Różnicowanie słoja przyrostu rocznego drewna sosny pospolitej ( Pinus sylwestris )
Jan Górski Katedra botaniki leśnej
Gr. II SGGW w Warszawie
I. Wstęp.
Badaniami objęto wyłącznie cewki systemu podłużnego sosny pospolitej (Pinus sylvestris). Komórki wytworzone przez podziały komórek kambialnych przechodzą poszczególne fazy: wzrostu (strefa w której komórki zwiększają swoje wymiary, głównie wymiar promieniowy), maturacji (strefa w której komórki odkładają wtórną ścianę komórkowej, jest jeszcze żywa), aby stać się w pełni wykształconą cewką podłużną o grubej, ścianie komórkowej jednak całkowicie pozbawiona żywego protoplastu.
Zaobserwowano, że w okresie wiosennym wytwarzane są cewki cienkościenne o dużej średnicy promieniowej, natomiast późnym latem tworzone są cewki grubościenne o małej średnicy promieniowej.
II. Cele
Celem tej pracy jest wyjaśnienie, co decyduje o takim zróżnicowaniu słoja rocznego. Czy decydujący wpływ na wielkość komórek i grubość wtórnych ścian komórkowych czas trwania fazy G i D, czy też intensywności procesów zachodzących w tych wazach.
III. Materiał
Materiał do analizy pobrany został ze stuletniego drzewostanu sosnowego z powierzchni doświadczalnej w Rogowie. Próbki pobierano za pomocą wbijaka (φ 5 mm), następnie sporządzono z nich preparaty mikroskopowe, utrwalono w alkoholu, zabarwiono zielenią świetlistą (nie zdrewniałe ściany barwią się na zielono czyli zielone aa komórki żywe) i safraniną (ściany zdrewniałe barwią się na czerwono czyli czerwone są martwe komórki) i zatopiono w balsamie kanadyjskim. Obserwacje prowadzono pod mikroskopem licząc komórki w tworzącym się słoju na podstawie wcześniej pobranych próbek.
W niniejszej pracy wzięto pod uwagę wyniki z czterech terminów badawczych. Są to:
termin 4-ty - 31 maj
termin 8-my - 30 lipiec
termin 12-ty - 19 sierpień
termin 16-ty - 28 wrzesień
IV. Wyniki
TAB.1 Liczba komórek w poszczególnych strefach rozwojowych drewna sosny, pomierzona w próbkach pobranych w 4 terminach
|
Data i numer obserwacji |
|
|
|
|
|
|
|
|
|
|
|
||||
nr |
31 maj (4) |
|
|
10 lipiec (8) |
|
19 sierpień (12) |
|
28 wrzesień (16) |
|
|||||||
drzewa |
Liczba komórek w strefie |
|
|
|
|
|
|
|
|
|
|
|
||||
|
C |
G |
D |
T |
C |
G |
D |
T |
C |
G |
D |
T |
C |
G |
D |
T |
15 |
9 |
5 |
5 |
0 |
7 |
4 |
9 |
11 |
6 |
5 |
9 |
18 |
6 |
2 |
12 |
22 |
20 |
8 |
5 |
2 |
0 |
8 |
5 |
5 |
6 |
5 |
4 |
6 |
12 |
4 |
3 |
13 |
20 |
57 |
8 |
6 |
2 |
0 |
7 |
4 |
4 |
8 |
6 |
4 |
6 |
14 |
4 |
3 |
8 |
16 |
159 |
7 |
8 |
3 |
0 |
6 |
5 |
4 |
6 |
5 |
4 |
7 |
16 |
5 |
3 |
8 |
29 |
123 |
9 |
8 |
1 |
0 |
7 |
3 |
4 |
4 |
7 |
3 |
12 |
13 |
5 |
0 |
12 |
27 |
21 |
8 |
6 |
1 |
0 |
6 |
5 |
7 |
8 |
5 |
4 |
8 |
15 |
4 |
3 |
11 |
24 |
39 |
5 |
9 |
4 |
0 |
5 |
4 |
8 |
10 |
4 |
4 |
11 |
17 |
4 |
3 |
8 |
25 |
37 |
7 |
10 |
1 |
0 |
7 |
6 |
10 |
12 |
7 |
4 |
14 |
17 |
7 |
3 |
7 |
15 |
92 |
8 |
7 |
4 |
0 |
7 |
4 |
5 |
12 |
2 |
4 |
15 |
21 |
5 |
2 |
9 |
21 |
58 |
8 |
5 |
2 |
0 |
5 |
3 |
5 |
8 |
5 |
5 |
8 |
13 |
5 |
3 |
10 |
23 |
35 |
7 |
8 |
3 |
0 |
6 |
4 |
4 |
12 |
5 |
5 |
14 |
21 |
4 |
3 |
14 |
19 |
112 |
7 |
8 |
3 |
0 |
7 |
6 |
7 |
11 |
6 |
6 |
9 |
18 |
3 |
4 |
16 |
16 |
167 |
8 |
7 |
2 |
0 |
6 |
6 |
8 |
8 |
6 |
4 |
7 |
9 |
5 |
3 |
17 |
21 |
68 |
7 |
14 |
8 |
0 |
6 |
6 |
10 |
20 |
6 |
6 |
19 |
27 |
6 |
1 |
11 |
30 |
średnia |
8 |
7 |
3 |
0 |
6 |
4 |
6 |
10 |
4 |
4 |
10 |
17 |
5 |
3 |
11 |
22 |
T |
|
|
|
0 |
|
|
|
10 |
|
|
|
17 |
|
|
|
22 |
T+D |
|
|
3 |
|
|
16 |
|
|
27 |
|
|
33 |
||||
T+D+G |
|
10 |
|
20 |
|
31 |
|
36 |
||||||||
T+D+G+C |
18 |
26 |
35 |
41 |
||||||||||||
|
|
|
|
10 |
|
|
11 |
|
|
5 |
|
|
|
|||
Czas pomiędzy poszczególnymi terminami badań wynosi 40 dni ( t= 40 dni).
Z przedstawionej tabeli 1 wynika, że średnio w ciągu badanego sezonu przyrosło 36 komórek drewna. Jeżeli założymy, że każda dojrzała komórka osiąga 40 *m w wymiarze promieniowym, to średnio przyrasta rocznie ok. 1,5mm drewna. Wieloletnie obserwacje udowadniają, że średni przyrost słoja rocznego uzależniony jest od częstości podziałów kambium. Analizując dane zawarte w tab.1 stwierdziliśmy, że ilość komórek kambium maleje wraz z upływem czasu.
Celem mojej pracy jest jednak odpowiedzenie na pytanie od czego zależy wielkość przyrostu:
1. Na pewno od długości okresu aktywności kambialnej. Podziały rozpoczynają się średnio w połowie kwietnia ( w koronie nieco wcześniej), a kończą na przełomie września i października. Razem trwa to pięć i pół do sześciu miesięcy. Długości okresu aktywności podziałowej kambium zależy również od cech genetycznych danego drzewa jak i o siedliska na jakim wegetuje. Zauważono również wyraźną zależność między wzrostem promieniowym a stanem wierzchołka - każdy czynnik hamujący wzrost powoduje ustanie wzrostu promieniowego różnicujących się cewek.
2. Zależy też od liczby komórek kambialnych na początku okresu wegetacyjnego - im jest ich więcej, tym większy jest przyrost.
3. Zależy także od tempa, czyli ich częstotliwości, podziałów komórek kambialnych. Tępo to było zagadnieniem' którym najpierw się zajęliśmy i uzyskaliśmy następujące wyniki:
Tempo tworzenia się słoja - V= ![]()
[komórki/dzień]
gdzie ![]()
- przyrost słoja miedzy 2 okresami pomiarowymi
t - czas pomiędzy kolejnymi pomiarami
V1=![]()
komórki/dzień
V2=![]()
komórki/dzień
V3=![]()
komórki/dzień
Dane do tych obliczeń pochodzą z tab. 1. Jak widać wyniki obrazują iż początkowo słój przyrastał szybciej a pod koniec okresu wegetacyjnego wyraźnie wolniej. Podobnie jak do obliczeń powyższych tak do tych poniżej dane pochodzą z tab. 1. Przedstawiają one intensywność podziałów komórkowych, mówiących nam co ile dni zachodzi podział. Jest ona odwrotnie proporcjonalna do tempa podziału. Przedstawia to wzór:
[dni/podział]
gdzie I to częstość podziałów komórkowych, c to szerokość strefy kambialnej, v to tempo tworzenia się słoja czyli v =. I tak:
I1=![]()
dni/podział czyli 1 podział co 23 dni
I2=![]()
dni/podział czyli 1 podział co 22 dni
I3=![]()
dni/podział czyli 1 podział co 44 dni.
Komórki dzielą się więc bardzo rzadko - około raz na miesiąc. Zakładając, że okres aktywności kambialnej trwa 6 miesięcy (t=6), a szerokość strefy kambialnej wynosi sześć komórek (c=6), powinniśmy ze wzoru c*t otrzymać: 6*6=36 komórek i taką właśnie liczbę określiłem z tabeli 1 na początku mojej pracy.
Następnie zastanawiałem się od czego może zależeć wymiar promieniowy i grubość ścian komórkowych cewek. Teoretycznie zależy ona może od :
czasu przebywania w strefie G i D, czyli od czasu trwania wzrostu i odkładania ściany komórkowej.
intensywności wzrostu komórek i odkładania ściany komórkowej.
W badanych preparatach wyróżniono kilka stref:
strefa kambium (C),
strefa różnicowania (G),
strefa maturacji (D),
strefa cewek dojrzałych (T).
Szerokość tych strefy uzależniona jest od tempa „wchodzenia” do niej komórek (A) i wychodzenia komórek ze stref do następnej (B).
Natomiast czas przebywania komórki w strefie zależy od:
szerokości strefy,
tempa „wchodzenia” komórek do stref.
Pomiędzy tempem wchodzenia do strefy (A), a tempem wychodzenia (B) z niej zachodzą następujące zależności:
A= B szerokość strefy nie zmienia się,
A< B strefa zwęża się,
A> B strefa rozszerza się
Na podstawie tych zależności jak i znajomości faktu, że tempo „wchodzenia” do strefy następnej jest równe tempu wychodzenia ze strefy poprzedniej, obliczyliśmy średni czas przebywania komórek w kolejnych strefach. Pierwszym krokiem było policzenie tempa wchodzenia do poszczególnych stref:
1. Tempo wchodzenia do strefy T(AT). At=![]()
=BD [komórki/ dobę]
AT1= 0,250 [komórki/ dobę] = BD1
AT2= 0,175 [komórki/ dobę] = BD2
AT3= 0,125 [komórki/ dobę] = BD3
2. Tempo wchodzenia do strefy D(AD).
AD=![]()
=BG [komórki/ dobę]
AD1= 0,325 [komórki/ dobę] = BG1
AD2= 0,275 [komórki/ dobę] = BG2
AD3= 0,150 [komórki/ dobę] = BG3
3. Tempo wchodzenia do strefy G(AG).
AG=![]()
= BC [komórki/ dobę]
AG1= 0,250 [komórki/ dobę] = BC1
AG2= 0,275 [komórki/ dobę] = BC2
AG3= 0,125 [komórki/ dobę] = BC3
Następnie policzyliśmy już same czasy przebywania komórek w poszczególnych strefach
a. Czas trwania strefy D (tD)
tD=D/BD ; D - szerokość strefy maturacji;
BD - tempo wchodzenia komórek ze strefy maturacji.
tD1= 18 dni
tD2= 46 dni
tD3= 84 dni
Czas przebywania komórek w strefie maturacji może dochodzić do dwóch miesięcy .
b. Czas trwania strefy G (tG)
tG= G/BG G - szerokość strefy wzrostu;
BG - tempo wychodzenia komórek ze strefy wzrostu
tG1= 17 dni
tG2= 15 dni
tG3= 23 dni
Na podstawie wyników powyższych obliczeń widać, że czas przebywania cewek drewna wczesnego w strefie wzrostu jest niewiele różniący się od czasu przebywania cewek drewna późnego. Natomiast czas przebywania w strefie maturacji jest zdecydowanie dłuższy dla cewek drewna późnego.
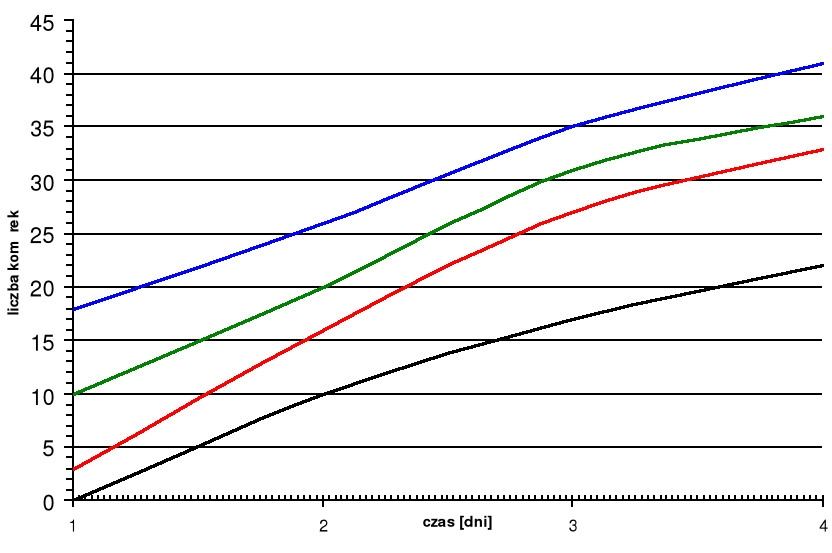
Rys.1. Szerokość poszczególnych stref w słoju w zależności od czasu od rozpoczęcia badań
V. Dyskusja:
Powyższy tok obliczeń jasno obrazuje, iż intensywność przyrostu słoja rocznego w trakcie sezonu wegetacyjnego zależy od liczby komórek strefy kambialnej i wyniósł on średnio około 36 komórek w ciągu sezonu. Na podstawie wyników tabeli 1 zauważono, że największy przyrost wykazały drzewa badane w pierwszym okresie badawczym 31 maj - 30 lipiec. Tak więc wiosną drzewa wytwarzają największą liczb* komórek kambialnych co powoduje zwiększenie przyrostu ksylemu. Wnioskujemy z tego, że potencjalne możliwości komórko-twórcze kambium z czasem maleją. Z ilością komórek kambialnych oraz częstości ich podziałów wiąże się także tempo przyrostu słoja czyli ilość produkowanych komórek w ciągu doby. Wielkość ta jest zmienna w zależności od pór roku. Wiosną jest ono największe, a wraz z upływem czasu zmniejsza si*. Swoje minimum osiąga jesienią.
Różnice w budowie cewek drewna wczesnego i drewna późnego zależą w głównej mierze od czasu przebywania w strefie maturacji (w której odbywa si* proces odkładania ściany komórkowej) oraz od intensywności wzrostu komórek. Stwierdzono, że cewki drewna późnego mają grubsze ściany komórkowe ponieważ ich czas przebywania w strefie maturacji jest bardzo długi (nawet do dwóch miesięcy). Na grubość ścian komórkowych niezbyt znacznie wpływa intensywność odkładania ścian komórek. Na wymiar promieniowy cewek drewna wczesnego wpływa przede wszystkim intensywność wzrostu. Wywnioskowano to z czasu przebywania komórek w strefie wzrostu. Okazało si*, że czas ten jest średnio jednakowy dla wszystkich cewek, zatem sądzić można, że wielkość cewek drewna wczesnego (w kierunku promieniowym) zależy od intensywności wzrostu.
Literatura:
Hejnowicz Z.,1973; „Anatomia rozwojowa drzew”: Państwowe Wydawnictwo Naukowe
Szweykowska A.,Szweykowski J., 1992: „Botanika”; Wydawnictwo Naukowe PWN
2
C
G
D
T
Wyszukiwarka
Podobne podstrony:
7535
7535
(7535) ck cw4 kofeina, nauka, chemia
7535
praca-magisterska-wa-c-7535, Dokumenty(2)
7535
7535
7535
7535
więcej podobnych podstron