2. USZKODZENIA DNA I ICH NAPRAWA
Mutacje i rekombinacje w DNA. Ewolucja na szczeblu molekularnym jest uwarunkowana akumulacja dziedzicznych zmian w genomach. Hugo de Vries wprowadzil na poczatku XX wieku termin "mutacja", na oznaczenie takich zmian badajac dziedziczenie cech u wiesiolka Lamarcka. Ironia losu chciala, ze efekty, ktore de Vries obserwowal byly spowodowane przegrupowaniami materialu genetycznego, ktorych to procesow w zasadzie nie obejmuje sie dzis terminem "mutacja" uzywajac na ich oznaczenie terminu "rekombinacja". Rozrozniamy kilka typow rekombinacji. Jeden z nich to rekombinacja zachodzaca podczas mejozy pomiedzy homologicznymi sekwencjami w DNA. Inny to reperacja uszkodzen DNA z wykorzystaniem sekwencji obecnych w chromosomie homologicznym lub na chromatydzie siostrzanej. Jeszcze inny to transpozycja polegajaca na wstawieniu jednego odcinka DNA w drugi. Wspolna ich cecha jest laczenie dwoch odcinkow DNA uprzednio ze soba niezwiazanych. Bedzie o nich mowa w innym rozdziale. Termin "mutacja" natomiast jest w zasadzie zarezerwowany dla trwalych zmian w sekwencji DNA nie wynikajacych z rekombinacji aczkolwiek pewna, historycznie uzasadniona niejednoznacznosc terminu "mutacja" przetrwala w literaturze genetycznej. Na przyklad osobniki u ktorych obserwuje sie zmiany spowodowane przez wstawienie transpozonu w gen nazywamy mutantami mimo, ze proces transpozycji zaliczany jest do rekombinacji. Podobnie, rearanzacje chromosomow takie jak rekombinacje pomiedzy chromosomami niehomologicznymi nazywa sie mutacjami chromosomowymi. Na pograniczu mutacji i rekombinacji sytuuja sie tzw. mutacje dynamiczne ktore polegaja na wydluzaniu sie lub skracaniu sekwencji mikrosatelitarnych. Tu bedziemy sie zajmowac uszkodzeniami DNA, ich naprawa oraz mutacjami polegajacymi na trwalej zmianie jednej pary zasad w chromosomie na inna jak rowniez krotkimi insercjami czy delecjami, ktore to mutacje bywaja wynikiem niedoskonalej naprawy tych uszkodzen.
Uszkodzenia DNA. Zasady w DNA sa reaktywne i stosunkowo latwo ulegaja uszkodzeniom spontanicznym, takim jak depurynacja czy dezaminacja, badz zachodzacym pod wplywem czynnikow mutagennych, takich jak promienie ultrafioletowe, czynniki alkilujace czy aktywne pochodne tlenu (tabela 2 - 1).
Glownym produktem reakcji DNA z kwantem energii UV jest dimer pirymidynowy posiadajacy czteroczlonowy pierscien cyklobutanu Py<>Py (ryc. 2 - 1).
Ryc. 2 - 1. Niektore uszkodzenia struktury DNA spowodowane promieniami UV. a) dimer tymidyny, b) fotoprodukt 6-4
Powstanie tego dimeru blokuje replikacje i transkrypcje i jesli nie zostanie naprawiony (ryc 2 - 2) powoduje smierc komorki. Innym czestym produktem reakcji fotochemicznej z udzialem zasad w DNA sa tzw fotoprodukty 6-4 (ryc. 2-1).
Ryc. 2 - 2. Reperacja dimerow pirymidynowych przez fotoliazy. 1) wzbudzenie systemu elektronowego czasteczki tetrahydrofolianu przez kwant swiatla widzialnego, 2) transfer energii z MTHF na FADH- 3) przekazanie elektronu na pierscien cyklobutanu, 4) odtworzenie FADH-
Szeroko rozpowszechnionymi mutagenami sa czynniki alkilujace, w szczegolnosci te, ktore metyluja zasady w DNA (ryc. 2 - 3). Podobnie jak w przypadku uszkodzen UV istnieja systemy odwracajace te uszkodzenia bez wycinania zasad z DNA (ryc. 2 - 4).
Ryc. 2 - 3. Przyklady zwiazkow alkilujacych
Ryc. 2 - 4. Schemat indukcji genu ada u E. coli.
Innym interesujacym uszkodzeniem w DNA sa addukty platyny. Zwiazki te, powstaja w wyniku traktowania komorek kompleksowymi pochodnymi platyny II lub IV wartosciowej. Pochodne te maja zastosowanie w klinice nowotworowej. W szczegolnosci zwiazek o nazwie « cisplatyna » (ryc. 2 - 5) tworzy zwiazki kowalencyjne z wieloma czasteczkami obecnymi w komorce. Jego biologiczna i terapeutyczna aktywnosc jest spowodowana glownie powstawaniem mostkow poprzecznych i podluznych w DNA (ryc 2 - 6, 2 - 7).
Najczestszy jest mostek pomiedzy atomami azotu N7 w pozycji 1,2 (sasiadujacych) puryn. Spotyka sie tez mostki 1,3 a takze mostki utworzone pomiedzy bardziej oddalonymi purynami. Analog typu trans- nie tworzy mostkow podluznych i jest nieaktywny jako lek aczkolwiek rowniez hamuje replikacje DNA. Cisplatyna jest znacznie bardziej toksyczna dla komorek, ktore maja nieczynne systemy reperacji DNA. W szczegolnosci komorki osob dotknietych fotodermatoza zwana "skora pergaminowata i barwnikowa" (xeroderma pigmentosum) pozbawione sa systemu naprawy uszkodzen przez wycinanie nukleotydow (NER, patrz ponizej) i sa bardzo wrazliwe na ten zwiazek. Okazalo sie, ze mostki podluzne 1,2 i 1,3 sa naprawiane wlasnie przez ten system. Komorki pochodzace od pacjentow z anemia Falconiego sa rowniez bardzo czule na cisplatyne.
Tlen, ktory z jednej strony jest niezbedny dla zycia jest rownoczesnie w duzej mierze toksyczny dla komorek, co zwiazane jest z tworzeniem jego silnie reaktywnych pochodnych a w szczegolnosci wolnych rodnikow. Powstaja one w komorkach glownie jako produkt uboczny metabolizmu (ryc. 2 - 8). Przyjmuje sie, ze w organizmie czlowieka powstaje rocznie okolo 2 kg anionorodnika O2Ż•
O2 + eŻ → O2Ż•
Jest to substancja wzglednie trudno reagujaca z DNA, ale w wyniku dysmutacji spontanicznej albo katalizowanej przez dysmutaze ponadtlenkowa daje H2O2
2 O2Ż• + 2 H+ → O2 + H2O2
W wyniku reakcji Fentona z nadtlenku wodoru H2O2 powstaje bardzo aktywny wolny rodnik hydroksylowy chetnie reagujacy z DNA
H2O2 + Fe++ → Fe+++ + [•OH] + OH¯
Uwaza sie, ze to nie sam ten rodnik dyfunduje i reaguje z DNA, ale, ze powstaje on in situ w tych miejscach, gdzie DNA jest zwiazane z jonami zelaza. Odgrywa tu role miedzy innymi NADH, ktory powoduje regeneracje Fe++ z Fe+++.
Ryc. 2 - 8. Losy tlenu w komorce. SOD dysmutaza ponadtlenkowa, GSH i GSSG zredukowana i utleniona forma glutationu.
Zarowno zasady jak i deoksyryboza ulegaja uszkodzeniom spowodowanym utlenieniem. Atak rodnika [•OH] na reszte rybozy powoduje oderwanie wodoru od wegla, przerwanie nici DNA i uwolnienie zasady. Atak rodnika na zasady powoduje 50 roznych uszkodzen. Najwiecej uwagi poswiecono formamido-pirymidynom oraz 7,8-dihydro-oksoguaninie (Tabela 2 - 1). Sposrod pirymidyn uwage skoncentrowano na glikolu tyminy oraz produktach jego hydrolizy. Zwiazki te sa usuwane z DNA przez specyficzne enzymy obecne w organizmach.
Najlepiej poznanym produktem utlenienia zasad w DNA jest 8-oksoG. Moze ona tworzyc pare zasad zarowno z C jak i z A, co moze byc przyczyna mutacji
Naprawa uszkodzen w DNA. W organizmie czlowieka zachodzi kilkadziesiat tysiecy roznych uszkodzen DNA w ciagu dnia. Uszkodzenia te w przewazajacej masie ulegaja naprawie przy pomocy licznych systemow naprawczych. Wiekszosc z tych systemow zostala wpierw poznana u E. coli po czym zwykle znajdowano podobne systemy w innych organizmach. Systemy naprawcze obecne w organizmach wyzszych sa bardziej skomplikowane niz u bakterii. Podlegaja one regulacji na poziomie organizmu, tkanki i komorki. U bakterii natomiast istnieje rodzaj centralnego ukladu regulacji systemow naprawy uszkodzen zwany systemem SOS. Najlepiej poznane systemy naprawy uszkodzen to :
-fotoreaktywacja, ktora jest systemem naprawy uszkodzen spowodowanych promieniami UV,
-metylotransferaza naprawiajaca alkilowany DNA
-system wycinania zasad z DNA (BER - Base Excission Repair),
-system wycinania nukleotydow z DNA (NER - Nucleotide Excission Repair),
-system reperacji zle sparowanych zasad (MMR - Mismatch repair),
-system omijania uszkodzen przez polimerazy o niskiej wiernosci kopiowania (TLS - translesion synthesis)
-niehomologiczne laczenie przerwanych nici DNA (NHEJ - non homologous end joining)
Fotoreaktywacja powoduje usuniecie uszkodzenia spowodowanego ekspozycja na krotkofalowe promieniowanie UV o dlugosci fali 200 - 300 nm wykorzystujac energie dostarczana przez promieniowanie o dlugosci fali 300 - 500 nm, a wiec przez dlugofalowy UV i przez swiatlo widzialne (ryc. 2 - 2). Ten mechanizm jest obecny w jednych gatunkach (E. coli, S. cerevisiae) a nieobecny w innych (H. influenzae, Homo).
Reakcja fotoreaktywacji polega na wiazaniu sie enzymu z produktem Py<>Py w etapie niezaleznym od swiatla, po ktorym nastepuje absorpcja fotonu bliskiego UV lub swiatla widzialnego, ktora z kolei powoduje rozszczepienie cyklobutanu, odtworzenie pirymidyn i dysocjacje enzymu. Fotoliazy zawieraja stechiometryczne ilosci chromoforow. Jednym z nich jest FADH (nukleotyd flawinowo adeninowy) a drugim albo metenylo-tetrahydrofolian (5,10 MTHF) albo 8-hydroksy-5-deazaryboflawina (8HDF). Z tego powodu dzielimy je na dwie klasy : folianowe i deazaflawinowe. Enzymy E. coli i S. cerevisiae naleza do klasy fotoliaz folianowych. 50 sposrod 150 aminokwasow konca karboksylowego fotoliaz jest identyczne dla wszystkich tych enzymow. Znajduje sie tam miejsce wiazania DNA oraz flawiny, ktora zapewnia specyficznosc wiazania do uszkodzonego DNA. FAD jest jedynym nukleotydem flawinowym znalezionym w fotoliazach. Nie moze on byc zastapiony przez zaden inny zwiazek np. ryboflawine. Folian znajduje sie w formie poliglutaminianu (3 - 8 reszt).
Reakcja rozszczepienia cyklobutanu zachodzi z rozna efektywnoscia w zaleznosci od sekwencji. Najefektywniej jest naprawiany dimer T<>T a najmniej efektywnie C<>C. Naprawa polega na absorpcji fotonu przez drugi chromofor (na rysunku MTHF) i transfer energii na flawine przez oddzialywanie dipol - dipol. *FADHŻ w stanie wzbudzonym przekazuje elektron dimerowi Py<>Py co powoduje dwuetapowe rozszczepienie pierscienia cyklobutanu. Powstajacy anion pirymidynowy jest nastepnie utleniany przez FADH• z odtworzeniem dwoch pirymidyn i funkcjonalnej formy flawiny.
Naprawa alkilowanego DNA przez metylotransferazy. U E. coli duza dawka zwiazku alkilujacego MNNG powoduje smierc komorek natomiast mala dawka indukuje opornosc na duza dawke. Stwierdzono, ze efekt ten wynika z indukcji systemow naprawczych a w szczegolnosci indukcji genu ada (ryc. 2 - 4) przez produkty naprawy uszkodzonego DNA. Bialko Ada jest syntetyzowane w malych ilosciach, ale konstytutywnie. Jest to metylotransferaza, ktora przenosi grupy metylowe z DNA na wlasne cysteiny obecne przy koncu karboksylowym oraz aminowym, dzialajac jako substrat w tej reakcji. W pierwszym etapie Cys 69 znajdujaca sie przy koncu aminowym bialka Ada jest metylowana przez fosfotrojestry obecne w alkilowanym DNA przy czym powstale alkilowane bialko indukuje transkrypcje szeregu genow, w tym glikozylazy usuwajacej uszkodzone zasady 3MetA, 3MetG i O2MetC. Indukuje rowniez swoja wlasna transkrypcje. W drugim etapie bialko Ada przenosi grupy metylowe z zasad na cysteine znajdujaca sie przy swoim koncu karboksylowym. Takie podwojnie metylowanie bialko nie ma juz wlasnosci aktywatora.
W komorkach ludzkich znajduje sie jedna O6MetG transferaza podobna do produktu genu ogt u E. coli. Jest ona syntetyzowana konstytutywnie i usuwa metyl z O6MetG i O6MetT a nie usuwa go z fosfotrojestrow. Wiele linii komorkowych nie syntetyzuje O6MetG transferazy co powoduje ich szczegolna podatnosc na dzialanie czynnikow alkilujacych. Niekiedy konwersja komorek z Mer+ do Mer- (nie syntetyzujacych O6MeG transferazy) jest wynikiem transformacji nowotworowej.
Naprawa uszkodzen w DNA z udzialem glikozylaz. W naprawie uszkodzen oksydatywnych biora udzial glikozylazy (tabela 2 - 2). Pierwsza opisana glikozylaza byla glikozylaza formamido-pirymidynowa z E. coli (bialko Fpg). Jak sie pozniej okazalo jest ona identyczna z produktem genu mutM, ktorego uszkodzenie powoduje akumulacje mutacji GC * TA. Funkcjonalne homologi tego bialka znaleziono u drozdzy i u czlowieka. (OGG1, 8-oksoguanino DNA glikozylaza 1). Wycina ono 8-OH-G z par 8-OH-G : C, 8-OH-G : T i 8-OH-G : G (ryc. 2 - 9).
Uszkodzone pirymidyny sa rowniez wycinane przez glikozylazy. Glikozylaza Nth wycina z DNA E. coli glikol tyminy, 5,6 dihydroksytymidyne, glikol uracylu itd. Strukturalne analogi tego bialka znaleziono u eukariotow, miedzy innymi u czlowieka. (NTG - endonuclease of tymine glycol). Po wycieciu uszkodzonej zasady przez glikozylaze istnieja dwie sciezki metaboliczne prowadzace do naprawy miejsca nieinstruktywnego (ryc. 2 - 10). Pierwsza z nich to przeciecie wiazania fosfodwuestrowego po stronie 5' od uszkodzenia, z utworzeniem 3' OH przez AP nukleaze (istnieje szereg nukleaz przecinajacych nic DNA w pozycji 5' w stosunku do miejsc AP, apurinic/apyrimidinic site). W nastepnym etapie 5' fosfodwuesteraza usuwa fosforan rybozy z ktorej glikozylaza usunela zasade, pozostawiajac 5'fosforan w nastepnej pozycji lancucha fosfodwuestrowego. Luka ta zostaje zapelniona przez polimeraze i ligaze III. Alternatywny szlak naprawy po przecieciu wiazania fosfodwuestrowego przez AP-endonukleaze to bezposrednia synteza kilku nukleotydow przez polimeraze δ. Wypchniety segment DNA jest wycinany przez specyficzna nukleaze FEN1. Aby ta reakcja zaszla niezbedna jest obecnosc klamry PCNA (proliferating cell nuclear antigen).
Jesli glikozylaza posiada aktywnosc liazy, tak jak bialko OGG1, mozliwa jest druga sciezka metaboliczna. Glikozylaza - liaza przecina wiazanie fosfodwuestrowe po stronie 3', co powoduje to, ze do wyciecia rybozy, ktora zostaje w tym procesie zlinearyzowana potrzebna jest aktywnosc 3' fosfodwuesterazy.
Ryc. 2 - 10. Schemat reakcji glikozylaz z uszkodzonym DNA.
Niedawno wyjaśniono w jaki sposób powstaje aktywny kompleks glikozylaz MutM i OGG1 z parą OG:C (oxoguanina - cytozyna). Trudność w tym przypadku polega na tym, że para ta nie deformuje w znaczny sposób helisy dwuniciowego DNA, podczas gdy w przypadku innych modyfikowanych zasad pary nukleotydów z reguły nie powstają w ogóle co silnie zmienia strukturę DNA i jest łatwe do wykrycia przez systemy reperacji.
Powstanie aktywnego kompleksu reperacyjnego powoduje wyślizgnięcie się reszty GO z podwójnej helicy DNA co jest wynikiem skoordynowanego działania trzech aminokwasów M77, R112 i F114. W początkowym stadium aminokwasy M77 i F114 dokonują inwazji DNA po stronie 3' GO, co zaburza oddziaływania typu base stacking, powoduje zgięcie się podwójnej spirali i obrót uszkodzonej zasady dookoła wiązania glikozydowego w pozycję syn . Następuje wówczas wejście argininy R112, w miejsce zajmowane dotychczas przez GO. Tworzy ona wiązania wodorowe z O2 i N3 cytozyny co w ostatecznym wyniku powoduje wysunięcie się GO poza podwójną spiralę i jej wejście do centrum aktywnego MutM (Ryc 2-11). Centrum to potrafi odróżnić OG od G i usuniąć zmodyfikowany nukleotyd.
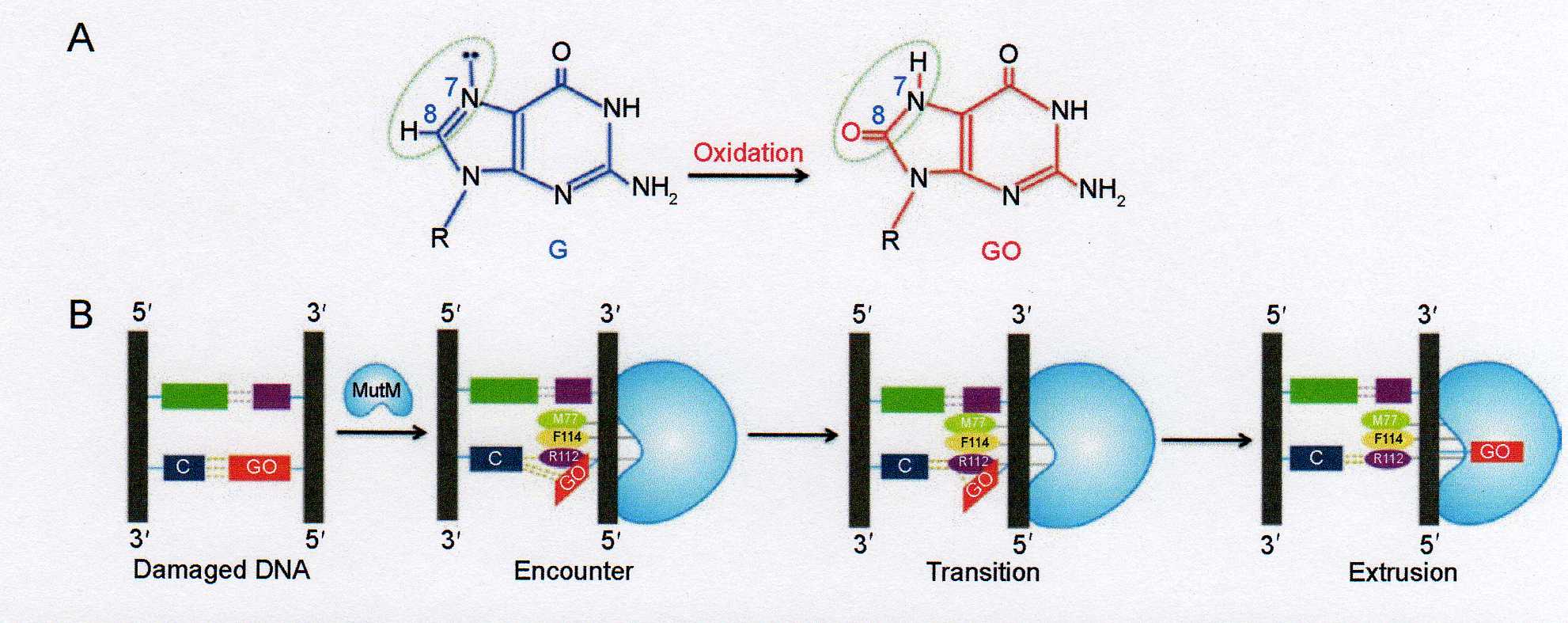
Ryc. 2-11 (a) powstawanie aktywnego kompleksu reperacyjnego MutM z parą OG:C
Ryc 2-11 (b) Konformacja anty i syn nukleotydu guanozyny
Naprawa uszkodzen przez wycinanie nukleotydow. Jest to proces bardziej skomplikowany niz wycinanie zasad przez glikozylazy. Wymaga on wspoldzialania szeregu bialek oraz zuzywa energie w postaci ATP, ale za to wykazuje mala specyficznosc wobec typu uszkodzenia. Wycinane moga byc fotodimery, addukty o roznej wielkosci podstawnika czy produkty utlenienia. U bakterii wycinany jest segment o dlugosci 12 - 13 nukleotydow, u eukariotow 27 - 29 nukleotydow.
Ryc. 2 - 12. Naprawa uszkodzen w DNA przez wyciecie nukleotydow (NER)
U E. coli naprawe zaczyna kompleks 2UvrA - UvrB (Ryc. 2 - 12) (UV resistant A i B) Powoduje on zgiecie DNA pod katem 130° zuzywajac ATP. (UvrA)2 nastepnie oddysocjowuje a do stabilnego kompleksu DNA-UvrB przylacza sie bialko UvrC zmieniajac konformacje kompleksu. Bialko UvrB, ktore wiaze ATP bez hydrolizy, nacina DNA w odleglosci 4 (5) nukleotydow po stronie 3' uszkodzenia. Nastepnie UvrC nacina DNA po stronie 5' w odleglosci 7 (8) nukleotydow od uszkodzenia. W nastepnym etapie helikaza II (UvrD) usuwa wyciety oligomer. UvrB pozostaje zwiazane z DNA do czasu wypelnienia luki przez polimeraze I i ligaze. U czlowieka NER jest inicjowane przez wiazanie sie kompleksu XPA - RPA (xeroderma pigmentozum, grupa komplementacji A, - replication protein A) do uszkodzenia. Po utworzeniu sie tego kompleksu dolacza sie kompleks TFIIH skladajacy sie z przynajmniej szesciu bialek (helikaza 5' - 3', XPD, helikaza 3' - 5', XPB, TTDA [trichotiodystrofia A], oraz bialka p34, p44 i p62). Kompleks ten zuzywa ATP i zgina DNA . XPC stabilizuje kompleks zgietego DNA z TFIIH a XPG dokonuje naciecia w odleglosci 3 - 5 nukleotydow po stronie 3' co z kolei powoduje zmiane konformacji kompleksu, przylaczenie sie ERCC1 (excission repair cross complementary gene 1) oraz XPF jak rowniez naciecie w odleglosci 20 - 24 nukleotydow po stronie 5'. Po wycieciu uszkodzonego odcinka kompleks oddysocjowuje pozostawiwszy w miejscu zawierajacym odcinek jednoniciowy DNA bialko RPA. Tworzy sie nowy kompleks RFC (replication factor C) ktory umieszcza klamre PCNA przy koncu 3' naciecia, po czym nastepuje przylaczenie sie polimerazy i synteza DNA . Ligaza konczy proces naprawy. U E. coli podobnie jak i u czlowieka transkrybowana nic DNA naprawiana jest szybciej niz nietranskrybowana, co wynika z tego, ze uszkodzenie stanowi przeszkode dla polimerazy RNA. Zatrzymana polimeraza wiaze sie z TRCF (transcription repair coupling factor) co powoduje oddysocjowanie polimerazy i transkryptu oraz przylaczenie sie kompleksu (UvrA)2 - UvrB. U czlowieka w procesie naprawy nici transkrybowanej uczestnicza bialka CSA i CSB (Cocayne syndrom, grupa komplementacji A lub B). Uczestnicza one w degradacji proteolitycznej polimerazy RNA zatrzymanej na DNA uszkodzonym przez promieniowanie UV. Jest to niezbedne dla podjecia transkrypcji po naprawie uszkodzenia.
Mostki podluzne 1,2 i 1,3 powstajace w DNA pod wplywem pochodnych platyny sa dobrym substratem dla systemu NER u czlowieka i w bakteriach. W ich usuwaniu biora udzial produkty genow XPA (odpowiedzialnego za rozpoznawanie uszkodzen) i XPE (o nieznanej funkcji). Mostki te sa znacznie bardziej aktywnie usuwane z nici transkrybowanej DNA co tlumaczy dlaczego niskie stezenie cisplatyny hamuje replikacje a nie hamuje transkrypcji. Opornosc na cisplatyne jest intensywnie badana poniewaz jest to jedna z przeszkod w szerszym stosowaniu tego leku w klinice. Jedna z przyczyn tej opornosci jest wzrost produkcji zwiazkow tiolowych w komorce, takich jak metalotioneina czy glutation. Zmniejszenie wrazliwosci na cisplatyne moze rowniez wynikac ze zwiekszonej syntezy polimerazy . Polimeraza ta ma jedyna w swoim rodzaju wlasciwosc mutagennego omijania mostkow 1,2 (GpG). Inne bialko o zwiekszonej ekspresji w komorkach opornych na cisplatyne to PCNA. Natomiast bialka posiadajace domene typu HMG (high mobility group) zwiekszaja czulosc komorki na cisplatyne (ryc. 2 - 7). Szereg bialek tego typu ma zdolnosc wiazania sie z czasteczka DNA w miejscu zgiecia spowodowanego utworzeniem mostka platynowego co oslania ten mostek przed mechanizmem naprawczym NER.
System SOS. Ekspozycja komorki bakterii E. coli na dzialanie czynnikow powodujacych uszkodzenia DNA indukuje transkrypcje co najmniej 42 genow rozsianych po genomie (ryc. 2 - 13). Produkty tych genow albo biora udzial w naprawie uszkodzen, albo tez zwiekszaja tolerancje komorki na te uszkodzenia (np. RecA, lexA, umuDC, polB, i uvrA). Podstawowymi elementami systemu SOS sa produkty genow recA i lexA. Produktem genu recA jest bialko, ktore bierze udzial nie tylko w regulacji systemu SOS, ale rowniez w innych procesach metabolicznych takich jak rekombinacja homologiczna czy odblokowanie widelek replikacyjnych zatrzymanych przez uszkodzenie DNA. Czasteczki bialka RecA wiaza sie z jednoniciowym DNA (ssDNA, single stranded DNA) rownoczesnie ulegajac polimeryzacji. Oligomer tworzacy sie w tej reakcji ma postac helikoidalnie skreconej nici. Jest on centralnym elementem strukturalnym posredniczacym w roznych reakcjach metabolizmu DNA. LexA natomiast jest bialkiem represorowym, ktory wiaze sie z sekwencja zwana kaseta SOS (SOS box) znajdujaca sie w sasiedztwie promotorow genow systemu SOS. Kompleks LexA z kaseta SOS uniemozliwia transkrypcje tych genow. Gdy DNA zostanie uszkodzony komorka podejmuje probe naprawy uszkodzenia stosujac rozne systemy naprawcze takie jak BER czy rekombinacja homologiczna. Gdy proba taka okaze sie nieudana, jednoniciowy odcinek DNA powstaly w jej wyniku wiaze bialko RecA. Kompleks RecA - ssDNA dziala jako koproteaza, ktora stymuluje autotrawienie represora LexA. To zmniejsza jego ilosc w komorce, powoduje jego dysocjacje z kaset SOS i indukcje regulonu SOS.
Bialko RecA jest rowniez jednym z bialek systemu SOS a wiec jest indukowane w wyniku autotrawienia LexA. System pozostaje aktywny tak dlugo, jak dlugo istnieje jednoniciowy DNA. Gdy jego ilosc w komorce sie zmniejsza, stezenie kompleksu RecA - ssDNA rowniez sie zmniejsza, co powoduje wzrost stezenia nienaruszonych czasteczek LexA w komorce, co z kolei pociaga za soba represje genow posiadajacych kasete SOS w regionie promotora.
Ryc. 2 - 13. System SOS u E. coli
Podstawowa funkcja mechanizmu SOS w komorce jest zapewnienie jej funkcjonowania po szoku jakim jest uszkodzenie DNA blokujace synteze replikatywna. SOS odblokowuje replikacje przez uruchomienie procesu syntezy o zmniejszonej wiernosci kopiowania (TLS - translesion synthesis). Cena jaka placi komorka za zwiekszenie szansy przezycia jest wzrost czestosci mutacji. Odbywa sie to w sposob nastepujacy: w wyniku autotrawienia LexA stymulowanego przez kompleks RecA - ssDNA zachodzi synteza RNA w operonie umuDC. Bialko UmuD, ktore jest jednym ze skladnikow kompleksu replikacyjnego podlega trawieniu w obecnosci RecA - ssDNA podobnie jak bialko LexA. W wyniku trawienia nastepuje odciecie 24 aminokwasow z konca aminowego UmuD. Dwie czasteczki tego nowego bialka zwanego UmuD' tworza razem z UmuC kompleks zwany polimeraza V, ktory to kompleks ma zdolnosc « przeslizgiwania sie » przez uszkodzone miejsce na DNA, w szczegolnosci przez miejsca nieinstruktywne, a wiec np. takie, w ktorym w matrycy brak jest zasady (najczesciej puryn). Pol V jest aktywna w obecnosci kompleksu RecA - ssDNA. Naprzeciw miejsc nieinstruktywnych Pol V wstawia w nowosyntetyzowana nic DNA przewaznie adenine. Niezaleznie od tego, synteza nieuszkodzonego DNA przez Pol V rowniez charakteryzuje sie zwiekszona iloscia mutacji. Obok Pol V zidentyfikowano caly szereg innych polimeraz, ktore syntetyzuja DNA na uszkodzonych matrycach przeslizgujac sie przez uszkodzenia generujac mutacje. Jedna z nich jest bialko DinB zwane inaczej polimeraza IV, ktora generuje substytucje oraz jednonukleotydowe delecje (-1 frameshift) w miejscach, gdzie w matrycy DNA znajduje sie zasada « wylamana » z szeregu. Geny kodujace polimerazy homologiczne do Pol IV i Pol V znaleziono w wielu organizmach miedzy innymi u czlowieka. Mozna mowic o superrodzinie mutagenizujacych polimeraz UmuC/DinB.
System SOS jest jednym z mechanizmow odpowiedzialnych za « mutacje adaptacyjne » tzn. za mutacje, ktore pojawiaja sie w warunkach gdy bakterie (lub ogolnie komorki) sa poddane selekcji, co nie znaczy jednak, ze mutacje te zachodza wylacznie w kierunku korzystnym dla komorek. W przypadku np. wyczerpania czynnikow odzywczych w srodowisku, komorki aktywuja system SOS, ktory przyspiesza powstanie mutantow zdolnych do zycia w zmienionych warunkach.
Mutacje mutatorowe. Podobna role spelniaja mutanty « mutatory » spontanicznie pojawiajace sie w populacjach. Zwykle zdolnosc do produkowania mutacji wynika z uszkodzenia ktoregos z systemow reperacji uszkodzen DNA. Wykazuja one wielokrotnie wyzsza czestosc mutacji niz szczep dziki. Mutatory w zasadzie znajduja sie w niekorzystnej sytuacji poniewaz ich zdolnosc przezycia zmniejsza sie w miare gromadzenia mutacji, z reguly niekorzystnych. Niemniej, ku zaskoczeniu badaczy okazalo sie, ze w populacjach bakteryjnych zyjacych w srodowiskach naturalnych spotyka sie zawsze pewna ilosc mutatorow. Uwaza sie obecnie, ze mutatory spelniaja role odkrywcow. Wsrod mutacji masowo przez nich produkowanych znajduja sie, choc rzadko i takie, ktore sa korzystne z punktu widzenia przezywalnosci. Gdy allel zawierajacy korzystna mutacje zostanie przekazany bakterii szczepu dzikiego (np. przez koniugacje) moze dac poczatek nowemu szczepowi bakterii lepiej przystosowanemu do zycia w zmienionym srodowisku niz szczep dotychczas tam zyjacy.
Tabela 2 - 1 Uszkodzenia w DNA
Tabela 2 - 2. Glikozylazy
Uszkodzenia DNA
-----------------------------------------------------------------------------------------------------------------
28
21
Ryc. 2 - 5. Pochodne platyny uzywane jako leki antynowotworowe.
cisplatyna
karboplatyna
NH3
NH3
NH3
NH3
5-MeC
Ryc. 2 - 6. Produkty reakcji zwiazkow platyny z DNA (a) mostek podluzny, (b) mostek poprzeczny, (c) addukt jednowartosciowy, (d) mostek DNA - bialko.
Ryc. 2 - 7 Oddzialywanie bialek posiadajacych domene HMG z DNA zawierajacym podluzne mostki platyny.
Ryc. 2 - 9. Mutacje powodowane przez 8 - Oksoguanine.
ROS (reactive oxygen species - reaktywne pochodne tlenu).
MTH ( MutT homologue - bialko ludzkie homologiczne do produktu genu MutT u E. coli hydrolizujace 8oksoGTP)
MYH (glikozylaza wycinajaca adenine sparowana z 8-oksoG. U E. coli produkt genu MutY).
OGG1, 2 (8-oksoGuanino DNA glikozylaza 1 lub 2. U E. coli role te spelnia produkt genu Mut M)
Mut M
Wyszukiwarka
Podobne podstrony:
[W] Badania Operacyjne (2009 02 21) wykład
2009 02 17 test egzaminacyjny n Nieznany (2)
2009-02-26, pedagogium, wykłady, Komunikacja społeczna
2009 02 16 test egzaminacyjny nr 3 Pomorski ZPN odpowiedzi A
SIMR-AN1-EGZ-2009-02-02a-rozw
2009 02 09
2009 02 Relacyjna baza danych HSQLDB [Bazy Danych]
2009 02 09 test egzaminacyjny nr 3 Pomorski ZPN odpowiedzi A
EDW 2009 02
efektywnosc plus 2009 02
pytania 2009-02-07, Procesy spalania
2009 02 Podstawowa kinezyterapia u kobiet po mastektomii
PRINTER Optionen 2009 02 27
SIMR-AN1-EGZ-2009-02-09a-rozw
2009 02 testy odpowiedzi
2009 02 Podstawy MySQL [Poczatkujacy]
25 Interakcje lekow w farmakoterapii bolu PTAiIT Lodz 2009 02 17 Woron J
2009 02 Metasploit Alternate Uses for a Penetration Test [Atak]
więcej podobnych podstron