Fitohormony
Wstęp
Regulacja procesów fizjologicznych może zachodzić poprzez:
regulację metaboliczną - regulację podstawowych przemian metabolicznych
regulację troficzną - poprzez zapewnienie lub ograniczenie dostępu substancji odżywczych do komórek
regulację z wykorzystaniem regulatorów wzrostu i rozwoju (zwanych także po prostu regulatorami wzrostu).
Regulatory wzrostu: przeważnie małocząsteczkowe, zróżnicowana budowa chemiczna, zdolne do przemieszczania się w roślinie. Należą do metabolitów wtórnych (nie uczestniczą w metabolizmie podstawowym. Regulują wzrost, podziały, różnicowanie się komórek, uczestniczą w odpowiedzi na czynniki środowiska np. stresy abiotyczne (światło, T, dostępność wody...) czy biotyczne (patogeny...)
Jak testuje się, czy dana substancja jest regulatorem wzrostu? Kryteria:
występowanie zależności pomiędzy stężeniem tej substancji a natężeniem danego procesu.
podanie substancji egzogennie inicjuje lub modyfikuje przebieg procesu
wywołanie zmiany stężenia substancji w roślinie modyfikuje przebieg procesu.
Jakie procesy można badać: wzrost, morfogenezę, zmiany zawartości konkretnych związków chemicznych, zmiany aktywności enzymu/ów,
Najprostsze do przeprowadzenia są eksperymenty z egzogennym podawaniem, jednak wtedy nie ma pewności, czy obserwowany efekt jest rzeczywistym efektem fizjologicznym (czy odpowiada reakcjom rośliny na zmiany stężenia endogennej substancji). Kryterium 1 - zależność natężenia procesu od stężenia wymaga dokładnej analizy ilościowej (endogenne regulatory wzrostu często występują w bardzo niskich stężeniach) oraz często (ze względu właśnie na małą zawartość) dużej ilości materiału roślinnego. Ostatnio wprowadza się metody immunochemiczne. Z kolei badanie zmian endogennego stężenia (kryterium 3) można wywołać traktując roślinę inhibitorem biosyntezy, transportu, czy katabolizmu tego regulatora. Ostatnio często przeprowadza się badania na mutantach lub roślinach transgenicznych - mogą one mieć np. upośledzoną biosyntezę, degradację lub transport hormonu albo niezdolny do przekazania sygnału receptor tegoż hormonu czy nieaktywny inny element kaskady sygnałowej.
Hormony roślinne - podgrupa regulatorów wzrostu.
są aktywne w bardzo małych stężeniach (już rzędu 10-6 mola/dm3)
występują powszechnie w roślinach.
Nie będące hormonami - substancje wzrostowe - działają w wyższych stężeniach (10-4 mol/dm3. Ich występowania i działanie często bywa ograniczone do danej grupy taksonomicznej, czasem nawet tylko do kilku gatunków (np. niektóre fenole czy terpenoidy pełnią funkcje substancji wzrostowych). Niektóre substancje wzrostowe powstają w odpowiedzi na zadziałanie czynnika stresowego np. ataku patogenu. Do tej grupy należą też fitoaleksyny, czy wytwarzane przez bakterie Rhizobium flawonoidy wywołujące specyficzną odpowiedź roślin motylkowych. Najczęściej są to związki małocząsteczkowe o prostej budowie (np. kwas salicylowy), choć są także bardziej skomplikowane: oligosacharydy, glikozydy fenolowe, polipeptydy.
Hormony roślinne mogą działać w miejscu ich syntezy, jak również w miejscach od nich odległych.
Obecnie stworzono szereg syntetycznych analogów hormonów, które mogą być trwalsze, wywoływać intensywniejszą odpowiedź, lub działać antagonistycznie (niektóre herbicydy, które są właśnie analogami hormonów). Znane są także związki będące np. inhibitorami biosyntezy naturalnych hormonów.
Koniugaty hormonów
Grupy: karboksylowa, aminowa i hydroksylowa hormonów mogą być wykorzystywane do tworzenia połączeń z cukrami, aminokwasami lub krótkimi peptydami - takie połączenia nazywamy koniugatami hormonów. Poza nielicznymi wyjątkami (dla cytokinin) koniugaty są nieaktywne biologicznie, ale przyłączenie podstawnika nie jest nieodwracalne. Odłączenie podstawnika w wyniku hydrolizy całkowicie przywraca aktywność. Tworzenie koniugatów jest jednym ze sposobów zmniejszania stężenia aktywnego hormonu, ale ma tę przewagę nad procesami degradacji, że inaktywacja hormonu w ten sposób jest odwracalna. Komórka może gromadzić pulę nieaktywnego związku i uwalniać aktywny hormon z koniugatu w razie zajścia takiej potrzeby. Powstawanie koniugatów zaobserwowano też po dodaniu egzogennego hormonu. Koniugaty mogą też pełnić rolę formy transportowej hormonów.
Hormony roślinne - fitohormony
Sześć klas - w każdej klasie związki o podobnej budowie i niemal identycznej aktywności.
Auksyny, gibereliny, cytokininy, kwas abscysynowy (ABA), kwas jasmonowy (JA), etylen,
Działanie plejotropowe - jeden hormon reguluje wiele procesów. Często też jeden hormon wywołuje odpowiedzi w różnych tkankach, czy organach, bywa, iż odpowiedź na ów hormon może być tkankowo/organospecyficzna. W większości przypadków jest też tak, że dany proces jest regulowany przez więcej niż jeden hormon. Hormony różnych klas działające na dany proces mogą działać synergistycznie lub antagonistycznie. Może być tak, że dla danego procesu jeden hormon odgrywa rolę dominującą, a pozostałe - są współdziałające.
Główny podział ze względu na funkcje:
Stymulatory wzrostu:
auksyny, gibereliny, cytokininy
Inhibitory wzrostu:
ABA, etylen, JA
Podział ze względu na biosyntezę:
Pochodne aminokwasów
auksyny, etylen
Pochodne zasad azotowych:
cytokininy
Pochodne izoprenu:
gibereliny, ABA, cytokininy (patrz biosynteza)
Pochodne kwasów tłuszczowych:
JA
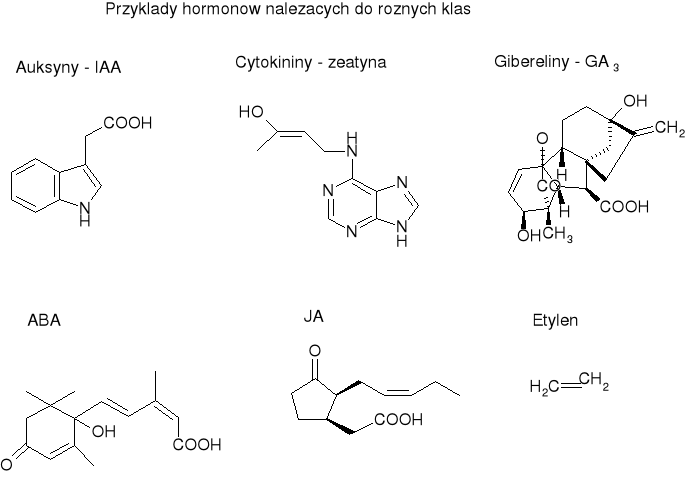
Auksyny
Pochodne tryptofanu.
Przykłady: IAA (kwas indolilo-3-octowy, pierwszy odkryty hormon roślinny, najpowszechniej występujący), nitryl kwasu indolilo-3-octowego, kwas 4-chloro-3-indolilooctowy. Większość auksyn to pochodne indolu, ale są też auksyny nieindolowe np. kwas fenylooctowy.
Syntetyczny analog: kwas 2,4-dichlorofenoksyoctowy (2,4-D), aktywny a stężeniach 1000x mniejszych niż IAA.
Co regulują, bądź za co odpowiadają:
wzrost elongacyjny (stymulują)
ukorzenianie (stymulują)
stymulacja tworzenia korzeni przybyszowych i korzeni bocznych
ruchy roślin - kontrola ruchów wzrostowych (często w różnych tropizmach np. fototropizm, grawitotropizm)
zjawisko dominacji wierzchołkowej
generalnie uważane za stymulatory wzrostu, ale ich aktywność stymulująca tylko dla pewnych zakresów stężeń, zakresy te są specyficzne dla danych organów (i tak np. w niskich stężeniach IAA stymuluje wzrost korzenia, ale już w nieco wyższych - hamuje). Korzenie są niezwykle wrażliwe na działanie auksyn, stężenia zewnętrznie stosowanych auksyn, optymalne dla wydłużania się koleoptyli, czy łodyg wielu roślin (ok. 10-5 moli/dm3)wywołują prawie całkowite zahamowanie wydłużania się korzeni, hamująco nie wpływają tylko bardzo małe stężenia , natomiast stężenia auksyny w zakresie 10-9 - 10-11 mola/dm3 stymulują wzrost korzeni niektórych roślin
do silnego wydłużania pędów niezbędna zawartość auksyn, ale też giberelin
regulacja różnicowania komórek
syntetyzowane w wierzchołkowej części liścia stymulują wzrost i rozwój tkanek przewodzących
starzenie się (opóźniają- np. opóźniają starzenie liści roślin drzewiastych)
nadmiar auksyn w pędzie stymuluje syntezę etylenu
hamują tworzenie warstwy odcinającej poprzedzające opadanie liści i owoców
są w stanie indukować tkankę merystematyczną
u niektórych roślin małe stężenia egzogennej auksyny stymulują kwitnienie a większe hamują, odpowiedź na auksynę silnie zależy też od etapu rozwoju kwiatu, przypuszcza się, że hamują samą indukcję kwitnienia ale z kolei stymulują morfogenezę kwiatu, po przejściu rośliny w fazę generatywną
małe stężenia auksyn przyspieszają a duże opóźniają starzenie się kwiatów
rozwój kwiatów
razem z giberelinami i cytokininami regulują aktywność podziałową kambium i felogenu podczas przyrostu na grubość
dojrzewanie owoców - stymulują rozrost zalążni razem z giberelinami, ale opóźniają - razem z giberelinami i cytokininami dojrzewanie owoców
Miejsca syntezy (generalnie - rosnące tkanki)
merystemy wierzchołkowe pędu i korzenia
młode liście, pąki, kwiaty, owoce,
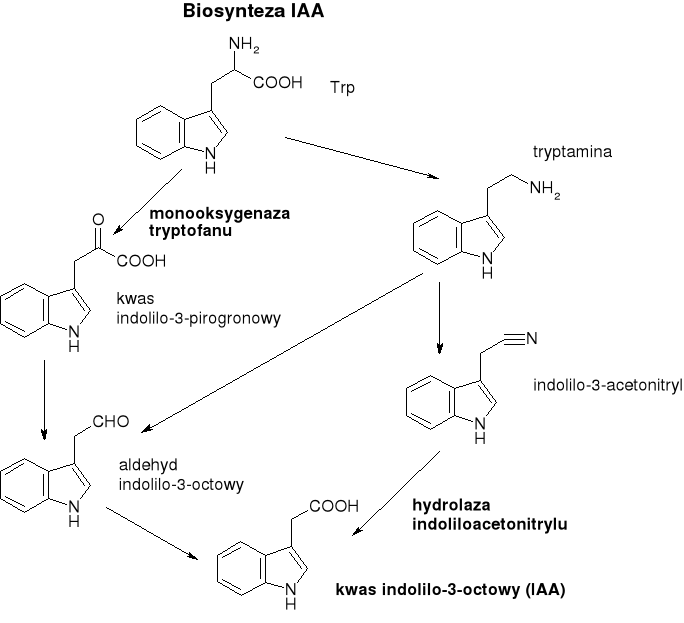
Biosynteza IAA:
W niektórych procesach fizjologicznych (np. kielowanie nasion, wzrost etiolowanych siewek) układ syntetyzujący IAA wykorzystuje Trp uwalniany w wyniku hydrolizy białek, natomiast zazwyczaj wykorzystywany jest Trp z puli wolnych aminokwasów. Trp - jest jednym z produktów szlaku kwasu szikimowego.
Deaminacja, dekarboksylacja i utlenienie tryptofanu - mogą przebiegać w różnej kolejności - powstają wtedy różne metabolity pośrednie. Udział poszczególnych szlaków może być różny w zależności od gatunku, organu czy stadium rozwoju. Najważniejsza z dróg: kolejno deaminacja, dekarboksylacja, utlenianie.
Przedostatni produkt pośredni - aldehyd indolilo-3-octowy może być gromadzony w niektórych roślinach w stosunkowo znacznych stężeniach (rzędu 10-5 mol/dm3)- być może u tych roślin ostatni krok biosyntezy reguluje produkcję hormonu, jednak u roślin testowych - i aldehyd i nitryl kwasu indolilo-3-octowego są aktywnymi stymulatorami wzrostu.
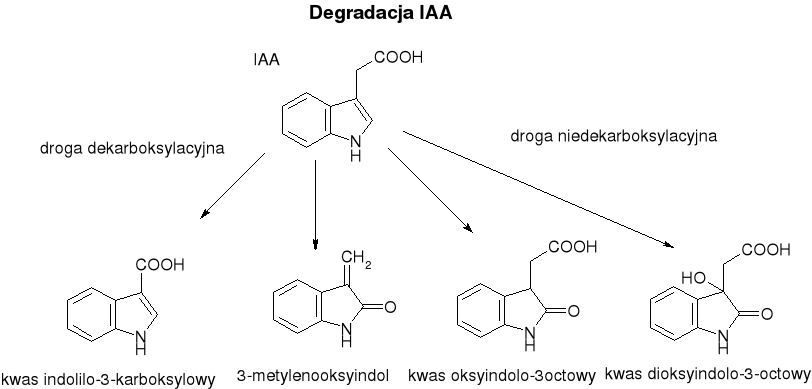
Degradacja: polega na modyfikacjach łańcucha bocznego lub układu indolowego prowadzących do utraty aktywności biologicznej. Najpowszechniej występuje oksydacyjna dekarboksylacja katalizowana przez kompleks enzymatyczny nazywany oksydazą IAA, w skład którego wchodzi peroksydaza zawierająca Mn (aktywność oksydazy IAA wymaga jonów Mg2+ oraz jest modulowana przez niektóre związki fenolowe). Są też inne drogi - niedekarboksylacyjne. IAA może też ulegać fotooksydacji z rozszczepieniem pierścienia indolowego od 3-metyleno-2-oksyindolu i indoliloaldehydu.
Koniugaty:
W ich powstawaniu bierze udział grupa karboksylowa IAA.
Powstają estry glikozylowe i amidy. Pochodne estrowe częściej w jednoliściennych (zboża), amidy - w dwuliściennych. Uwalnianie auksyn z koniugatów przez hydrolizę.
Powstawanie koniugatów poznano dokładnie dla kukurydzy:
UDP-glukoza + IAA -> glukozyd IAA
Potem - transestryfikacja do estru mezo-inozytolowego, który ulega kolejnym glikozylacjom.
Etylen
Jego prekursorem jest metionina.
Co reguluje, bądź za co odpowiada:
wzrost elongacyjny (hamują, sprzyjają natomiast wzrostowi na grubość) (czynniki hamujące wydłużanie pędu - etylen, ABA, światło, kinetyna - stymulują jednocześnie jego grubienie)
dojrzewanie owoców - przyspiesza procesy dojrzewania, zwłaszcza owoce klimakteryczne wykazują zdolność do indukowanego przez sam etylen wytwarzania tego hormonu
starzenie się (przyspieszają) (starzejące się liście wydzielają zwiększone ilości etylenu)
regulacja odpowiedzi na stres
udział w stymulowaniu opadania liści (razem z ABA)
regulacja kiełkowania
regulacja etiolacji
przejście w stan spoczynku
stymulacja wzrostu korzenia na grubość
udział w tworzeniu aerenchymy
hamuje kwitnienie wielu roślin dnia krótkiego
przyspiesza więdnięcie kwiatów (wrażliwość na etylen zależy także od stadium rozwojowego kwiatu i zazwyczaj zwiększa się w miarę dojrzewania i starzenia się kwiatów)
Miejsce syntezy:
prawdopodobnie we wszystkich żywych komórkach
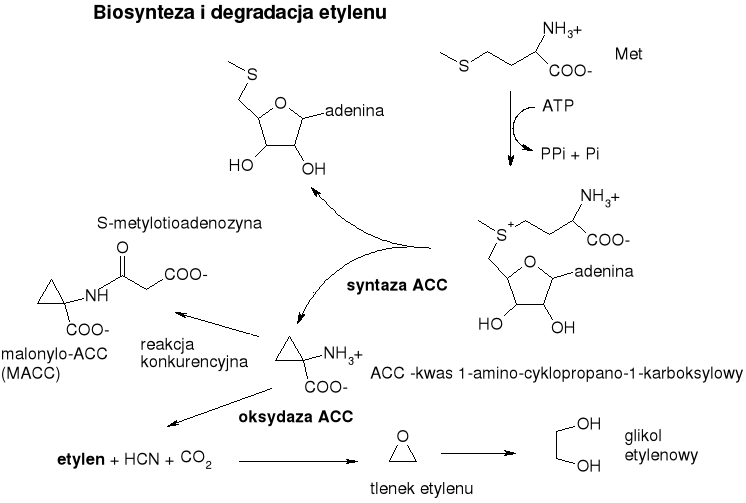
Biosynteza:
Metionina nie pochodzi z białek komórkowej tylko z puli wolnej metioniny.
Kluczowy produkt pośredni to S-adenozylometionina (SAM)
Powstawaniu etylenu towarzyszy powstawanie równomolowej ilości cyjanowodoru.
Bezpośredni prekursor etylenu - ACC (kwas 1-amino-cyklopropano-1-karboksylowy)
Aktywność kluczowego enzymu szlaku biosyntezy etylenu - syntazy ACC jest pod kontrolą innych hormonów: IAA indukuje syntezę tego enzymu - stymulacja produkcji etylenu, ABA - przeciwdziała. Zatem - mamy tutaj swego rodzaju ujemne sprzężenie zwrotne. Sygnał stymulujący (auksyna) indukuje wzrost syntezy sygnału hamującego. Inny sygnał hamujący hamuje syntezę etylenu.
Droga biosyntezy etylenu zawiera wspólny odcinek ze szlakiem biosyntezy grupy substancji wzrostowych - mianowicie z SAM mogą powstawać aminy biogenne (są to poliaminy - związki alifatyczne zawierające kilka grup aminowych) np. putrescyna, spermina, spermidyna. Poliaminy wykazują wielokierunkową aktywność biologiczną, występują i są aktywne w stężeniach znacznie większych niż hormony.
Oksydaza ACC związana z tonoplastem.
Ujemne sprzężenie zwrotne - etylen hamuje oksydazę ACC a stymuluje malonylotransferazę ACC.
Degradacja: utlenienie do tlenku etylenu, a potem glikolu etylenowego.
Nie tworzy koniugatów. Ale może je tworzyć jego prekursor - ACC - amid z kwasem malonowym - (malonylo-ACC, MACC). Powstawaniu MACC sprzyja zahamowanie syntezy etylenu. Hydrolizę MACC do ACCC stwierdzano sporadycznie, stąd nie uważa się MACC za „rezerwuar” etylenu.
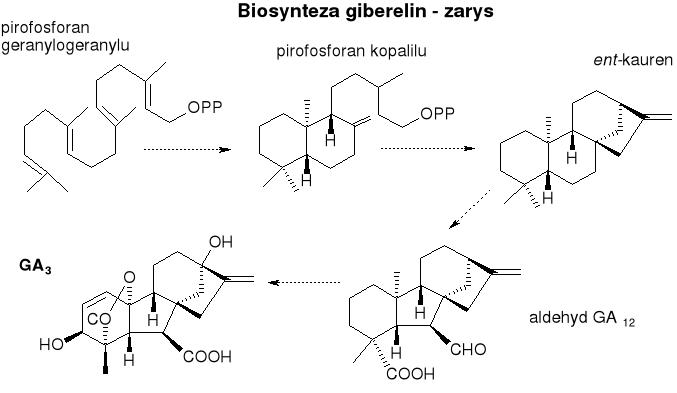
Gibereliny
Pochodne izoprenu.
Liczna rodzina - do tej pory poznano ponad 80. Pierwszą giberelinę (kwas giberelowy) obecnie nazywaną GA3 odkryto... u grzyba Gibberella fujikuroi, który wywoływał powstawanie narośli na zakażonych roślinach - dlatego właśnie, że syntetyzował substancję będącą hormonem roślinnym, która stymulowała wzrost tkanek.
Pochodne diterpenu ent-giberelanu. Zawierają trójpierścieniowy układ gibanu. Różnią się między sobą: stopniem utlenienia, obecnością lub brakiem pierścienia laktonowego, podstawnikami itp. Mają podobną aktywność biologiczną.
Za główne gibereliny roślin wyższych uważa się: GA1, GA4, GA9.
Co reguluje, bądź za co odpowiada:
spoczynek - jest jednym fitohormonów współdziałających przy ustępowaniu spoczynku
kiełkowanie nasion (podczas kiełkowania odpowiadają za aktywację materiałów zapasowych)
podane egzogennie przełamują karłowatość siewek
regulacja kwitnienia. Są w stanie wywołać kwitnienie w warunkach nieindukcyjnych u wielu, choć nie wszystkich roślin dnia długiego oraz u niektórych roślin wymagających wernalizacji (wernalizacja, jaryzacja - indukcyjne działanie obniżonej temperatury na kwitnienie roślin), choć są też gatunki roślin, u których hamują. Zaobserwowano odpowiedzi specyficzne na daną konkretną giberelinę, ważny okazał się także czas podawania egzogennych hormonów. U roślin długiego dnia po zastosowaniu indukcyjnego fotoperiodu wzrastają stężenia endogennych giberelin, wzrasta też wrażliwość wierzchołków wzrostu na ich działanie
wzrost
zawiązywanie owoców
stymulują podziały komórkowe w subapikalnej strefie merystemu, oraz wydłużanie się komórek, aktywne są jednak tylko w obecności auksyn, cytokininy mogą hamować to działanie giberelin
razem z cytokininami przeciwdziałają starzeniu, ponadto stężenie giberelin i cytokinin spada w starzejących się organach
podanie egzogenne stymuluje wzrost
gibereliny i cytokininy opóźniają dojrzewanie i starzenie się owoców
prawdopodobnie wytwarzane w wierzchołku wzrostu korzenia gibereliny hamują tworzenie korzeni bocznych w jego pobliżu, gibereliny mogą też hamować ukorzenianie np. siewek fasoli
spoczynek bulw ziemniaka ustępuje w wyniku działania gibereliną
Miejsca syntezy:
młode liście
wierzchołki wzrostu
korzenie
dojrzewające nasiona i owoce
pręciki kwiatów
zarodek w nasieniu
Transport - przez łyko i drewno
Biosynteza:
Pierwsze etapy - wspólne dla wszystkich terpenoidów. Pirofosforan izopentenylu i jego izomer pirofosforan dimetyloallilu powstają na szlaku kwasu mewalonowego z 3 cz. acetylo-CoA.
Takie jednostki izoprenowe mogą kondensować ze sobą dając dłuższe łańcuchy, one następnie są przekształcane w szereg ważnych związków roślinnych.
Z 4 reszt izoprenowych - pirofosforan geranylogeranylu - dalej - kolejne cyklizacje - powstaje ent-kauren. Potem - szereg przekształceń w tym utlenienia.
Bezpośredni prekursor wszystkich giberelin - aldehyd GA12.
Dalsze reakcje - utleniania, oksydacyjnej dekarboksylacji, hydroksylacji, odwodorowania (z utworzeniem wiązania podwójnego), dehydratacji z utworzeniem dodatkowego pierścienia laktonowego...
Degradacja:
Gibereliny najbardziej utlenione są bardzo słabo aktywne biologicznie, zaś końcowe produkty dalszego ich utleniania są już całkowicie nieaktywne i uważa się je za produkty metabolicznej dezaktywacji.
Koniugaty:
Przede wszystkim pochodne cukrowe, choć prawdopodobnie mogą też powstawać amidy (aminokwasy lub krótkie peptydy związane z grupą karboksylową przy węglu C7) oraz estry kwasu octowego i propionowego.
Pochodne cukrowe - do tej pory odkryto tylko pochodne glukozy, związanej z grupą hydroksylową przy 2,3 albo 12 węglu, albo - estry glikozylowe gdzie glukoza estryfikuje grupę karboksylową C7.
Donorem reszty cukrowej w syntezie koniugatów jest UDP-glukoza.
Uwalnianie giberelin z koniugatów głównie przez β-glukozydazy.
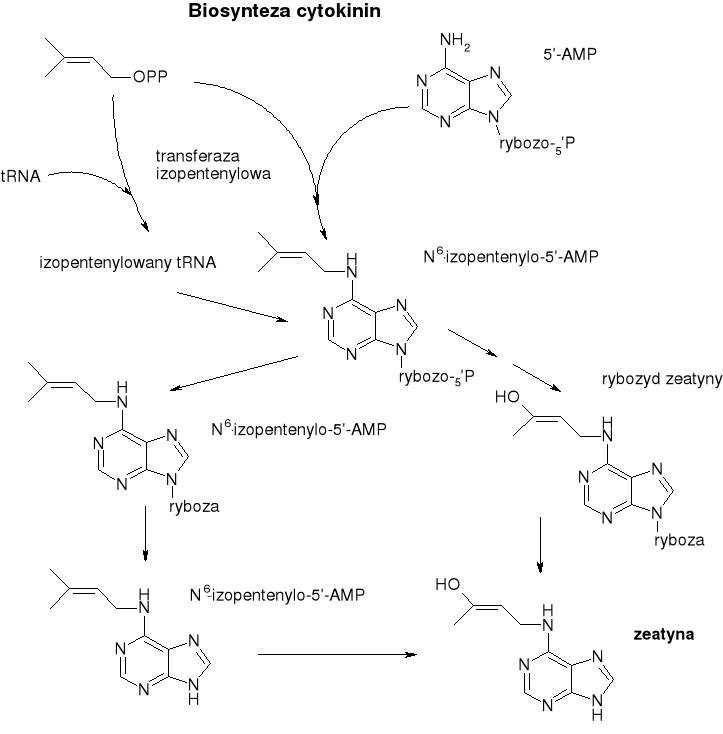
Cytokininy
Powstają w wyniku kondensacji adeniny z aktywowaną jednostką izoprenową.
Przykład: zeatyna ( (γ-hydroksymetylo, γ-metyloalliloamino)puryna, pierwsza poznana) kinetyna.
Ta klasa liczy niewiele związków, wszystkie są izopentenylowymi pochodnymi adeniny,
W odróżnieniu od pochodnych cukrowych innych hormonów glikozydy (rybozydy - bo najczęściej przyłączana ryboza) cytokinin i ich fosforany (rybotydy) są aktywne biologicznie.
Co reguluje, bądź za co odpowiada:
podziały komórkowe (stymulują)
różnicowanie komórek (reg.)
wzrost (stymulująco)
starzenie (przeciwdziałają)
mobilizacja substancji pokarmowych, regulacja transportu asymilatów
spoczynek (przeciwdziałają wchodzeniu w stan spoczynku)
wraz z giberelinami, auksynami, etylenem i ABA kontrolują rozwój tkanek blaszki liściowej poza przewodzącymi (tam głównie auksyna działa)
egzogennie podane cytokininy niwelują hamujący wpływ auksyn na rozwój pędów bocznych
mają różny wpływ na kwitnienie u różnych roślin, częściej jednak działają stymulująco, ich efekt zależy od obecności innych substancji wzrostowych
przedłużają trwałość ciętych kwiatów
indukcja powstawania merystemów z tkanek parenchymatycznych
inicjacja powstawania kalusa
proliferacja pączków
Miejsce syntezy:
głównie w korzeniach
owoce
młode liście
Transport - przez łyko i drewno
Biosynteza:
Cząsteczka pirofosforanu izopent-2-enylu kondensuje z resztą adeniny znajdującą się w cząsteczce 5'-AMP lub w cząsteczce tRNA w wyniku działania odpowiedniej transferazy (syntetaza cytokininowa).
Z samej biosyntezy wynika powszechność występowania cytokinin w formie rybozydów i rybotydów (obok formy niezwiązanej powstałej w wyniku hydrolizy produktu kondensacji.
Nukleotydy o budowie cytokinin wchodzą w skład niektórych rodzajów tRNA (najczęściej tRNA przenoszący reszty Ser i Tyr i sąsiadują z antykodonem). Postulowano bezpośredni udział tych hormonów w regulacji syntezy białka.
Wiele syntetycznych analogów cytokinin (jak 6-benzyloadenina) wykazuje aktywność biologiczną podobną do naturalnych hormonów. Zastosowanie praktyczne.
Degradacja: (zarówno naturalnych, jak i syntetycznych analogów) - polega na oksydacyjnym oderwaniu reszty izopentenylowej (enzym katalizujący - oksydaza cytokininowa) a następnie włączeniu adeniny do ogólnego metabolizmu rośliny.
Koniugaty:
Jak było wspomniane rybozydy i rybotydy zeatyny i izopentenyloadeniy są w pełni aktywne biologicznie.
Znane są jednak też nieaktywne pochodne - N-glukozydy (reszta glukozy podstawiona w pozycji N9 lub N7) alanylozeatyna (reszta alaniny w pozycji N9) i O-glukozyd zeatyny (reszta glukozy związana z grupą hydroksylową łańcucha izoprenowego). Ten ostatni wykazuje niewielką aktywność cytokiniy.
Do syntezy N-glukozydów wykorzystywana UDP- i UTP-glukoza. Glukozylotransferazy przeprowadzające tą reakcję są regulowane przez auksynę.
Uwaga - do tej pory nie wykryto enzymów hydrolizujących N-glukozydy, są one wyjątkowo trwałe.
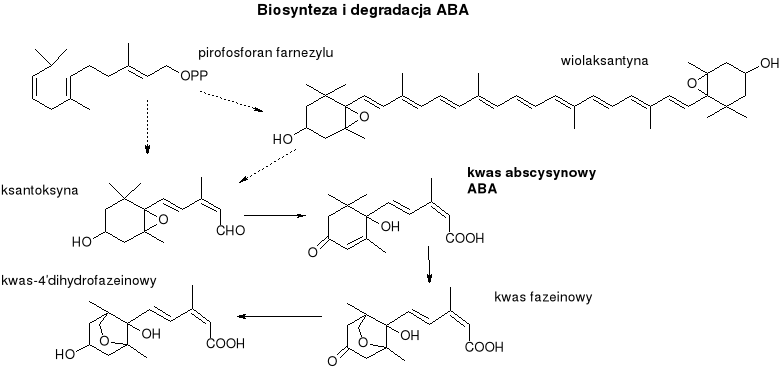
ABA
Pochodne izoprenu. Należą do seskwiterpenów.
Kiedyś nazywano go dorminą (od ang. dormancy).
Oprócz ABA pewną aktywność biologiczną wykazuje ksantoksyna, bezpośredni prekursor w biosyntezie.
Co reguluje, bądź za co odpowiada:
kiełkowanie (hamowanie)
spoczynek (wchodzenie w stan spoczynku, utrzymywanie tegoż stanu) (gdy rośliny wchodzą w stan spoczynku spada stężenie cytokinin i giberelin natomiast wzrasta stężenie ABA, gdy spoczynek ustępuje ABA spada a wzrastają cytokininy i gibereliny)
indukowanie opadania owoców i liści - stymuluje tworzenie warstwy odcinającej
regulacja odpowiedzi na stres (zwłaszcza wodny)
starzenie (przyspiesza)
indukuje zamykanie się szparek
wzrost (hamowanie)
morfogeneza
hamowanie wzrostu korzeni
dojrzewanie owoców - przyspiesza, stymuluje wytwarzanie w owocach etylenu
przyspiesza więdnięcie kwiatów
Miejsca syntezy:
dojrzałe liście i owoce
korzenie
nasiona
pąki
W komórkach zawierających plastydy.
Biosynteza:
Reszty izoprenowe dostarczane przez szlak mewalonowy. Kondensacja trzech jednostek izoprenowych - powstaje pirofosforan farnezylu, dalej cyklizacja, utlenienie, izomeryzacja do cis-ABA (forma aktywna hormonu).
Inna droga - oksydacyjna degradacja karotenoidów.
Obie drogi funkcjonują w roślinach, ale dla różnych gatunków, tkanek, warunków środowiska różny ich udział.
Degradacja:
Zaczyna się od utlenienia jednej z grup metylowych przy C6, po czym następuje natychmiastowa cyklizacja do kwasu fazeinowego., a potem do kwasu dihydrofazeinowego (oba kwasy są nieaktywne biologicznie).
Koniugaty:
Główny - ester glukozowy, ale także występuje glukozyd ABA.
Produkty degradacji ABA też mogą ulegać estryfikacji przez glukozę.
Obecność cytokinin sprzyja przechodzeniu ABA w koniugaty.
Transport przez floem (bardziej) i ksylem
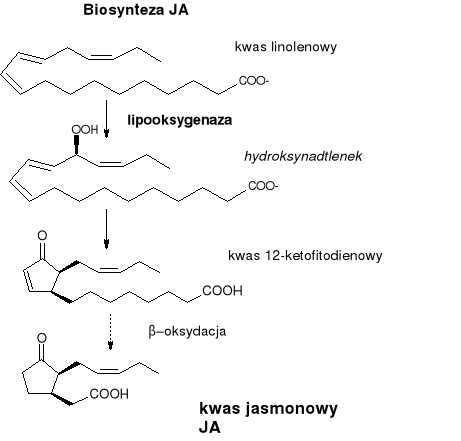
Jasmonidy
Pochodne kwasów tłuszczowych.
Główny przedstawiciel - JA, inne: ester metylowy kwasu jasmonowego, kilka streoizomerów.
Co reguluje, bądź za co odpowiada:
ochrona przed chorobami i owadami
regulacja akumulacji białek zapasowych w nasieniu
dojrzewanie owoców wzrost (reg.)
starzenie się (przyspiesza)
odpowiedź na stresy
rozwój (reg.)
fotosynteza (reg.)
niektóre przemiany metaboliczne (reg.)
Miejsca biosyntezy:
prawdopodobnie w większości żywych komórek roślinnych
Biosynteza:
Prekursor - kwas linolenowy uwalniany z lipidów roślinnych wyniku hydrolizy.
Następnie - utlenienie (enzym - lipooksygenaza), potem odwodnienie, cyklizacja i β-oksydacja.
Degradacja:
Słabo poznana. Ma charakter oksydacyjny, pierwszy etap - utlenianie grupy metylowej nienasyconego łańcucha bocznego.
W wyniku przekształceń kwasów tłuszczowych powstaje także wiele roślinnych substancji fizjologicznie czynnych, np. kwas traumatynowy (zwany hormonem przyrannym), triakontanol (składnik włosków kutykularnych, stymulator wzrostu).
Koniugaty:
Estry z cukrami - głównie glukozą i gencjobiozą, są nieaktywne. Także nieaktywne są amidy z aminokwasami (Val, Ile, Leu). Glikozylacji ulegają także produkty oksydacyjnego katabolizmu JA.
3) Uwagi pozostałe
Ostatnio często do hormonów zalicza się brasinosteroidy (BS). Około 40 związków, występują powszechnie w roślinach wyższych, aktywne w stężeniach rzędu 1 μg/dm3.
Najczęściej występujący (i najwcześniej wykryty): brasinolid.
Funkcje:
wzrost łagiewki pyłkowej
przyspieszanie wzrostu łodyg
rozwój blaszki liściowej
regulacja wzrostu i rozwoju
regulacja fotosyntezy i niektórych przemian metabolicznych
regulacje odpowiedzi na stresy i infekcje
Aktywność biologiczna brasinosteroidów zależy w znacznym stopniu od równoczesnej obecności innych hormonów: auksyn, giberelin, cytokinin.
Biosynteza BS:
Szlak kwasu mewalonowego -> dostarcza reszt izoprenowych -> kondensacje i cyklizacje -> tworzenie układu steroidowego -> prekursor - kampesterol -> reakcje utleniania - katastron - już aktywny BS -> dalsze utlenianie układu pierścieniowego i łańcucha bocznego -> prowadzi do pozostałych członków rodziny BS
Glikozylacja grup hydroksylowych łańcucha bocznego BS prowadzi do utraty aktywności biologicznej. Niewiele wiadomo o produktach degradacji tych substancji.
Hormony - regulują wzrost i rozwój, umożliwiają reakcje na bodźce.
Niektóre pasożyty wytwarzają hormony bądź ich analogi i wydzielają w stężeniach znacznie przewyższających fizjologiczne do komórek gospodarza, co prowadzi do rozregulowania metabolizmu rośliny.
Np. wspomniana wcześniej Giberella powoduje chorobę ryżu.
Bakterie z rodzaju Agrobacterium - powodują powstawanie rakowatych narośli (tumorów) na roślinach. Podczas infekcji odchodzi do przenoszenia kopii fragmentu genomu bakterii do genomu gospodarza (zostało to wykorzystane przy produkcji roślin transgenicznych, gdzie Agrobacterium jest wykorzystywane do wprowadzania transgenu), w tymże fragmencie są geny enzymów uczestniczących w syntezie auksyn (monooksydaza tryptofanu i hydrolaza indoliloacetonitrylu) i cytokinin (transferaza izopentenylowa). W zainfekowanych komórkach dochodzi do ekspresji tych enzymów, co prowadzi do nadprodukcji obu hormonów i w konsekwencji do chaotycznego wzrostu zainfekowanych tkanek.
Dla większości badanych procesów zależność między stężeniem hormonu (lub logarytmem tej wartości) a efektem jego działania ma charakter prostoliniowy, przynajmniej dla pewnego zakresu stężeń (zwykle obejmującym do 2 rzędów wielkości). Dotyczy to zarówno dawek egzogennych i stężeń endogennego hormonu.
Procesy kształtujące stężenie hormonów w komórce (WAŻNE)
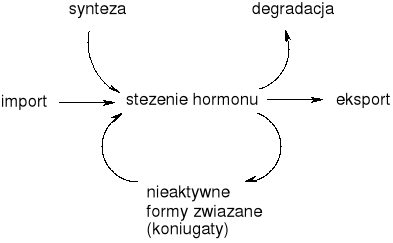
Fitohormony działają na poziomie zarówno wewnątrz, jak i międzykomórkowym. W zjawiskach morfogenezy są przekaźnikami koordynującymi procesy różnicowania, wzrostu i rozwoju. Z reguły hormony są syntetyzowane w określonych miejscach, a następnie przemieszczają się do innych tkanek i organów.
Mała specyficzność działania poszczególnych grup hormonów roślinnych. Hormony współdziałają ze sobą w skomplikowany sposób, regulując morfogenezę i rozwój roślin.
Badania: rodzaj tkanki lub organu powstającego w hodowli tkankowej zależy od równowagi stężeń fitohormonów obecnych w pożywce. Różnicowanie kalusa w korzeń albo pęd - zależy od ilościowego stosunku auksyn do cytokinin.
Kalus tytoniu.
IAA [mg/dm3] |
2 |
2 |
0,02 |
kinetyna [mg/dm3] |
0,2 |
0,02 |
1 |
organogeneza |
brak |
tworzenie korzeni |
tworzenie pędów |
Auksyny i gibereliny wpływają na podziały komórek kambium i regulują różnicowanie powstających tkanek przewodzących, przewaga auksyn - tworzenie drewna, gibereliny - tworzenie łyka. Duże stężenia IAA powodują zakładanie drewna wczesnego, zaś małe - późnego.
Egzogennie podane auksyny i gibereliny a rozwój kwiatu - auksyny - niedorozwój pręcikowia, stymulacja rozwoju słupkowia, gibereliny - stymulacja rozwoju pręcikowia. Cytokininy stymulują rozwój kielicha i słupkowia, etylen stymuluje rozwój słupkowia, ABA - hamuje rozwój pręcikowia.
W przypadku niektórych roślin traktowanie ich auksynami, bądź giberelinami powoduje powstawanie owoców partenokarpicznych (beznasiennych). We wczesnej fazie rozwoju owoców w młodych tkankach sporo cytokinin, auksyn i giberelin. Z drugiej strony nadmiar auksyny podanej egzogennie w bardzo wczesnym okresie rozwoju owoców może powodować ich przedwczesne opadanie.
Dojrzewanie owoców mięsistych - przekształcanie skrobi w cukry proste, hydroliza substancji pektynowych -> prowadzi do mięknięcia owocu, zmiany zabarwienia -> skutek rozkładu chlorofilu, wzrost syntezy karotenoidów, flawonoidów, betalain
Owoce klimakteryczne i nieklimakteryczne.
Klimakteryczne - w okresie dojrzewania ok. 5 x wzrasta intensywność wymiany gazowej, po czym następuje spadek aktywności oddechowej podczas późniejszych okresów dojrzewania i starzenia się owoców.
Nieklimakteryczne - niewielka, stała aktywność oddechowa w ciągu całego okresu dojrzewania.
Podczas zimnej stratyfikacji nasion ABA zanika w zarodkach, wzrasta natomiast ilość giberelin, cytokinin i JA. Wzrasta też etylen. Auksyny intensywnie wzrastają dopiero podczas wzrostu siewki.
Regulacja odpowiedzi na stres (więcej na stronach 658-673 Kopcewicza):
W wyniku zadziałania czynnika stresowego spada stężenie stymulatorów wzrostu (giberelin, cytokinin, czasem auksyn), natomiast zwiększa się stężenie hormonów hamujących wzrost elongacyjny, przyspieszających dojrzewanie i starzenie tkanek lub indukujących wchodzenie w stan spoczynku - ABA, JA, etylen -są one nazywane hormonami stresowymi, odpowiadają za uruchomienie mechanizmów ochronnych
Kryteria oceny, czy dany hormon uczestniczy w reakcjach prowadzących do zwiększenia odporności na stres:
działanie czynnika stresotwórczego indukuje bądź przyspiesza działanie hormonu, lub powoduje jego uwalnianie z nieaktywnej formy
zahamowanie syntezy hormonu zapobiega wystąpieniu odpowiedzi na czynnik stresowy lub ją osłabia
u mutantów charakteryzujących się niezdolnością od syntezy danego hormonu lub niewrażliwością na jego działanie nie występuje poszukiwana odpowiedź na czynnik stresowy
zwiększenie podaży hormonu powoduje zmiany metaboliczne i modyfikacje strukturalne w komórkach podobne do tych, które wywołuje czynnik stresotwórczy
reakcja na hormon pozwala osobnikowi na takie modyfikacje wzrostu lub rozwoju, które przyczyniają się od jego przetrwania
ABA - odgrywa dużą rolę w przetrwaniu okresów mrozu i suszy przez rośliny wieloletnie zapadające w stan spoczynku. Stężenie ABA wzrasta w odpowiedzi na suszę, niską temperaturę, zasolenie. Działanie ABA umożliwia dostosowanie osmotyczne komórek do niedoboru wody w środowisku.
Etylen - stymulacja syntezy w odpowiedzi na czynniki stresowe.
Wiadomo, że niektóre czynniki stresowe powodują zwiększenie zawartości endogennego JA, ale mechanizmy uczestniczenia JA w odpowiedzi na stresy są słabo poznane.
Transport hormonów
Auksyny, gibereliny, cytokininy, ABA - wykryto w soku ksylemowym i we floemie. Hormony (oraz ich koniugaty) mogą przemieszczać się w roślinie. O tym, skąd dokąd ów transport decydują oczywiście miejsce produkcji hormonu oraz położenie i właściwości komórek docelowych.
Wykazano iż estry inozytolowe IAA nagromadzone w ziarniaku zbóż przemieszczają się akropetalnie (w kierunku wierzchołka) w rosnącym pędzie, co prowadzi do uwolnienia auksyny w wierzchołku wzrostu.
Koniugaty z cukrami są lepiej rozpuszczalne niż wolne hormony ze względu na zwiększoną polarność cząsteczki.
Glikozylowe koniugaty giberelin wykryto w soku płaczu drzew.
Etylen z kolei jako gaz najczęściej po prostu dyfunduje przez symplast i apoplast. Prekursor syntezy etylenu - ACC przemieszcza się we floemie. Podobne własności (lotność i niepolarność) wykazuje ester metylowy JA, stąd postuluje się podobny sposób transportu.
Bazypetalny (skierowany ku podstawie) polarny transport auksyn w tkankach pędu. Wbrew gradientowi stężenia, szybkość 10-15 mm/h. Jest niezależny od przemieszczania się auksyny w naczyniach (które może zachodzić z większą szybkością). Bazypetalny transport auksyn jest hamowany przez inhibitory oddechowe (znane są również specyficzne inhibitory polarnego transportu IAA - TIBA - kwas trijodobenzoesowy i NPA - kwas naftyloftalamowy). Tylko w przypadku auksyn polarny transport został w pełni udokumentowany, badania nad innymi hormonami nie dały jednoznacznych wyników.
Dzięki polarnemu transportowi - przepływ auksyny z wierzchołka pędu do korzenia - jest to ważny mechanizm determinujący polarność organizmu i jego skoordynowany rozwój.
Jaka jest hipoteza: że niezdysocjowana IAA (apoplast jest kwaśny i IAA jako słaby kwas występuje w nim w postaci niezdysocjowanej) może przenikać przez błony komórkowe do wnętrza komórek. W panującym wewnątrz komórek zasadowym pH IAA dysocjuje - powstaje anion IAA-, który nie może samorzutnie przenikać przez błony. Konieczna jest zatem obecność wyspecjalizowanego transportera, który aktywnie „wyrzuca” IAA- z komórek. Transportery IAA- w tkankach zdolnych do polarnego transportu auksyn znajdują się na dolnej stronie komórek . Są też inne hipotezy - że IAA- jest aktywnie pakowany do pęcherzyków, a potem te pęcherzyki są uwalniane po konkretnej stronie komórki.
Wykazano też obecność przenośników GA w błonie komórkowej komórek aleuronowych ziarniaków zbóż i przenośnika ABA - w błonie komórek szparkowych.
Kompetencja komórek
Gromadzenie się hormonu w komórce - nazywamy jego sekwestracją.
Komórki docelowe odznaczają się dużą wrażliwością na hormon.
Mówi się o kompetencji komórek lub tkanek względem danego hormonu i danej odpowiedzi fizjologicznej (jako, że hormony roślinne działają plejotropowo).
Wysoka kompetencja komórki wobec określonego hormonu jest warunkiem pełnienia przez nią roli komórki docelowej.
Przykład takich komórek: komórki aleuronowe ziarniaków zbóż reagujące na giberelinę indukcją syntezy wielu enzymów, komórki strefy odcinającej w ogonkach liści, odpowiadające na etylen syntezą i wydzielaniem celulazy i innych enzymów hydrolitycznych.
Wrażliwość na dany hormon jest różna w różnych tkankach i organach, zmienia się też wraz z wiekiem rośliny. Reakcja na dany hormon może też być różna w zależności od tkanki docelowej.
Receptory hormonów (podrozdział streszczony bardzo pobieżnie).
Postuluje się obecność białkowych receptorów hormonów.
Wiązanie hormonu przez receptor musi być
odwracalne
o dużym powinowactwie do hormonu występującego w bardzo małym stężeniu
specyficzne w stosunku do hormonów tej samej grupy i ich aktywnych analogów
ograniczone do tkanek kompetentnych w stosunku do danego hormonu
związane ze specyficzną odpowiedzią fizjologiczną rośliny na hormon
podobne stężenie hormonu powinno wysycać receptor w tkance i powodować jej maksymalną odpowiedź fizjologiczną
Odkryte przykłady:
białka wiążące auksynę (ABP - auxin binding proteins)
Szereg doświadczeń z wykorzystaniem koleoptyli zbóż i mutantów (np. tytoniu) - ich opis, omówienie wyników i dokładniejsze informacje o ABP znajdują się w podręczniku na stronie 158.
wykazano obecność receptorów giberelin, ale dokładniej poznano tylko receptory z warstwy aleuronowej nasion owsa.
etylen - odkryto i przebadano (na Arabidopsis) białko ETR1 (aktywność kinazy histydynowej; kinazy histydynowe występują u roślin, gdzie odgrywają ważną rolę w transdukcji sygnałów), mechanizm działania etylenu należy do najlepiej poznanych. Są też znane inne receptory etylenu (ERS u Arabidopsis, TXTR-14 u pomidora).
Cytokininy - szerokie badania, CK1 - posiada domeny kinazy histydynowej. Badania na Arabidopsis.
Brasinosteroidy, też doświadczenia na Arabidopsis, białko BRI1 - transbłonowa kinaza serynowo-treoninowa.
ABA - prowadzi się badania, ale nie wyodrębniono (do czasu wydania podręcznika) odpowiedzialnego za wiązanie tego hormonu białka
Mechanizmy działania hormonów (podrozdział streszczony bardzo pobieżnie).
Kinetyka reakcji rośliny na hormon pozwala wyróżnić:
odpowiedzi szybkie
odp. powolne
Doświadczenie ze wzrostem elongacyjnym hipokotylu soi po dodaniu auksyny. W kilka minut po dodaniu hormonu rozpoczyna się pierwsza faza wzrostu, która słabnie po około 3 h, za to mniej więcej wtedy rozpoczyna się druga faza wzrostu o kinetyce odmiennej od poprzedniej. Wzrost tkanki w drugiej fazie hamowany przez inhibitory syntezy białka (np. aktynomycyna D), które nie hamują wzrostu w fazie pierwszej.
Wskazywałoby to na fakt, iż możliwa jest sytuacja, w której hormon oddziałuje na proces na dwa sposoby, różniące się szybkością i mechanizmem.
Do typowych szybkich odpowiedzi należą : polaryzacja błon (po zadziałaniu IAA lub ABA), aktywacja przez hormony niektórych enzymów, regulacja transportu jonów.
Do najlepiej poznanych szybkich odpowiedzi ma hormony należy regulacja transportu jonów przez błony.
W wielu tkankach (koleoptyl, łodyga) auksyna powoduje wzrost komórek przez osłabienie wiązań pomiędzy składnikami ściany (mikrofibryle celulozowe, pektyny, hemicelulozy). Równocześnie obserwuje się zmniejszenie pH apoplastu. Koncepcja „wzrostu kwasowego”:
Związanie auksyny przez receptor wywołuje odpowiedź polegającą na wydzielaniu H+ do apoplastu. Obniżenie pH ściany komórkowej powoduje osłabienie niekowalencyjnych wiązań pomiędzy niektórymi składnikami ściany oraz aktywację niektórych enzymów hydrolitycznych (hemicelulaz), co prowadzi do rozluźnienia struktury ściany, zmniejszenia ciśnienia turgorowego i do wzrostu komórki.
Argumenty za:
korelacja wzrostu komórki indukowanego przez IAA z kwasowością apoplastu
zakwaszenie apoplastu zwiększa szybkość wzrostu pod nieobecność IAA
wprowadzenie obojętnego buforu uniemożliwia indukcję wzrostu przez IAA
pozbawienie środowiska soli potasowych hamuje wzrost indukowany przez IAA (eksportowi H+ z komórek musi towarzyszyć import innego kationu i zazwyczaj jest to K+).
Argumenty przeciw:
Ale nie zawsze występują ilościowe zależności pomiędzy stężeniem hormonu, spadkiem pH a zwiększeniem szybkości wzrostu.
Wzrost na grubość indukowany przez etylen można zahamować dodając do środowiska obojętny bufor.
Regulowany przez ABA wypływ kationów potasowych z komórek szparkowych w warunkach suszy - jeden z głównych mechanizmów powodujących zamykanie się szparek.
Często obserwowaną odpowiedzią na hormon jest otwarcie kanałów wapniowych w błonie komórkowej, co prowadzi do zwiększenia wewnątrzkomórkowego stężenia jonów wapnia. Efekt ten obserwowano w różnych systemach w odpowiedzi na podanie auksyn, giberelin, cytokinin, ABA czy JA. Najczęściej wapń jest tu wtórnym przekaźnikiem.
Regulacja selektywnej ekspresji genów przez hormony.
Mechanizm długofalowych odpowiedzi roślin na hormony wiąże się ze zmianami ekspresji i degradacji konkretnych białek.
Element warunkujący odpowiedź na ABA - ABRE - minimum 11 nt, towarzyszy promotorom kilku poznanych genów kodujących białka indukowane przez ten hormon.
ERE - sekwencja związana z odpowiedzią na etylen - w promotorach β-1,3-glukanazy i chitynazy tytoniu.
GARE - element warunkujący odpowiedź na giberelinę - w promotorze jednego z genów α-amylazy, tylko 6 nt,
AuxRE - element warunkujący odpowiedź na auksynę, też 6 nt.
Inne
Kwas salicylowy - funkcje:
- ochrona przed pasożytami
- akumulacja białek PR (pathogen related)
Fitohormony oddziałują ze specyficznymi receptorami. W różnych miejscach różne receptory.
Aktywne w bardzo niskich stężeniach. Związki niskocząsteczkowe.
Auksyny warunkują wszelkie tropizmy.
Oświetlona część łodygi wzrasta wolniej.
Test wygięciowy - wczesny test na obecność auksyn, reakcja bardzo czuła - nawet na niewielkie stężenia.
Ucinamy koleoptyl i na jego wierzchołku kładziemy asymetrycznie bloczek agarowy. Kąt wygięcia proporcjonalny do stężenia IAA w bloczku.
Światło rozkłada IAA - po stronie oświetlonej jest go mniej.
Auksyny - reakcje kondensacji z:
- makrocząsteczkami np. białkami
- inozytolem
- kwasem asparaginowym
- amoniakiem - powstaje acetamid
- glukozą - powstaje glukozyd
Auksyny syntetyczne - przydatne w hodowlach in vitro. Część syntetycznych analogów auksyn to herbicydy
Różne gatunki roślin różnią się wrażliwością na auksyny. Stężenie dla jednych stymulujące, dla innych może już hamować wzrost elongacyjny.
Wrażliwości organów na auksyny (generalnie)
- najbardziej wrażliwe są korzenie - zakresy stężeń 10-11-10-7 mola/dm3
- potem - pąki 10-7-10-5
- najmniej wrażliwe są łodygi 10-5-10-3
Tuż po przekroczeniu maksimum czułości zaczyna się gwałtowne hamowanie wzrostu przez auksyny.
Pierwsze białka transportujące auksyny - AUX, geny tych białek podobne do genów transportujących aa, 10 helis transbłonowych.
Geotropizm korzenia. W poziomo położonym korzeniu ruch jonów wapniowych w dół. Za wapniem w jakiś sposób auksyna.
Zakwaszenie ściany komórkowej powoduje aktywację obecnych tam enzymów.
Jeśli potraktujemy roślinę egzogenną auksyną w dowolnych miejscach zaczną jej wyrastać korzenie przybyszowe.
Etylen
Przykłady owoców klimakterycznych: awokado, banan, gruszka
Egzogenne podanie etylenu powoduje anormalny przyrost na grubość.
Etiolowana siewka - występuje tzw. haczyk apikalny - czubek siewki wygięty jest w dół. Jak zadziałamy światłem haczyk się wyprostowuje, wytworzenie haczyka możemy odwrócić działając auksyną i etylenem.
Dojrzewanie owoców - czasem wcale niepotrzebne cytokininy, za to auksyny i gibereliny.
I faza - proliferacja komórek, owoc mały, twardy, gorzko-kwaśny. Faza podziałów kończy się, gdy owoc jeszcze malutki.
Potem - wzrasta objętość komórek, rusza synteza barwników i cukrów, osłabienie ścian komórkowych.
Etylen syntetyzowany także w odpowiedzi na stres mechaniczny, czy infekcję patogenu.
Rośliny wieloletnie klimatu umiarkowanego - gdy okres ciemności przekracza pewien okres krytyczny - synteza inhibitorów wzrostu.
ABA - wyłącza procesy wegetacyjne i przygotowuje do spoczynku.
Na wiosnę zaś:
- rozkład inhibitora
- gwałtowna produkcja stymulatorów.
Hamowanie wzrostu siewki jako test na stężenie.
Działaniem ABA można wywołać efekt krótkich dni.
Nawet drobne zmiany struktury cząsteczki wywołują utratę aktywności.
Utlenianie ksantoksyny powstających w wyniku rozszczepienia neoksantyny powoduje powstawanie ABA.
ABA może łatwo wchodzić w koniugację z monosacharydami np. ester glukozowy może powstawać.
Mutanty bez ABA - przedwczesne kiełkowanie nasion, podatność na więdnięcie.
Stres wodny - stężenie ABA w liściach wzrasta 50x
Znaczny wzrost stężenia ABA towarzyszy akumulacji materiałów zapasowych w nasieniu a potem dehydratacji.
W warunkach stresu wodnego ABA stymuluje syntezę specjalnych białek bogatych w Gly i Lys, hydrofilowe, zatrzymują wodę w postaci związanej na powierzchni.
2 typy uśpienia nasion:
- przez okrywę nasienną i inne tkanki okrywające zarodek - taka okrywa ogranicza wymianę gazową, uniemożliwia pobieranie wody i zatrzymuje w nasieniu inhibitory.
- uśpienie zarodka wywołane przez odpowiedni stosunek ABA/GA
Regulacja kwitnienia - egzogenne gibereliny mogą wywoływać efekt długiego dnia a ABA - krótkiego.
21
Wyszukiwarka
Podobne podstrony:
Fizjologia roślin fitohormony
fitohormony2, Materiały =), Fizjologia roślin
Fizjologia roślin wykłady, Biologia, fizjologia roślin
fizjo - wyk+éady, Leśnictwo UP POZNAŃ 2013, Fizjologia roślin drzewiastych
Zes. 7, AR Poznań - Leśnictwo, gleboznawstwo LP, Fizjologia roślin, tylko ściągi
instr X 4, Fizjologia Roślin, Biologia podstawowa
wykład 6 fizj roślin, biologia, fizjologia roślin
wyklad 4 fizj roślin, biologia, fizjologia roślin
Pytania z Fizjologii roślin z poniedziałku 3 września
Spr gosp kom, Biologia UWr, II rok, Fizjologia Roślin
fizjologia roslin - testy1+2, UG, FIZJO ROŚLIN, FIZJO, fizjo
instr V 1, Fizjologia Roślin, Biologia podstawowa
fizjo-kiełkowanie-folie111111, Leśnictwo UP POZNAŃ 2013, Fizjologia roślin drzewiastych
zadania fizjologia roślin
Fizjologia roślin
Fizjologia roślin
fizjologia roslin, Szkoła Rolnictwo studia, Szkoła, Materiały studia, fizjo roslin
więcej podobnych podstron