Mięsień poprzecznie prążkowany na obu końcach jest przyczepiony do ściegien. Zbiór komórek tworzy pęczki.
- Komórka mięśniowa poprzecznie prążkowana:
* wielojądrzasta
* cylindryczna
*otoczone sarkolemmą (pobudliwą błoną komórkową)
* wnętrze komórki wypełnia sarkoplazma (cytoplazma) i pęczki włókienek mięśniowych
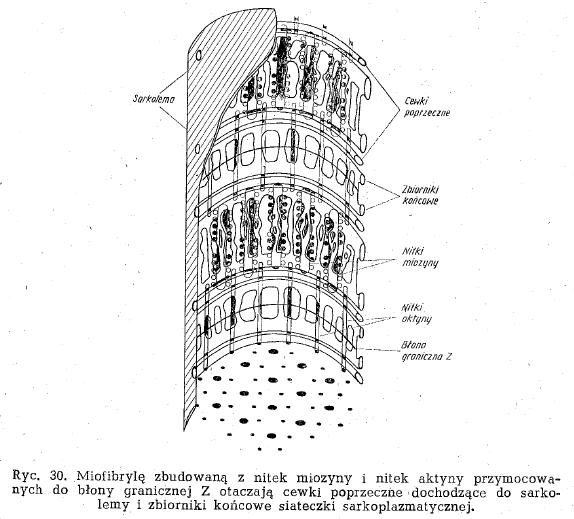
- włókienko mięśniowe (miofibryla):
* mają ciemne prążki anizotropowe (silniej załamujące światło)
* i jasne prążki izotropowe (słabiej załamujące światło)
* naprzemiennie ułożone jasne i ciemne prążki dają poprzeczne prążkowanie
* składa się z cienkich (aktynowych i tropomiozynowych) i grubych (miozynowych) nitek białek kurczliwych
* filamenty grube zbudowane są z dwóch ciężkich (splecionych ślimakowato z jednego końca tworzą ogon a z drugiego dwie głowy) i czterech lekkich łańcuchów polipeptydowych
* filamenty cienkie tworzą cztery sznury skręconych ślimakowato paciorków
* na tropomiozynie jest troponina z podjednostkami: T, I, C
* podjednostka T - łączenie troponiny z tropomiozyną
* podjednostka I - duże powinowactwo do aktyny
* a podjednostka C - do jonów wapnia
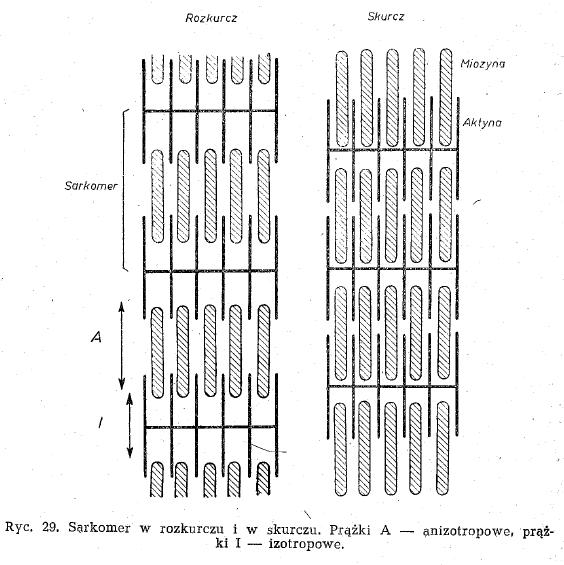
- Sarkomer - cały prążek anizotropowy (grube miozynowe) i dwie połówki izotropowego (cienkie aktynowe) doczepione do błony granicznej Z.
- Błona Z - dzieli prążki anizotropowe dwóch sąsiadujących sarkomerów. Jest do niej zaczepiona aktyna w postaci grzebieni a między nią wchodzi miozyna (otaczana 6 nitkami aktyny)
- w czasie skracania się komórki mięśniowej aktyna wsuwa się między miozynę chowając prążki aktynowe (izotropowe), a w czasie rozkurczu wysuwają się spomiędzy miozyny
- układ sarkotubularny - struktura pośrednicząca w przenoszeniu pobudzenia wewnątrz komórki mięśniowej. Składa się z:
* poprzecznych cewek, których końce dochodzą do błony komórkowej, są też między miofibrylami na granicy prążków A i B
* siateczki sarkoplazmatycznej - otacza miofibryle, prostopadle do cewek poprzecznych. W miejscu styku siateczki z cewką tworzy zbiornik końcowy z jonami wapnia
- Pompa wapniowa - w czasie depolaryzacji błony cewek otwierają się kanały wapniowe w zbiorniku końcowym, wapń wypływa między nitki białek kurczliwych i łączy się z podjednostką C troponiny. Podczas rozkurczu wsysa wolne jony wapniowe do zbiorników końcowych (rozsunięcie się nitek)
1. Mechanizm skurczu:
- bodziec - acetylocholina na synapsach nerwowo mięśniowych
- depolaryzacja
- aktywacja dla dokomórkowych jonów sodowych
- przesuwanie fali depolaryzacyjnej wzdłuż cewek
- uwolnienie jonów wapnia ze zbiornika końcowego, które łączą się z C Troponiną. Zmniejszenie powinowactwa z aktyną
- cząsteczki aktyny stykają się z głowami miozyny
- aktywacja enzymatyczna miozyny
- hydroliza ATP (cząsteczki aktyny stykają się z głowami miozyny)
- zmiana konformacji aktyny względem nici miozyny
- powrót do pierwotnego położenia nitki cienkie aktyny wsuwają
- zmiana konformacji aktyny względem nici miozyny się w grube miozynowe
- powrót do pierwotnego położenia (Model ślizgowy)
- sprzężenie mechaniczno-chemiczne - czerpanie energii z hydrolizy ATP dla czynności mechanicznej kom. Mięśniowej
- sprzężenie elektromechaniczne - czynność elektryczna ponadprogowa oddziałuje na czynność mechaniczną komórki, lub poszczególnych jej elementów (kurczenie komórki maksymalne)
- okres bezwzględnej wrażliwości - depolaryzacja błony mięśnia trwa 1-3ms i w tym czasie jest nie wrażliwa
Rodzaje skurczów:
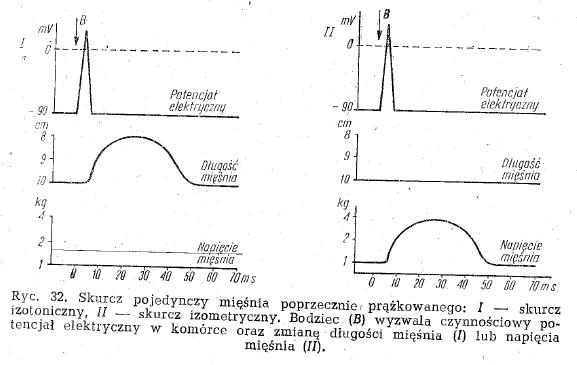
- tężcowy izotoniczny - komórki mięśniowe skracają się i cały mięsień ulega skróceniu, jego napięcie nie zmienia się. Przyczepy mięśnia przybliżają się do siebie
- tężcowy izometryczny - zwiększenie napięcia mięśnia bez zmian jego długości. Przyczepy nie zmieniają położenia.
- tężcowy auksotoniczny - skracanie się mięśnia i jego napięcie. Zbliżenie przyczepów
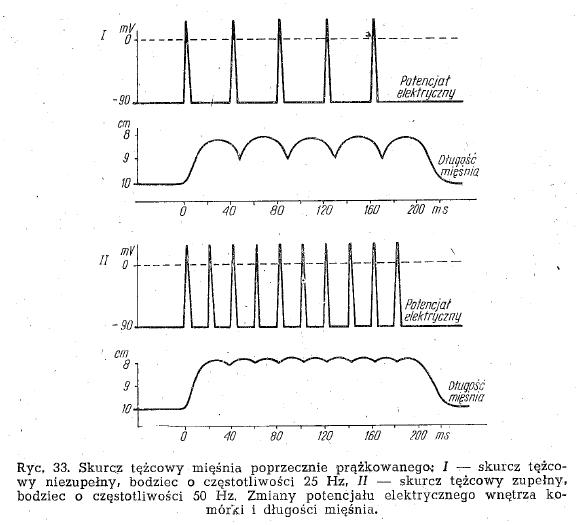
- sumowanie się skurczów pojedynczych - powtarzające się bodźce nadprogowe wywołują kolejne skurcze, które zaczynają się na siebie nakładać. Pojawia się skurcz tężcowy zupełny lub niezupełny.
- skurcz tężcowy zupełny - bodźce pobudzają mięsień w odstępach krótszych niż trwa pojedynczy skurcz
- są mięśnie szybko i wolno kurczące się (nierozciągnięte)
- nierozciągnięty mięsień słabo się kurczy, za bardzo rozciągnięty też
- skurcz maksymalny - pobudzone są wszystkie komórki mięśniowe
- energia z hydrolizy ATP w czasie skurczu
- resynteza ATP z rozpadu glukozy (glikoliza tlenowa)
- dysocjacja mioglobiny - podobna do hemoglobiny, uwalnia związany tlen kiedy zapotrzebowanie na tlen przerasta dostarczanie tlenu.
- glikoliza beztlenowa - podczas niej resyntezuje się ATP, w warunkach beztlenowych, mniej skuteczna
- gdy gromadzą się mleczany, spada pH
Powstawanie energii cieplnej:
- w procesie spoczynkowego metabolizowania wewnątrzkomórkowego
- w czasie reakcji chemicznych, podczas których aktywuje się miozyna
- w czasie skracania się komórek mięśniowych
- w procesie rozkurczu i wydłużania się komórki mięśniowej
- w procesie odnowy związanej z resyntezą ATP i działaniem pompy sodowo potasowej
- mięśnie poprzecznie prążkowane unerwiane są przez komórki nerwowe w ruchowej części pnia mózgowia i rdzenia kręgowego.
- im bardziej wyspecjalizowany mięsień tym więcej komórek go unerwia
- jednostka motoryczna - komórka nerwowa i wszystkie komórki mięśniowe przez nią unerwiane. Działają niesynchronicznie.
Siła skurczu mięśnia zależy od:
- liczby jednostek motorycznych podczas skurczu
- częstotliwości z jaką jednostki motoryczne są pobudzane
- stopnia rozciągnięcia mięśnia przed skurczem
- napięcie mięśniowe spoczynkowe - nawet jak mięśnie nie pracują to są w skurczu tężcowym izometrycznym długo utrzymującym się.
Napięcie mięśniowe jest regulowane przez:
- nadrzędne ośrodki ruchowe w ośrodkowym układzie nerwowym
- samoregulację
- Samoregulacja - rozciągnięcie mięśnia wyzwala odruchowe napięcie (skurcz zginaczy powoduje rozciąganie mięśni antagonistycznych - prostowników i zwiększanie ich napięcia)
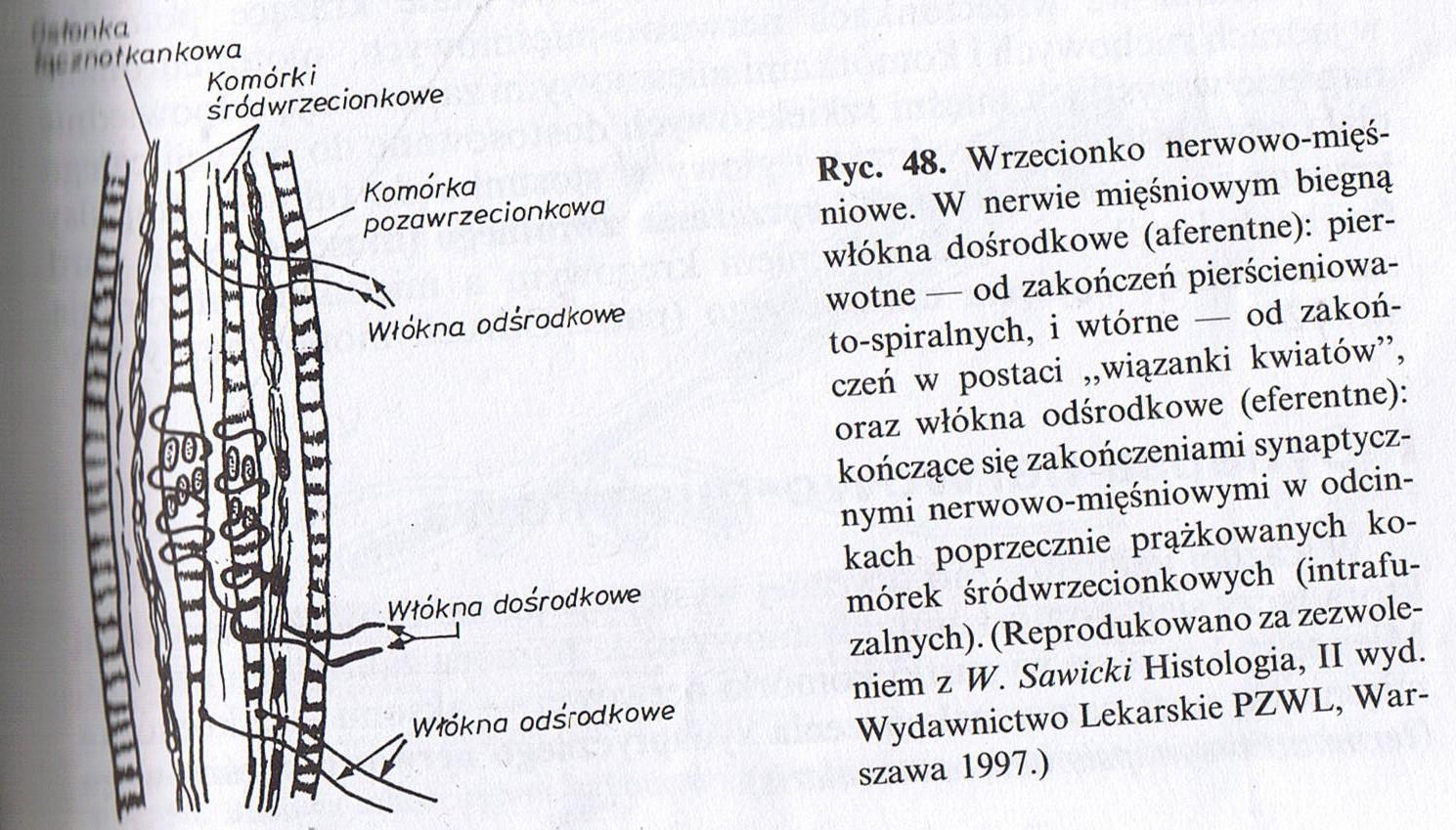
- komórki mięśniowe ekstrafuzalne (większość mięsni) - mają jednolitą budowę, są skupione w pęczki i oba ich kończe są przyczepione do ścięgien. Unerwione długimi neuronami Alpha
- komórki mięśniowe intrafuzalne - w części środkowej (niekurczącej się) nie maja poprzecznego prążkowania, są skupione w pęczki (wrzeciona nerwowo mięśniowe) otoczone łącznotkankową torebką. Unerwiane przez małe neurony gamma.
- wrzeciona nerwowo mięśniowe - przyczepiają się końcami do komórek ekstrafuzalnych, znajdują się w nich receptory wrażliwe na rozciąganie, regulowane przez neurony gamma
Mechanizm samoregulacji:
- pobudzenie receptorów we wrzecionach nerwowo-mięśniowych rozciągnięciem mięśnia
- wysyłanie impulsów do neuronu alpha w jądrach ruchowych pnia mózgowia/rdzenia kręgowego
- neurony alpha wysyłają impulsy do komórek ekstrafuzalnych i kurczą je
a) skurcz izotoniczny komórek ekstrafuzalnych zmniejsza pobudliwość receptorów we wrzecionach
- zmniejszenie napięcia mięśnia
b) skurcz izometryczny nie zmniejsza pobudliwości receptorów i napięcie mięśniowe, utrzymuje się długo
- sprzężenie zwrotne - impulsy krążą po zamkniętej pętli między receptorami wrzeciona nerwowo mięśniowego, motoneuronami w jądrach ruchowych i komórkami mięśniowymi. Zapewnia optymalne napięcie mięśni, niezbędne to utrzymania postawy. A krążąc pomiędzy rdzeniem kręgowym a mięśniami utrzymuje samoregulację napięcia mięśniowego.
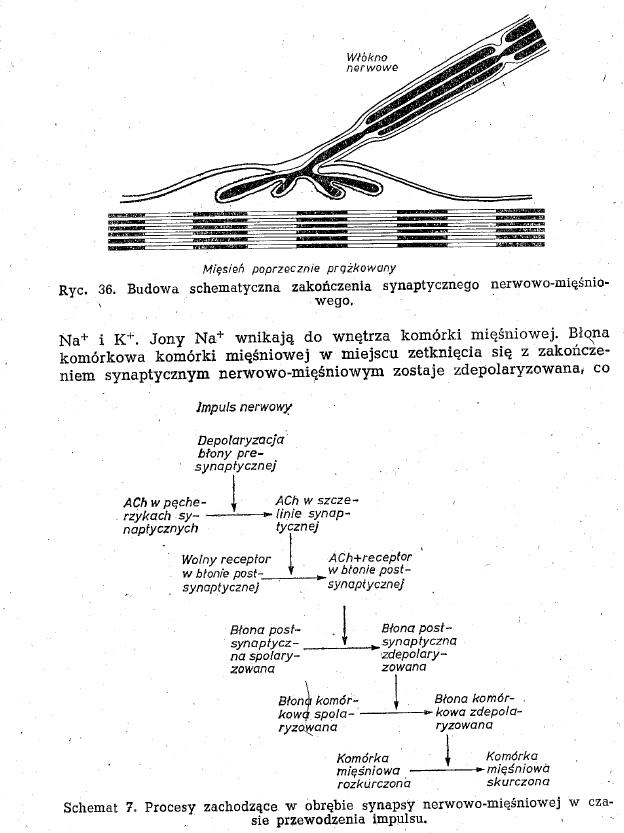
- zakończenie synaptyne nerwowo mięśniowe - miejsce stuki aksonu z komórką mięśniową. Tworzą je włókna nerwowe A przy unerwianej komórce tracą osłonkę i rozdzielają się na wiele stopek końcowych.
- błona komórkowa jest w miejscu styku z neuronem pofałdowana, zagłębiona i tworzy rynienki otaczające każdą stopkę końcową
- szczelina synaptyczna - jest pomiędzy błoną neuronu postsynaptyczną a pofałdowaną błoną komórki mięśniowej
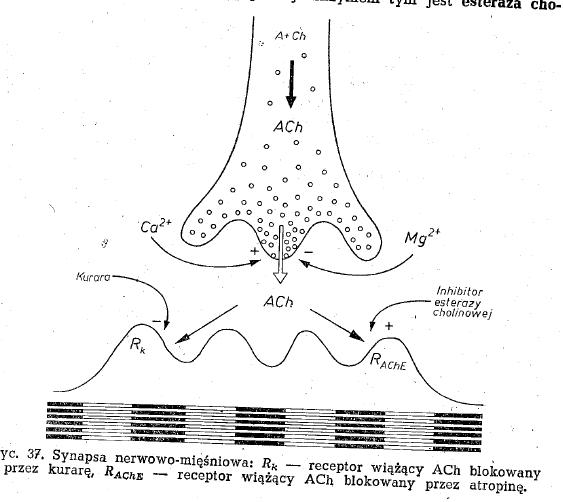
-sprzężenie elektrowydzielnicze - proces polegający na zainicjowaniu przez falę depolaryzacji uwalniania mediatora synaptycznego do szczeliny synaptycznej(stopki końcowe mają pęcherzyki synaptyczne z acetylocholiną).
- acetylocholina - wpływa na błonę postsynaptyczną tak, że otwiera kanały dokomórkowego prądu jonów sodowych
- potencjał zakończenia synaptycznego nerwowo-mięśniowego - wahanie potencjału elektrycznego w okolicy zetknięcia błony mięśnia z zakończeniem synaptycznym nerwowo-mięśniowym, spowodowane depolaryzacją.
- kurara - blokuje receptory acetylocholinowe, a konkretniej łączenie się Ach z receptorem, co hamuje depolaryzację
- esteraza cholinowa - jest w błonie postsynaptycznej rozkłada Ach i kwas octowy
- inhibitory esterazy cholinowej - hamują działanie esterazy i zwiększają wrażliwość błony na działanie acetylocholiny (mniej jej trzeba żeby zdepolaryzować błonę.
- Jony wapniowe - warunkuje uwalnianie Ach z zakończenia synaptycznego nerwowo-mięśniowego
- jony magnezowe - hamuje uwalnianie Ach
- komórka mięśniowa gładka - nie ma jednostek kurczliwych (sarkomerów). Ma nitki kurczliwe ułożone wzdłuż komórki. W czasie skurczu nitki te skracają się (aktyna nachodzi na miozynę)
- kalmodulina - jest w cytoplazmie, łączy się z jonami wapniowymi i aktywuje jeden z łańcuchów lekkich miozyny. Następuje hydroliza ATP i zmiana konformacji miozyny w stosunku do nitki grubej miozyny, co powoduje przesuwanie się nitek cienkich aktyny wzdłuż miozynowych
Komórki mięśniowe gładkie:
- wielojednostkowe mięśnie gładkie - poszczególne komórki kurczą się niezależnie od siebie , pobudzenie nie przenosi się z jednej komórki na drugą. Są w ścianach naczyń krwionośnych i tęczówce
- trzewne mięśnie gładkie - tworzą warstwy lub pierścienie równolegle ułożonych komórek, w których pobudzenie przenosi się z jednej komórki na drugą, dzięki połączeniom szczelinowym. Tworzą czynnościowe syncytia. Są w ścianach przewodu pokarmowego, moczowodzie, pęcherzu moczowym, macicy. Spełniają dwa typy czynności mechanicznych - skurcze i zmiany napięcia.
- wnętrze komórek mięśniowych gładkich ma potencjał spoczynkowy -50mV
- zmniejszenie potencjału następuje, jak otwierają się kanały dla prądu jonów Ca2+ i następuje ich napływ do komórki.
- hyperpolaryzacja - komórka ma zmniejszoną pobudliwość i wnętrze ujemne (-65mV). Pozostaje w rozkurczu. Spowodowana jest ucieczką jonów K+ z komórki lub utrudnionym wchodzeniem Ca2+ do komórki.
Skurcz komórek mięśniowych gładkich poprzedza:
- potencjał czynnościowy iglicowy (50ms)
- potencjał iglicowy przechodzący plateau depolaryzacji (100ms-1s)
- w komórkach częściowo depolaryzowanych skurcz nie jest poprzedzany potencjałem czynnościowym
- skurcz poprzedzany potencjałem czynnościowym zaczyna się po 200ms od początku depolaryzacji
Czynniki kurczliwości mięśni gładkich:
- samoistne pobudzenie występujące rytmicznie w kom. Trzewnych. Fala się przesuwa po całym mięśniu. Rozrusznik pozostałych komórek. Wyzwala 1 skurcz na 5s.
- czynnika miejscowego mechanicznego lub chemicznego działającego na komórki (rozciąganie, zmiany pH, zwiększanie prężności CO2)
- przekaźników chemicznych przenoszonych drogą humoralną (hormony rdzenia nadnerczy)
- przekaźników chemicznych wydzielonych z aksonów neuronów należących do układu autonomicznego (działają jednocześnie na kilka komórek)
- noradrenalina - wydzielana na zakończeniach neuronów zazwojowych układu współczulnego, rozkurcza trzewne mięśnie przewodu pokarmowego. działa na receptory adenergiczne alpha (jony Ca2+ wychodzą z komórki, następuje rozkurcz) i beta (aktywacja cyklazy adenylanowej, jony Ca2+ są wiązane w komórce, spada ich stężenie w komórce. Rozkurcz komórek)
- acetylocholina - wydzielana na zakończeniach neuronów układu przywspółczulnego, kurczy trzewne mięśnie przewodu pokarmowego. Łącząc się z receptorami muskarynowymi m3 i m4 aktywuje fosfolipazę C i uwalnianie inozytolofosforanu (IP3), co wywołuje wzrost stężenia Ca2+ w komórkach i ich skurcz
- stan mięśni jest wypadkową działania antagonistycznych noradrenaliny i acetylocholiny
- na mięśnie wielojednostkowe gładkie w tętnicach acetylocholina działa rozkurczająco a noradrenalina kurcząco
Mięsień sercowy składa się z:
- układu przewodzącego - brak prążkowania, mają dużo sarkoplazmy i glikogenu, leżą pod wsierdziem
- mięśnia przedsionków
- mięśnia komór
- odgrodzone pierścieniami włóknistymi
- jest to mięsień z włókienkami mięśniowymi i elementami kurczliwymi tworzącymi sarkom er
- wstawka - błony komórkowe ściśle do siebie przylegają w miejscu prążków Z tworząc pozazębianą błonę. Pobudzenie przenosi się z jednej komórki na drugą dzięki niej i połączeniom komunikującym, odpowiada maksymalnie i jest syncytium fizjologicznym
- potencjał spoczynkowy wynosi -90mV
- dokomórkowy wolny prąd jonów potasowych i sodowych - potas i sód ciągle wychodzą na zewnątrz (K+) i do wewnątrz (Na+)
- pompa sodowo potasowa utrzymuje odpowiednie stężenie jonów w komórce i na zewnątrz jej, dzięki czemu utrzymany jest potencjał spoczynkowy
Czynność mięśnia sercowego:
- pobudzenie komór
- faza 0 - bardzo szybka depolaryzacja (spowodowana napływem jonów sodowych i wapniowych)
- faza 1 - nieznaczna depolaryzacja (w przedsionkach, odpływ jonów K, a w komorach napływem Cl-)
- faza 2 - utrzymanie stałej depolaryzacji przez 300ms (równowaga między stężeniem K+ i Ca2+)
- faza 3 - powrót do potencjału spoczynkowego (uciekanie K+ i powrót elektroujemności)
* w fazie 0, 1, 2 jony wapniowe napływają do sarkoplazma i zbiorników sarkoplazmatycznych
- glikokaliks - tu magazynowane są jony wapniowe
- okres bezwzględnej refrakcji (niewrażliwości) - obejmuje depolaryzację i fazę 1 i 2 repolaryzacji
- okres względnej refrakcji (niewrażliwości) - obejmuje fazę 3 repolaryzacji
- oba te okresy są dłuższe niż trwa skurcz, więc w mięśniu sercowym nie występuje skurcz tężcowy
- prawo Starlinga - większa ilość krwi wpływającej do serca w trakcie rozkurczu (objętość końcoworozkurczowa) powoduje wypłynięcie większej ilości krwi w trakcie skurczu (objętość wyrzutowa). Innymi słowy, siła skurczu mięśnia jest wprost proporcjonalna do długości początkowej jego włókien. Długość włókien mięśniowych zależy od stopnia wypełnienia komór serca krwią, a ta z kolei od dopływu krwi do serca.
- energia do skurczu czerpana jest z kwasów tłuszczowych, węglowodanów, aminokwasów, cał ketonowych, kwasu mlekowego i kwasu pirogronowego
układ przewodzący mięśnia sercowego tworzy:
- węzeł zatokowo-przedsionkowy (Keitha- Flacka) - 72 razy na minutę się pobudza, narzucają rytm reszcie
- węzeł przedsionkowo-komorowy (Aschoffa-Tawary)
- pęczek przedsionkowo-komorowy (Paladino-Hisa) - ma dwie odnogi kończące się komórkami mięśniowymi przewodzącymi (Purkinjego)
- rozrusznik - komórki same pobudzają się
- prepotencja rozrusznika - samoistne pobudzenie komórek powodowane jest powolną depolaryzacją błony między potencjałami czynnościowymi. Występuje na skutek napływu jonów wapniowych aż do osiągnięcia potencjału progowego wywołując potencjał czynnościowy (depolaryzację)
Depolaryzacja -> węzeł zatokowo przedsionkowy -> mięsień przedsionków -> węzeł i pęczek przedsionkowo-komorowy -> komórki mięśniowe przewodzące -> mięsień komór
- komórki unerwiające serce nie wywołują w nim skurczów, tylko wyzwalają transmiter przyspieszający albo zwalniający akcję serca.
- acetylocholina działająca na receptor m2, zwiększa wychodzenie jonów K+ i wydłuża czas trwania prepotencjału. Serce bije wolno
- noradrenalina działająca z receptorami beta-adenergicznymi, zwiększa napływ jonów Ca2+ do rozrusznika, czas prepotencjału zmniejsza się, serce bije szybciej.
- układ współczulny przeważa nad układem przywspółczulnym, dlatego że po zablokowaniu obu receptorów dla noradrenaliny i acetylocholiny, częstość skurczów przyspiesza w porównaniu do częstości występującej w spoczynku
Wyszukiwarka
Podobne podstrony:
5322
5322
5322
5322
5322
04025 Anleitung Asientascheid 5322
5322
więcej podobnych podstron