BŁONY BIOLOGICZNE
Transport bierny = dyfuzja
cechy transportu:
zgodny z gradientem stężeń
bez udziału energii pochodzącej z hydrolizy ATP
rodzaje dyfuzji:
prosta - zachodzi bezpośrednio przez błonę, cząsteczki nie potrzebują dodatkowych struktur, żeby się przez nią przedostać (dotyczy niewielkich obojętnych cząsteczek, np. gazów oraz substancji rozpuszczalnych w tłuszczach - etanol, hormony steroidowe)
ułatwiona - wymaga obecności:
Dyfuzja ułatwiona przez kanały jonowe - substancje transportowane (np. woda, jony) nie łączą się z białkiem tworzącym kanał
kanały mogą być:
specyficzne - odróżniają anion od kationu
selektywne - odróżniają rodzaj kationu
budowa kanałów:
4,5 lub 6 podjednostek białkowych, których oś symetrii wyznacza drogę dla substancji
im więcej podjednostek tym szersza ta droga
kanały bramkowane elektrycznie (na przykładzie kanału sodowego) - bramkowanie zachodzi dzięki różnicy potencjałów między zewnętrzną w wewnętrzną stroną błony komórkowej:
depolaryzacja powoduje otwarcie kanału jonowego
przez otwarty kanał napływają jony (np. Na+)
po krótkim czasie kanał zamyka się powodując repolaryzację błony
po 2-5 ms kanał nadal jest zamknięty, ale podatny na pobudzenie
jeżeli nastąpi kolejna depolaryzacja to się otworzy
kanały bramkowane chemicznie (ligandem)
posiadają one specyficzne miejsca do których może przyłączyć się agonista lub antagonista
przyłączenie się agonisty powoduje otwarcie kanału (acetylocholina zwiększa przewodnictwo Na+/K+)
przyłączenie się antagonisty powoduje zamknięcie się kanału
depolaryzacja powoduje otwarcie kanału jonowego
przez otwarty kanał napływają jony (np. Na+)
po krótkim czasie kanał zamyka się powodując repolaryzację błony
po 2-5 ms kanał nadal jest zamknięty, ale podatny na pobudzenie
jeżeli nastąpi kolejna depolaryzacja to się otworzy
Naturalnym agonistą tego receptora jest acetylocholina. Do agonistów egzogennych należą nikotyna i muskaryna, go antagonistów min. atropina, dekstrometorfan, pirenzepina
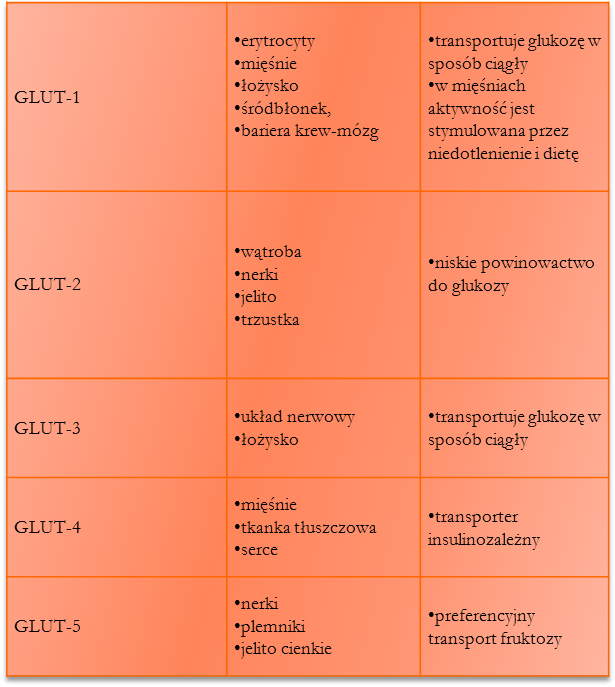
Dyfuzja ułatwiona przez białka transportowe - podczas transportu tworzy się przejściowe wiązanie kowalencyjne między białkiem a substratem (np. glukozą)
rodzaje transporterów GLUT:
Transport aktywny
cechy:
zachodzi wbrew gradientowi stężeń
niezgodny z równowagą termodynamiczną (ΔG >0)
z udziałem nośnika
wykorzystuje energię pochodzącą z: hydrolizy ATP, ruchu elektronów lub światła
rodzaje:
transport aktywny pierwotnie
transport aktywny wtórnie
translokacja grup
Transport aktywny pierwotnie - transport w którym cała zużywana energia jest wykorzystywana na wytworzenie wiązań kowalencyjnych w nośniku
pompa sodowo-potasowa:
transport na zasadzie antyportu
składa się z dwóch podjednostek β (nieaktywnych) oraz dwóch podjednostek α do których przyłącza się ATP
ATP-aza Na+/K+ jest integralną częścią pompy - enzymem odpowiedzialnym za hydrolizę ATP w celu uzyskania energii
mechanizm działania:
funkcje:
pompa wapniowa
utrzymuje niski poziom wapnia cytozolowego (czyli wypompowuje wapń na zewnątrz komórki lub do cytozolu w przypadku pompy w retikulum)
Rola wapnia w komórce: cząsteczka sygnałowa (dlatego nie może być go za dużo)
występuje w retikulum endoplazmatycznym i błonach plazmatycznych
pompa protonowa
transport na zasadzie antyportu
znajduje się w komórkach okładzinowych żołądka
transport jonów H+ do światła żołądka i wprowadzenie K+ do komórki
transport na zasadzie antyportu
składa się z dwóch podjednostek β (nieaktywnych) oraz dwóch podjednostek α do których przyłącza się ATP
ATP-aza Na+/K+ jest integralną częścią pompy - enzymem odpowiedzialnym za hydrolizę ATP w celu uzyskania energii
mechanizm działania:
funkcje:
- kanału jonowego, przez który mogą przechodzić substancje
- transporteru (np. GLUT)
Receptor acetylocholinowy składa się z 5 podjednostek, każda z nich z domeny zewnątrzkomórkowej, segmentu transbłonowego i segmentu wewnątrz komórki.
Białka transportowe (BT) umożliwiają przenoszenie przez błonę substancji, które nie rozpuszczają się w lipidach. Wszystkie białka transportowe są białkami integralnymi, wielokrotnie przebijającymi błonę.
- ATP fosforyluje i tym samym uaktywnia ATP-azę
- do podjednostki α pompy wiązane są trzy jony Na+
- hydroliza ATP do ADP
- zmiana konformacji białka
- jony sodu zostają przetransportowane na zewnątrz komórki i uwolnione z kompleksu
- przyłączenie 2 jonów potasu do podjednostki α
- defosforylacja wywołuje ponowną zmianę konformacji białka
- jony potasu przedostają się do wnętrza komórki
- uwolnienie jonów K+ po przyłączeniu się ATP
- kontroluje objętość komórki
- niezbędny do pobudzenia nerwów i mięśni (chwilowe zatrzymanie pompy Na/K powoduje depolaryzację błony komórkowej i przekaz sygnału na dalsze komórki)
- siła napędowa do wtórnego transportu aktywnego
Pompy jonowe klasy P
(zależne od ATP):
• (Na+, K+)-ATPaza, w błonach plazmatycznych większości komórek
zwierzęcych, antyportowa wymiana Na+/K+
• (H+, K+)-ATPaza, bierze udział w wytwarzaniu kwaśnego soku żołądkowego,
mechanizm antyportowy. Katalizuje zależny od ATP transport H+z komórek
okładzinowych żołądka (do światła żołądka), wprowadzając K+ do komórki.
• Ca2+-ATPaza, w retikulum endoplazmatycznym (ER) i błonach plazmatycznych
wielu komórek. Katalizuje zależny od ATP transport Ca2+ z cytozolu, do
światła ER lub na zewnątrz komórki. Mogą być antyporterami, przenoszącymi
H+ w przeciwnym kierunku. Ca2+-ATPazy utrzymują niski poziom cytozolowego
wapnia, umożliwiając działanie Ca2+ jako cząsteczki sygnałowej.
Podczas fosforylacji oksydacyjnej, w wyniku szeregu reakcji redoks, elektrony przenoszone są ze zredukowanych nukleotydów, NADH i FADH2, na pełniący funkcję akceptora elektronów tlen. Zachodzące reakcje prowadzą do zmagazynowania energii, służącej następnie do syntezy ATP. Szereg reakcji redoks zachodzi na kompleksach białkowych znajdujących się w mitochondriach. Zestaw enzymów biorących udział w przenoszeniu elektronów określa się jako łańcuch oddechowy.
Energia uwalniana podczas transportu elektronów w łańcuchu oddechowym zużywana jest do przenoszenia protonów przez wewnętrzną błonę mitochondrialną. Energia potencjalna gromadzona jest w postaci gradientu pH i potencjału elektrycznego w poprzek błony. Zgromadzona w tej formie energia wykorzystywana jest przez kompleks enzymatyczny syntazy ATP, który pozwala protonom przejść przez błonę zgodnie z gradientem stężeń. Enzym ten zamienia jednocześnie energię gradientu pH i elektrycznego na energię wiązań chemicznych ATP, wytwarzanego przez przyłączenie do ADP reszty kwasu ortofosforowego, czyli reakcji fosforylacji. Niezwykłość reakcji syntezy ATP związana jest z obracaniem się części enzymu napędzanej przepływającymi protonami, przypominając działanie silnika elektrycznego. Obrót części enzymu odłącza wytworzoną cząsteczkę ATP.
pompa sodowo-potasowa:
- ATP fosforyluje i tym samym uaktywnia ATP-azę
- do podjednostki α pompy wiązane są trzy jony Na+
- hydroliza ATP do ADP
- zmiana konformacji białka
- jony sodu zostają przetransportowane na zewnątrz komórki i uwolnione z kompleksu
- przyłączenie 2 jonów potasu do podjednostki α
- defosforylacja wywołuje ponowną zmianę konformacji białka
- jony potasu przedostają się do wnętrza komórki
- uwolnienie jonów K+ po przyłączeniu się ATP
- kontroluje objętość komórki
- niezbędny do pobudzenia nerwów i mięśni (chwilowe zatrzymanie pompy Na/K powoduje depolaryzację błony komórkowej i przekaz sygnału na dalsze komórki)
- siła napędowa do wtórnego transportu aktywnego
Miejsce wiązania ATP znajduje się na podjednostce α. Na tej podjednostce, na powierzchni skierowanej do środowiska zewnątrzkomórkowego, znajdują się również miejsca wiązania dla steroidów kardiotonicznych (np. digitoksygeniny, uabainy, marinobufageniny), które hamują aktywność pompy przez blokowanie defosforylacji. Powoduje to wzrost stężenia jonów wapnia w mięśniu sercowym i skurcz mięśnia (bicie serca). Zwiększenie siły skurczu.
Wyszukiwarka
Podobne podstrony:
Wersja 8 zeszły rok2, rok numer trzy, biochemia, seminaria
Kolokwium 2 (sem) - pytania, rok numer trzy, chemia leków, seminaria
podział sem 3, rok numer trzy, chemia leków, seminaria
Morfologia, rok numer trzy, farmakognozja, sprawdziany = kolokwia
spr 2(1), rok numer trzy, farmakognozja, sprawdziany = kolokwia
spr 1(1), rok numer trzy, farmakognozja, sprawdziany = kolokwia
klo farmakologia II(1), rok numer trzy, farmakognozja, sprawdziany = kolokwia
gnozja egzamin gr 2, rok numer trzy, farmakognozja, egzamin
Pytania - spr 3, rok numer trzy, farmakognozja, sprawdziany = kolokwia
Test z ćwiczeń 7-10, rok numer trzy, tpl, kolokwia
pytania egzamin 2009, rok numer trzy, farmakognozja, egzamin
spr 4, rok numer trzy, farmakognozja, sprawdziany = kolokwia
Biolomolo Ćwiczenia, rok numer trzy, biolomolo
Gnozja - wykłady by me, rok numer trzy, farmakognozja, wykłady
TPL - wykłady z receptury jałowej, rok numer trzy, tpl, wykłady
Sprawdzian 1, rok numer trzy, farmakognozja, sprawdziany = kolokwia
spr 1, rok numer trzy, farmakognozja, sprawdziany = kolokwia
więcej podobnych podstron