowane-
3260
Rz´Êl hakowata (
Callitriche hamulata), w∏osiecznik p´dzelkowaty (Batrachium penicillatum
)
i rdestnica st´piona (
Potamogeton obtusifolius)
w korycie Starej Radwi (Rosnowo, pow. Koszalin)

97
Tytu∏ rozdzia∏u Tytu∏ rozdzia∏u Tytu∏ rozdzia∏u Tytu∏
P
oradniki ochrony siedlisk i gatunk
ów
Nizinne i podgórskie rzeki ze zbiorowiskami w∏osieniczników
go wyró˝nienia podtypów siedliska. Niektóre przes∏anki do
ewentualnej póêniejszej klasyfikacji zostanà wskazane
w opisie odmian siedliska. Nie nale˝y jednak oczekiwaç –
nawet po dok∏adniejszym zbadaniu – mo˝liwoÊci zapropo-
nowania równie zró˝nicowanego podzia∏u, jak w europej-
skim obszarze klimatu atlantyckiego.
Umiejscowienie siedliska w polskiej
klasyfikacji fitosocjologicznej
Omawiane siedlisko wed∏ug polskiej klasyfikacji obejmuje
jedynie zwiàzek
Ranunculion fluitantis, zaliczany do klasy
Potametea, przy czym jedynie jeden z gatunków uznawa-
nych za charakterystyczne dla tej klasy (
Elodea canadensis)
wyst´puje jako sk∏adnik zbiorowisk tego siedliska.
Pozycja syntaksonomiczna siedliska w polskiej klasyfikacji
przedstawia si´ nast´pujàco:
Klasa
Potametea
Rzàd
Potametalia
Zwiàzek
Ranunculion fluitantis
Zespo∏y:
Ranunculetum fluitantis w∏osienicznika rzecz-
nego
Ranunculo–Callitrichetum hamulatae rz´Êli
hakowatej i w∏osieniczników
Ranunculo–Sietum erecto–submersi podwodnej
formy potocznika i w∏osieniczników
Stan poznania tych zespo∏ów na obszarze Polski jest dale-
ce niewystarczajàcy, a przysz∏e badania prawdopodobnie
doprowadzà do znaczàcej rewizji tej klasyfikacji. M. in. wy-
mieniony w definicji siedliska wg Natura 2000 zwiàzek
Callitricho–Batrachion (opisywany jako charakterystyczny
dla rzek z wyraênymi przep∏ywami ni˝ówkowymi, a wi´c ty-
powymi dla klimatu Polski) nie zosta∏ uwzgl´dniony w pol-
skiej klasyfikacji fitosocjologicznej. W europejskich bada-
niach roÊlinnoÊci zanurzonej rzek podkreÊla si´ jednak
cz´sto nieadekwatnoÊç metodyki fitosocjologicznej do kla-
syfikacji zbiorowisk roÊlin wód p∏ynàcych.
Bibliografia
3260 – Rivi¯res des étages planitiaire ∫ montagnard avec
végétation du
Ranunculion fluitantis
et du
Callitricho–Ba-
trachion
In: Ministére de l’écologie et du développement du-
rable: Le réseau écologique européen Natura 2000. Cahier
d habitats, Habitats humides: 191–222. http: //natura2000.
environnement. gouv. fr
ADAMEC L., ONDOK P. 1992. Water alkalization due to photo-
synthesis of aquatic plants: the dependence on total alkalini-
ty. Aquatic Botany 43: 93–98.
ANDRZEJEWSKI W., NAGENGAST B. 2002. Bioró˝norodnoÊç rezerwa-
tu przyrody „S∏onawy” na rzece We∏nie. W: Puchalski W., PAWEL-
CZUK J. K. (red.) Bliskie Naturze Kszta∏towanie Dolin Rzecznych.
Wydawn. Uczeln. Politechniki Koszaliƒskiej, Koszalin: 80–81.
BAATTRUP–PEDERSEN A., LARSEN S. E., RIIS T. 2003. Composi-
tion and richness of macrophyte communities in small Da-
nish streams – influence of environmental factors and weed
cutting. Hydrobiologia 495: 171–179.
BAATTRUP–PEDERSEN A., RIIS T. 1999. Macrophyte diversity and
composition in relation to substratum characteristics in regu-
lated and unregulated Danish streams. Freshwater Biology
42: 375–385.
BAATTRUP–PEDERSEN A., RIIS T., HANSEN H. O. FRIBERG N.
2000. Restoration of a Danish headwater stream: short–term
changes in plant species abundance and composition. Aqua-
tic Conserv.: Mar. Freshw. Ecosyst. 10: 13–23.
BAATTRUP–PEDERSEN A., SKRIVER J., WIBERG–LARSEN P. 2000.
Weed–cutting practice and impact on trout density in Danids
lowland streams. Verh. Internat. Verein. Limnol. 27:
674–677.
BARENDREGT A., WASSEN M. J. 1994. Surface water chemistry
of the Biebrza River with special emphasis on nutrient flow
and vegetation. W: Report 2: Biebrza Wetlands Research of
the Utrecht University presented at the Workshop „Biebrza
Wetlands”, October 1993, IMUZ, Falenty. Red. M. J. Wassen,
H. Okruszko. Falenty, Utrecht: 133–146.
BARRAT–SEGRETAIN M. H., AMOROS C. 1995. Influence of flo-
od timing on the recovery of macrophytes in a former river
channel. Hydrobiologia 316: 91–101.
BARRAT–SEGRETAIN M. H., BORNETTE G., 2000. Regeneration
and colonization abilities of aquatic plant fragments: effect of
disturbance seasonality. Hydrobiologia 421: 31–39.
BARRAT–SEGRETAIN M. H., HENRY C. P., BORNETTE G. 1999.
Regeneration and colonization of aquatic plant fragments in
relation to the disturbance frequency of their habitats. Arch.
Hydrobiol. 145: 111–127.
BERNEZ I., HAURY J. 1996. Downstream effects of hydroelectric
impoundment on river macrophyte communities. In: Leclerc
M., Capra H., Valentin S., Boudreault A., Cote Y. (eds) Eco-
hydraulics 2000 Québec, INRS–Eau Québec: A13–A24.
BIA¸OKOZ W. 1999. Biebrzaƒski Park Narodowy – Plan Ochro-
ny. Operat Ochrony Ekosystemów Wodnych.
BODNER M. 1994. Inorganic carbon source for photosynthesis in
the aquatic macrophytes Potamogeton natans and Ranuncu-
lus fluitans. Aquatic Botany 48:109–120.
BORNETTE G., AMOROS C., LAMOUROUX N. 1998. Aquatic
plant diversity in riverine wetlands: the role of connectivity.
Freshwater Biology 39: 267–283.
BORNETTE G., GUERLESQUIN M., HENRY P. H. 1996. Are the
Characeae able to indicate the origin of groundwater in for-
mer river channels? Vegetatio 125: 207–222.
BRYLI¡SKA M. (red.) 2000. Ryby s∏odkowodne Polski. PWN, War-
szawa, s. 521.
CARBIENER R., TRÉMOLIÉ RES M., MERCIER J. L., ORTSCHEIT A.
1990. Aquatic macrophyte communities as bioindicators of
eutrophication in calcareous oligosaprobe stream waters
(Upper Rhine plain, Alsace). Vegetatio 86: 71–88.
DAJDOK Z., PROåKÓW J. 2003. Flora wodna i b∏otna Dolnego
3260

Âlàska na tle zagro˝eƒ i mo˝liwoÊci ochrony. W: Kàcki Z.
(red.) Zagro˝one gatunki flory naczyniowej Dolnego Âlàska.
PTPP „Pro Natura”, Wroc∏aw:131–150.
DUFF J. H., HENDRICKS S. P., JACKMAN A. P., TRISKA F. J. 2002.
The effect of Elodea canadensis beds on porewater chemi-
stry, microbial respiration, and nutrient retention in the Shin-
gobee River, Minnesota, North America. Verh. Internat. Vere-
in. Limnol. 28: 214–222.
DYDUCH–FALNIOWSKA A., HERBICH J., HERBICHOWA M.,
MRÓZ W., PERZANOWSKA J. 2002. Wdra˝anie koncepcji
sieci NATURA 2000 w Polsce w latach 2001–2003. Materia-
∏y instrukta˝owe dla wojewódzkich zespo∏ów realizacyjnych:
Krótka charakterystyka typów siedlisk przyrodniczych o zna-
czeniu europejskim, wyst´pujàcych w Polsce. Kraków-
–Gdaƒsk 2002.
EICHENBERGER E., WEILENMANN H. U. 1982. The growth of
Ranunculus fluitans Lam. in artificial canals. In: Symoens J. J.,
Hooper S. S., Compere P. (eds.) Studies on aquatic vascular
plants. Royal Botanical Soc. Belgium, Brussels: 324–332.
EUROPEAN COMMISSION, DG ENVIRONMENT, NATURE AND
BIODIVERSITY 2003. NATURA 2000. Interpretation Manual
of European Union Habitats. EUR 25.
FJORBACK C., KRONVANG B. 2002. Interaction between aqu-
atic macrophytes and stream hydraulics in a Danish stream:
implications of different weed–cutting methods. Verh. Inter-
nat. Verein. Limnol. 28: 1299–1303
GREULICH S., BORNETTE G.1999. Competitive abilities and re-
lated strategies in four aquatic plant species from an inter-
mediately disturbed habitat. Freshwater Biology 41:
493–506.
HARRISON S. S. C. 2000. The importance of aquatic margins to
invertebrates in English chalk streams. Arch. Hydrobiol. 149:
213–240.
HARRISON S. S. C., HARRIS I. T., CROEZE A., WIGGERS R. 2000.
The influence of bankside vegetation on the distribution of
aquatic insects. Verh. Internat. Verein. Limnol. 27:
1480–1484.
HATTON–ELLIS T. W., GRIEVE N. 2003. Ecology of watercourses
characterised by
Ranunculion fluitantis and Callitricho–Batra-
chion vegetation. Conserving Natura 2000 Rivers Ecology
Series No. 11, English Nature, Peterborough, s.64,
http: //www.riverlife.org.uk
HAURY J., 1996. Assessing functional typology involving water
quality, physical features and macrophytes in a Normandy ri-
ver. Hydrobiologia 340: 43–49.
HAURY J., AĪDARA L. G. 1999. Macrophyte cover and standing
crop in the River Scorff and its tributaries (Brittany, northwe-
stern France): scale, patterns and process. Hydrobiologia
415: 109–115.
HAURY J., DUTARTRE A., BINESSE F., CODHANT H., VALKMAN G.
2001. Macrophyte biotypologies of rivers in Lozere – France.
Verh. Internat. Verein. Limnol. 27: 3510–3517.
HERBICH J., GÓRSKI W. 1993. Specyfika, zagro˝enia i problemy
ochrony przyrody dolin ma∏ych rzek Pomorza. W: Tomia∏ojç
L. (red.) Ochrona przyrody i Êrodowiska w dolinach nizinnych
rzek Polski. Wydawn. Instytutu Ochrony Przyrody PAN, Kra-
ków: 167–188.
JASNOWSKI M., JASNOWSKA J., FRIEDRICH S. 1986. RoÊlinnoÊç
rzeczna, torfowiskowa i êródliskowa projektowanego Drawieƒ-
skiego Parku Narodowego. W: Agapow L., Jasnowski M. (red.)
Przyroda projektowanego Drawieƒskiego Parku Narodowego.
Gorzowskie Towarzystwo Naukowe, Gorzów Wlkp.: 69–94.
JO¡CA T., DÑBROWSKA B. B. , PUCHALSKI W. 2002. Ekologicz-
ne efekty podpi´trzeƒ ma∏ych rzek na przyk∏adzie Skotawy.
W: Puchalski W., Pawelczuk J. K. (red.): Bliskie Naturze
Kszta∏towanie Dolin Rzecznych. Wydawn. Uczeln. Politechni-
ki Koszaliƒskiej, Koszalin: 109–111.
KAENEL B. R., BUEHRER H., UEHLINGER U. 2000. Effects of aqu-
atic plant management on stream metabolism and oxygen
balance in streams. Freshwater Biology 45: 85–95.
KING J. J., O’GRADY M. F., CURTIN J. 2000. The experimental
drainage maintenance (EDM) programme: engineering and
fisheries management interactions in drained Irish salmonid
channels. Verh. Internat. Verein. Limnol. 27: 1480–1484.
KRASKA M. 1998. RoÊlinnoÊç rzek Drawy i P∏ocicznej w Drawieƒ-
skim Parku Narodowym. Operat Ochrony Ekosystemów
Wodnych Drawieƒskiego Parku Narodowego. Zak∏ad Ochro-
ny Wód Uniwersytetu im. A. Mickiewicza, Poznaƒ.
LIFE IN UK RIVERS 2003. Monitoring watercourses characterised by
Ra-
nunculion fluitantis
and
Callitricho–Batrachion
vegetation com-
munities. Conserving Natura 2000 Rivers Monitoring Series No. 11,
English Nature, Peterborough, s. 30, http://www.riverlife.org.uk
LUCASSEN E. C. H. E. T., SMOLDERS A. J. P., ROELOFS J. G. M.
2000. Increased groundwater levels cause iron toxicity in
Glyceria fluitans
(L.). Aquatic Botany 66: 321–327.
MABERLY S. C., MADSEN T. V. 2002. Use of bicarbonate ions as
a source of carbon in photosynthesis by
Callitriche herma-
phroditica.
Aquatic Botany 73: 1–7.
MADSEN T. V. 1991. Inorganic carbon uptake kinetics of the stre-
am macrophyte
Callitriche cophocarpa
Sendt. Aquatic Bo-
tany 40: 321–332.
MADSEN T. V. CEDERGREEN N. 2002. Sources of nutrients to ro-
oted submerged macrophytes growing in a nutrient–rich stre-
am. Freshwater Biology 47: 283–291.
MADSEN T. V., CHAMBERS P. A. JAMES W. F., KOCH E. W., WEST-
LAKE D. F. 2001. The intewraction between water movement,
sediment dynamics and submersed macrophytes. Hydrobio-
logia 444: 71–84.
MADSEN T. V., HAHN P., JOHANSEN J. 1998. Effects of inorga-
nic carbon supply on the nitrogen requirement of two sub-
merged macrophytes, Elodea canadensis and
Callitriche
cophocarpa.
Aquatic Botany 62: 95–106.
MATUSZKIEWICZ W. 2001. Przewodnik do oznaczania zbioro-
wisk roÊlinnych Polski. PWN, Warszawa.
MIREK Z., PI¢KO–MIRKOWA H., ZAJÑC A., ZAJÑC M. Vascular
Plants of Poland – A Checklist. Krytyczna lista roÊlin naczynio-
wych Polski. http: //bobas.ib–pan.krakow.pl/czek/check.htm
NIELSEN S. L., SAND–JENSEN K.1993. Photosynthetic implica-
tions of heterophylly in
Batrachium peltatum
(Schrank)
Presl. Aquatic Botany 44: 361–371.
98
P
oradniki ochrony siedlisk i gatunk
ów
Wody s∏odkie i torfowiska
3260

99
Tytu∏ rozdzia∏u Tytu∏ rozdzia∏u Tytu∏ rozdzia∏u Tytu∏
P
oradniki ochrony siedlisk i gatunk
ów
NORMANN H. D. 1967. Experiments on the uptake of phosphate
by
Ranunculus fluitans
L Am. Arch. Hydrobiol. 33: 243–54
NOWAK A. (red.) 2001. Ostoje przyrody Natura 2000 w woje-
wództwie opolskim (koncepcja regionalna) . Prace Opolskie-
go Towarzystwa Przyjació∏ Nauk Wydz. III. Opole, s.173.
OBRDLÍK P., RAST G., NIEZNA¡SKI P., HAMPLOVÁ V., DÖPKE M.,
GÜNTHER–DIRINGER D. 2003. Graniczne meandry Odry –
fenomen o znaczeniu europejskim (raport za okres od mar-
ca 2001 do kwietnia 2003). WWF, Rastatt, s. 66.
O’HARE M. T., MURPHY K. J. 1999. Invertebrate hydraulic micro-
habitat and community structure in
Callitriche stagnalis
Scop. patches. Hydrobiologia 415: 169–176.
PROåKÓW J. 2004.
Batrachium penicillatum
Dumort. (
Ranun-
culaceae) na Dolnym Âlàsku. Acta Botanica Silesiaca 1:
000–000 (w druku)
PROåKÓW J., SZELÑG Z. 2001.
Batrachium penicillatum
Du-
mort. – W∏osienicznik (jaskier) p´dzelkowaty. W: Kaêmiercza-
kowa R., Zarzycki K. (red.): Polska czerwona ksi´ga roÊlin.
Paprotniki i roÊliny kwiatowe. Inst. Botaniki im. W. Szafera
PAN, Inst. Ochrony Przyrody PAN, Kraków: 148–149.
PUCHALSKI W. 2000. The capability of bottom communities to
decrease fluctuations of nutrient concentrations in small stre-
ams. Verh. Internat. Verein. Limnol. 27: 1371–1375.
RIIS T., BIGGS B. J. F. 2003. Hydrologic and hydraulic control of
macrophyte establishment and performance in streams. Lim-
nol. Oceanogr. 48: 1488–1497.
RIIS T., SAND–JENSEN K. 2001. Historical changes in species
composition and richness accompanying perturbation and
eutrophication of Danish lowland streams over 100 years.
Freshwater Biology 46: 269–280.
RIIS T., SAND–JENSEN K., LARSEN S. E. 2001. Plant distribution
and abundance in relation to physical conditions and location
within Danish stream systems. Hydrobiologia 448: 217–228.
RIIS T., SAND–JENSEN K., VESTERGAARD O. 2000. Plant com-
munities in lowland Danish streams: species composition
and environmental factors. Aquatic Botany 66: 255–272.
ROBACH F., THIÉBAULT G., MULLER S., TRÉMOLIÉRES M. 1996.
A reference system for continental running waters: plant
communities as bioindicators of increasing eutrophication in
alkaline and acidic waters in north–eastern France. Hydro-
biologia 340: 67–76.
RUTKOWSKI L. 1993. Nowe i ma∏o znane zbiorowiska roÊlinne Bo-
rów Tucholskich. W: Rejewski M., Nienartowicz A., Boiƒski M.:
Bory Tucholskie. Walory przyrodnicze – Problemy ochrony –
Przysz∏oÊç. UMK, Toruƒ: 79–88.
SAND–JENSEN K. 1998. Influence of submerged macrophytes
on sediment composition and near–bed flow in lowland stre-
ams. Freshwater Biology 39: 663–679.
SAND–JENSEN K. ANDERSEN K., ANDERSEN T. 1999. Dynamic
properties of recruitment, expansion and mortality of macro-
phyte patches in streams. Internat. Rev. Hydrobiol. 84:
497–508.
SAND–JENSEN K., PEDERSEN O. 1999. Velocity gradients and
turbulence around macrophyte stands in streams. Freshwater
Biology 42: 315–328.
SCHNITZLER A., EGLIN I., ROBACH F., TRÉMOLIÉRES M. 1996.
Response of aquatic macrophyte communities to levels of
P and N nutrients in an old swamp of the upper Rhine plain
(Eastern France). Écologie 27: 51–61.
THIÉBAUT G., MULLER S. 1999. A macrophyte communities se-
quence as an indicator of eutrophication and acidification le-
vels in weakly mineralised streams in north–eastern France.
Hydrobiologia 410: 17–24.
TRÉMOLIERES M., EGLIN I., ROECK U., CARBIENER R. 1993. The
exchange process between river and groundwater on the
central Alsace floodplain (eastern France) : I. The case of the
canalised river Rhine. Hydrobiologia 254: 133–148.
TURA¸A K. 1970. Cytotaxonomical studies in
Ranunculus flu-
itans
Lam. and
R. penicillatus
(Dumort.) Bab. from lower
Silesia (Poland) . Preliminary report. Acta Biol. Cracov., Ser.
Bot. 13: 119–123+Plat. 20–23.
URBAN D. 1996. Zbiorowiska wodne i szuwarowe doliny Ciemi´-
gi. Annales UMCS, Sectio E, 17: 119–137.
WERPACHOWSKI C. 2000. Lista roÊlin naczyniowych Kotliny Bie-
brzaƒskiej ze szczególnym uwzgl´dnieniem Biebrzaƒskiego
Parku Narodowego. Parki Narodowe i Rezerwaty Przyrody,
19.4: 19–52.
WHITE D. S., HENDRICKS S. P. 2000. Lotic macrophytes and sur-
face–subsurface exchange processes. In: Jones J. B. , Mulhol-
land P. J. (eds.) Streams and ground waters, Academic Press,
San Diego: 363–379.
WIEGLEB G., HERR W. 1985. The occurrence of communities with
species of
Ranunculus
subgenus
Batrachium
in central Eu-
rope – preliminary remarks. Vegetatio 59: 235–241.
WRIGHT J. F., CLARKE R. T., GUNN R. J. M., WINDER J. M.,
KNEEBONE N. T., DAVY–BOWKER J. 2003. Response of
the flora and macroinvertebrate fauna of a chalk stream
site to changes in management. Freshwater Biology 48,
s. 894–911.
ZALEWSKI M., PUCHALSKI W., FRANKIEWICZ P., BIS B. 1993. Ri-
parian ecotones and fish communities in rivers – intermedia-
te complexity hypothesis. In: Cowx I. G. (Ed.): Rehabilitation
of Inland Fisheries, Fishing New Books, Blackwell Sci. Publ.,
Oxford, s.152–160.
ZA¸USKI T. 2003. Materia∏y do planu ochrony rezerwatu „Piekie∏-
ko”. Biuro Us∏ug Ekologicznych i Urzàdzeniowo-LeÊnych
OPERAT, Toruƒ.
˚UKOWSKI W. 2001.
Groenlandia densa
(L.) Fourr. – Rdestni-
ca g´sta. W: Kaêmierczakowa R., Zarzycki K. (red.) Polska
czerwona ksi´ga roÊlin. Paprotniki i roÊliny kwiatowe. Inst.
Botaniki im. W. Szafera PAN, Inst. Ochrony Przyrody PAN,
Kraków: 407–408.
Wojciech Puchalski
Nizinne i podgórskie rzeki ze zbiorowiskami w∏osieniczników
3260

100
P
oradniki ochrony siedlisk i gatunk
ów
Wody s∏odkie i torfowiska
Wody s∏odkie i torfowiska
B. Opis podtypu
Nizinne i podgórskie rzeki ze
zbiorowiskami w∏osieniczników
Kod Physis: 24.4
Cechy diagnostyczne siedliska
Cechy obszaru
Siedlisko jest okreÊlane przez roÊlinnoÊç zanurzonà, przy-
twierdzonà do uziarnionych osadów mineralnych (piasek,
˝wir, drobne kamienie) na dnie koryt strumieni i rzek o in-
tensywnym przep∏ywie wody i zauwa˝alnym zasilaniu przez
wody podziemne lub wymianie wód mi´dzy strefà koryta
a interaktywnà strefà migrujàcych wód gruntowych (hypo-
reicznà). Wyst´puje na odcinkach erozyjnych i przejÊcio-
wych cieków wodnych niskiej lub Êredniej rz´dowoÊci, w ni-
˝ej po∏o˝onych odcinkach akumulacyjnych zanika.
Do najwa˝niejszych czynników Êrodowiskowych okreÊlajàcych
wyst´powanie, rozmieszczenie i struktur´ siedliska nale˝à:
¸agodne warunki klimatyczne w pó∏roczu zimowym.
Gatunki roÊlin tworzàce dominujàce zbiorowiska (rz´Êle,
w∏osieniczniki) nie tylko nie zamierajà (lub zamierajà jedy-
nie cz´Êciowo) na zim´, ale jeszcze w listopadzie zachowu-
jà aktywnoÊç fotosyntetycznà; mszaki
(Fontinalis, Scapa-
nia) sà zdolne do fotosyntezy równie˝ przez ca∏à zim´.
Wiele z reprezentowanych tu gatunków roÊlin nie tworzy zi-
mowych organów przetrwalnych, niekiedy równie˝ nie roz-
mna˝a si´ generatywnie, tak wi´c przezimowanie p´dów
wegetatywnych w niezamarzajàcym cieku jest warunkiem
przetrwania zimy. Z tego wzgl´du zasi´g geograficzny sie-
dliska w Polsce jest ograniczony do Pomorza, zachodniej
Wielkopolski i Âlàska.
Sezonowa stabilnoÊç hydrologiczna rzek. Wyst´pu-
jàce tu roÊliny z jednej strony wymagajà co najmniej
umiarkowanego przep∏ywu wody (zasilanie w pobierane
z wody przez liÊcie substancje biogenne, przeciwdzia∏anie
sedymentacji zawiesiny na powierzchni roÊlin), z drugiej -
zwi´kszony przep∏yw wezbraniowy powoduje mechanicznà
fragmentacj´ i odrywanie p´dów oraz erozj´ dna. Z tego
wzgl´du preferowane sà zimowe maksima odp∏ywu (gdy
p´dy roÊlin sà najs∏abiej rozwini´te), a w lecie brak wez-
braƒ spowodowanych nawalnymi deszczami i brak d∏ugo-
trwa∏ych okresów ni˝ówek. Pod tym wzgl´dem najkorzyst-
niejsze warunki wyst´pujà w rzekach Pomorza.
Zasilanie koryt wodami podziemnymi. Ten czynnik
∏àczy w sobie kilka mechanizmów. Du˝a retencja gruntowa
w zlewni, warunkujàca ten typ zasilania, oprócz czynników
klimatycznych decyduje o wy˝ej opisanej stabilnoÊci hydro-
logicznej. W okresie zimowym dop∏yw wód podziemnych
podnosi temperatur´ wody koryta rzecznego i zapobiega
jego zamarzaniu. Dlatego te˝ na obszarach o klimacie kon-
tynentalnym wyst´powanie omawianego siedliska jest
ograniczone do cieków êródliskowych. Wody podziemne
dostarczajà te˝ niezb´dnych dla roÊlin substancji biogen-
nych, a ich intensywna wymiana zapobiega ich odtlenieniu
i powstawaniu w nich toksycznych zredukowanych zwiàz-
ków (amoniak, siarkowodór, Fe
++
). Na tej zasadzie najlepiej
3260
1
Podwodna forma potocznika wàskolistnego (
Berula erecta fo. submersa) w korycie rzeki Grabowej (Polanów)

101
Tytu∏ rozdzia∏u Tytu∏ rozdzia∏u Tytu∏ rozdzia∏u Tytu∏
P
oradniki ochrony siedlisk i gatunk
ów
Nizinne i podgórskie rzeki ze zbiorowiskami w∏osieniczników
wykszta∏cone zbiorowiska roÊlinne obserwuje si´ w rzekach
charakteryzujàcych si´ najwi´kszym Êrednim odp∏ywem
jednostkowym w przeliczeniu na km
2
powierzchni zlewni
(Grabowa na Pomorzu) . Dobrze te˝ rozwijajà si´ w inten-
sywnie zasilanych wy∏àcznie wodami podziemnymi starych
korytach rzecznych przy obiektach hydroenergetycznych,
gdzie wody powierzchniowe zosta∏y poprowadzone równo-
leg∏ym kana∏em derywacyjnym. Dla zrealizowania zasilania
podziemnego dno rzeki musi byç zbudowane z osadów
o du˝ym przewodnictwie hydrologicznym; dno ilaste lub or-
ganiczne wyklucza rozwój omawianego siedliska. Krytycz-
nym okresem dla przetrwania roÊlin mo˝e byç sucha jesieƒ,
gdy du˝a zawartoÊç zwiàzków organicznych z dekompozy-
cji roÊlin làdowych, niedostatecznie rozcieƒczona przez wo-
dy gruntowe, prowadzi do ich odtlenienia i toksycznoÊci
zredukowanych rozpuszczonych substancji.
Dost´p Êwiat∏a. Wi´kszoÊç roÊlin naczyniowych charak-
terystycznych dla tego siedliska wymaga dobrych warun-
ków Êwietlnych i s∏abo si´ rozwija na odcinkach zacienio-
nych przez drzewa i wysokà zielnà roÊlinnoÊç brzegowà.
Dost´p Êwiat∏a ma szczególne znaczenie w przypadku do-
p∏ywu zredukowanych wód podziemnych o du˝ej zawarto-
Êci materii organicznej, prowadzàcego do odtlenienia stre-
fy korzeniowej roÊlin w osadach dennych. Intensywna foto-
synteza staje si´ wtedy warunkiem doprowadzenia tlenu
do osadów i przeciwdzia∏ania toksycznoÊci wody Êródosa-
dowej. W zacienionych odcinkach rzek o niskiej trofii do-
brze rozwijajà si´ mszaki i krasnorosty.
Dost´pnoÊç wolnego dwutlenku w´gla. Wi´kszoÊç
wyst´pujàcych w tym siedlisku roÊlin ca∏kowicie zanurzo-
nych mo˝e asymilowaç jedynie wolny CO
2
i nie ma zdolno-
Êci przyswajania wodorow´glanów. Z tà w∏aÊciwoÊcià zwià-
zane sà opisywane z Europy Zachodniej odmiennoÊci flory-
styczne podtypów zwiàzanych z dominujàcymi w zlewni
ska∏ami krystalicznymi i osadowymi (innymi istotnymi czyn-
nikami mo˝e byç równie˝ zawartoÊç wapnia, odczyn i zwià-
zana z niskim pH potencjalna toksycznoÊç glinu i metali
ci´˝kich). Efektywnym êród∏em CO
2
sà wody podziemne
zasilajàce koryto rzeki lub procesy dekompozycji na obsza-
rach podmok∏ych w dolinie (Biebrza). Wyczerpanie CO
2
przez fitoplankton rozwijajàcy si´ w eutroficznych jeziorach
i zbiornikach zaporowych, po∏àczone tak˝e ze wzrostem
temperatury wody, powoduje zanik omawianego siedliska
na odcinkach rzek poni˝ej zbiorników wód stojàcych.
Dost´pnoÊç zwiàzków biogennych (N, P). Rozpusz-
czone nieorganiczne formy azotu i fosforu pobierane sà
zarówno z wody poprzez liÊcie, jak z osadów przez korze-
nie. Pozbawione korzeni p´dy oderwane od pod∏o˝a nie
obni˝ajà intensywnoÊci pobierania zwiàzków biogennych.
Istniejà obserwacje wskazujàce na zanikanie zbiorowisk
roÊlinnych omawianego siedliska w Polsce jako skutek eu-
trofizacji wód rzecznych, a z drugiej strony w Danii wyka-
zano, ˝e liczba jego stanowisk nie zmniejszy∏a si´ w ciàgu
ostatnich 100 lat mimo znacznie intensywniejszej eutrofi-
zacji ni˝ u nas. SprzecznoÊç ta wynika z faktu, ˝e wzrost
trofii wody rzeki powoduje intensywniejszy wzrost p´dów
roÊlin przy niezmienionej masie korzeni. To z kolei zwi´k-
sza podatnoÊç na mechaniczne niszczenie i wyrywanie ro-
Êlin przez letnie fale wezbraniowe, charakterystyczne dla
niestabilnych hydrologicznie rzek klimatu kontynentalne-
go, a rzadkie w klimacie oceanicznym.
Poszczególne gatunki roÊlin zbiorowisk w∏osieniczników
i rz´Êli charakteryzujà si´ zró˝nicowanà efektywnoÊcià i dy-
namikà asymilacji ró˝nych form azotu (NH
4
+
, NO
3
–
i NO
2
–
)
i fosforanów. Osiàgajà wysokà aktywnoÊç metabolicznà
w ni˝szych temperaturach ni˝ inne hydrofity, stàd ich liczne
wyst´powanie w wodach ch∏odnych, klasyfikowanych zwykle
jako oligo–mezotroficzne. Niedobór substancji biogennych
w wodach o niskiej trofii jest kompensowany ich dop∏ywem
z wód podziemnych i wymianà wody w szybko p∏ynàcym cie-
ku. W rzekach cieplejszych latem (i równoczeÊnie opisywa-
nych jako eutroficzne) przegrywajà konkurencj´ z bardziej
efektywnymi w wy˝szej temperaturze rdestnicami i moczar-
kà, które ponadto nie sà uzale˝nione od dost´pnoÊci wolne-
go CO
2
. Ten czynnik staje si´ kolejnà przyczynà zaniku
omawianego siedliska w wodach o wysokiej produktywno-
Êci.
PrzezroczystoÊç wody. Ma∏y ∏adunek zawiesiny niesiony
wodami rzeki jest istotnym czynnikiem warunkujàcym ist-
nienie siedliska. M´tnoÊç wody i sedymentujàce na po-
wierzchni p´dów czàstki pogarszajà warunki Êwietlne dla
roÊlin, zawiesina glinokrzemianowa absorbuje limitujàce
rozwój roÊlin fosforany, a drobne czàstki osadzajàce si´ na
dnie powodujà jego kolmatacj´ i ograniczenie dop∏ywu
wód podziemnych. Stagnacja wód w osadach i brak wy-
miany gazowej prowadzi do odtlenienia osadów i ich tok-
sycznoÊci dla roÊlin.
Fizjonomia i struktura zbiorowisk
Na siedlisko sk∏adajà si´ p∏aty ró˝nych (cz´sto niedok∏adnie
zdefiniowanych fitosocjologicznie) zbiorowisk roÊlin zakorze-
nionych w dnie cieku (lub przytwierdzonych do pod∏o˝a –
w przypadku mszaków i glonów), o p´dach zanurzonych
w wodzie i unoszàcych si´ w pràdzie wody. Poszczególne ga-
tunki roÊlin mogà równie˝ wytwarzaç liÊcie p∏ywajàce po po-
wierzchni wody (w warunkach spowolnionego przep∏ywu wo-
dy) lub p´dy cz´Êciowo wystajàce nad powierzchni´ wody
(przy brzegu cieku), co mo˝e uniezale˝niaç roÊliny od wolne-
go CO
2
rozpuszczonego w wodzie. P∏aty roÊlin w nurcie, po-
przedzielane liniami szybkiego pràdu wody, sà zwykle jedno-
gatunkowe, na tym samym odcinku rzeki mogà wyst´powaç
obok siebie p∏aty ró˝nych gatunków. Pokrycie dna p∏atami
roÊlin mo˝e wynosiç od kilku do ponad 80%. P´dy roÊlin mo-
gà mieç ró˝nà d∏ugoÊç – od kilku cm mszaków (
Scapania sp.)
lub kilkunastu cm kontrastujàco jasnozielonych rz´Êli do kil-
ku metrów w przypadku w∏osienicznika rzecznego
Batra-
chium fluitans; ten gatunek, o ile w ogóle wyst´puje, stanowi
g∏ówny element strukturotwórczy siedliska.
Podstawowà formà ˝yciowà roÊlin w tym siedlisku sà roÊliny
zimozielone, niewytwarzajàce organów spichrzowych ani
3260
1

form przetrwalnikowych, rozmna˝ajàce si´ wegetatywnie
przez fragmentacj´ p´dów i ich przytwierdzanie do pod∏o˝a
w innym miejscu i zakorzenianie. Rozmna˝anie generatywne
ma mniejsze znaczenie, o ile w ogóle wyst´puje. Oprócz tej
formy ˝yciowej mogà wyst´powaç zanurzone w wodzie eko-
typy (fo.
submersa) gatunków roÊlin posiadajàcych k∏àcza,
których formy typowe wyst´pujà w zbiorowiskach brzegów
wód lub w wodach stojàcych; w tym siedlisku rozmna˝ajà si´
one równie˝ wegetatywnie lub przez nasiona przeniesione
z ich typowych siedlisk. Niekiedy obserwuje si´ w strefach za-
stoiskowych p∏ywajàce na powierzchni p´dy rz´s.
W siedlisku mogà wyst´powaç równie˝ osiàgajàce
znaczne rozmiary p∏aty glonów makroskopowych. Naj-
cz´Êciej spotyka si´ przytwierdzone do twardego pod∏o-
˝a, zwykle kamieni, plechy krasnorostów –
Batracho-
spermum sp. lub Lemanea sp., lub zielenicy Cladophora
glomerata oraz rozwijajàce si´ na dnie piaszczystym p∏a-
ty
Vaucheria sp. Na powierzchni wi´kszych kamieni za-
uwa˝alne sà krwistoczerwone plamy krasnorostu
Hilde-
brandtia rivularis. Na nieporoÊni´tym przez roÊliny
i nieerodowanym piaszczystym dnie cieków, szczególnie
w miejscach umiarkowanego wysi´ku bogatych w fosfor
wód podziemnych, wyraênie widoczny jest bràzowy nalot
wielogatunkowych zbiorowisk okrzemek ze zwykle domi-
nujàcymi gatunkami z rodzajów
Nitzschia i Navicula.
W miejscach wyp∏ywu zredukowanych wód podziemnych
tworzà si´ obfite naloty bakterii siarkowej
Lamprocystis
roseo–persicina.
Omawiane siedlisko jest miejscem szczególnie licznie,
w porównaniu z innymi siedliskami rzecznymi, wyst´pu-
jàcej fauny bezkr´gowców wodnych. Poszczególne ich
gatunki wykazujà zauwa˝alne preferencje w zale˝noÊci
od charakterystycznej dla gatunku roÊliny morfologii p´-
dów i ich aktualnej produkcji. Najwa˝niejszymi repre-
zentantami fauny sà skorupiaki (
Gammarus sp., Asellus
sp.), ma∏e ma∏˝e (
Sphaeriidae), larwy j´tek, widelnic,
chruÊcików oraz muchówek z rodziny
Simmulidae i Chi-
ronomidae. RoÊliny wodne nie stanowià znaczàcej bazy
pokarmowej; bezkr´gowce te ˝ywià si´ zakumulowanà
materià organicznà, mikroskopowymi glonami lub odfil-
trowywanà zawiesinà. Bezkr´gowce sà podstawowà ba-
zà pokarmowà dla ryb rzecznych.
Wi´kszoÊç gatunków ryb zwiàzanych z szybko p∏ynàcymi
rzekami preferuje siedliska nieporoÊni´te roÊlinnoÊcià,
z dnem ˝wirowym i kamienistym. Obszary dna poroÊni´-
te roÊlinnoÊcià sà êród∏em pokarmu dla ryb, schronie-
niem dla narybku, a tak˝e poÊrednio wp∏ywajà na po-
wstawanie preferowanego przez ryby uk∏adu bystrzy i za-
stoisk w nurcie rzeki i zró˝nicowania morfologii cieku.
W przypadku zarówno zbyt s∏abego, jak i zbyt intensyw-
nego poroÊni´cia koryta rzeki przez roÊlinnoÊç zag´sz-
czenie ryb spada, a w drugim przypadku cenne reofilne
gatunki ryb ust´pujà i zostajà zastàpione przez gatunki fi-
tofilne, o mniejszym znaczeniu w kategoriach funkcji eko-
systemu, w´dkarstwa i ochrony przyrody.
Reprezentatywne i dominujàce gatunki
roÊlin
I. RoÊliny zimozielone, o p´dach zanurzonych lub z p∏ywa-
jàcymi liÊçmi, nietworzàce organów spichrzowych:
w∏osienicznik rzeczny
Batrachium fluitans *
w∏osienicznik skàpopr´cikowy
Batrachium tricho–
phyllum *
w∏osienicznik wodny
Batrachium aquatile–
ssp. pseudofluitans *
w∏osienicznik p´dzelkowaty
Batrachium penicillatum1
rz´Êl hakowata
Callitriche hamulata *
rz´Êl d∏ugoszyjkowa
Callitriche cophocarpa
rdestniczka g´sta
Groenlandia densa *1
rdestnica nawodna
Potamogeton nodosus *3
zdrojek (mech) wodny
Fontinalis antypyretica
wàtrobowiec
Scapania
undulata
II. Formy zanurzone (fo.
submersa) roÊlin brzegowych
z apikalnà strefà wzrostu
potocznik wàskolistny
Berula erecta *
przetacznik bobowniczek
Veronica beccabunga *
przetacznik bobownik
Veronica anagalis–
aquatica *
rukiew wodna
Nasturtium officinale *
III. Formy zanurzone (fo.
submersa) roÊlin brzegowych
i wodnych z rozwini´tymi k∏àczami i bazalnà strefà wzrostu
∏àczeƒ baldaszkowaty
Butomus umbellatus *
je˝og∏ówka pojedyncza
Sparganium emersum
grà˝el ˝ó∏ty
Nuphar lutea *
IV. RoÊliny niecharakterystyczne, niekiedy licznie wyst´pujà-
ce w siedlisku lub w strefach przejÊciowych
moczarka kanadyjska
Elodea canadensis
rdestnica k´dzierzawa
Potamogeton crispus
rdestnica alpejska
Potamogeton alpinus
rdestnica przeszyta
Potamogeton perfoliatus
mi´ta wodna
Mentha aquatica
rz´sa drobna
Lemna minor
rz´sa trójrowkowa
Lemna trisulca
ramienica
Chara fragilis 2
Uwagi: * gatunki dominujàce i równoczeÊnie charaktery-
styczne w znaczeniu fitosocjologicznym
(1) gatunki roÊlin notowane w Polsce na pojedynczych,
reliktowych stanowiskach
(2) charakterystyczna dla siedlisk rzecznych w zachod-
niej Europie, w Polsce bardzo rzadka w tym siedlisku
(3) tylko liÊcie p∏ywajàce.
Odmiany siedliska
S∏abe poznanie siedliska na obszarze Polski nie pozwala
na Êcis∏e wyró˝nienie jego odmian. Na uwag´ zas∏uguje
potencjalna mo˝liwoÊç wyró˝nienia odmian, które po do-
k∏adniejszym zbadaniu mogà okazaç si´ nawet podtypami
siedliska.
• Odmiana typowa, z p∏atami co najmniej kilku gatun-
ków reprezentatywnych i dominacjà iloÊciowà w∏osie-
niczników, szczególnie w. rzecznego
B. fluitans. W stre-
102
P
oradniki ochrony siedlisk i gatunk
ów
Wody s∏odkie i torfowiska
Wody s∏odkie i torfowiska
3260
1

103
Tytu∏ rozdzia∏u Tytu∏ rozdzia∏u Tytu∏ rozdzia∏u Tytu∏
P
oradniki ochrony siedlisk i gatunk
ów
fach brzegowych rozwini´te p∏aty roÊlin z II i III grupy.
Wyst´puje w rzekach pó∏nocno–zachodniej i zachodniej
Polski.
• Odmiana zubo˝ona, z wyst´pujàcymi pojedynczymi
gatunkami w∏osieniczników, rz´Êli lub innych gatunków
I grupy, mo˝liwa obecnoÊç gatunków II i III grupy. Cz´sto
wyst´puje na Âlàsku i w obszarze granicznym area∏u wy-
st´powania.
• Odmiana kontynentalna, z dominacjà potocznika
i mo˝liwym wyst´powaniem gatunków II grupy oraz rz´-
Êli, zwykle bez w∏osieniczników, wyst´puje na odcinkach
êródliskowych cieków prawdopodobnie na terenie ca∏e-
go kraju, szczególnie na obszarach krasowych zbudowa-
nych ze ska∏ wapiennych (Jura Krakowsko–Cz´stochow-
ska, Roztocze) .
• Odmiana „liÊci wst´gowatych”. Tworzona g∏ównie
przez zbiorowiska roÊlin III grupy, tu nale˝à m.in. opisywa-
ne z Drawy zaroÊla podwodnej formy ∏àczenia baldaszko-
watego
Butomus umbellatus. Wyró˝niona ze wzgl´du na
odmiennà fizjonomi´, sposób zimowania (k∏àcza), rozmna-
˝ania i zasiedlania nowych area∏ów, mniejsze zag´szczenie
bezkr´gowców i wi´kszà odpornoÊç na wykaszanie roÊlin-
noÊci. Prawdopodobnie nie uzale˝niona od dop∏ywu wód
podziemnych.
• Odmiana wy˝ynna z obszarów ska∏ krystalicz-
nych. Dno kamieniste, poroÊni´te przez mszaki lub
B.
penicillatum, na odcinkach zacienionych mogà domino-
waç plechowe krasnorosty (
Batrachospermum monilifor-
me), tworzàc siedlisko o wyjàtkowo niskim zag´szczeniu
bezkr´gowców. Wody o niskiej trofii, ze szczególnym de-
ficytem fosforu i du˝ej podatnoÊci na zakwaszenie. Wy-
st´puje w Sudetach i Górach Âwi´tokrzyskich.
Mo˝liwoÊci pomy∏ki z innymi siedliskami
Wykaz siedlisk Natura 2000 nie obejmuje ˝adnych innych
siedlisk rozwijajàcych si´ w nurcie rzek.
Z innych, nieobj´tych ochronà zbiorowisk mo˝liwe jest pomy-
lenie w∏osieniczników z p∏atami rdestnicy grzebieniastej
Pota-
mogeton pectinatus o podobnej fizjonomii, rozwijajàcej si´
(wraz z innymi gatunkami rdestnic i moczarkà) w nurcie eutro-
ficznych rzek, zwykle na dnie ilasto–piaszczystym, nieprze-
puszczalnym dla wód podziemnych. Rdestnice obumierajà je-
sienià i zimujà w postaci organów przetrwalnikowych w dnie
rzeki.
Mo˝liwe jest równie˝ pomylenie wst´gowatych liÊci pod-
wodnych ∏àczenia baldaszkowatego
Butomus umbellatus
i je˝og∏owki pojedynczej
Spargasnium emersum z innymi
gatunkami roÊlin o podobnym pokroju, wyst´pujàcymi
w rzekach o wysokiej trofii na dnie ilastym lub mulistym
(strza∏ka wodna
Sagittaria sagittifolia, manna jadalna Gly-
ceria fluitans, manna mielec G. maxima).
W stojàcych, a niekiedy równie˝ wolno p∏ynàcych wodach
dolin rzecznych (starorzecza, kana∏y, rozlewiska) wyst´pujà
inne gatunki w∏osieniczników, rz´Êli i mchów, które mogà byç
mylone z charakterystycznymi dla omawianego siedliska.
PrzejÊcia mi´dzy „êródliskiem” (7220) a po∏o˝onym poni-
˝ej, stanowiàcym omawiane siedlisko „ciekiem êródlisko-
wym” mogà byç p∏ynne.
Identyfikatory fitosocjologiczne
Zwiàzek
Ranunculion fluitantis
Zespo∏y:
Ranunculetum fluitantis w∏osienicznika rzecznego
Ranunculo–Callitrichetum hamulatae rz´Êli
hakowatej i w∏osieniczników
Ranunculo–Sietum erecto–submersi podwodnej
formy potocznika i w∏osieniczników
Dynamika roÊlinnoÊci
RoÊlinnoÊç omawianego siedliska rozprzestrzenia si´ przez
mechaniczne odrywanie fragmentów p´dów (przez wody
wezbraniowe, zwierz´ta, sp∏ywajàce rzekà ga∏´zie drzew)
i nast´pne ich zakotwiczenie w dnie (z wykorzystaniem nie-
równoÊci dna lub istniejàcych przeszkód w nurcie (kamie-
nie, ga∏´zie) i ukorzenienie w sprzyjajàcych warunkach
(opisanych wy˝ej). Masowe powstawanie nowych p∏atów
nast´puje po powodziach nawet w dolnych odcinkach
rzek; wraz z póêniejszym ograniczeniem dop∏ywu wód
podziemnych (lub pogorszeniem ich jakoÊci) i kolmatacjà
dna przez sedymentujàce osady drobnoczàsteczkowe p∏a-
ty te w wi´kszoÊci zanikajà.
Tworzàcy si´ p∏at przyczynia si´ do sedymentacji wleczonego
rumowiska i zawiesiny, przez co poziom powierzchni dna we-
wnàtrz p∏atu podnosi si´, a z czasem równie˝ zmienia si´ je-
go sk∏ad granulometryczny na bardziej drobnoczàsteczkowy,
równie˝ o wi´kszej zawartoÊci materii organicznej. Mo˝e to
doprowadziç do kolmatacji dna i zmniejszenia dop∏ywu wód
podziemnych lub ich chemicznego zredukowania, co z kolei
prowadzi do rozpadu i zaniku p∏atu; oderwane p´dy zako-
rzeniajà si´ w innym miejscu. Równie˝ mechaniczna erozja
dna z jednej strony p∏atu i akumulacja z drugiej przyczynia
si´ do stopniowej „w´drówki” p∏atu. Rozprzestrzenianie si´
p∏atów na ca∏à szerokoÊç cieku prowadzi do zwi´kszenia ich
podatnoÊci na erozj´ nawet przy mniejszych wezbraniach.
Znaczàca erozja p∏atów obserwowana jest w okresie zimo-
wym (zw∏aszcza przy wystàpieniu zlodzenia), nie nast´epu-
je jednak ich masowe zamieranie na zim´.
Relacja mi´dzy powierzchnià zaj´tà przez p∏aty poszczególnych
gatunków jest zmienna i zale˝na od zmieniajàcych si´ warun-
ków hydrologicznych. Generalnie, w∏osieniczniki dominujà
w latach i strefach o wysokim przep∏ywie na osadach gruboziar-
nistych, rz´Êle w warunkach s∏abszego przep∏ywu i na przepusz-
czalnych osadach drobnoziarnistych. W latach suchych wynu-
rzone formy typowe gatunków amfibiotycznych przewa˝ajà nad
formami zanurzonymi i dochodzi do wytworzenia nasion.
Brak jest danych wskazujàcych na istnienie naturalnej, kierun-
kowej sukcesji w tym siedlisku, uzale˝nionym od umiarkowane-
go poziomu perturbacji. Rozwój roÊlinnoÊci brzegowej (wyso-
Nizinne i podgórskie rzeki ze zbiorowiskami w∏osieniczników
3260
1
104
P
oradniki ochrony siedlisk i gatunk
ów
Wody s∏odkie i torfowiska
Wody s∏odkie i torfowiska
kich helofitów, a w szczególnoÊci krzewów i drzew) prowadzi do
znaczàcego ograniczenia, a nawet zaniku omawianego siedli-
ska; w odmianie wy˝ynnej na ska∏ach krystalicznych nast´puje
przejÊcie do dominacji krasnorostów.
Dynamika powiàzana z dzia∏alnoÊcià cz∏owieka b´dzie
uwzgl´dniona przy opisie zagro˝eƒ.
Siedliska przyrodnicze zale˝ne lub
przylegajàce
Brak siedlisk zale˝nych.
Siedliska przylegajàce: zbiorowiska ∏àkowe, szuwarowe w kory-
cie rzeki (
Sparganio–Glycerion fluitantis, Physis 53.4) i na tera-
sach (
Magnocaricion, Physis 53.2) , nadrzeczne zbiorowiska
welonowe (Physis 37.71), zaroÊla wierzbowe (Physis 44.12)
oraz zbiorowiska leÊne: ∏´gi (91E0) i olsy (Physis 44.3) dolin
rzecznych, inne lasy zboczy dolin. Zbiorowiska êródliskowe
(7220) mogà wyst´powaç powy˝ej, sà êród∏em zasilania w wo-
d´, mogà byç êród∏em diaspor wyst´pujàcych gatunków roÊlin
oraz refugium dla zwierzàt bezkr´gowych. Innym siedliskiem
przylegajàcym mogà byç ˝wirowe brzegi z roÊlinnoÊcià pionier-
skà przy korytach rzek podgórskich (Physis 24.22) lub brzegi
piaszczyste w dolinach rzek nizinnych (Physis 24.3), wa˝ne ze
wzgl´du na ich znaczenie w migracji wód podziemnych.
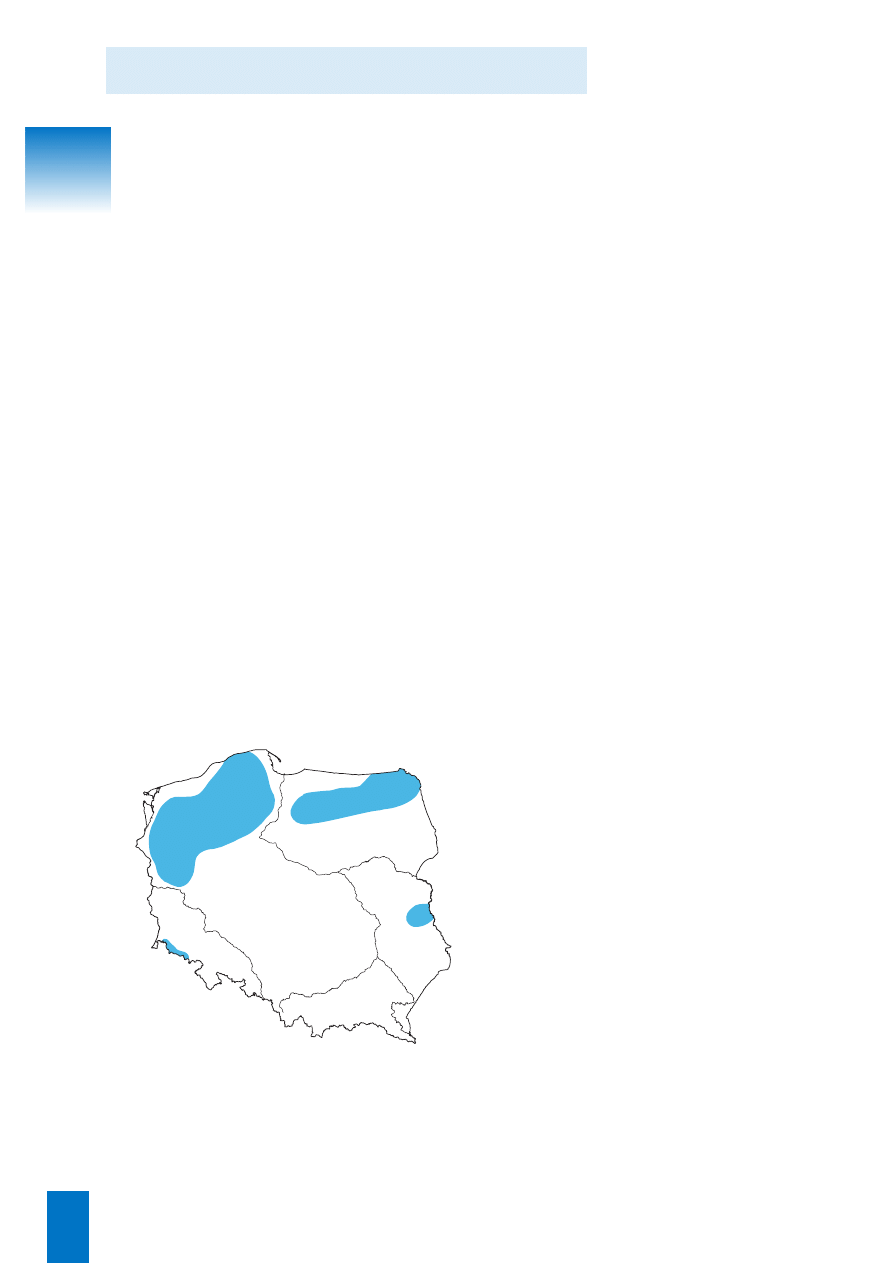
Rozmieszczenie geograficzne
i mapa rozmieszczenia
Na mapie przedstawiono prawdopodobny zasi´g odmia-
ny typowej i zubo˝onej.
Znaczenie ekologiczne i
biologiczne
• Znaczàce (nawet 10–krotne) zmniejszenie tempa erozji
wg∏´bnej koryta rzeki.
• Poprawa jakoÊci wody – rola aktywnego w ciàgu ca∏ego
roku filtru zatrzymujàcego zwiàzki biogenne i zawiesin´
z wód koryta rzek i wód podziemnych poprzez wbudowy-
wanie w mas´ roÊlin, osadów i larw owadów, które po
przeobra˝eniu wynoszà je ze Êrodowiska wodnego.
• forowanie st´˝eƒ fosforanów w wodzie – stabilizacja Êro-
dowiska chemicznego rzeki.
• Rola siedliskotwórcza dla du˝ego zag´szczenia bezkr´-
gowców wodnych – ró˝norodnoÊç fauny, pokarm dla ryb.
• Preferowane strefy wyst´powania i rozrodu ryb, m.in.
pstràga potokowego (
Salmo trutta m. fario).
• Siedlisko (i tworzàce je gatunki roÊlin) na wschodniej
granicy zasi´gu przebiegajàcej przez Polsk´.
• Wy∏àczne wyst´powanie gatunków roÊlin: krytycznie za-
gro˝onego (CR,
Groenlandia densa) i nara˝onego (VU,
Batrachium penicillatum) oraz cz´Êciowe – gatunku obj´-
tego cz´Êciowà ochronà gatunkowà
(Nuphar lutea) .
• Wyst´powanie gatunków bezkr´gowców, minogów, ryb, pta-
ków i ssaków z za∏àcznika II Dyrektywy Siedliskowej i Za∏àczni-
ka I Dyrektywy Ptasiej (w cz´Êci obj´tych ochronà gatunkowà
w Polsce oraz wpisanych na polskà czerwonà list´ zwierzàt)
Gatunki z za∏àcznika II Dyrektywy
Siedliskowej
minóg rzeczny
Lampetra fluviatilis
OG cz´Êc.
minóg strumieniowy
Lampetra planeri
OG
kie∏b Kesslera
Gobio Kessleri
OG
∏osoÊ atlantycki
Salmo salar
koza
Cobitis taenia
OG
g∏owacz bia∏op∏etwy
Cottus gobio
OG
bóbr
Castor fiber
OG cz´Êc.
wydra
Lutra lutra
OG
∏àtka ozdobna
Coenagrion ornatum OG CR
trzepla zielona
Ophiogomphus cecilia OG
skójka gruboskorupowa
Unio crassus
OG EN
Gatunki z Za∏àcznika I Dyrektywy Ptasiej
zimorodek
Alcedo atthis
OG
OG – gatunki obj´te ochronà gatunkowà w Polsce (OG –
ochrona Êcis∏a, OG cz´Êc. – ochrona cz´Êciowa)
Stany, w jakich znajduje si´ siedlisko
Stany uprzywilejowane
• Najbardziej zró˝nicowane gatunkowo stanowiska od-
miany typowej siedliska.
• Stanowiska, na których elementami flory sà rzadkie ga-
tunki reliktowe.
• Odcinki rzek, gdzie ochrona siedliska b´dzie elementem
ochrony cennych populacji ryb (lub ca∏e rzeki dla ryb
w´drownych).
• Strumienie i cieki êródliskowe zaroÊni´te przez zbiorowi-
sko z dominacjà potocznika; nie jest to szczególna oso-
bliwoÊç florystyczna, ale ochrona jest zalecana ze wzgl´-
du na wyjàtkowe znaczenie ekologiczne.
3260
1

105
Tytu∏ rozdzia∏u Tytu∏ rozdzia∏u Tytu∏ rozdzia∏u Tytu∏
P
oradniki ochrony siedlisk i gatunk
ów
Nizinne i podgórskie rzeki ze zbiorowiskami w∏osieniczników
Inne obserwowane stany
Nawet opisana powy˝ej odmiana typowa omawianego
siedliska stanowi stan zubo˝ony w porównaniu do stanów
charakterystycznych dla atlantyckiej strefy klimatycznej za-
chodniej Europy. Ró˝nice sà zauwa˝alne w:
• morfologii roÊlin: w Polsce tylko skarla∏e formy niektó-
rych gatunków (w tym
B. fluitans)
• braku we florze Polski wielu gatunków o atlantyckim za-
si´gu wyst´powania lub ograniczeniu ich wyst´powania
do nielicznych, izolowanych stanowisk reliktowych
• ograniczeniu wyst´powania niektórych charakterystycz-
nych dla tego rzecznego siedliska gatunków w Polsce je-
dynie do jezior o niskiej trofii
• braku innych gatunków tworzàcych siedlisko nie zwiàza-
nych z atlantyckà strefà klimatycznà
Zubo˝enie struktury siedliska wynika g∏ównie z odmienno-
Êci warunków klimatycznych. Zauwa˝alne obecnie i pro-
gnozowane dynamiczne zmiany klimatu w skali kontynen-
tu mogà w znaczàcy sposób przesuwaç granice zasi´gu
siedliska na terenie Polski i przekszta∏caç jego struktur´.
S∏abo poznane sà zbiorowiska, opisane tu prowizorycznie jako
odmiana kontynentalna, ró˝niàce si´ sk∏adem od typowych
siedlisk atlantyckich i ubo˝sze florystycznie. Nie przesàdza to
jednak, ˝e powinny byç uznane za mniej cenne z ekologiczne-
go punktu widzenia. Definicja omawianego siedliska jest na ty-
le szeroka, ˝e w niektórych krajach (np. W. Brytanii) w∏àcza si´
w jego zakres wi´kszoÊç zbiorowisk roÊlin zanurzonych, nieza-
le˝nie od wyst´powania wyró˝nionych gatunków charaktery-
stycznych. PodkreÊla si´ fakt, ˝e te same gatunki w ró˝nych stre-
fach swojego zasi´gu mogà mieç odmienne preferencje siedli-
skowe i funkcje ekologiczne.
Tendencje do przemian siedliska w
skali kraju i potencjalne zagro˝enia
Fragmentaryczne dane i brak ciàg∏oÊci obserwacji nie po-
zwalajà na opis przemian siedliska zachodzàcych w ró˝-
nych sytuacjach w perspektywie d∏ugoterminowej. Charak-
terystyczne cechy jego ekologii i biologii gatunków pozwa-
lajà na okreÊlenie potencjalnych zagro˝eƒ i kierunków an-
tropogenicznych przemian siedliska w ró˝nych sytuacjach
presji. Najwa˝niejsze zagro˝enia w skali zlewni mogà wy-
nikaç z:
1. Rozwoju intensywnej gospodarki rolnej w zlewni,
w szczególnoÊci wielkotowarowej hodowli zwierzàt zanie-
czyszczajàcej gleb´ i wody gruntowe, uprawy roÊlin z za-
stosowaniem wysokich dawek ∏atwo wyp∏ukiwanych z gle-
by nawozów i herbicydów.
2. Odprowadzania nieoczyszczonych lub niedostatecznie
oczyszczonych Êcieków bytowo–gospodarczych do rzeki
lub do gleby w dolinie (nawet z pojedynczych gospodarstw
po∏o˝onych nad rzekà).
Zanieczyszczenia organiczne wód podziemnych, prowa-
dzàce do ich odtlenienia, powodujà zahamowanie rozwoju
korzeni (co zwi´ksza podatnoÊç na mechaniczne niszczenie
przez wody wezbraniowe) i zamieranie p´dów roÊlin jesie-
nià (niemo˝liwoÊç przetrwania zimy). Wysoka trofia wody
przy ograniczonym przep∏ywie prowadzi do nadmiernego
zarastania koryta rzeki przez p∏aty roÊlin, w konsekwencji
do ograniczenia dost´pnej przestrzeni i bazy pokarmowej
dla ryb reofilnych oraz powstawania szkodliwych dla nich
nocnych deficytów tlenowych. Drastycznie wzrasta podat-
noÊç siedliska na perturbacje. Ca∏kowite pokrycie dna rze-
ki przez roÊliny nale˝y uznaç za objaw degradacji siedliska!
3. Prowadzenia gospodarki powodujàcej erozj´ wodnà
gleby, ze sp∏ukiwaniem zawiesiny do cieków wodnych,
w tym upraw roÊlin okopowych na stokach i eksploatacji
kopalin (piasków, ˝wirów i kredy) w dolinie z odprowadze-
niem wód o wysokiej m´tnoÊci do rzeki
4. Zabiegów melioracyjnych w dolinie, je˝eli przyczynià si´
one do ograniczenia zasobów lub jakoÊci wód podziem-
nych lub wzrostu zró˝nicowania sezonowego ich dop∏ywu
do koryta rzeki.
5. Kanalizacji koryta rzecznego, prowadzàcej do ujednolice-
nia struktury dna i ograniczenia interakcji z wodami hypore-
icznymi oraz zanieczyszczania koryta odpadami sta∏ymi.
Odwodnienie zlewni, obni˝enie poziomu i tempa migracji wód
podziemnych, erozja gleb i zastàpienie dop∏ywu gruntowego
sp∏ywem powierzchniowym powodujà w pierwszej kolejnoÊci
zmiany struktury siedliska z dominacji w∏osieniczników na do-
minacj´ rz´Êli; w efekcie mniejszà zdolnoÊç usuwania z wody
fosforanów (zwykle odpowiedzialnych za eutrofizacj´ wód)
i toksycznych azotynów. W dalszej kolejnoÊci zmniejszenie ob-
j´toÊci przep∏ywu rzek, sedymentacja osadów drobnoczàstecz-
kowych (ilastych i organicznych) przy równoczesnym wzroÊcie
trofii wód powodujà zanik lub zastàpienie zbiorowisk omawia-
nego siedliska przez zbiorowiska rdestnic (g∏ównie P. pectina-
tus) i moczarki. Ich p´dy jesienià zamierajà, a ich dekompozy-
cja jest zwiàzana z uwolnieniem du˝ego ∏adunku zwiàzków bio-
gennych. Obni˝enie obj´toÊci przep∏ywu cieku (zmniejszenie
wydajnoÊci êródlisk) prowadzi równie˝ do zastàpienia form za-
nurzonych gatunków grupy II i III przez ich formy typowe (wy-
nurzone) , a nast´pnie do zaniku gatunków grupy II.
6. Budowy zbiorników zaporowych na odcinkach rzek zaj-
mowanych przez siedlisko oraz powy˝ej strefy jego wyst´-
powania, a tak˝e podpi´trzania rzek w tej strefie.
Zagro˝enie o szczególnie istotnym znaczeniu, powoduje
natychmiastowe efekty zbli˝one do opisanych wy˝ej,
w wyniku wzrostu temperatury, spadku zawartoÊci CO
2
i sedymentacji zawiesiny.
7. Niew∏aÊciwego prowadzenia lub – niekiedy równie˝ –
zaniechania zabiegów piel´gnacji roÊlinnoÊci wodnej
i brzegowej.
Cz´ste wycinanie (grabienie) roÊlin w korycie rzeki mo˝e
prowadziç do zastàpienia gatunków grupy I przez je˝og∏ów-
k´. Zarastanie brzegów cieków przez roÊlinnoÊç drzewiastà
i wysokà zielnà w warunkach wysokiej trofii prowadzi do
zmniejszenia powierzchni p∏atów roÊlin zanurzonych
(w skrajnym przypadku do zaniku), a w niskiej – do zastà-
3260
1

pienia roÊlin naczyniowych przez mszaki i krasnorosty. We
wszystkich przypadkach w konsekwencji nast´puje znaczà-
ce zmniejszenie zag´szczenia fauny bezkr´gowców.
W skali regionalnej zagro˝enia siedliska mogà wynikaç ze
zmian klimatu w kierunku jego kontynentalizacji. Na granicy
zasi´gu mo˝na spodziewaç si´ tu szczególnej wra˝liwoÊci.
Ponadto zakwaszenie wód i gleb w zlewni na obszarach zbu-
dowanych ze ska∏ krystalicznych powoduje zauwa˝alnà elimi-
nacj´ mszaków; w okresie roztopów wiosennych wody mogà
osiàgaç letalny poziom toksycznoÊci dla ryb i bezkr´gowców.
U˝ytkowanie gospodarcze
i potencja∏ produkcyjny
Siedlisko nie jest bezpoÊrednio u˝ytkowane gospodarczo.
Jego istnienie i ochrona poÊrednio prowadzi do osiàgni´-
cia wa˝nych celów zwiàzanych z u˝ytkowaniem wód i wy-
maganych przez obowiàzujàce prawo lub wzgl´dy ekono-
miczne, takich jak:
• Ochrona i poprawa jakoÊci wód. Siedlisko to stanowi
najskuteczniejszy z mo˝liwych (w warunkach intensywne-
go przep∏ywu wody) filtrów biologicznych, redukujàcych
st´˝enia i ∏adunki zanieczyszczeƒ w wodzie koryta rzeki
i przesiàków hyporeicznych. W szczególnoÊci z parame-
trów decydujàcych o jakoÊci wody ulegajà obni˝eniu st´-
˝enia: wszystkich form fosforu i azotu, zawiesiny, chloro-
filu, agresywnego CO
2
, metali ci´˝kich, niektórych zwiàz-
ków organicznych. W odró˝nieniu od innych zbiorowisk
roÊlin, filtr ten jest aktywny w ciàgu ca∏ego roku, choç
mo˝liwe sà sezonowe ró˝nice jego skutecznoÊci.
• Przeciwdzia∏anie erozji dennej. Rozwój siedliska na odcin-
ku cieku obni˝a (nawet 10–krotnie) tempo erozji dna
w porównaniu z odcinkami nieporoÊni´tymi, prowadzi do
zatrzymywania wleczonego rumowiska i zawiesiny. Po-
Êrednio wp∏ywa na dolinowà retencj´ wody przez utrzymy-
wanie zwierciad∏a wód podziemnych w dolinie rzeki.
• Gwarancja dobrego stanu ekologicznego rzeki, wyma-
ganego zaleceniami Ramowej Dyrektywy Wodnej Unii
Europejskiej.
• Wzrost produkcji ryb i atrakcyjnoÊci w´dkarskiej cieku.
Realizowany poÊrednio przez produkcj´ bazy pokarmo-
wej i zró˝nicowanie mikrosiedliskowe cieku w warun-
kach cz´Êciowego pokrycia dna przez roÊlinnoÊç, sprzy-
ja rozwojowi populacji cennych gospodarczo gatunków
ryb – pstràga potokowego, lipienia, brzany i ∏ososia.
• Utrzymanie atrakcyjnoÊci turystyczno - rekreacyjnej rze-
ki, szczególnie dla turystyki kajakowej.
Nale˝y jednak zwróciç uwag´, ˝e nadmierny rozwój roÊlin-
noÊci w silnie zeutrofizowanych ciekach (równie˝ nieko-
rzystny z przyrodniczego punktu widzenia) mo˝e stwarzaç
zagro˝enia dla realizacji celów ochrony przeciwpowodzio-
wej (udro˝nienie cieku), zagospodarowania rybackiego
i w´dkarskiego (utrzymanie licznych populacji ryb ∏ososio-
watych) oraz ochrony zespo∏ów ryb reofilnych.
Ochrona
Przypomnienie o wra˝liwych cechach
Siedlisko utrzymujàce si´ w stabilnych hydrologicznie cie-
kach o intensywnym lub umiarkowanym pràdzie wody, za-
silanych niezanieczyszczonymi wodami podziemnymi
przez mineralne osady denne, z zapewnionà dost´pnoÊcià
wolnego dwutlenku w´gla i Êwiat∏a, z wodà nie przegrza-
nà, nie m´tnà i nie nadmiernie zeutrofizowanà. Kluczowe
znaczenie ma równie˝ ∏agodnoÊç klimatu – makroklimat
zbli˝ony do atlantyckiego lub mikroklimat êródlisk z ma∏y-
mi rocznymi amplitudami temperatury i zasilania w wod´.
Zalecane metody ochrony
Skuteczna ochrona siedliska mo˝e byç realizowana poprzez
komplementarne dzia∏ania prowadzone w skalach: ca∏ej
zlewni, doliny rzecznej oraz koryta rzecznego, oparte na roz-
poznaniu uwarunkowaƒ hydrogeologicznych i biogeoche-
micznych przynajmniej w skali doliny. Same dzia∏ania lokal-
ne zwykle nie przyniosà zadowalajàcych trwa∏ych rezultatów.
Do najwa˝niejszych zadaƒ ochronnych nale˝y zaliczyç:
W skali zlewni:
• Zlewniowà i dolinowà retencj´ wody w celu z∏agodzenia
zró˝nicowania przep∏ywu poprzez zwi´kszanie lesistoÊci
zlewni, ochron´ i odtwarzanie mokrade∏. Retencja zbiorni-
kowa z powierzchniowym odp∏ywem z zapory odniesie tu
skutki odwrotne do zamierzonych; w przypadku odp∏ywu
dennego ze stratyfikowanego zbiornika efektem mo˝e byç
nadmierny rozwój w∏osieniczników poni˝ej zapory i koniecz-
noÊç ich wycinania dla zachowania równowagi siedliska.
• Zalesienie górnych, I–rz´dowych odcinków cieków (po-
wy˝ej strefy wyst´powania siedliska) , o ile nie spowodu-
je to trwa∏ego odtlenienia wody w tych ciekach. Nie sto-
sowaç na obszarach podatnych na zakwaszenie.
• Ochron´ wód podziemnych przed zanieczyszczeniami,
w szczególnoÊci organicznymi, a tak˝e ograniczenie od-
p∏ywu substancji biogennych do wód gruntowych.
• Uregulowanie gospodarki Êciekowej – budowa i uspraw-
nienie oczyszczalni Êcieków, uszczelnienie szamb, likwi-
dacja odp∏ywów Êcieków z gospodarstw rolnych do do-
liny rzeki.
• Ochron´ gleb dolin rzecznych i ich stoków przed erozjà.
W skali doliny rzecznej:
• Koszenie ∏àk, wykaszanie helofitów, czyli wysokiej roÊlinnoÊci
brzegowej (je˝eli konieczne, równie˝ usuwanie nagromadzo-
nych drobnoczàsteczkowych osadów i k∏àczy szybko odra-
dzajàcych si´ helofitów), wycinanie nadbrze˝nych drzew
i krzewów od strony po∏udniowej i wschodniej; pozostawianie
roÊlinnoÊci drzewiastej i helofitów od strony pó∏nocnej, w stre-
fach przenikania wód koryta do gruntu oraz przy ujÊciach za-
nieczyszczonych cieków, sp∏ywów powierzchniowych i wód
podziemnych, je˝eli poprawi∏oby to ich zdolnoÊci samo-
oczyszczenia lub ograniczy∏oby dop∏yw zanieczyszczeƒ i za-
wiesiny. Nale˝y zwróciç równie˝ uwag´ na znaczenie kumu-
lujàcych si´ opad∏ych liÊci drzew w obni˝aniu trofii strumienia.
106
P
oradniki ochrony siedlisk i gatunk
ów
Wody s∏odkie i torfowiska
Wody s∏odkie i torfowiska
3260
1

• Tworzenie (odtwarzanie) równoleg∏ych do cieku ciàgów
starorzeczy lub kana∏ów lateralnych na skraju doliny,
gdy sp∏ywajàce z wysoczyzny wody podziemne i po-
wierzchniowe wnoszà du˝e ∏adunki substancji organicz-
nych, zawiesiny i zwiàzków biogennych.
• Renaturyzacja skanalizowanych koryt rzecznych w celu
zró˝nicowania struktury dna i zintensyfikowania wymia-
ny wód powierzchniowych i hyporeicznych.
• Poni˝ej ju˝ istniejàcych, stratyfikowanych zbiorników zapo-
rowych z mo˝liwoÊcià upustu dennego – odprowadzanie
wód hypolimnetycznych w okresach wysokiej produkcji fi-
toplanktonu i wysokiej temperatury wody w epilimnionie.
• Je˝eli budowa nowego zbiornika powy˝ej siedliska jest
konieczna, uwzgl´dniç mo˝liwoÊç uzyskania w nim stra-
tyfikacji termicznej i konstrukcji zapory z upustem den-
nym. Je˝eli nie spowoduje to innych ekologicznie niepo-
˝àdanych efektów, to tego typu zbiornik, stabilizujàcy
przep∏ywy poni˝ej zapory, w sprzyjajàcych warunkach
móg∏by nawet umo˝liwiç rozwój siedliska w rzece poza
klimatycznie uwarunkowanà granicà zasi´gu.
W skali koryta rzecznego:
• Utrzymywanie ma∏ych nierównoÊci dna jako miejsca
zakotwiczenia odrywanych p´dów roÊlin, pozostawia-
nie w korycie zwalonych pni i ga∏´zi drzew, je˝eli nie
tamujà przep∏ywu wody.
• Usuwanie du˝ych przeszkód, hamujàcych przep∏yw wo-
dy i prowadzàcych do zamulania dna koryta rzeki.
• Dosadzanie p´dów roÊlin w celu odtworzenia zniszczo-
nego siedliska: zwróciç uwag´ na niezb´dne warunki –
przep∏yw wody, dop∏yw i jakoÊç wód podziemnych,
struktur´ osadów, oÊwietlenie.
• Przerzedzanie nadmiernego pokrycia dna roÊlinnoÊcià
w zeutrofizowanych ciekach – jak wyjaÊniono wy˝ej, jest
to zabieg majàcy na celu ochron´ stabilnoÊci siedliska!
Najlepsze efekty, godzàce interesy ochrony siedliska,
ochrony przeciwpowodziowej i zespo∏ów ryb reofilnych ,
uzyskuje si´, przeprowadzajàc wycinanie roÊlin w uk∏a-
dzie warkocza diagonalnych ci´ç, uwzgl´dniajàcych roz-
mieszczenie istniejàcych p∏atów (rys. rycina B) .
• Naturalnym sposobem redukcji nadmiernie rozrastajàcej si´
roÊlinnoÊci zanurzonej w zeutrofizowanej rzece mo˝e byç
pozostawienie lub nawet dosadzenie roÊlinnoÊci drzewiastej
przy brzegach, co ograniczy dost´p Êwiat∏a do koryta.
• Usuwanie nadmiaru zakumulowanych na dnie cieku osa-
dów ilastych i organicznych (mechaniczne lub przez regu-
lacj´ spadku rzeki) w przypadku, gdy obserwuje si´ zaha-
mowanie rozwoju p∏atów typowych dla siedliska gatunków
roÊlin i sukcesj´ w kierunku dominacji rdestnic (
P. pectina-
tus, P. perfoliatus, P. crispus), moczarki i je˝og∏ówki.
• Utrzymanie mo˝liwie stabilnego poziomu wody, obj´to-
Êci i pr´dkoÊci przep∏ywu; przeciwdzia∏anie erozji wg∏´b-
nej. W przypadku, gdy nastàpi∏o d∏ugotrwa∏e, nieodwra-
calne zmniejszenie obj´toÊci przep∏ywu, mo˝na zmniej-
szyç szerokoÊç koryta lub zwi´kszyç, gdy pr´dkoÊç prze-
p∏ywu jest zbyt du˝a.
• W systemach hydroenergetycznych, gdzie odci´te stare ko-
ryta rzeczne zasilane wodami podziemnymi zostajà zasie-
dlone przez rz´Êle i w∏osieniczniki, nale˝y zastosowaç ich
okresowe, krótkotrwa∏e przep∏ukiwanie wodà ze zbiornika
w celu usuni´cia nadmiaru osadów organicznych.
Inne czynniki mogàce wp∏ywaç na sposób
ochrony
Potrzeba uwzgl´dnienia wymagaƒ ochrony przeciwpowodzio-
wej: niedopuszczanie do nadmiernego zarastania cieku (prze-
ciwdzia∏anie eutrofizacji lub mechaniczne usuwanie roÊlin),
cz´Êciowe usuwanie roÊlinnoÊci brzegowej (sprzyjajàce równie˝
rozwojowi siedliska).
Zadania ochrony ryb ∏ososiowatych: równie˝ utrzymywanie
umiarkowanego pokrycia dna przez roÊlinnoÊç, z zastrze˝e-
niem powstrzymywania wycinania roÊlin i innych prac regula-
cyjnych w okresie tar∏a i rozwoju m∏odego narybku, najlepszy
okres lipiec–wrzesieƒ. Podobnie przy realizacji celów ochrony
innych gatunków ryb reofilnych.
Ochrona populacji wydry i zimorodka: pozostawiç fragmenty
brzegów poroÊni´te roÊlinnoÊcià drzewiastà.
Mo˝liwy jest lokalny konflikt mi´dzy wskazaniami dzia∏aƒ
ochronnych omawianego siedliska i zaleceniami utrzymania ro-
ÊlinnoÊci drzewiastej na brzegach rzeki (wskazania ochrony nad-
rzecznych siedlisk ∏´gowych, zapobieganie erozji bocznej koryta)
– do rozwiàzania przy planowaniu ochrony przez tworzenie mo-
zaiki siedlisk, uwzgl´dniajàcej lokalne przyrodnicze i gospodar-
cze uwarunkowania optymalizacji gospodarki przestrzennej.
W przysz∏oÊci mo˝e pojawiç si´ potrzeba dostosowania tech-
nik ochrony do efektów regionalnych i globalnych wieloletnich
zmian klimatycznych i hydrologicznych. W tym kontekÊcie na-
le˝y ju˝ obecnie objàç szczególnà ochronà strefy wysi´ków
wód podziemnych przy brzegach rzek, w zastoiskach i w sta-
rorzeczach, gdzie poprzez umo˝liwienie generatywnego roz-
mna˝ania roÊlin (dla grupy II i III – rozwoju form typowych)
wzrasta prawdopodobieƒstwo otrzymania genotypów lepiej
107
Tytu∏ rozdzia∏u Tytu∏ rozdzia∏u Tytu∏ rozdzia∏u Tytu∏
P
oradniki ochrony siedlisk i gatunk
ów
Nizinne i podgórskie rzeki ze zbiorowiskami w∏osieniczników
3260
1
P∏aty roÊlin
Miejsce dosadzania roÊlin
Bez wycinania
Dwie diagonalne linie wyci´cia
Jedna meandrujàca linia wyci´cia
A
B
C
Rysunek wed∏ug zamieszczonego w pracy: Fjorback C. , Kronvang
B., 2002. Interaction between aquatic macrophytes and stream hy-
draulics in a Danish stream: implications of different weed–cutting
methods. Verh. Internat. Verein. Limnol. 28: 1299–1303.

108
P
oradniki ochrony siedlisk i gatunk
ów
Wody s∏odkie i torfowiska
Wody s∏odkie i torfowiska
przystosowujàcych si´ do warunków Êrodowiska. Dzia∏ania
ochronne powinny zmierzaç do ochrony jakoÊci wód pod-
ziemnych oraz przeciwdzia∏ania sedymentacji frakcji ilastej
i organicznej w tych strefach i ich nadmiernemu zacienianiu.
Przyk∏ady obszarów obj´tych dzia∏aniami
ochronnymi
Bierna ochrona rezerwatowa, np. Drawieƒski Park Narodowy,
rezerwat „S∏onawy” na We∏nie, Rezerwat na Rzece Grabowej
Monitoring populacj
i Batrachium penicillatum w Sudetach.
Eksperymentalne wyci´cie drzew i krzewów wzd∏u˝ brzegu
górnej Lubrzanki w Górach Âwi´tokrzyskich (1993).
Inwentaryzacje, doÊwiadczenia,
kierunki badaƒ
• Badania taksonomiczno–morfologiczne prowadzàce do
opracowania kluczy do oznaczania gatunków z rodzajów
Batrachium i Callitriche w stanie wegetatywnym, a tak˝e
popularne opisy form zanurzonych roÊlin brzegowych
• Badania rozmieszczenia gatunków (zw∏aszcza nierozró˝-
nialnych z w/w. rodzajów) i struktury zbiorowisk roÊlin-
nych siedliska na tle warunków Êrodowiskowych, opra-
cowanie i rewizja systemu jednostek syntaksonomicz-
nych.
• Powiàzanie wyst´powania, charakterystyki i produktyw-
noÊci dominujàcych gatunków roÊlin z czynnikami hy-
drologicznymi, klimatycznymi i troficznymi, ze szczegól-
nym uwzgl´dnieniem interakcji z wodami podziemnymi
i hyporeicznymi.
• Dynamika p∏atów roÊlin: procesy zakorzeniania, wzrostu
p∏atu i jego rozpadu, sedymentacji osadów i kolmatacji
dna na tle parametrów hydrodynamicznych; okreÊlenie
krytycznych dla poszczególnych roÊlin parametrów hydro-
dynamiki cieku z uwzgl´dnieniem warunków troficznych.
• Strategie ˝yciowe gatunków roÊlin w odniesieniu do
zró˝nicowanych warunków trofii i perturbacji w ró˝nych
porach roku, z uwzgl´dnieniem uwarunkowaƒ rozmna-
˝ania generatywnego i kie∏kowania nasion.
• Ekofizjologia glonów makroskopowych, mszaków i po-
w∏ok okrzemkowych jako podstawa okreÊlenia ich funk-
cji biocenotycznych i znaczenia wskaênikowego.
• Dynamika produkcji biomasy, pobierania i uwalniania
substancji biogennych przez roÊliny i osady denne w ró˝-
nych warunkach trofii i potencja∏u redox.
• Charakterystyka struktury i funkcjonowania populacji
i zbiorowisk na granicy zasi´gu geograficznego i na re-
liktowych stanowiskach wyspowych.
• OkreÊlenie trendów w zmianach struktury i rozmieszcze-
nia siedliska na tle zmian klimatycznych – na podstawie
wieloletnich danych monitoringowych.
• OkreÊlenie optymalnych biologicznie, ekologicznie i hy-
drologicznie technicznych metod zrównowa˝onego go-
spodarowania w korytach i dolinach rzek w klimatycz-
nych i geologicznych warunkach Polski.
Monitoring naukowy
Parametry do oceny: powierzchnia p∏atów poszczegól-
nych gatunków roÊlin i glonów makroskopowych, ich roz-
mieszczenie w segmencie cieku – mapka z zaznaczeniem wy-
sokiej roÊlinnoÊci brzegowej; morfologiczne wskaêniki kondy-
cji roÊlin, ocena intensywnoÊci zasilania podziemnego; cha-
rakterystyka chemiczna wód koryta i hyporeicznych (alkalicz-
noÊç, tlen, pH, potencja∏ redox, przewodnictwo elektr. , nie-
organiczne formy N i P, Fe rozp., Corg. lub ChZT); zag´sz-
czenie i struktura fauny bezkr´gowej.
Czas: 3 x w roku – maj/czerwiec, sierpieƒ, listopad dla scha-
rakteryzowania faz rozwoju (i ewentualnego zamierania) ro-
Êlin; dodatkowo oceniç znaczenie ewentualnych anomalii hy-
drologicznych (du˝e wezbrania, d∏ugotrwa∏e ni˝ówki).
Metody monitoringu: szkicowe kartowanie roÊlinnoÊci, zdj´cia
fotograficzne jako uzupe∏nienie szkiców, piezometry do pobierania
wód gruntowych, ∏apaczki przesiàków lub mini piezometry do okre-
Êlania pionowego gradientu hydraulicznego, standardowe metody
analiz chemicznych i pomiarów instrumentalnych, próby bezkr´-
gowców pobierane czerpakiem Surbera lub siatkà, oznaczenia
podstawowych grup funkcjonalnych bezkr´gowców.
Dla szybkiej oceny siedliska mo˝na oceniaç pokrycie po-
szczególnych gatunków roÊlin na co najmniej kilku reprezen-
tatywnych powierzchniach koryta rzeki o d∏ugoÊci 10 m, wy-
branych z ok. 500–metrowego odcinka rzeki.
Mo˝liwoÊci zdiagnozowania stanu ochrony: do póê-
niejszego opracowania na podstawie przeprowadzonych ba-
daƒ; stan poznania siedliska niewystarczajàcy. Wst´pnie mo˝-
na zaproponowaç uwzgl´dnienie nast´pujàcych wskaêników:
• % pokrycia powierzchni koryta przez roÊlinnoÊç,
• stosunek „pokrycie w sierpniu” / „pokrycie w listopadzie”
(o ile mi´dzy terminami badaƒ nie by∏o wi´kszych wezbraƒ
i nie prowadzono wycinania roÊlinnoÊci) , okreÊlajàcy de-
strukcj´ siedliska przez toksyczne wody podziemne.
• stosunek „powierzchnia p∏atów rdestnic, moczarki i je˝o-
g∏ówki” / „powierzchnia p∏atów roÊlin grupy I i II”, okreÊla-
jàcy degradacj´ siedliska spowodowanà nadmiernà kol-
matacjà dna, eutrofizacjà, niedoborem CO
2
, wysokà tem-
peraturà lub zbyt intensywnym wycinaniem roÊlin.
• stosunek „powierzchnia p∏atów rz´Êli” / ”powierzchnia
p∏atów w∏osieniczników”, jako czu∏y wskaênik zmian
re˝imu hydrologicznego i sedymentacyjnego (tylko na
stanowiskach odmiany typowej i zubo˝onej) .
Wojciech Puchalski
3260
1
Wyszukiwarka
Podobne podstrony:
3260 Nizinne i podgorskie rzeki ze zbiorowiskami wlosieniczn
3150 Starorzecza i naturalne eutroficzne zbiorniki wodne ze zbiorowiskami z Nympheion
OGÓLNE ZASADY KORZYSTANIA ZE ZBIORÓW W CZYTELNIACH
Innowacje Pedagogiczne – wybór materiałów ze zbiorów PBW
3130 Brzegi lub osuszone dna zbiornikow wodnych ze zbiorowis
3150 Starorzecza i naturalne eutroficzne zbiorniki wodne ze zbiorowiskami z Nympheion
Jędrzej Giertych Roman Dmowski pisarz polityczny , artykuł pochodzi ze zbiorowej pracy pt Pamięci R
Autyzm materiały ze zbiorów biblioteki w Chełmie (publikacje książkowe i artykuły) Kopia
filozofia, Relatywizm heraklitejski - nie można wejść do tej samej rzeki - chodzi o to że rzeka płyn
więcej podobnych podstron