skanuj0005
Prometafaza
Na początku prometafazy otoczka jądrowa ulega fragmentacji. Zapoczątkowuje ją akty wacja białkowej kinazy fazy M, która fosforyluje i zmienia konformację białek blaszki gęstej przy wewnętrznej błonie otoczki jądrowej. W tym czasie zanika również jąderko(a) przy udziale fosforylacji białka — nukleoliny. W dzielącej się komórce widoczne są dwie tzw. gwiazdy (czyli pary centriol, od których odchodzą promieniście mikrotubule). Kondensujące chromosomy są łączone z elementami wrzeciona podziałowego. Łączenie to odbywa się w sposób zapewniający przejście każdej siostrzanej chromatydy do jednego bieguna komórki, tj. do jednej potomnej komórki po dokonaniu się mitozy. Odbywa się to przez związanie mikrotubul kinetochorowych z kinetochorem każdej siostrzanej chromatydy, a następnie oddziaływanie tych mikrotubul z innymi elementami wrzeciona. W wyniku takiego oddziaływania każda siostrzana chromatyda jest orientowana prostopadłe w stosunku do długiej osi wrzeciona podziałowego, a jej kinetochor kieruje się ku jednemu z dwóch biegunów komórki. Oddziaływanie takie prowadzi do przesuwania chromosomów ku płaszczyźnie centralnej (równikowej) wrzeciona. Procesy te trwają 10-20 min i polegają na oscylacyjnych ruchach chromosomów, które wynikają z przypadkowych oddziaływań mikrotubul kinetochorowych z mikrotubulami biegunowymi.
Metafaza
W czasie metafazy chromosomy ulegają dalszej kondensacji, która kończy się wtedy, gdy chromosomy ustawiają się w płaszczyźnie równikowej komórki. Jest to typowa konfiguracja chromosomów metafazy: chromosomy oglądane z góry od strony bieguna tworzą figurę podobną do gwiazdy (gwiazda macierzysta), natomiast oglądane z boku tworzą figurę przypominającą płytkę (płytka równikowa) (rys. 20.3). W metafazie chromosomy pozostają w stałych pozycjach w stosunku do wrzeciona podziałowego. Jednocześnie odbywa się przemieszczanie składników cytoplazmy ku biegunom komórki. Przemieszczanie to zachodzi z szybkością ok. 1 pm na min i ma na celu przekazanie do komórek potomnych składników cytoplazmy w możliwie równych ilościach. W wyniku kondensacji każdy chromosom metafazowy składa się z 30 nm włókienek chromatyny, które tworzą struktury wyższego rzędu w postaci włókien o średnicy 200-400 nm. Te ostatnie prawdopodobnie poprzez spiralizację wytwarzają 700-800 nm chromatydy. W chromosomach metafazowych stopień upakowania nici DNA jest 1:5 000 — 1:10 000. Oznacza to, że DNA jądra komórkowego po replikacji, długości ok. 4 m skraca się do długości 200-400 pm. Ta skrócona, upakowania helisa DNA zawarta jest w komórkach somatycznych człowieka w 46 chromosomach. Pod koniec metafazy chromosomy układaj ą się na obwodzie wrzeciona podziałowego, a każdy z nich ulega częściowemu rozdzieleniu na dwie chromatydy.
Anafaza
W anafazie następuje nagłe rozdzielenie siostrzanych chromatyd w mieiscŁch_££ntromerów-Na oba kinetochory tego samego chromosomu działają takie same siły skierowane ku różnym hegunom wskutek ojidziaływanjajmifag^ kinetochorowych biegunowymi. Przerwanie ciągłości centromerów powoduje ruch siostrzanych chromatyd ku biegunom. Przypuszcza się, że jednoczesne i nagłe rozdzielenie chromosomów odbywa się poprzez syntezę DNA. Ruch chromosomów odbywa się z różną szybkością zależną od wielu czynników (rodzaj komórki, tempe-
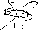
ratura, itp.) ale średnio wynosi 2,5 |lm na minutę. Na ogół ruch rozpoczyna się jednocześnie dla wszystkich chromosomów i postępuje z taką samą szybkością dla całego zestawu chromosomów. Szybkość ta jest odwrotnie proporcjonalna do odległości chromosomów od biegunów, tzn. maleje w miarę zbliżania się do biegunów.
Mechanizm ruchu chromosomów w prometafazie i metafazie oraz w anafazie nie jest w pełni wyjaśniony. Przypuszcza się, że zajmowanie pozycji w płaszczyźnie równikowej przez chromosomy jest wynikiem równowagi między siłami działającymi na siostrzane chromosomy (chromatydy połączone w centromerze) ze strony mikrotubul kinetochorowych obu biegunów. Natomiast ruch chromosomów ku biegunom komórki jest wypadkową dwóch ruchów: wydłużania się wrzeciona podziałowego (rys. 20.5), któremu towarzyszy oddalanie się od siebie centriol biegunowych oraz mchu samych chromosomów odbywającego się przez pociąganie.
Udział obu mechanizmów w oddalaniu się od siebie chromosomów siostrzanych jest różny u różnych organizmów. Pod koniec anafazy odległość między biegunami może, w komórkach niektórych gatunków, wydłużać się 15-krotnie w stosunku do odległości na początku anafazy. W komórkach ssaków oba mechanizmy odgrywają mniej więcej jednakową rolę. Wydłużanie wrzeciona podziałowego w czasie anafazy odbywa się najprawdopodobniej przy pomocy mechanizmu ślizgowego: leżące obok siebie i równolegle do siebie mikrotubule biegunowe obu biegunów przesuwają się względem siebie, w przeciwnych kierunkach (rys. 20.5). Energia do ruchu pochodzi z ATP. Wydłużaniu wrzeciona podziałowego towarzyszy wydłużanie mikrotubul.
wczesna anafaza
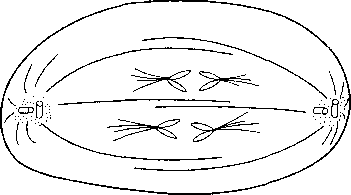
późna anafaza
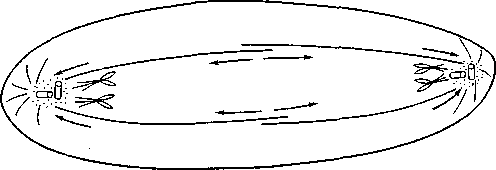
pociąganie
pchanie
pociąganie
Rys. 20.5. Schemat pokazujący popychanie (przez wydłużanie się wrzeciona podziałowego) i pociąganie (przez oddziaływanie mikrotubul kinetochorowych z mikrotubulami biegunowymi lub zbiornikami siateczki śródplazmatycznej gładkiej) chromosomów w anafazie
347
Wyszukiwarka
Podobne podstrony:
62591 skanuj0053 (35) Na początku dwudziestolecia międzywojennego teoria czasu wolnego się przedmiot
skanuj0031 6 6 9 7 8 Rys. 8.1. Budowa jądra komórkowego. 1 — otoczka jądrowa, 2 —
Prometafaza biegun wrzeciona fragmenty otoczki jądrowej aktywnie poruszające się chromosomy
Prometafaza biegun wrzeciona fragmenty otoczki jądrowej aktywnie poruszające się chromosomy
więcej podobnych podstron