bullock (9)

Fizjologia błony komórkowej i komórki
9
A

B
glukoza ^ Q l'l‘*
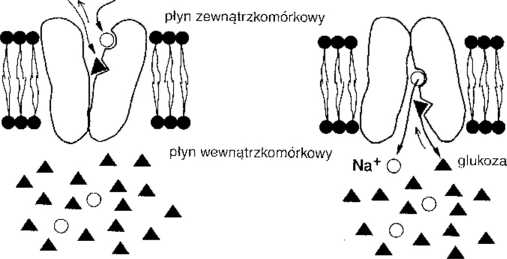
RYC. 1-5. Glukoza jest transportowana przez błonę komórkową w procesie wtórnego transportu czynnego (tzn. transportu aktywnego wykorzystującego pośrednie źródło energii). (Ą) Powinowactwo cząsteczek nośnika do glukozy wzrasta w obecności jonów Na", co oznaczono odpowiednią długością strzałek. Cząsteczka glukozy przechodzi przez błonę razem z jonem Na'. (B) Przeniesiony do wnętrza komórki NaT odłącza się od nośnika, zmniejszając jego powinowactwo do glukozy i powodując również jej odłączenie od nośnika.
(1) pompa wapniowa (Ca3+) w siateczce sarkoplazmatycznej (SR) komórek mięśniowych, warunkująca utrzymanie stężenia jonów Ca1' poniżej 0,1 jU.mol/1,
(2) pompa potasowo-wodorowa (K -H ) komórek błony śluzowej żołądka, umożliwiająca wydzielanie jonów H+ do światła żołądka podczas trawienia.
Zjawiska wtórnego transportu czynnego wykorzystują energię zgromadzoną pod postacią przezbłonowego gradientu stężenia jonów Na' do transportu substancji przeciw ich gradientowi stężeń (ryc. 1-5).
a. Działanie. Wiele jonów i substancji odżywczych transportowanych przeciw gradientowi elektrochemicznemu wykorzystuje mechanizm wtórnego transportu czynnego zależnego od jonów Na'.
(1) Glukoza i aminokwasy są reabsorbowane z kanalików bliższych nerek i wchłaniane ze światła jelit z wykorzystaniem mechanizmu transportu wtórnie aktywnego zależnego od jonów Na+.
(2) Jony wapnia są usuwane z cytoplazmy kardiomiocytów (i innych komórek mięśniowych) w procesie wtórnego transportu czynnego zależnego od jonów Na+, zwanego wymiennikiem Na+-Ca2+. Wymiennik sodowo-wapniowy zużywa energię zmagazynowaną w trzech jonach sodowych do przetransportowania jednego jonu wapnia na zewnątrz komórki. Ten mechanizm - we współdziałaniu z pompą Ca1" siateczki sarkoplazmatycznej, opisaną w rozdz. III C 1 b (1) - prowadzi do rozkurczu mięśnia.
(3) Jony H+ powstające w komórce w wyniku procesów metabolicznych są usuwane na zewnątrz w procesie wtórnego transportu aktywnego zależnego od jonów Na+. Ten mechanizm jest istotny dla utrzymania normalnego pH wnętrza komórki, a także dla reabsorpcji dwuwęglanów z kanalików bliższych nerek.
b. Gradient elektrochemiczny. Energia zmagazynowana w gradiencie elektrochemicznym jonów Na+ jest sumą potencjału błonowego EM (energii elektrycznej powodującej napływ jonów sodowych do komórki) oraz potencjału równowagi (Nern-sta), EJ0n (ekwiwalent energii gradientu stężeń w postaci energii elektrycznej).
Wyszukiwarka
Podobne podstrony:
bullock (5) Fizjologia błony komórkowej i komórki 5 RYC. 1-3. Jeśli błona półprzepuszczalną oddziela
bullock (3) Fizjologia błony komórkowej i komórki J 3 a. Białka zewnętrzne, związa
bullock (7) 7 Fizjologia błony komórkowej i komórki gdzie i oraz f odnoszą się do początkowej i końc
bullock (11) Fizjologia błony komórkowej i komórki 11 W ostatnim czasie pacjentka zaobserwowała u s
342 (19) Ryc. 1X9. Schemat modelu elektrycznego błony komórkowej. W modelu gradienty stężeń poszczeg
histologia wykład�9 zlewanie sie ffuzja) błony komórkowej owocytu i plemnika Zaraz po wniknięciu p
skanuj0101 (15) trycznej. Ultradźwięki wywierają również wpływ na potencja* tryczny błony komórkowej
Slajd12 (51) Powstanie błony komórkowej nieprzepuszczalnej dla cząsteczek być może było ogromnym
Slajd33 (30) Jest to raptowna depolaryzacja, a następnie repolaryzacja błony komórkowej Składa się z
SNC01760 Uważa się że kształt erytrocytów zależy między innymi odprewtdtowej budowy błony komórkowej
skanowanie0017 83 ^odstawowe właściyyości błony komórkowej masie cząsteczkowej około 100 kDa. Łańcuc
więcej podobnych podstron