608
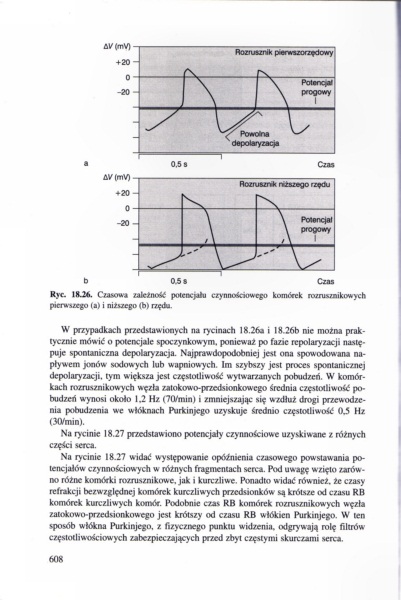
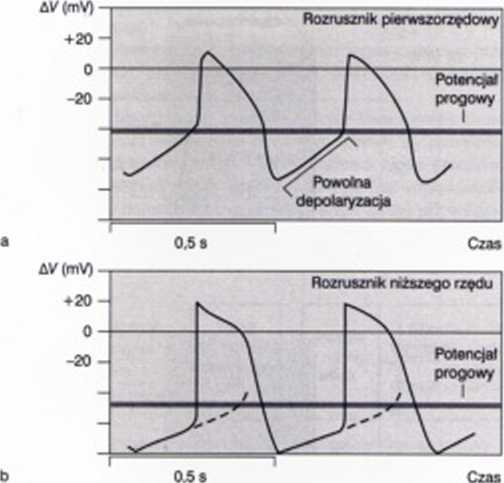
Ryc. 1RJ*. C/iwui zależność potencjału czynnościowego komórek ru/rus/nikowych pierwszego (a) i ni/wrego (b) rzędu
W przypadkach przedstawionych na rycinach 18.26* i 18.266 nic można praktycznie mówić o potencjale spoczynkowym, ponieważ po fazie repolaryzacji następuje spontaniczna depolary zacja. Najprawdopodobniej jest ona spowodowana napływem jonów sodowych lub wapniowych. Im szybszy jest proces spontanicznej depolaryzacji, tym większa jest częstotliwość wylwar/anych pobudzeń. W komórkach rozrusznikowych węzła zatokowo-przedsionkowego średnia częstotliwość pobudzeń wynosi około 1.2 Hz (70/min) i zmniejszają się w zdłuż drogi przewodzenia pobudzenia we włóknach Purkinjego uzyskuje średnio częstotliwość 0.5 Hz (30/min).
Na rycinie 18.27 przedstawiono potencjały czynnościowe uzyskiwane z różnych części serca.
Na rycinie 18.27 widać występowanie opóźnienia czasowego powstawania potencjałów czynnościowych w różnych fragmentach serca. Pod uwagę wzięto zarówno różne komórki rozrusznikowe, jak i kurczliwe. Ponadto widać również, że czasy refrakcji bezwzględnej komórek kurczliwych przedsionków są krótsze od czasu RB komórek kurczliwych komór. Podobnie czas RB komórek rozrusznikowych węzła zatokowo-przedsionkowego jest krótszy od czasu RB włókien Purkinjego. W ten sposób włókna Purkinjego. z fizycznego punktu widzenia, odgry wają rolę filtrów częstotliwościowych zabezpieczających przed zbyt częstymi skurczami serca.
608
Wyszukiwarka
Podobne podstrony:
P1010376 F Fizjologiczne podstawy elektrokardiografii Ryc. 4. Potencja) czynnościowy komórek węzła
Serce 1 Prawidłowe odpowiedzi SERCE I. Potencjał czynnościowy komórek roboczych mi
f17 Ryc.19. Wykres zależności potencjału pola elektrostatycznego w danym punkcie od jego odległości
365 (16) Częstottwoóć inxxiMCji t [Hz) Ryc. 14.18. Zależność potencjału receptorowego i częstotliwoś
Układ Krążenia0032 o Ryc. 95. Linie łączące punkty o równych potencjałach na powierzchni ciała zależ
Slajd44 (21) Depolaryzacja i Prawidłowy potencjał czynnościowy: depolaryzacja zależna od k
A) prawidłowe przewodzenie potencjał czynnościowy dalsza część tkanki przewodzącej Ryc. 5.14.
Ryc. UL27. Typami potencjały czynnościowe w różnych częściach wi. Ciągłymi kinami zaznaczono potencj
Mięśnie0023 8 A Schemat objaśniający stosunki między potencjałem czynnościowym a skurczem Ryc. 24. A
004 (53) i Mv Ryc. 191. Kształt potencjału czynnościowego komórki m. sercowego. W okresie repolaryza
004 (53) i Mv Ryc. 191. Kształt potencjału czynnościowego komórki m. sercowego. W okresie repolaryza
Ryc. 14.14. Zjawnko wnw»wj w czasie. U góry potencjał czynnościowy docierający do Mony preiytuptyczn
fizjologia12 ktrona 1 1. Powstanie potencjału czynnościowego polega na: a)
więcej podobnych podstron